
Using Science and Technology to Unveil The Hidden Delicacy Terfezia arenaria, a Desert Truffle




Abstract
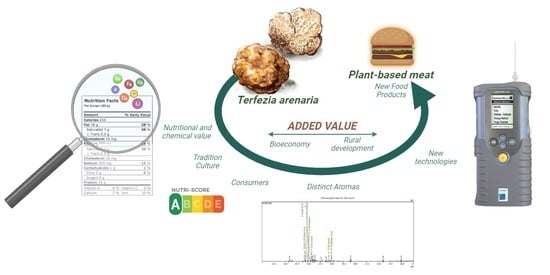
1. Introduction
2. Materials and Methods
2.1. Terfezia arenaria Samples
2.2. Comparing T. arenaria’s Nutritional Value with That of Other Edible Mushrooms and Truffles, and Meat
2.3. Volatiles Profile by GC–MS
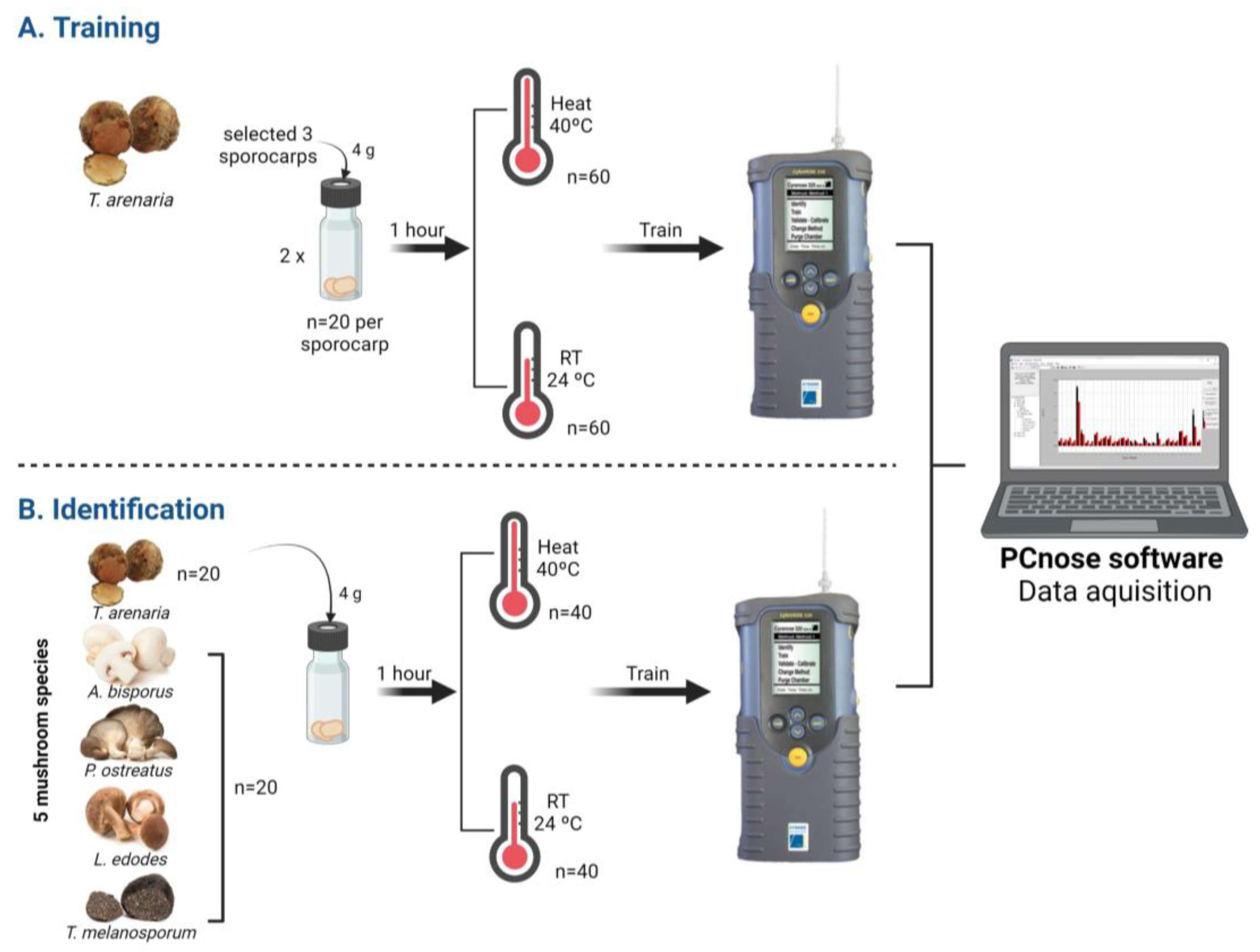
2.4. First Steps in Developing a Non-Destructive and Rapid Identification Method for T. arenaria
2.4.1. Phase 1: E-Nose Training
2.4.2. Phase 2: E-Nose Identification Accuracy
2.5. Statistical Analysis
3. Results and Discussion
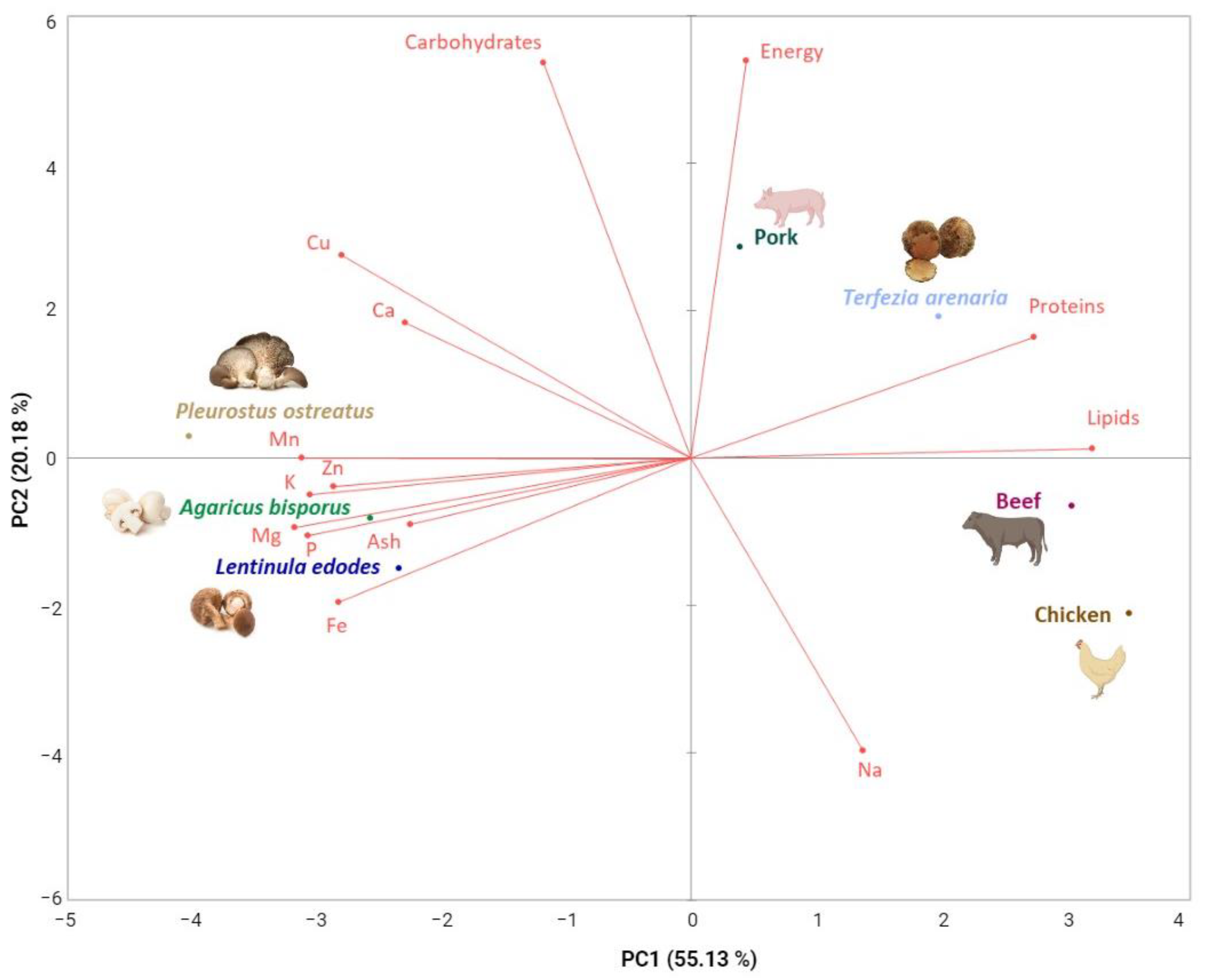
3.1. Showcasing T. arenaria’s Nutritional Value

{kind=link}
{kind=link}
{kind=link}
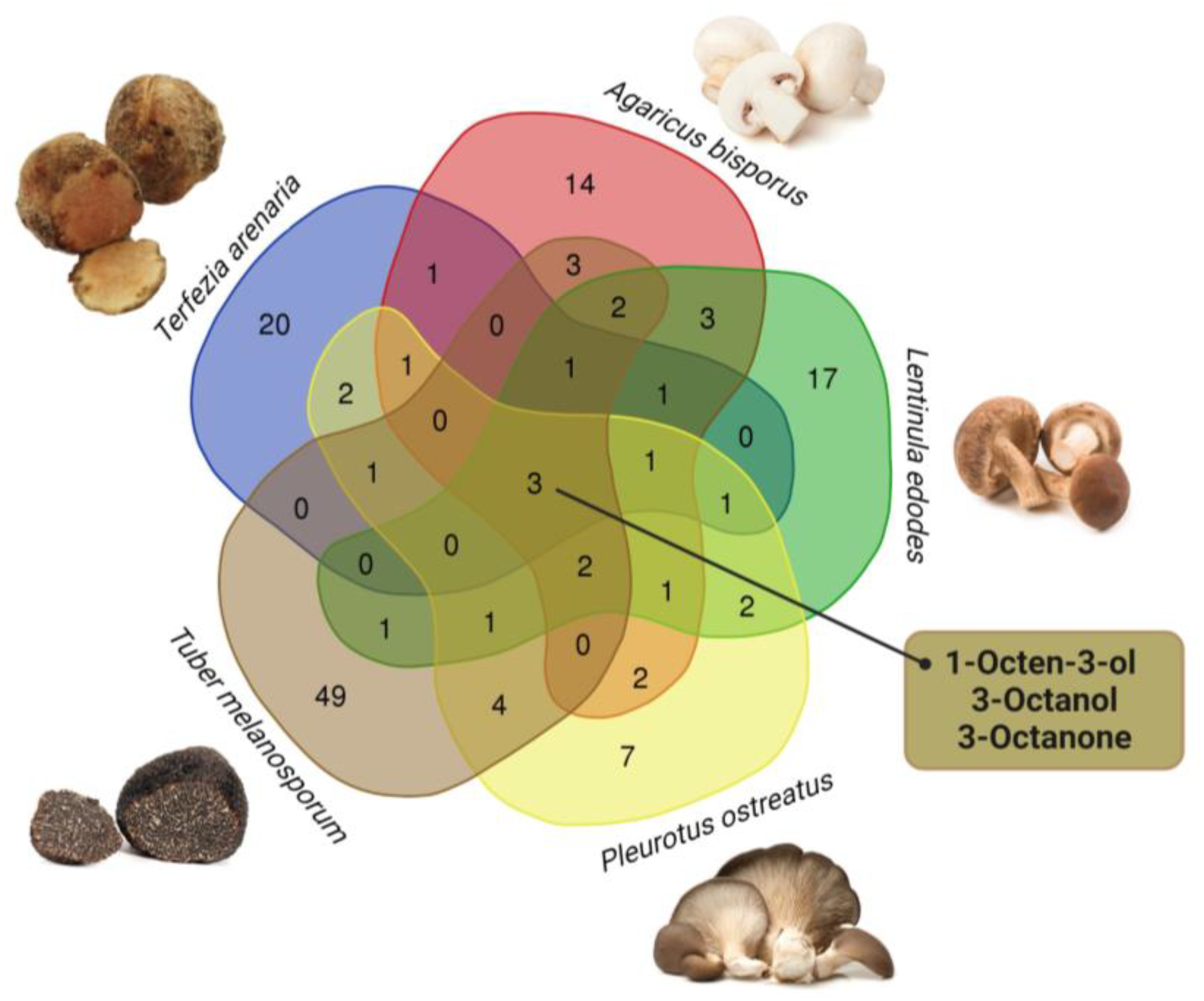{kind=link}
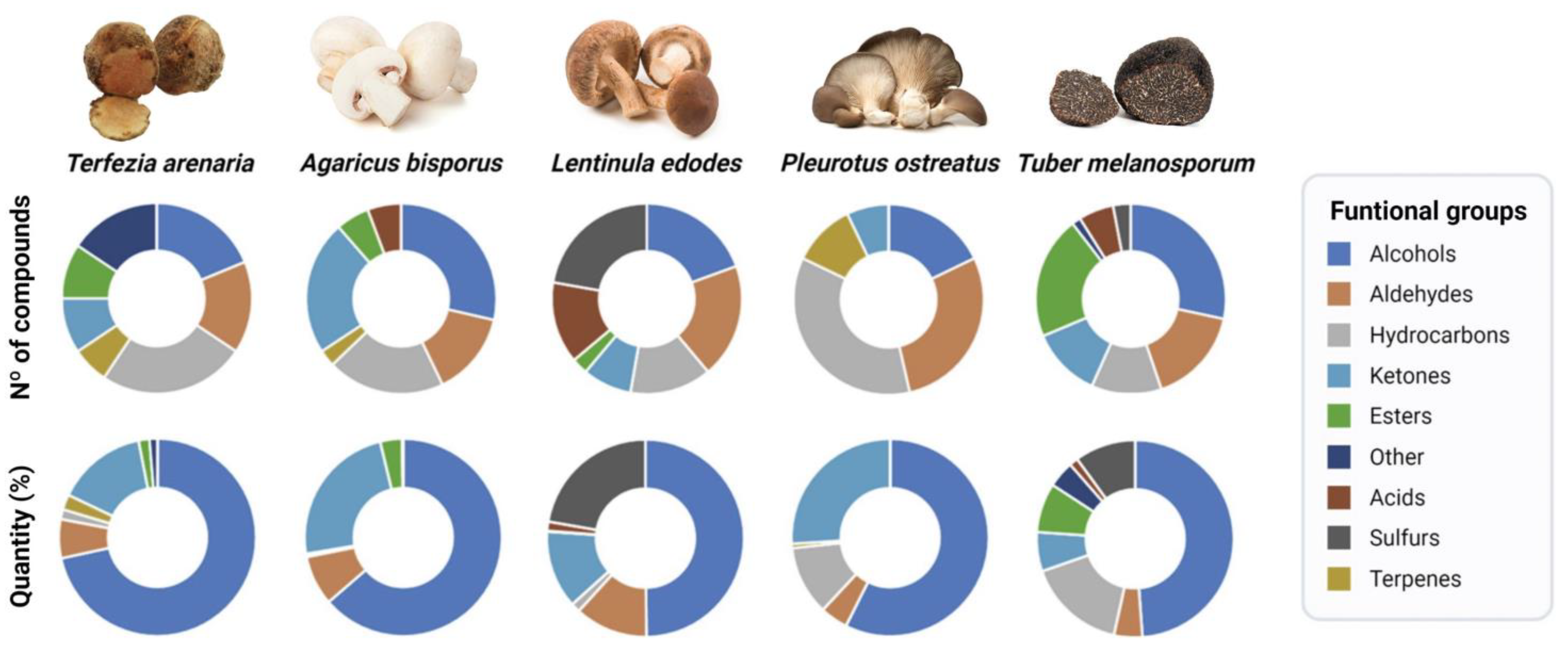{kind=link}
{kind=link}
{kind=link}
| Moisture | Ash | Proteins | Lipids | Carbohydrates | Fibre | Energy | References | ||
|---|---|---|---|---|---|---|---|---|---|
| % fw | g/100 g | g/100 g | g/100 g | g/100 g | g/100 g | kcal/100 g | |||
| Desert Truffles | Terfezia arenaria a | 77 | 7.3 | 14 | 2.2 | 67 | 10 | 387 | This study |
| n.a | 4.3 | 23 | 5.1 | 77 | n.a | 394 | [23] | ||
| Terfezia claveryi a | 73 | 4.3 | 16 | 7.0 | 65 | 8 | n.a | [68] | |
| 83 | 15.3 | 32 | 2.8 | 46 | n.a | 338 | [21] | ||
| Terfezia boudieri a | n.a | 12.9 | 17 | 6.4 | 60 | 4 | n.a | [24] | |
| 78 | 4.5 | 26 | 8.0 | 62 | n.a | n.a | [20] | ||
| Terfezia olbiensis a | 80 | 15.3 | 36 | 3.2 | 48 | n.a | 366 | [21] | |
| Commercial mushrooms | Agaricus bisporus a | 91 | 12.7 | 19 | 2.0 | 67 | 10 | 360 | [69] |
| 90 | 9.4 | 25 | 2.3 | 64 | n.a | 374 | [70] | ||
| Pleurotus ostreatus a | 91 | 7.8 | 18 | 2.6 | 71 | 14 | 382 | [69] | |
| n.a | 9.3 | 9 | 1.3 | 70 | 11 | n.a | [71] | ||
| 89 | 6.7 | 13 | 2.5 | 78 | n.a | 383 | [70] | ||
| Lentinula edodes a | n.a | 3.8 | 18 | 0.9 | 30 | 32 | 264 | [72] | |
| 94 | 6.7 | 16 | 1.8 | 74 | 15 | 382 | [69] | ||
| 88 | 7.4 | 17 | 2.1 | 73 | n.a | 381 | [70] | ||
| Tuber melanosporum a | n.a | 0.0 | 22 | 2.3 | 75 | n.a | 411 | [23] | |
| Meat | Pork b | 13 | 37.0 | 2.4 | n.a | 390 | [83] | ||
| 64 | 0.9 | 18 | 17.5 | n.a | n.a | 228 | [90] | ||
| Beef b | 20 | 4.2 | 2.0 | n.a | 126 | [83] | |||
| 63 | 0.8 | 18 | 19.4 | n.a | n.a | 243 | [90] | ||
| Chicken b | 19 | 1.3 | 9.4 | n.a | 167 | [83] | |||
| 75 | 1.0 | 18 | 7.2 | n.a | n.a | 133 | [90] |
| Minerals | Terfezia arenaria | Agaricus bisporus | Lentinula edodes | Pleurotus ostreatus | Tuber melanosporum | Pork | Beef | Chicken |
|---|---|---|---|---|---|---|---|---|
| Major essential elements | ||||||||
| Ca | 26 | 580 | 438 | 730 | 817 | 60 | 70 | 60 |
| K | 3695 | 38,400 | 21,700 | 14,244 | 7356 | 3180 | 2730 | 3020 |
| Mg | 128 | 1300 | 1330 | 2800 | 241 | 190 | 164 | 205 |
| Na | 23 | 491 | 144 | 35 | 67 | 540 | 550 | 630 |
| P | 1407 | 8210 | 4080 | 6204 | 2678 | 1730 | 1440 | 1660 |
| S | 299 | n.a | n.a | n.a | n.a | n.a | n.a | n.a |
| Essential trace elements | ||||||||
| Cr | 0.9 | 7.0 | 0.3 | n.a | n.a | n.a | n.a | n.a |
| Cu | 6.6 | 34.2 | 7.1 | 39 | 18 | 0.7 | 0.6 | 0.4 |
| Fe | 19.6 | 49.4 | 35.7 | 130 | 12 | 7.9 | 19.6 | 5.9 |
| Mn | 1.4 | 6.6 | 19.3 | 14 | 1 | <0.125 | <0.125 | 0.1 |
| Mo | 0.6 | 0.3 | 0.2 | <0.01 | n.a | n.a | n.a | n.a |
| Ni | 0.1 | 0.7 | 0.1 | 0.7 | n.a | n.a | n.a | n.a |
| Se | 0.5 | 1.7 | 1.1 | 0.3 | n.a | n.a | n.a | n.a |
| Zn | 11.0 | 51.5 | 76.3 | 110.4 | 37 | 22.3 | 38.5 | 11.8 |
| Non-essential elements | ||||||||
| Al | 10.7 | 17.9 | 5.8 | n.a | n.a | n.a | n.a | n.a |
| Li | 0.4 | <0.1 | 0.1 | 0.3 | n.a | n.a | n.a | n.a |
| Elements with detrimental health effects | ||||||||
| As | 0.1 | 0.3 | 0.5 | <0.1 | n.a | n.a | n.a | n.a |
| Ba | 0.3 | 2.8 | 1.7 | n.a | n.a | n.a | n.a | n.a |
| References | This study | [75] | [76] | [91,92,93] | [94] | [90] | [90] | [90] |
| Formula | RT | Area (%) | Name of Compounds | Funtional Groups | Pathway | Odor |
|---|---|---|---|---|---|---|
| C8H16O | 15.847 | 57.21 | 1-Octen-3-ol | Alcohols | Lipoxygenase–linoleic acid | Mushroom like |
| C8H16O | 16.007 | 12.86 | 3-Octanone | Ketones | Lipoxygenase–linoleic acid | Green apple-like |
| C6H12O | 7.412 | 4.29 | Hexanal | Aldehydes | Lipoxygenase–linoleic acid | Green, grassy |
| C8 H16 O | 17.952 | 3.22 | (Z)-2-Octen-1-ol | Alcohols | Lipoxygenase–linoleic acid | Green, citrus |
| C10H16 | 14.379 | 1.87 | α-Pinene | Terpenes | Monoterpenoid biosynthesis | Woody, resinous |
| C8H18O | 16.256 | 1.54 | 3-Octanol | Alcohols | Lipoxygenase–linoleic acid | Floral, fatty |
| C8H14O | 15.658 | 1.38 | (5Z)-Octa-1,5-dien-3-ol | Alcohols | Lipid metabolism | Sweet or floral |
| C8 H16 O2 | 16.549 | 0.98 | Pentyl propanoate | Ester | n.a | Fruity, sweet |
| C8 H14 O | 17.735 | 0.86 | I-2-Octenal | Aldehydes | Lipoxygenase–linoleic acid | Fatty, nutty |
| C21H41IO2 | 24.933 | 0.39 | Propionic acid, 3-iodo-, octadecyl ester | Ester | n.a | n.a |
| C10 H16 | 17.086 | 0.35 | Limonene | Terpenes | Monoterpenoid biosynthesis | Citrus |
| C8H9N | 18.411 | 0.32 | Pyridine, 5-ethenyl-2-methyl- | Other compounds | n.a | Pungent, fish-like |
| C18H37ClO2S | 25.156 | 0.3 | 1-Octadecanesulphonyl chloride | Other compounds | n.a | Strong and pungent |
| C21H42O2 | 25.127 | 0.29 | Henicosanoic acid | Other compounds | n.a | Odorless |
| C32H66 | 25.155 | 0.28 | Dotriacontane | Hydrocarbons | n.a | Odourless |
| C8 H16 | 17.479 | 0.26 | Caprylene (1-octene) | Hydrocarbons | n.a | Petroleum-like |
| C8H15NO3 | 20.904 | 0.23 | 2-Octanone, 1-nitro- | Ketones | n.a | Sweet |
| C12H24O3 | 23.046 | 0.22 | Propanoic acid, 2-methyl-, 3-hydroxy-2,2,4-trimethylpentyl ester | Ester | n.a | Mild, fruity or sweet |
| C8H8O | 17.417 | 0.21 | Benzeneacetaldehyde | Aldehydes | Phenylalanine metabolism | Sweet, floral |
| C14 H30 | 24.692 | 0.21 | Tetradecane | Hydrocarbons | n.a | Gasoline-like to odorless |
| C14 H30 | 24.692 | 0.21 | Eicosane-7-hexyl | Hydrocarbons | n.a | n.a |
| C13 H22 O3 Si2 | 18.831 | 0.19 | Benzaldehyde, 2,5-bis[(trimethylsilyl)oxy] | Aldehydes | n.a | n.a |
| C16H34 | 24.134 | 0.15 | Hexadecane | Hydrocarbons | Fatty acid degradation | Odourless |
| C6H13ClO | 23.201 | 0.14 | Chlorohexanol | Alcohols | n.a | Odorless |
| C10H22 | 24.654 | 0.13 | 3,3,5-Trimethylheptane | Hydrocarbons | n.a | Gasoline-like |
| C7H7NO2 | 24.979 | 0.13 | Anthranilic acid | Other compounds | L-tryptophan-kynurenine | Odorless |
| C9H18O | 20.273 | 0.12 | Nonanal | Aldehydes | n.a | Fruity, waxy |
| C8H10O2 | 23.461 | 0.11 | Tyrosol | Other compounds | Tyrosine metabolism | Floral, phenolic |
| C16H32 | 23.812 | 0.1 | 1-Dodecanol | Alcohols | n.a | Waxy, fatty |
| C20H41Cl | 25.336 | 0.1 | 1-chloroeicosane | Hydrocarbons | n.a | n.a |
| C13H22O | 23.68 | 0.09 | Geranylacetone | Ketones | Ketone Body Metabolism | Sweet, floral, fruity |
| C20 H42 | 23.201 | 0.08 | Eicosane | Hydrocarbons | n.a | Odourless |
3.2. Volatiles Profile by GC–MS
3.3. First Steps in Developing a Non-Destructive and Rapid Identification Method for T. arenaria
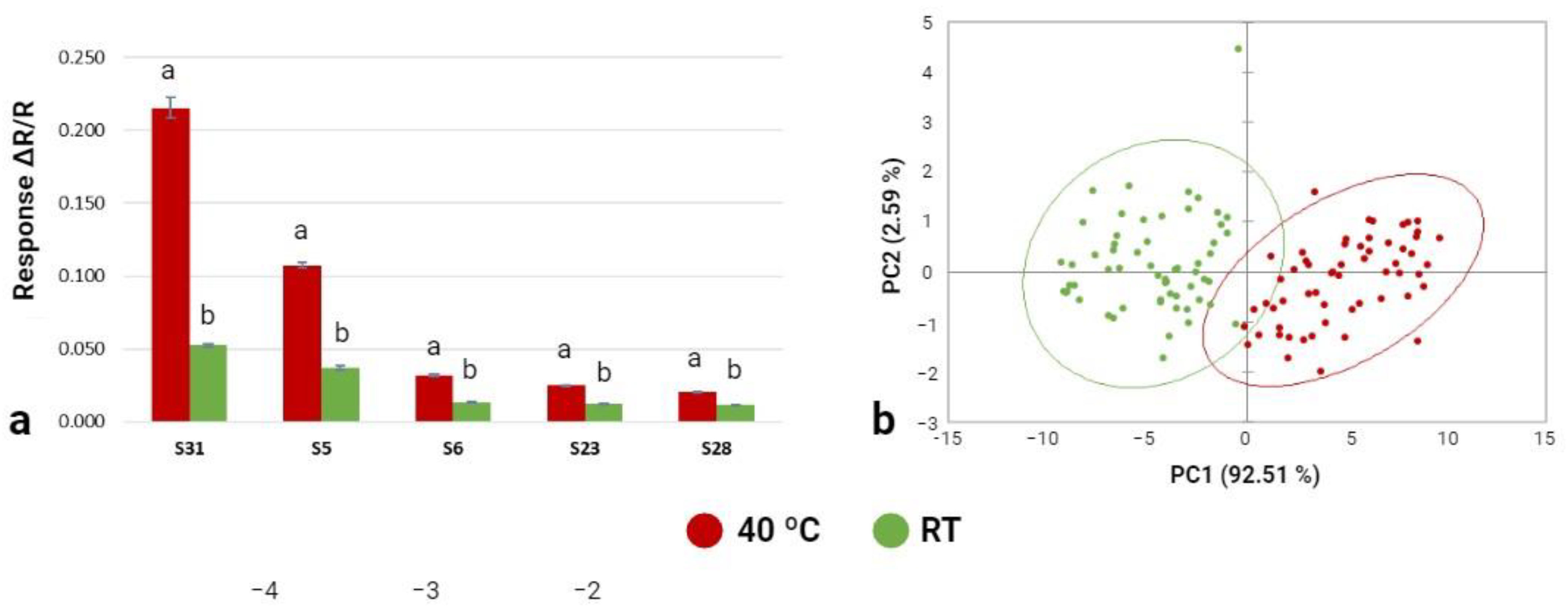
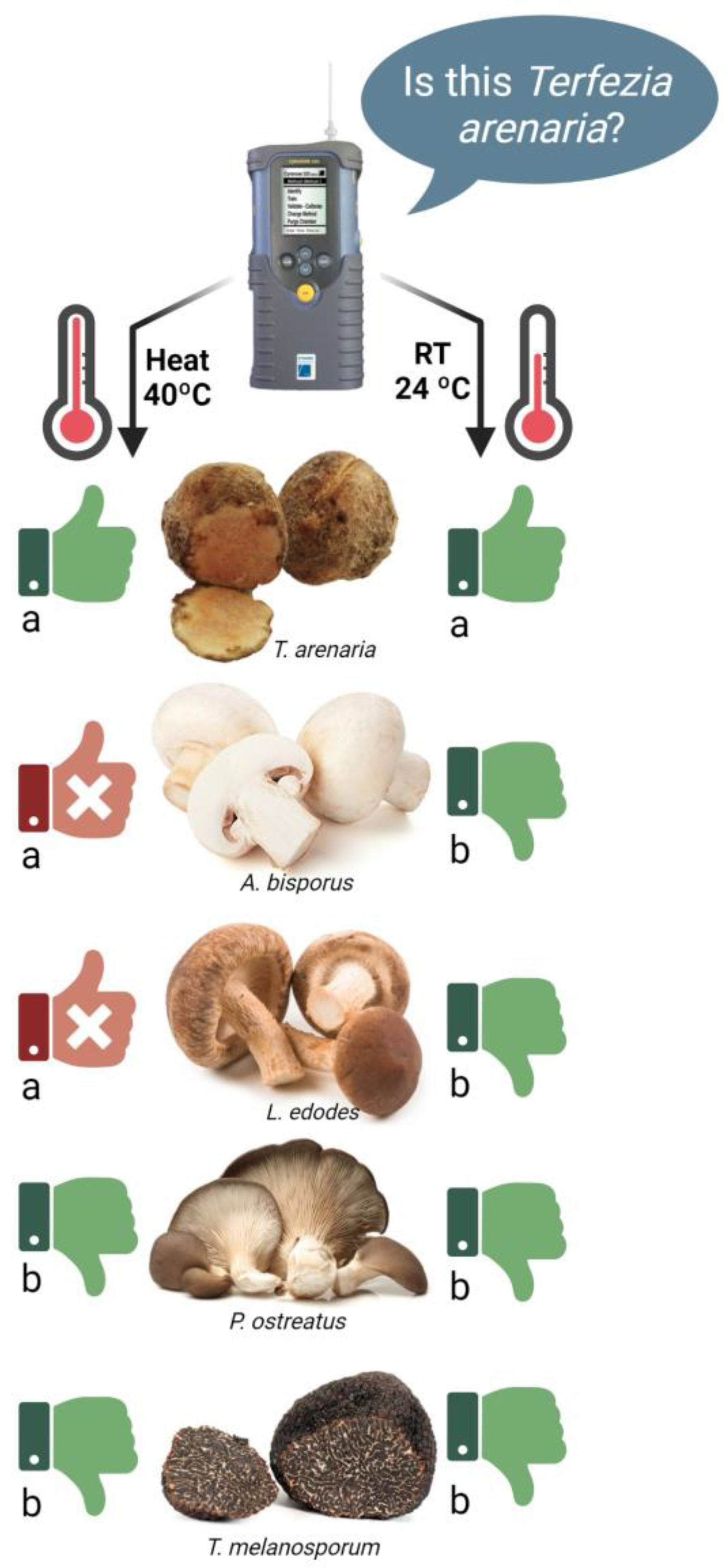
3.3.1. Phase 1: Electronic Nose Training
3.3.2. Phase 2: E-Nose Identification Accuracy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Köberle, A.C. Food Security in Climate Mitigation Scenarios. Nat. Food 2022, 3, 98–99. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.; Corrêa, A.; Cruz, C. Sustainable Production of Ectomycorrhizal Fungi in the Mediterranean Region to Support the European Green Deal. Plants People Planet 2023, 5, 14–26. [Google Scholar] [CrossRef]
- Boa, E. Wild Edible Fungi. A Global Overview of Their Use and Importance to People. In Non-Wood Forest Products; FAO: Rome, Italy, 2004; Volume 17, ISBN 9251051577. [Google Scholar]
- Ferreira, I.; Dias, T.; Cruz, C. The Potential of Ectomycorrhizal Fungi to Modulate below and Aboveground Communities May Be Mediated by 1-Octen-3-Ol. J. Fungi 2023, 9, 180. [Google Scholar] [CrossRef] [PubMed]
- de Frutos, P. Changes in World Patterns of Wild Edible Mushrooms Use Measured through International Trade Flows. For. Policy Econ. 2020, 112, 102093. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 4 August 2023).
- Pérez-Moreno, J.; Guerin-Laguette, A.; Rinaldi, A.C.; Yu, F.; Verbeken, A.; Hernández-Santiago, F.; Martínez-Reyes, M. Edible Mycorrhizal Fungi of the World: What is Their Role in Forest Sustainability, Food Security, Biocultural Conservation and Climate Change? Plants People Planet 2021, 3, 471–490. [Google Scholar] [CrossRef]
- Pérez-Moreno, J.; Mortimer, P.; Xu, J.; Karunarathna, S.; Li, H.; Pérez-Moreno, J.; Mortimer, P.; Xu, J.; Karunarathna, S.; Li, H. Global Perspectives on the Ecological, Cultural and Socioeconomic Relevance of Wild Edible Fungi. Stud. Fungi 2021, 6, 408–424. [Google Scholar] [CrossRef]
- Andrino, A.; Navarro-Ródenas, A.; Marqués-Gálvez, J.E.; Morte, A. The Crop of Desert Truffle Depends on Agroclimatic Parameters during Two Key Annual Periods. Agron. Sustain. Dev. 2019, 39, 51. [Google Scholar] [CrossRef]
- Oliach, D.; Morte, A.; Sánchez, S.; Navarro-Ródenas, A.; Marco, P.; Gutiérrez, A.; Martín-Santafé, M.; Fischer, C.; Albisu, L.M.; García-Barreda, S.; et al. Las Trufas y Las Turmas. In Los Productos Forestales no Madereros en España: Del Monte a la Industria; Sánchez-González, M., Calama, R., Bonet, J.A., Eds.; INIA, Ministerio de Economía Industria y Competitividad: Madrid, Spain, 2020; pp. 283–324. ISBN 9788474985856. [Google Scholar]
- Bradai, L.; Neffar, S.; Amrani, K.; Bissati, S.; Chenchouni, H. Ethnomycological Survey of Traditional Usage and Indigenous Knowledge on Desert Truffles among the Native Sahara Desert People of Algeria. J. Ethnopharmacol. 2015, 162, 31–38. [Google Scholar] [CrossRef]
- Shavit, E. The History of Desert Truffle Use; Springer: Berlin/Heidelberg, Germany, 2014; pp. 217–241. [Google Scholar]
- Chevalier, G. The European Desert Truffles. In Desert Truffles. Soil Biology; Kagan-Zur, V., Roth-Bejerano, N., Sitrit, Y., Morte, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 38, pp. 121–141. [Google Scholar] [CrossRef]
- Morte, A.; Arenas, F.; Marqués-Gálvez, J.E.; Andrino, A.; Guarnizo, Á.L.; Gutiérrez, A.; Berná, L.M.; Pérez-Gilabert, M.; Rodríguez, A.; Navarro-Ródenas, A. Desert Truffles (Terfezia spp.) Breeding. In Advances in Plant Breeding Strategies: Vegetable Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; Volume 10, pp. 479–504. [Google Scholar] [CrossRef]
- Shavit, E. Truffles Roasting in the Evening Fires. Pages Hist. Desert Truffles Fungi 2008, 1, 18–23. [Google Scholar]
- Gadallah, M.G.E.; Ashoush, I.S. Value Addition on Nutritional and Sensory Properties of Biscuit Using Desert Truffle (Terfezia claveryi) Powder. Food Nutr. Sci. 2016, 7, 1171–1181. [Google Scholar] [CrossRef]
- Najjaa, H.; Abdelkbir, R.; Ben Arfa, A.; Doria, E.; Tlili, H.; Zouari, N.; Neffati, M. Improved Sensory Quality and Antioxidant Capacity of Wheat Bread Supplemented with the Desert Truffle Terfezia boudieri Flour. Anal. Lett. 2021, 54, 867–883. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Farag, M.A.; Yosri, N.; Sabir, J.S.M.; Saeed, A.; Al-Mousawi, S.M.; Taha, W.; Musharraf, S.G.; Patel, S.; El-Seedi, H.R. Truffles: From Islamic Culture to Chemistry, Pharmacology, and Food Trends in Recent Times. Trends Food Sci. Technol. 2019, 91, 193–218. [Google Scholar] [CrossRef]
- Martínez-Tomé, M.; Maggi, L.; Jiménez-Monreal, A.M.; Murcia, M.A.; Marí, J.A.T. Nutritional and Antioxidant Properties of Terfezia and Picoa; Springer: Berlin/Heidelberg, Germany, 2014; pp. 261–273. [Google Scholar]
- Hamza, A.; Zouari, N.; Zouari, S.; Jdir, H.; Zaidi, S.; Gtari, M.; Neffati, M. Nutraceutical Potential, Antioxidant and Antibacterial Activities of Terfezia boudieri Chatin, a Wild Edible Desert Truffle from Tunisia Arid Zone. Arab. J. Chem. 2016, 9, 383–389. [Google Scholar] [CrossRef]
- Kıvrak, İ. Analytical Methods Applied to Assess Chemical Composition, Nutritional Value and In Vitro Bioactivities of Terfezia olbiensis and Terfezia claveryi from Turkey. Food Anal. Methods 2015, 8, 1279–1293. [Google Scholar] [CrossRef]
- Al Obaydi, M.F.; Hamed, W.M.; Al Kury, L.T.; Talib, W.H. Terfezia boudieri: A Desert Truffle With Anticancer and Immunomodulatory Activities. Front. Nutr. 2020, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Tejedor-Calvo, E.; Amara, K.; Reis, F.S.; Barros, L.; Martins, A.; Calhelha, R.C.; Venturini, M.E.; Blanco, D.; Redondo, D.; Marco, P.; et al. Chemical Composition and Evaluation of Antioxidant, Antimicrobial and Antiproliferative Activities of Tuber and Terfezia Truffles. Food Res. Int. 2021, 140, 110071. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.; Mohamed, M.A.; Hami, M.A. Libyan Truffles Terfezia boudieri Chatin: Chemical Composition and Toxicity. J. Food Sci. 1981, 46, 927–929. [Google Scholar] [CrossRef]
- Amara, K.; Reis, F.S.; Barros, L.; Skhiri, F.; Martins, A.; Ferreira, I.C.F.R. Nutritional Values, Chemical Characterization and Cytotoxicity in Human Tumor Cell Lines of Desert Truffles. In Proceedings of the 8th Journées Scientifiques Internationales sur la Valorisation des Bioressources, Monastir, Tunisia, 5–7 May 2017. [Google Scholar]
- Benaceur, F.; Chaibi, R.; Berrabah, F.; Neifar, A.; Leboukh, M.; Benaceur, K.; Nouioua, W.; Rezzoug, A.; Bouazzara, H.; Gouzi, H.; et al. Purification and Characterization of Latent Polyphenol Oxidase from Truffles (Terfezia arenaria). Int. J. Biol. Macromol. 2020, 145, 885–893. [Google Scholar] [CrossRef]
- Harir, M.; Bendif, H.; Yahiaoui, M.; Bellahcene, M.; Zohra, F.; Rodríguez-Couto, S. Evaluation of Antimicrobial Activity of Terfezia arenaria Extracts Collected from Saharan Desert against Bacteria and Filamentous Fungi. 3 Biotech 2019, 9, 281. [Google Scholar] [CrossRef]
- Moreno, G.; Alvarado, P.; Manjón, J.L. Hypogeous Desert Fungi. In Desert Truffles: Phylogeny, Physiology, Distribution and Domestication; Springer: Berlin/Heidelberg, Germany, 2013; pp. 3–20. [Google Scholar]
- Ammarellou, A.; Wang, Y.; Nematzadeh, G.; Tajick, M. Non-Mediterranean Asian Desert Countries; Springer: Berlin/Heidelberg, Germany, 2014; pp. 173–192. [Google Scholar]
- Dafri, A.; Beddiar, A. Morphological Characterisation of the Mycorrhizal Symbiosis between Tuberaria guttata (L.) Fourr and Terfezia arenaria (Moris) Trappe. Symbiosis 2018, 75, 149–154. [Google Scholar] [CrossRef]
- Brenko, A.; Vidale, E.; Oliach, D.; Marois, O.; Andrighetto, N.; Stara, K.; de Aragón, J.M.; Bonet, J.A. Short Communication: Edible Wild Mushrooms of the Northern Mediterranean Area–Sectorial Analysis and Future Perspectives. For. Syst. 2022, 31, eSC05. [Google Scholar] [CrossRef]
- Egli, S.; Peter, M.; Buser, C.; Stahel, W.; Ayer, F. Mushroom Picking Does Not Impair Future Harvests–Results of a Long-Term Study in Switzerland. Biol. Conserv. 2006, 129, 271–276. [Google Scholar] [CrossRef]
- Peintner, U.; Schwarz, S.; Mešić, A.; Moreau, P.-A.; Moreno, G.; Saviuc, P. Mycophilic or Mycophobic? Legislation and Guidelines on Wild Mushroom Commerce Reveal Different Consumption Behaviour in European Countries. PLoS ONE 2013, 8, 63926. [Google Scholar] [CrossRef] [PubMed]
- Harki, E.; Farah, A.; Bouseta, A. Volatile Compounds from Four Species of Moroccan Truffles. Vice Ed. Chief Vice Redacteur Chef 2010, 12, 10. [Google Scholar]
- Farag, M.A.; Fathi, D.; Shamma, S.; Shawkat, M.S.A.; Shalabi, S.M.; El Seedi, H.R.; Afifi, S.M. Comparative Metabolome Classification of Desert Truffles Terfezia claveryi and Terfezia boudieri via Its Aroma and Nutrients Profile. LWT 2021, 142, 111046. [Google Scholar] [CrossRef]
- Kamle, M.; Bar, E.; Lewinsohn, D.; Shavit, E.; Roth-Bejerano, N.; Kagan-Zur, V.; Barak, Z.; Guy, O.; Zaady, E.; Lewinsohn, E.; et al. Characterization of Morphology, Volatile Profiles, and Molecular Markers in Edible Desert Truffles from the Negev Desert. J. Agric. Food Chem. 2017, 65, 2977–2983. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Wen, Y.; Wu, W.; Zhang, L.; Salman Farid, M.; Shan, S.; Wen, J.; Farag, M.A.; Zhang, Y.; Zhao, C. The Flavors of Edible Mushrooms: A Comprehensive Review of Volatile Organic Compounds and Their Analytical Methods. Crc Cr Rev Food Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.M.; Angeloni, S.; Nzekoue, F.K.; Abouelenein, D.; Sagratini, G.; Caprioli, G.; Torregiani, E. An Overview on Truffle Aroma and Main Volatile Compounds. Molecules 2020, 25, 5948. [Google Scholar] [CrossRef]
- Lubes, G.; Goodarzi, M. GC–MS Based Metabolomics Used for the Identification of Cancer Volatile Organic Compounds as Biomarkers. J. Pharm. Biomed. Anal. 2018, 147, 313–322. [Google Scholar] [CrossRef]
- Zhou, Y.; Abbas, F.; Wang, Z.; Yu, Y.; Yue, Y.; Li, X.; Yu, R.; Fan, Y. HS–SPME–GC–MS and Electronic Nose Reveal Differences in the Volatile Profiles of Hedychium Flowers. Molecules 2021, 26, 5425. [Google Scholar] [CrossRef]
- Zhou, J.; Feng, T.; Ye, R. Differentiation of Eight Commercial Mushrooms by Electronic Nose and Gas Chromatography-Mass Spectrometry. J. Sens. 2015, 2015, 374013. [Google Scholar] [CrossRef]
- Guo, Q.; Adelina, N.M.; Hu, J.; Zhang, L.; Zhao, Y. Comparative Analysis of Volatile Profiles in Four Pine-Mushrooms Using HS-SPME/GC-MS and E-Nose. Food Control 2022, 134, 108711. [Google Scholar] [CrossRef]
- Gholami, R.; Aghili Nategh, N.; Rabbani, H. Evaluation the Effects of Temperature and Packaging Conditions on the Quality of Button Mushroom during Storage Using E-Nose System. J. Food Sci. Technol. 2023, 60, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Hu, Z.; Liang, M.; Song, L.; Wu, W.; Li, R.; Li, Z.; Zhang, J. Evaluation of the Flavor Compounds of Pleurotus eryngii as Affected by Baking Temperatures Using HS-SPME-GC–MS and Electronic Nose. J. Food Process. Preserv. 2022, 46, e17056. [Google Scholar] [CrossRef]
- Chilo, J.; Pelegri-Sebastia, J.; Cupane, M.; Sogorb, T. E-Nose Application to Food Industry Production. IEEE Instrum. Meas. Mag. 2016, 19, 27–33. [Google Scholar] [CrossRef]
- Falasconi, M.; Concina, I.; Gobbi, E.; Sberveglieri, V.; Pulvirenti, A.; Sberveglieri, G. Electronic Nose for Microbiological Quality Control of Food Products. Int. J. Electrochem. 2012, 2012, 715763. [Google Scholar] [CrossRef]
- Mota, I.; Teixeira-Santos, R.; Cavaleiro Rufo, J. Detection and Identification of Fungal Species by Electronic Nose Technology: A Systematic Review. Fungal Biol. Rev. 2021, 37, 59–70. [Google Scholar] [CrossRef]
- Pei, F.; Yang, W.; Ma, N.; Fang, Y.; Zhao, L.; An, X.; Xin, Z.; Hu, Q. Effect of the Two Drying Approaches on the Volatile Profiles of Button Mushroom (Agaricus bisporus) by Headspace GC–MS and Electronic Nose. LWT Food Sci. Technol. 2016, 72, 343–350. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.; Li, M.; Hao, T.; Lin, S. The Dynamic Changes in Product Attributes of Shiitake Mushroom Pilei and Stipes during Dehydration by Hot Air Drying. J. Food Process. Preserv. 2021, 45, e15648. [Google Scholar] [CrossRef]
- Ma, N.; Pei, F.; Yu, J.; Wang, S.; Ho, C.T.; Su, K.; Hu, Q. Valid Evaluation of Volatile Flavor Composition of Fresh and Dehydrated Tuber indicum with Different Drying Methods. CyTA J. Food 2018, 16, 413–421. [Google Scholar] [CrossRef]
- Song, Y.; Hu, Q.; Wu, Y.; Pei, F.; Kimatu, B.M.; Su, A.; Yang, W. Storage Time Assessment and Shelf-Life Prediction Models for Postharvest Agaricus bisporus. LWT 2019, 101, 360–365. [Google Scholar] [CrossRef]
- Portalo-Calero, F.; Arroyo, P.; Suárez, J.I.; Lozano, J. Triangular Test of Amanita Mushrooms by Using Electronic Nose and Sensory Panel. Foods 2019, 8, 414. [Google Scholar] [CrossRef]
- Portalo-Calero, F.; Lozano, J.; Meléndez, F.; Arroyo, P.; Suárez, J.I. Identification of Poisonous Mushrooms by Means of a Hand-Held Electronic Nose. Proceeding 2019, 14, 33. [Google Scholar] [CrossRef]
- Keshri, G.; Challen, M.; Elliott, T.; Magan, N. Differentiation of Agaricus Species and Other Homobasidiomycetes Based on Volatile Production Patterns Using an Electronic Nose System. Mycol. Res. 2003, 107, 609–613. [Google Scholar] [CrossRef]
- Gómez, I.; Lavega González, R.; Tejedor-Calvo, E.; Pérez Clavijo, M.; Carrasco, J. Odor Profile of Four Cultivated and Freeze-Dried Edible Mushrooms by Using Sensory Panel, Electronic Nose and GC-MS. J. Fungi 2022, 8, 953. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, M.; Adhikari, B. Advances of Electronic Nose and Its Application in Fresh Foods: A Review. Crit. Rev. Food Sci. Nutr. 2017, 58, 2700–2710. [Google Scholar] [CrossRef] [PubMed]
- AOAC. AOAC Official Methods of Analysis, 15th ed.; Association of Official Agricultural Chemists: Washington, DC, USA, 1990; pp. 136–138. [Google Scholar]
- Mędyk, M.; Chudzińska, M.; Barałkiewicz, D.; Falandysz, J. Specific Accumulation of Cadmium and Other Trace Elements in Sarcodon imbricatus Using ICP-MS with a Chemometric Approach. J Environ Sci Heal B 2017, 52, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Medicine, I. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; The National Academies Press: Washington, DC, USA, 2005; ISBN 978-0-309-08525-0. [Google Scholar]
- SCHER Assessment of the Tolerable Daily Intake of Barium; European Commission Director General Health and Consumer: Brussels, Belgium, 2012.
- Scientific Opinion on Arsenic in Food. EFSA J. 2009, 7, 1351. [CrossRef]
- Dietary Reference Values for Nutrients Summary Report. EFSA Support. Publ. 2017, 14, e15121E. [CrossRef]
- Turck, D.; Bohn, T.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Knutsen, H.K.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Peláez, C.; et al. Scientific Opinion on the Tolerable Upper Intake Level for Selenium. EFSA J. 2023, 21, e07704. [Google Scholar] [CrossRef]
- Nutri-Score. Available online: https://www.santepubliquefrance.fr/en/nutri-score (accessed on 9 August 2023).
- Splivallo, R.; Ebeler, S.E. Sulfur Volatiles of Microbial Origin Are Key Contributors to Human-Sensed Truffle Aroma. Appl. Microbiol. Biotechnol. 2015, 99, 2583–2592. [Google Scholar] [CrossRef] [PubMed]
- Sensigents. Available online: https://www.sensigent.com/products/cyranose.html (accessed on 5 June 2023).
- Santos, J.P.; Lozano, J.L.; Aleixandre, M.; Sayago, I.; Fernández, M.J.; Arés, L.; Gutiérrez, J.; Horrillo, M.D.C. Discrimination of Different Aromatic Compounds in Water, Ethanol and Wine with a Thin Film Sensor Array. Sens. Actuators B Chem. 2004, 103, 98–103. [Google Scholar] [CrossRef]
- Murcia, M.A.; Martínez-Tomé, M.; Vera, A.; Morte, A.; Gutierrez, A.; Honrubia, M.; Jiménez, A.M. Effect of Industrial Processing on Desert Truffles Terfezia claveryi Chatin and Picoa juniperi Vittadini): Proximate Composition and Fatty Acids. J. Sci. Food Agric. 2003, 83, 535–541. [Google Scholar] [CrossRef]
- Jacinto-Azevedo, B.; Valderrama, N.; Henríquez, K.; Aranda, M.; Aqueveque, P. Nutritional Value and Biological Properties of Chilean Wild and Commercial Edible Mushrooms. Food Chem. 2021, 356, 129651. [Google Scholar] [CrossRef] [PubMed]
- Roncero-Ramos, I.; Mendiola-Lanao, M.; Pérez-Clavijo, M.; Delgado-Andrade, C. Effect of Different Cooking Methods on Nutritional Value and Antioxidant Activity of Cultivated Mushrooms. Int J Food Sci Nutr. 2016, 68, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Ekute, B. Nutritional Profile of Two Nigerian Edible Mushrooms: Pleurotus ostreatus and Pleurotus pulmonarius. J. Appl. Sci. Environ. Manag. 2019, 22, 1745–1747. [Google Scholar] [CrossRef]
- Yu, Q.; Guo, M.; Zhang, B.; Wu, H.; Zhang, Y.; Zhang, L. Analysis of Nutritional Composition in 23 Kinds of Edible Fungi. J. Food Qual. 2020, 2020, 8821315. [Google Scholar] [CrossRef]
- Kalač, P. A Review of Chemical Composition and Nutritional Value of Wild-Growing and Cultivated Mushrooms. J. Sci. Food Agric. 2013, 93, 209–218. [Google Scholar] [CrossRef]
- Rybakowski, J.K.; Ferensztajn-Rochowiak, E. Mini-Review: Anomalous Association between Lithium Data and Lithium Use. Neurosci. Lett. 2022, 777, 136590. [Google Scholar] [CrossRef]
- Siwulski, M.; Niedzielski, P.; Budka, A.; Budzyńska, S.; Kuczyńska-Kippen, N.; Kalač, P.; Sobieralski, K.; Mleczek, M. Patterns of Changes in the Mineral Composition of Agaricus Bisporus Cultivated in Poland between 1977 and 2020. J. Food Compos. Anal. 2022, 112, 104660. [Google Scholar] [CrossRef]
- Siwulski, M.; Budka, A.; Budzyńska, S.; Gąsecka, M.; Kalač, P.; Niedzielski, P.; Mleczek, M. Mineral Composition of Traditional and Organic-Cultivated Mushroom Lentinula edodes in Europe and Asia–Similar or Different? LWT 2021, 147, 111570. [Google Scholar] [CrossRef]
- Kalac, P. Mineral Composition and Radioactivity of Edible Mushrooms; Academic Press: Cambridge, MA, USA, 2019; ISBN 0128176067. [Google Scholar]
- Manzi, P.; Gambelli, L.; Marconi, S.; Vivanti, V.; Pizzoferrato, L. Nutrients in Edible Mushrooms: An Inter-Species Comparative Study. Food Chem. 1999, 65, 477–482. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current Overview of Mushroom Production in the World. In Edible and Medicinal Mushrooms; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 5–13. [Google Scholar]
- Reyna, S.; Garcia-Barreda, S. Black Truffle Cultivation: A Global Reality. For. Syst. 2014, 23, 317–328. [Google Scholar] [CrossRef]
- WHO. Global Action Plan for the Prevention and Control of Noncommunicable Diseases 2013–2020; World Health Organization: Geneva, Switzerland, 2013; p. 102. ISBN 9789241506236.
- van den Akker, K.; Bartelet, D.; Brouwer, L.; Luijpers, S.; Nap, T.; Havermans, R. The Impact of the Nutri-Score on Food Choice: A Choice Experiment in a Dutch Supermarket. Appetite 2022, 168, 105664. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, R. A Review on Nutritional Advantages of Edible Mushrooms and Its Industrialization Development Situation in Protein Meat Analogues. J. Future Foods 2023, 3, 1–7. [Google Scholar] [CrossRef]
- Kwasny, T.; Dobernig, K.; Riefler, P. Towards Reduced Meat Consumption: A Systematic Literature Review of Intervention Effectiveness, 2001–2019. Appetite 2022, 168, 105739. [Google Scholar] [CrossRef] [PubMed]
- Andreani, G.; Sogari, G.; Marti, A.; Froldi, F.; Dagevos, H.; Martini, D. Plant-Based Meat Alternatives: Technological, Nutritional, Environmental, Market, and Social Challenges and Opportunities. Nutrients 2023, 15, 452. [Google Scholar] [CrossRef] [PubMed]
- Guinard, J.X.; Myrdal Miller, A.; Mills, K.; Wong, T.; Lee, S.M.; Sirimuangmoon, C.; Schaefer, S.E.; Drescher, G. Consumer Acceptance of Dishes in Which Beef Has Been Partially Substituted with Mushrooms and Sodium has been Reduced. Appetite 2016, 105, 449–459. [Google Scholar] [CrossRef]
- Li, J.; Silver, C.; Gómez, M.I.; Milstein, M.; Sogari, G. Factors Influencing Consumer Purchase Intent for Meat and Meat Substitutes. Future Foods 2023, 7, 100236. [Google Scholar] [CrossRef]
- Lang, M. Consumer Acceptance of Blending Plant-Based Ingredients into Traditional Meat-Based Foods: Evidence from the Meat-Mushroom Blend. Food Qual. Prefer. 2020, 79, 103758. [Google Scholar] [CrossRef]
- De Cianni, R.; Pippinato, L.; Mancuso, T. A Systematic Review on Drivers Influencing Consumption of Edible Mushrooms and Innovative Mushroom-Containing Products. Appetite 2023, 182, 106454. [Google Scholar] [CrossRef] [PubMed]
- Food Data Central. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/1999627/nutrients (accessed on 11 May 2023).
- Koutrotsios, G.; Danezis, G.; Georgiou, C.; Zervakis, G.I. Elemental Content in Pleurotus Ostreatus and Cyclocybe cylindracea Mushrooms: Correlations with Concentrations in Cultivation Substrates and Effects on the Production Process. Molecules 2020, 25, 2179. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Zakil, F.; Xuan, L.H.; Zaman, N.; Alan, N.I.; Salahutheen, N.A.A.; Sueb, M.S.M.; Isha, R. Growth Performance and Mineral Analysis of Pleurotus Ostreatus from Various Agricultural Wastes Mixed with Rubber Tree Sawdust in Malaysia. Bioresour. Technol. Rep. 2022, 17, 100873. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.L.; Wang, C.H. The Effects of Different Substrates on the Growth, Yield, and Nutritional Composition of Two Oyster Mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2018, 43, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, T.; Kinoshita, A.; Kusumoto, N.; Nakano, S.; Nakamura, N.; Yamanaka, T. Component Features, Odor-Active Volatiles, and Acute Oral Toxicity of Novel White-Colored Truffle Tuber japonicum Native to Japan. Food Sci. Nutr. 2020, 8, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Rodriguez, E.; Rivera-Macias, L.E.; Adame-Alvarez, R.M.; Torres, J.M.; Heil, M. Shared Weapons in Fungus-Fungus and Fungus-Plant Interactions? Volatile Organic Compounds of Plant or Fungal Origin Exert Direct Antifungal Activity In Vitro. Fungal Ecol. 2018, 33, 115–121. [Google Scholar] [CrossRef]
- Combet, E.; Henderson, J.; Eastwood, D.C.; Burton, K.S. Eight-Carbon Volatiles in Mushrooms and Fungi: Properties, Analysis, and Biosynthesis. Mycoscience 2006, 47, 317–326. [Google Scholar] [CrossRef]
- Maga, J.A. Mushroom Flavor. J. Agric. Food Chem. 1981, 29, 1–4. [Google Scholar] [CrossRef]
- Zhang, H.; Peng, J.; Zhang, Y.R.; Liu, Q.; Pan, L.Q.; Tu, K. Discrimination of Volatiles of Shiitakes (Lentinula Edodes) Produced during Drying Process by Electronic Nose. Int. J. Food Eng. 2020, 16, 20190233. [Google Scholar] [CrossRef]
- Feng, T.; Yang, M.; Ma, B.; Zhao, Y.; Zhuang, H.; Zhang, J.; Chen, D. Volatile Profiles of Two Genotype Agaricus Bisporus Species at Different Growth Stages. Food Res. Int. 2021, 140, 109761. [Google Scholar] [CrossRef]
- Tagkouli, D.; Bekiaris, G.; Pantazi, S.; Anastasopoulou, M.E.; Koutrotsios, G.; Mallouchos, A.; Zervakis, G.I.; Kalogeropoulos, N. Volatile Profiling of Pleurotus eryngii and Pleurotus ostreatus Cultivated on Agricultural and Agro-Industrial by-Products. Foods 2021, 10, 1287. [Google Scholar] [CrossRef] [PubMed]
- Tasaki, Y.; Kobayashi, D.; Sato, R.; Hayashi, S.; Joh, T. Variations in 1-Octen-3-Ol and Lipoxygenase Gene Expression in the Oyster Mushroom Pleurotus Ostreatus According to Fruiting Body Development, Tissue Specificity, Maturity, and Postharvest Storage. Mycoscience 2019, 60, 170–176. [Google Scholar] [CrossRef]
- Splivallo, R.; Ottonello, S.; Mello, A.; Karlovsky, P. Truffle Volatiles: From Chemical Ecology to Aroma Biosynthesis. New Phytol. 2011, 189, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.B.; Zhang, Z.Y.; Xin, G.; Sun, B.X.; Bao, X.J.; Wei, Y.Y.; Zhao, X.M.; Xu, H.R. Advances in Umami Taste and Aroma of Edible Mushrooms. Trends Food Sci. Technol. 2020, 96, 176–187. [Google Scholar] [CrossRef]
- Czapski, G.A.; Czubowicz, K.; Strosznajder, R.P. Evaluation of the Antioxidative Properties of Lipoxygenase Inhibitors. Pharmacol. Rep. 2012, 64, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Nkadimeng, S.M.; Steinmann, C.M.L.; Eloff, J.N. Anti-Inflammatory Effects of Four Psilocybin-Containing Magic Mushroom Water Extracts in Vitro on 15-Lipoxygenase Activity and on Lipopolysaccharide-Induced Cyclooxygenase-2 and Inflammatory Cytokines in Human U937 Macrophage Cells. J. Inflamm. Res. 2021, 14, 3729. [Google Scholar] [CrossRef] [PubMed]
- Mashima, R.; Okuyama, T. The Role of Lipoxygenases in Pathophysiology; New Insights and Future Perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef]
- Szydłowska-Tutaj, M.; Szymanowska, U.; Tutaj, K.; Domagała, D.; Złotek, U. The Addition of Reishi and Lion’s Mane Mushroom Powder to Pasta Influences the Content of Bioactive Compounds and the Antioxidant, Potential Anti-Inflammatory, and Anticancer Properties of Pasta. Antioxidants 2023, 12, 738. [Google Scholar] [CrossRef]
- Szydłowska-Tutaj, M.; Szymanowska, U.; Tutaj, K.; Domagała, D.; Złotek, U. Influence of Addition of Dried Maitake and Enoki Mushrooms on Antioxidant, Potentially Anti-Inflammatory, and Anti-Cancer Properties of Enriched Pasta. Appl. Sci. 2023, 13, 8183. [Google Scholar] [CrossRef]
- Darwish, R.S.; Shawky, E.; Nassar, K.M.; Rashad ElSayed, R.M.; Hussein, D.E.; Ghareeb, D.A.; El Sohafy, S.M. Differential Anti-Inflammatory Biomarkers of the Desert Truffles Terfezia claveryi and Tirmania nivea Revealed via UPLC-QqQ-MS-Based Metabolomics Combined to Chemometrics. LWT 2021, 150, 111965. [Google Scholar] [CrossRef]
- Choo, K.S.O.; Bollen, M.; Dykes, G.A.; Coorey, R. Aroma-Volatile Profile and Its Changes in Australian Grown Black Périgord Truffle (Tuber melanosporum) during Storage. Int. J. Food Sci. Technol. 2021, 56, 5762–5776. [Google Scholar] [CrossRef]
- Mohd Ali, M.; Hashim, N.; Abd Aziz, S.; Lasekan, O. Principles and Recent Advances in Electronic Nose for Quality Inspection of Agricultural and Food Products. Trends Food Sci. Technol. 2020, 99, 1–10. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, I.; Dias, T.; Mouazen, A.M.; Cruz, C. Using Science and Technology to Unveil The Hidden Delicacy Terfezia arenaria, a Desert Truffle. Foods 2023, 12, 3527. https://doi.org/10.3390/foods12193527
Ferreira I, Dias T, Mouazen AM, Cruz C. Using Science and Technology to Unveil The Hidden Delicacy Terfezia arenaria, a Desert Truffle. Foods. 2023; 12(19):3527. https://doi.org/10.3390/foods12193527
Chicago/Turabian StyleFerreira, Inês, Teresa Dias, Abdul M. Mouazen, and Cristina Cruz. 2023. "Using Science and Technology to Unveil The Hidden Delicacy Terfezia arenaria, a Desert Truffle" Foods 12, no. 19: 3527. https://doi.org/10.3390/foods12193527
APA StyleFerreira, I., Dias, T., Mouazen, A. M., & Cruz, C. (2023). Using Science and Technology to Unveil The Hidden Delicacy Terfezia arenaria, a Desert Truffle. Foods, 12(19), 3527. https://doi.org/10.3390/foods12193527