

Exploiting Potential Probiotic Lactic Acid Bacteria Isolated from Chlorella vulgaris Photobioreactors as Promising Vitamin B12 Producers
 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Lactic Acid Bacteria from Photobioreactor’s Microbiota
2.2. Genetic Identification of Bacterial Strains
2.3. Determination of Vitamin B12 Biosynthesis by Lactic Acid Bacteria
2.4. Next-Generation Sequencing (NGS) and Bioinformatics Analysis
2.5. Assessment of Lactic Acid Bacteria Probiotic Potential
2.5.1. In Vitro Gastrointestinal Survival Analysis
2.5.2. Evaluation of the Safety Profile of B12-Producing Strains
Antibiotic Susceptibility Evaluation
Virulence Genes, Hemolytic Activity, and Biogenic Amine Production
2.6. Statistical Analysis
3. Results and Discussion
3.1. Identification of Lactic Acid Bacteria
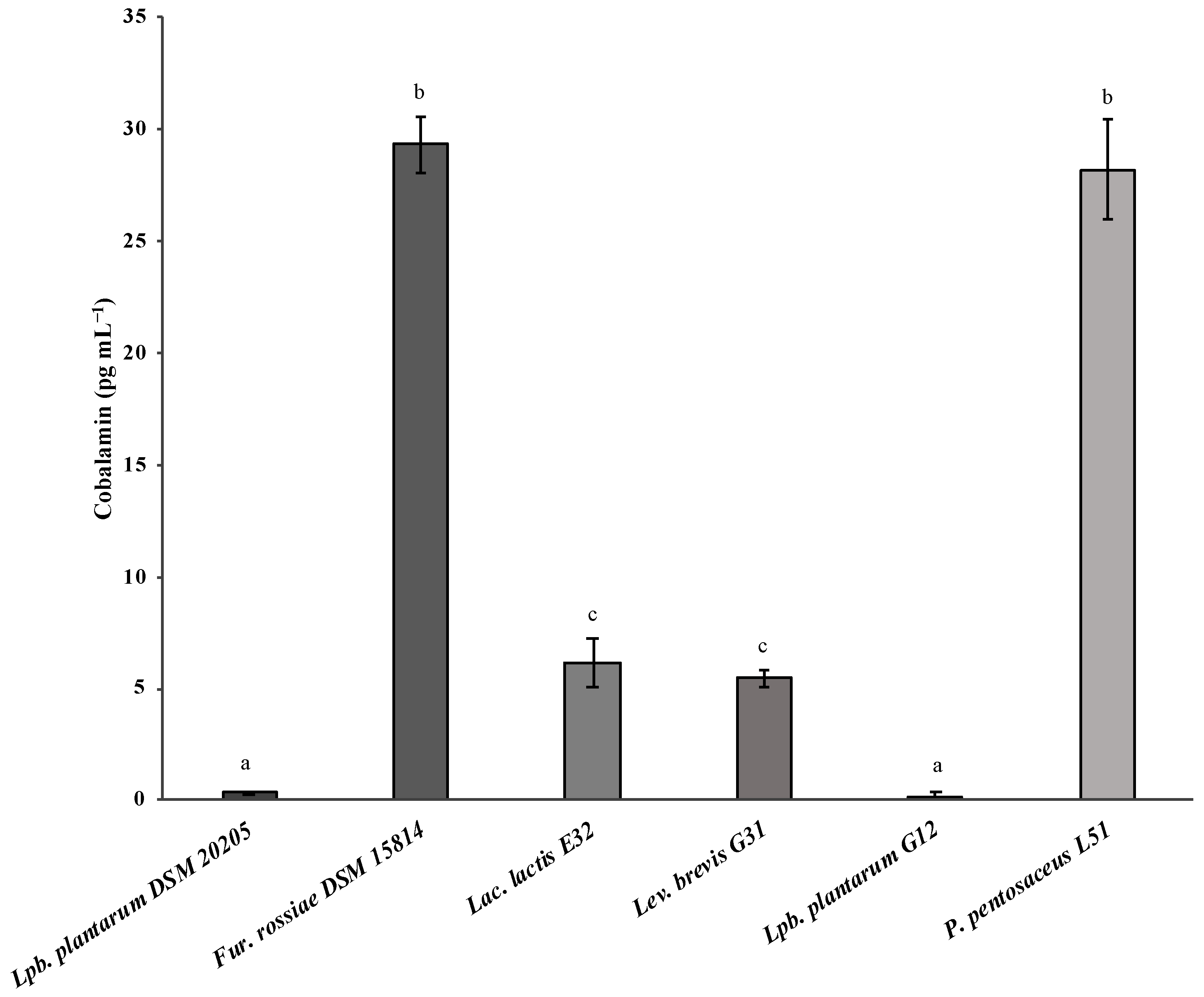
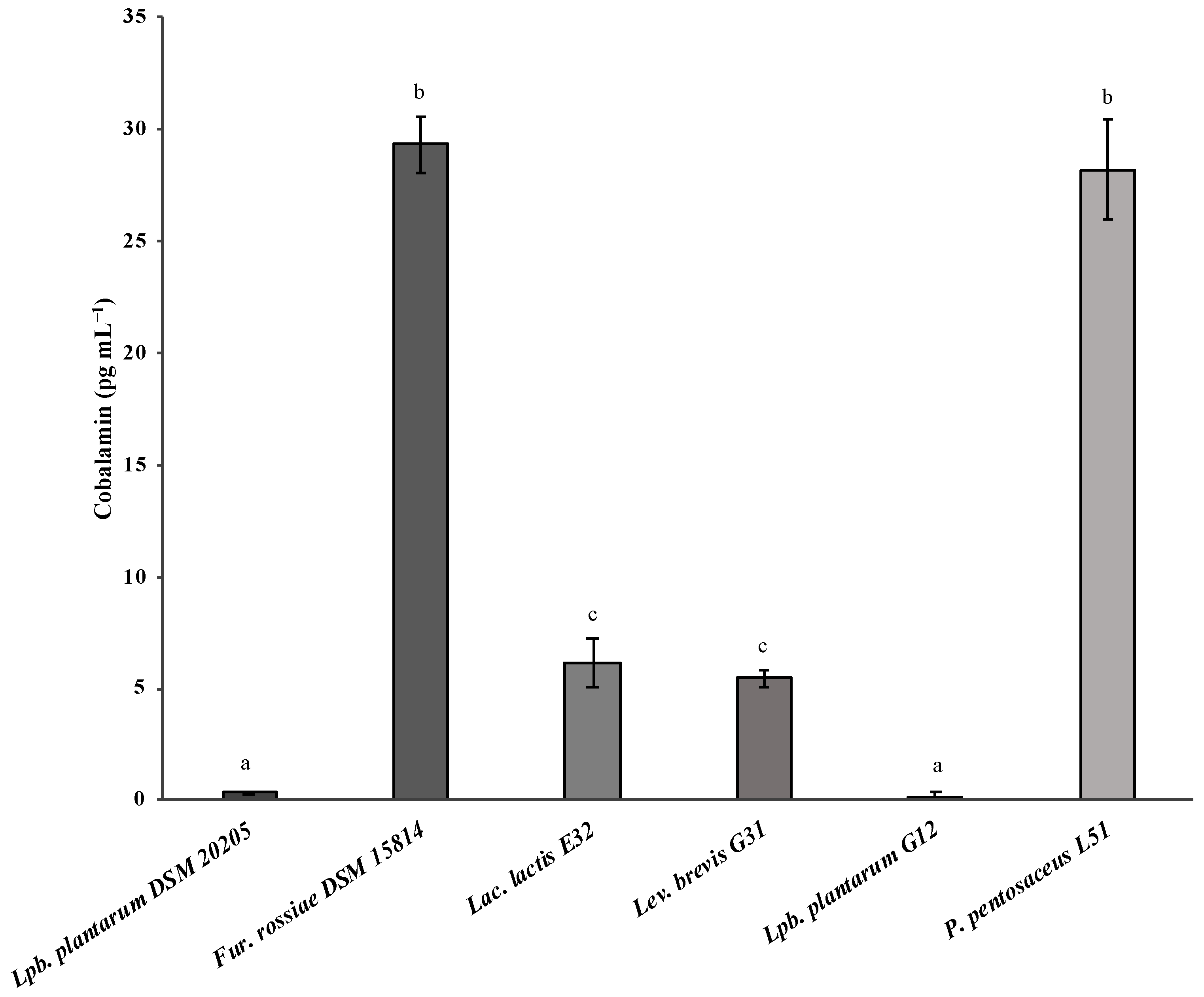
3.2. Evaluation of Vitamin B12 Biosynthesis by Lactic Acid Bacteria
3.3. Genotypic Confirmation of Vitamin B12 Biosynthesis
3.4. Lactic Acid Bacteria Probiotic Potential
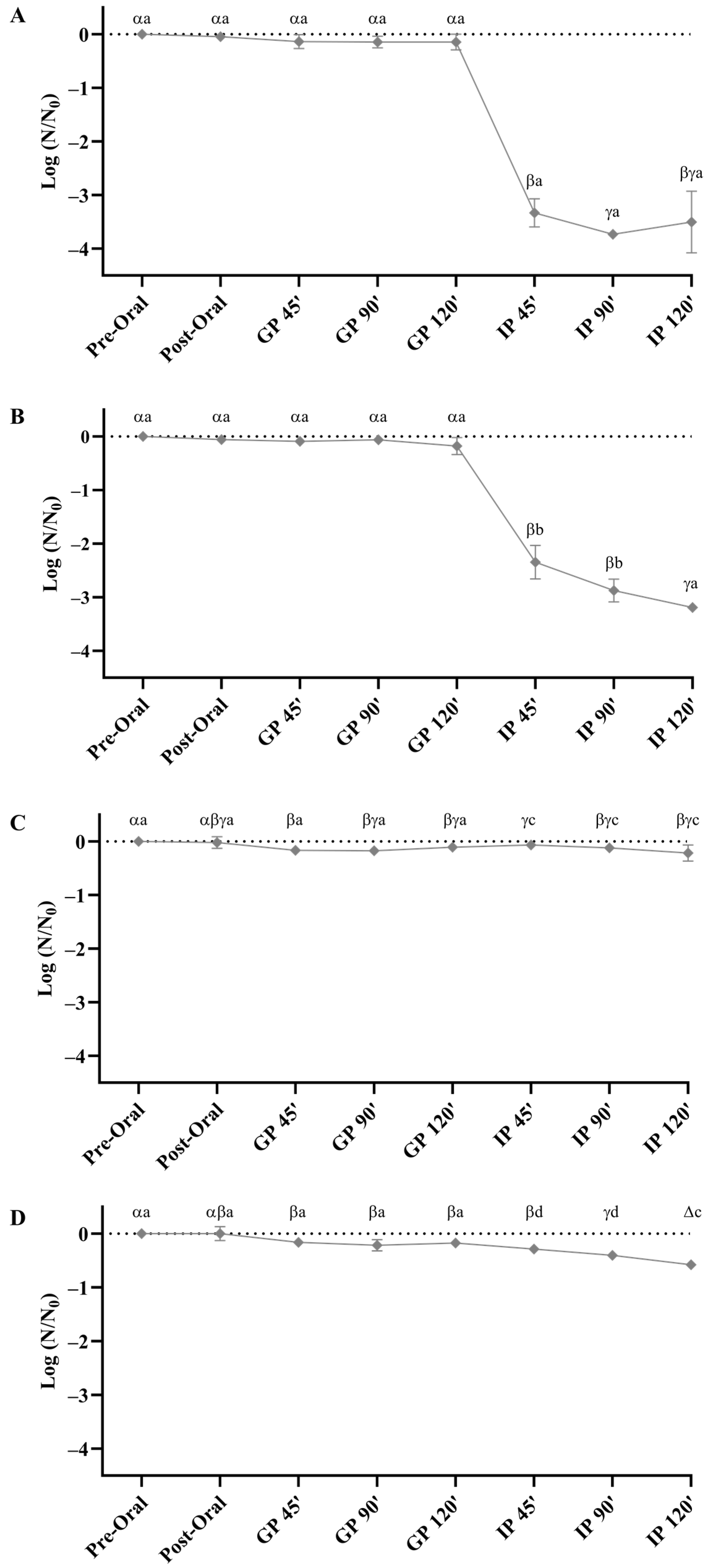
3.4.1. Evaluation of In Vitro Gastrointestinal Survival
3.4.2. Evaluation of the Safety Profile of the B12-Producing Strains
Antibiotic Susceptibility Evaluation
Virulence Genes, Hemolytic Activity, and Biogenic Amine Production Evaluation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watanabe, F.; Bito, T. Vitamin B(12) Sources and Microbial Interaction. Exp. Biol. Med. (Maywood) 2018, 243, 148–158. [Google Scholar] [CrossRef]
- Mądry, E.; Lisowska, A.; Grebowiec, P.; Walkowiak, J. The Impact of Vegan Diet on B-12 Status in Healthy Omnivores: Five-Year Prospective Study. Acta Sci. Pol. Technol. Aliment. 2012, 11, 209–212. [Google Scholar]
- Shipton, M.J.; Thachil, J. Vitamin B12 Deficiency—A 21st Century Perspective. Clin. Med. 2015, 15, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Grimley Evans, J.; Schneede, J.; Nexo, E.; Bates, C.; Fletcher, A.; Prentice, A.; Johnston, C.; Ueland, P.M.; Refsum, H.; et al. Vitamin B12 and Folate Deficiency in Later Life. Age Ageing 2004, 33, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, R.; Lester, S.E.; Babatunde, T. The Prevalence of Cobalamin Deficiency among Vegetarians Assessed by Serum Vitamin B12: A Review of Literature. Eur. J. Clin. Nutr. 2014, 68, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, G.; Laganà, A.S.; Rapisarda, A.M.C.; La Ferrera, G.M.G.; Buscema, M.; Rossetti, P.; Nigro, A.; Muscia, V.; Valenti, G.; Sapia, F.; et al. Vitamin B12 among Vegetarians: Status, Assessment and Supplementation. Nutrients 2016, 8, 767. [Google Scholar] [CrossRef] [PubMed]
- Langan, R.C.; Goodbred, A.J. Vitamin B12 Deficiency: Recognition and Management. Am. Fam. Physician 2017, 96, 384–389. [Google Scholar]
- Martens, J.H.; Barg, H.; Warren, M.J.; Jahn, D. Microbial production of vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef]
- Warren, M.J.; Raux, E.; Schubert, H.L.; Escalante-Semerena, J.C. The Biosynthesis of Adenosylcobalamin (Vitamin B12). Nat. Prod. Rep. 2002, 19, 390–412. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Kang, J.; Zhang, D. Microbial Production of Vitamin B(12): A Review and Future Perspectives. Microb. Cell Fact. 2017, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Raux, E.; Schubert, H.L.; Roper, J.M.; Wilson, K.S.; Warren, M.J. Vitamin B12: Insights into Biosynthesis’s Mount Improbable. Bioorg. Chem. 1999, 27, 100–118. [Google Scholar] [CrossRef]
- Bykhovskiĭ, V.I.; Zaĭtseva, N.I.; Eliseev, A.A. Tetrapyrroles: Diversity, biosynthesis, biotechnology. Prikl. Biokhim. Mikrobiol. 1998, 34, 3–21. [Google Scholar] [PubMed]
- Kang, Z.; Zhang, J.; Zhou, J.; Qi, Q.; Du, G.; Chen, J. Recent Advances in Microbial Production of δ-Aminolevulinic Acid and Vitamin B12. Biotechnol. Adv. 2012, 30, 1533–1542. [Google Scholar] [CrossRef]
- Taranto, M.P.; Vera, J.L.; Hugenholtz, J.; De Valdez, G.F.; Sesma, F. Lactobacillus reuteri CRL1098 Produces Cobalamin. J. Bacteriol. 2003, 185, 5643–5647. [Google Scholar] [CrossRef]
- Santos, F.; Vera, J.L.; Lamosa, P.; de Valdez, G.F.; de Vos, W.M.; Santos, H.; Sesma, F.; Hugenholtz, J. Pseudovitamin B(12) Is the Corrinoid Produced by Lactobacillus reuteri CRL1098 under Anaerobic Conditions. FEBS Lett. 2007, 581, 4865–4870. [Google Scholar] [CrossRef]
- Santos, F.; Vera, J.L.; van der Heijden, R.; Valdez, G.; de Vos, W.M.; Sesma, F.; Hugenholtz, J. The Complete Coenzyme B12 Biosynthesis Gene Cluster of Lactobacillus reuteri CRL1098. Microbiology 2008, 154, 81–93. [Google Scholar] [CrossRef]
- De Angelis, M.; Bottacini, F.; Fosso, B.; Kelleher, P.; Calasso, M.; Di Cagno, R.; Ventura, M.; Picardi, E.; van Sinderen, D.; Gobbetti, M. Lactobacillus rossiae, a Vitamin B12 Producer, Represents a Metabolically Versatile Species within the Genus Lactobacillus. PLoS ONE 2014, 9, e107232. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Laiño, J.E.; del Valle, M.J.; Vannini, V.; van Sinderen, D.; Taranto, M.P.; de Valdez, G.F.; de Giori, G.S.; Sesma, F. B-Group Vitamin Production by Lactic Acid Bacteria-Current Knowledge and Potential Applications. J. Appl. Microbiol. 2011, 111, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Russo, P.; Dueñas, M.T.; López, P.; Spano, G. Lactic Acid Bacteria Producing B-Group Vitamins: A Great Potential for Functional Cereals Products. Appl. Microbiol. Biotechnol. 2012, 96, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Olivares, M.; Marín, M.L.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Characterization of a Reuterin-Producing Lactobacillus coryniformis Strain Isolated from a Goat’s Milk Cheese. Int. J. Food Microbiol. 2005, 104, 267–277. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production Potency of Folate, Vitamin B(12), and Thiamine by Lactic Acid Bacteria Isolated from Japanese Pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef] [PubMed]
- Basavanna, G.; Prapulla, S.G. Evaluation of Functional Aspects of Lactobacillus fermentum CFR 2195 Isolated from Breast Fed Healthy Infants’ Fecal Matter. J. Food Sci. Technol. 2013, 50, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.C.; Vannini, V.; Bonacina, J.; Font, G.; Saavedra, L.; Taranto, M.P. Cobalamin Production by Lactobacillus coryniformis: Biochemical Identification of the Synthetized Corrinoid and Genomic Analysis of the Biosynthetic Cluster. BMC Microbiol. 2016, 16, 240. [Google Scholar] [CrossRef]
- Bhushan, B.; Tomar, S.K.; Mandal, S. Phenotypic and Genotypic Screening of Human-Originated Lactobacilli for Vitamin B12 Production Potential: Process Validation by Micro-Assay and UFLC. Appl. Microbiol. Biotechnol. 2016, 100, 6791–6803. [Google Scholar] [CrossRef]
- Watanabe, F.; Nakano, Y.; Tamura, Y.; Yamanaka, H. Vitamin B12 Metabolism in a Photosynthesizing Green Alga, Chlamydomonas reinhardtii. Biochim. Biophys. Acta 1991, 1075, 36–41. [Google Scholar] [CrossRef]
- ISO 15214:1998; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Mesophilic Lactic Acid Bacteria—Colony Count Technique at 30 °C. ISO (International Organization for Standardization): Geneva, Switzerland, 1998.
- Lavermicocca, P.; Valerio, F.; Lonigro, S.L.; De Angelis, M.; Morelli, L.; Callegari, M.L.; Rizzello, C.G.; Visconti, A. Study of Adhesion and Survival of Lactobacilli and Bifidobacteria on Table Olives with the Aim of Formulating a New Probiotic Food. Appl. Environ. Microbiol. 2005, 71, 4233–4240. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA Sequencing—Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Ferreira Da Silva, M.; Vaz-Moreira, I.; Gonzalez-Pajuelo, M.; Nunes, O.C.; Manaia, C.M. Antimicrobial Resistance Patterns in Enterobacteriaceae Isolated from an Urban Wastewater Treatment Plant. FEMS Microbiol. Ecol. 2007, 60, 166–176. [Google Scholar] [CrossRef]
- Li, P.; Gu, Q.; Yang, L.; Yu, Y.; Wang, Y. Characterization of Extracellular Vitamin B12 Producing Lactobacillus plantarum Strains and Assessment of the Probiotic Potentials. Food Chem. 2017, 234, 494–501. [Google Scholar] [CrossRef]
- Kelleher, B.P.; Broin, S.D. Microbiological Assay for Vitamin B12 Performed in 96-Well Microtitre Plates. J. Clin. Pathol. 1991, 44, 592–595. [Google Scholar] [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Genomes and Mini-Metagenomes from Highly Chimeric Reads BT—Research in Computational Molecular Biology; Deng, M., Jiang, R., Sun, F., Zhang, X., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 158–170. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST Static in Vitro Simulation of Gastrointestinal Food Digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Guidance on the Characterisation of Microorganisms Used as Feed Additives or as Production Organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef] [PubMed]
- Eaton, T.J.; Gasson, M.J. Molecular Screening of Enterococcus Virulence Determinants and Potential for Genetic Exchange between Food and Medical Isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Semedo, T.; Almeida Santos, M.; Martins, P.; Silva Lopes, M.F.; Figueiredo Marques, J.J.; Tenreiro, R.; Barreto Crespo, M.T. Comparative Study Using Type Strains and Clinical and Food Isolates to Examine Hemolytic Activity and Occurrence of the cyl Operon in Enterococci. J. Clin. Microbiol. 2003, 41, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Bover-Cid, S.; Holzapfel, W.H. Improved Screening Procedure for Biogenic Amine Production by Lactic Acid Bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Britt, N.S.; Potter, E.M. Clinical Epidemiology of Vancomycin-Resistant Enterococcus gallinarum and Enterococcus casseliflavus Bloodstream Infections. J. Glob. Antimicrob. Resist. 2016, 5, 57–61. [Google Scholar] [CrossRef]
- Monticelli, J.; Knezevich, A.; Luzzati, R.; Di Bella, S. Clinical Management of Non-Faecium Non-Faecalis Vancomycin-Resistant Enterococci Infection. Focus on Enterococcus gallinarum and Enterococcus casseliflavus/flavescens. J. Infect. Chemother. 2018, 24, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.; Bito, T. Determination of Cobalamin and Related Compounds in Foods. J. AOAC Int. 2018, 101, 1308–1313. [Google Scholar] [CrossRef]
- Madhu, A.N.; Giribhattanavar, P.; Narayan, M.S.; Prapulla, S.G. Probiotic Lactic Acid Bacterium from Kanjika as a Potential Source of Vitamin B12: Evidence from LC-MS, Immunological and Microbiological Techniques. Biotechnol. Lett. 2010, 32, 503–506. [Google Scholar] [CrossRef]
- Stupperich, E.; Nexø, E. Effect of the Cobalt-N Coordination on the Cobamide Recognition by the Human Vitamin B12 Binding Proteins Intrinsic Factor, Transcobalamin and Haptocorrin. Eur. J. Biochem. 1991, 199, 299–303. [Google Scholar] [CrossRef]
- Morita, H.; Toh, H.; Fukuda, S.; Horikawa, H.; Oshima, K.; Suzuki, T.; Murakami, M.; Hisamatsu, S.; Kato, Y.; Takizawa, T.; et al. Comparative Genome Analysis of Lactobacillus reuteri and Lactobacillus fermentum Reveal a Genomic Island for Reuterin and Cobalamin Production. DNA Res. 2008, 15, 151–161. [Google Scholar] [CrossRef]
- Schatz, M.C.; Delcher, A.L.; Salzberg, S.L. Assembly of Large Genomes Using Second-Generation Sequencing. Genome Res. 2010, 20, 1165–1173. [Google Scholar] [CrossRef]
- Sohn, J.-I.; Nam, J.-W. The Present and Future of de novo Whole-Genome Assembly. Brief. Bioinform. 2018, 19, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Bove, P.; Gallone, A.; Russo, P.; Capozzi, V.; Albenzio, M.; Spano, G.; Fiocco, D. Probiotic Features of Lactobacillus plantarum Mutant Strains. Appl. Microbiol. Biotechnol. 2012, 96, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Bove, P.; Russo, P.; Capozzi, V.; Gallone, A.; Spano, G.; Fiocco, D. Lactobacillus plantarum Passage through an Oro-Gastro-Intestinal Tract Simulator: Carrier Matrix Effect and Transcriptional Analysis of Genes Associated to Stress and Probiosis. Microbiol. Res. 2013, 168, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Gheziel, C.; Russo, P.; Arena, M.P.; Spano, G.; Ouzari, H.-I.; Kheroua, O.; Saidi, D.; Fiocco, D.; Kaddouri, H.; Capozzi, V. Evaluating the Probiotic Potential of Lactobacillus plantarum Strains from Algerian Infant Feces: Towards the Design of Probiotic Starter Cultures Tailored for Developing Countries. Probiotics Antimicrob. Proteins 2019, 11, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Damodharan, K.; Lee, Y.S.; Palaniyandi, S.A.; Yang, S.H.; Suh, J.-W. Preliminary Probiotic and Technological Characterization of Pediococcus pentosaceus Strain KID7 and In Vivo Assessment of Its Cholesterol-Lowering Activity. Front. Microbiol. 2015, 6, 768. [Google Scholar] [CrossRef]
- Cao, Z.; Pan, H.; Tong, H.; Gu, D.; Li, S.; Xu, Y.; Ge, C.; Lin, Q. In Vitro Evaluation of Probiotic Potential of Pediococcus pentosaceus L1 Isolated from Paocai—A Chinese Fermented Vegetable. Ann. Microbiol. 2016, 66, 963–971. [Google Scholar] [CrossRef]
- Faye, T.; Tamburello, A.; Vegarud, G.E.; Skeie, S. Survival of Lactic Acid Bacteria from Fermented Milks in an in Vitro Digestion Model Exploiting Sequential Incubation in Human Gastric and Duodenum Juice. J. Dairy Sci. 2012, 95, 558–566. [Google Scholar] [CrossRef]
- Fernandez, B.; Hammami, R.; Savard, P.; Jean, J.; Fliss, I. Pediococcus acidilactici UL5 and Lactococcus lactis ATCC 11454 Are Able to Survive and Express Their Bacteriocin Genes under Simulated Gastrointestinal Conditions. J. Appl. Microbiol. 2014, 116, 677–688. [Google Scholar] [CrossRef]
- Adouard, N.; Magne, L.; Cattenoz, T.; Guillemin, H.; Foligné, B.; Picque, D.; Bonnarme, P. Survival of Cheese-Ripening Microorganisms in a Dynamic Simulator of the Gastrointestinal Tract. Food Microbiol. 2016, 53, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Clarke, S.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-Wide Assessment of Antibiotic Resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2019, 85, e01738-18. [Google Scholar] [CrossRef] [PubMed]
- Singla, V.; Mandal, S.; Sharma, P.; Anand, S.; Tomar, S.K. Antibiotic Susceptibility Profile of Pediococcus spp. from Diverse Sources. 3 Biotech 2018, 8, 489. [Google Scholar] [CrossRef] [PubMed]
- Stefańska, I.; Kwiecień, E.; Jóźwiak-Piasecka, K.; Garbowska, M.; Binek, M.; Rzewuska, M. Antimicrobial Susceptibility of Lactic Acid Bacteria Strains of Potential Use as Feed Additives—The Basic Safety and Usefulness Criterion. Front. Vet. Sci. 2021, 8, 687071. [Google Scholar] [CrossRef] [PubMed]
- Devirgiliis, C.; Zinno, P.; Perozzi, G. Update on Antibiotic Resistance in Foodborne Lactobacillus and Lactococcus Species. Front. Microbiol. 2013, 4, 301. [Google Scholar] [CrossRef] [PubMed]
- Gómez, N.C.; Ramiro, J.M.P.; Quecan, B.X.V.; de Melo Franco, B.D.G. Use of Potential Probiotic Lactic Acid Bacteria (LAB) Biofilms for the Control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coli O157: H7 Biofilms Formation. Front. Microbiol. 2016, 7, 863. [Google Scholar] [CrossRef]
- Zeng, H.; Liu, Y.; Huang, K.; Chen, H.; Yang, B.; Wang, J. Lactiplantibacillus plantarum A1, C1 and C10 Are Potential Probiotics Isolated from Pineapple Residual Silage. Microorganisms 2023, 11, 29. [Google Scholar] [CrossRef]
- Hossain, M.I.; Kim, K.; Rahaman Mizan, M.F.; Toushik, S.H.; Ashrafudoulla, M.; Roy, P.K.; Nahar, S.; Jahid, I.K.; Choi, C.; Park, S.H.; et al. Comprehensive Molecular, Probiotic, and Quorum-Sensing Characterization of Anti-Listerial Lactic Acid Bacteria, and Application as Bioprotective in a Food (Milk) Model. J. Dairy Sci. 2021, 104, 6516–6534. [Google Scholar] [CrossRef]
- Zommiti, M.; Bouffartigues, E.; Maillot, O.; Barreau, M.; Szunerits, S.; Sebei, K.; Feuilloley, M.; Connil, N.; Ferchichi, M. In Vitro Assessment of the Probiotic Properties and Bacteriocinogenic Potential of Pediococcus pentosaceus MZF16 Isolated from Artisanal Tunisian Meat Dried Ossban. Front. Microbiol. 2018, 9, 2607. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic Diversity, Safety and Technological Characterization of Lactic Acid Bacteria Isolated from Artisanal Pico Cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Feng, S.; Wang, H.; Lin, X.; Liang, H.; Zhang, S.; Chen, Y.; Ji, C. Probiotic Properties of Lactobacillus plantarum and Application in Prebiotic Gummies. LWT 2023, 174, 114357. [Google Scholar] [CrossRef]
- Iosca, G.; Fugaban, J.I.I.; Özmerih, S.; Wätjen, A.P.; Kaas, R.S.; Hà, Q.; Shetty, R.; Pulvirenti, A.; De Vero, L.; Bang-Berthelsen, C.H. Exploring the Inhibitory Activity of Selected Lactic Acid Bacteria against Bread Rope Spoilage Agents. Fermentation 2023, 9, 290. [Google Scholar] [CrossRef]
- Pumriw, S.; Luang-In, V.; Samappito, W. Screening of Probiotic Lactic Acid Bacteria Isolated from Fermented Pak-Sian for Use as a Starter Culture. Curr. Microbiol. 2021, 78, 2695–2707. [Google Scholar] [CrossRef] [PubMed]
- Özogul, F.; Hamed, I. The Importance of Lactic Acid Bacteria for the Prevention of Bacterial Growth and Their Biogenic Amines Formation: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1660–1670. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on Risk Based Control of Biogenic Amine Formation in Fermented Foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Bacterial Strains | Number of Isolates |
|---|---|
| Lactococcus lactis | 95 |
| Enterococcus casseliflavus | 16 |
| Lactiplantibacillus plantarum | 14 |
| Pediococcus pentosaceus | 3 |
| Levilactobacillus brevis | 2 |
| Lactic Acid Bacteria | Vitamin B12 Content | Reference |
|---|---|---|
| Lev. brevis | 0.6 pg mL−1 | [21] |
| P. pentosaceus | 0.4–0.6 pg mL−1 | [21] |
| Lpb. plantarum CN-225 | 2.0 pg mL−1 | [21] |
| Lpb. plantarum | 13 ng of g−1 DW | [42] |
| Lpb. plantarum BHM10 | 0.5–0.8 pg mL−1 | [24] |
| Lpb. plantarum BCF20 | 0.5–0.8 pg mL−1 | [24] |
| Lac. lactis E31 | 6.18 ± 1.08 pg mL−1 | This work |
| Lev. brevis G31 | 5.47 ± 0.37 pg mL−1 | |
| P. pentosaceus L51 | 28.19 ± 2.27 pg mL−1 |
| Lac. lactis E32 | Lev. brevis G31 | Lpb. plantarum G12 | P. pentosaceus L51 | |||||
|---|---|---|---|---|---|---|---|---|
| E-Value | Identity | E-Value | Identity | E-Value | Identity | E-Value | Identity | |
| hemL | 4.00 × 10−24 | 27% | 4.00 × 10−24 | 27% | 1.00 × 10−28 | 31% | 0 | 0% |
| cobA | 0 | 0% | 0 | 0% | 0 | 0% | 4.00 × 10−28 | 38% |
| cbiT | 7.00 × 10−5 | 26% | 2.00 × 10−5 | 26% | 2.00 × 10−4 | 26% | 7.00 × 10−4 | 27% |
| cobD | 4.00 × 10−9 | 23% | 3.00 × 10−9 | 23% | 8.00 × 10−17 | 26% | 1.00 × 10−8 | 32% |
| cobC | 9.00 × 10−7 | 30% | 9.00 × 10−7 | 30% | 6.00 × 10−9 | 30% | 3.00 × 10−6 | 24% |
| LAB Strains | Antibiotic Susceptibility | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AMP | CHL | CLI | ERY | GEN | KAN | STR | TET | VAN | ||
| Lac. lactis E32 | MIC | 0.25 | 4 | 0.5 | 0.5 | 32 | 32 | 32 | 0.5 | 0.5 |
| Cut-off | 2 | 8 | 8 | 1 | 32 | 64 | 32 | 4 | 4 | |
| Lev. brevis G31 | MIC | 0.25 | 4 | 0.5 | 0.5 | 16 | 32 | 64 | 0.25 | 0.5 |
| Cut-off | 2 | 4 | 4 | 1 | 16 | 32 | 64 | 8 | n.r. | |
| Lpb. plantarum G12 | MIC | 0.25 | 4 | 8 | 0.5 | 8 | 64 | 32 | 8 | R. |
| Cut-off | 2 | 8 | 8 | 1 | 16 | 64 | n.r. | 32 | n.r. | |
| P. pentosaceus L51 | MIC | 2 | 4 | 0.125 | 0.25 | 16 | 64 | 64 | 8 | R. |
| Cut-off | 4 | 4 | 4 | 1 | 16 | 64 | 64 | 8 | n.r. | |
| LAB Strain | Haemolysis | Biogenic Amine-Producer Phenotype | |||
|---|---|---|---|---|---|
| Tyramine | Histamine | Cadaverine | Putrescine | ||
| Lac. lactis E32 | γ | N | N | N | N |
| Lev. brevis G31 | γ | N | N | N | N |
| Lpb. plantarum G12 | α | N | N | N | N |
| P. pentosaceus L51 | γ | N | N | N | N |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, M.; Maciel, C.; Cruz, P.; Darmancier, H.; Nogueira, T.; Costa, M.; Laranjeira, J.; Morais, R.M.S.C.; Teixeira, P. Exploiting Potential Probiotic Lactic Acid Bacteria Isolated from Chlorella vulgaris Photobioreactors as Promising Vitamin B12 Producers. Foods 2023, 12, 3277. https://doi.org/10.3390/foods12173277
Ribeiro M, Maciel C, Cruz P, Darmancier H, Nogueira T, Costa M, Laranjeira J, Morais RMSC, Teixeira P. Exploiting Potential Probiotic Lactic Acid Bacteria Isolated from Chlorella vulgaris Photobioreactors as Promising Vitamin B12 Producers. Foods. 2023; 12(17):3277. https://doi.org/10.3390/foods12173277
Chicago/Turabian StyleRibeiro, Mónica, Cláudia Maciel, Pedro Cruz, Helena Darmancier, Teresa Nogueira, Margarida Costa, Joana Laranjeira, Rui M. S. C. Morais, and Paula Teixeira. 2023. "Exploiting Potential Probiotic Lactic Acid Bacteria Isolated from Chlorella vulgaris Photobioreactors as Promising Vitamin B12 Producers" Foods 12, no. 17: 3277. https://doi.org/10.3390/foods12173277
APA StyleRibeiro, M., Maciel, C., Cruz, P., Darmancier, H., Nogueira, T., Costa, M., Laranjeira, J., Morais, R. M. S. C., & Teixeira, P. (2023). Exploiting Potential Probiotic Lactic Acid Bacteria Isolated from Chlorella vulgaris Photobioreactors as Promising Vitamin B12 Producers. Foods, 12(17), 3277. https://doi.org/10.3390/foods12173277