
Clipping Effect on the Grain Nitrogen and Protein Fractions of Ancient and Old Wheats Grown in a Mediterranean Environment


Abstract
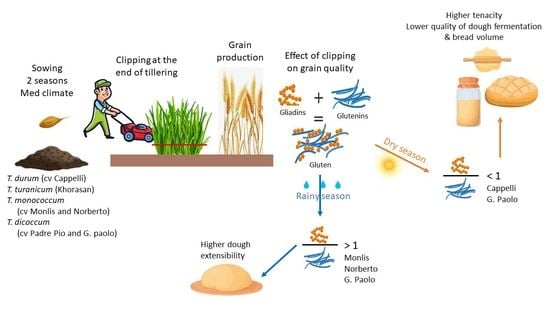
1. Introduction
2. Material and Methods
2.1. Site, Soil, and Agronomic Management
2.2. Factors, Treatment, and Design of the Experiment
2.3. Meteorological Data
2.4. Grain Weight and Nitrogen Content
2.5. Protein Characterization
2.5.1. Sequential Extraction of Gliadin and Glutenin for Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis
2.5.2. Extraction of Extractable and Un-Extractable Polymeric Proteins for Size-Exclusion High-Performance Liquid Chromatography (SE-HPLC) Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Weather Conditions and Phenology
3.2. Grain Nitrogen Content, Protein Composition, and Quality
3.3. Relationship between Genotypic Variation in Grain Nitrogen Content and Protein Fractions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mefleh, M. Cereals of the Mediterranean Region: Their Origin, Breeding History and Grain Quality Traits. In Cereal-Based Foodstuffs: The Backbone of Mediterranean Cuisine; Springer International Publishing: New York, NY, USA, 2021; pp. 1–18. [Google Scholar]
- Boukid, F.; Vittadini, E. Ancient Grains and Pseudocereals: Nutritional Value and Food Applications. In Ancient Grains in Modern Soils; Cooper, R., Ed.; Destech Publications, Inc.: Lancaster, PA, USA, 2022. [Google Scholar]
- Cheng, A. Review: Shaping a Sustainable Food Future by Rediscovering Long-Forgotten Ancient Grains. Plant Sci. 2018, 269, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Mefleh, M.; Motzo, R.; Samson, M.F.; Morel, M.H.; Giunta, F. N Partitioning between Gluten Fractions in a Set of Italian Durum Wheat Cultivars: Role of the Grain N Content. Foods 2020, 9, 1684. [Google Scholar] [CrossRef]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Adhikari, S.; Kumari, J.; Jacob, S.R.; Prasad, P.; Gangwar, O.P.; Lata, C.; Thakur, R.; Singh, A.K.; Bansal, R.; Kumar, S.; et al. Landraces-Potential Treasure for Sustainable Wheat Improvement. Genet. Resour. Crop Evol. 2022, 69, 499–523. [Google Scholar] [CrossRef]
- Van Frank, G.; Rivière, P.; Pin, S.; Baltassat, R.; Berthellot, J.-F.; Caizergues, F.; Dalmasso, C.; Gascuel, J.-S.; Hyacinthe, A.; Mercier, F.; et al. Genetic Diversity and Stability of Performance of Wheat Population Varieties Developed by Participatory Breeding. Sustainability 2020, 12, 384. [Google Scholar] [CrossRef]
- Cadeddu, F.; Motzo, R.; Mureddu, F.; Giunta, F. Ancient Wheat Species Are Suitable to Grain-Only and Grain plus Herbage Utilisations in Marginal Mediterranean Environments. Agron. Sustain. Dev. 2021, 41, 15. [Google Scholar] [CrossRef]
- Giunta, F.; Mefleh, M.; Pruneddu, G.; Motzo, R. Role of Nitrogen Uptake and Grain Number on the Determination of Grain Nitrogen Content in Old Durum Wheat Cultivars. Agronomy 2021, 11, 42. [Google Scholar] [CrossRef]
- Cadeddu, F.; Motzo, R.; Mureddu, F.; Giunta, F. Effects of Clipping on the Nitrogen Economy of Four Triticum Species Grown in a Mediterranean Environment. Field Crops Res. 2021, 267, 108162. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, T.; Peter Angerer, J.; Hou, F. Grazing Seasons and Stocking Rates Affects the Relationship between Herbage Traits of Alpine Meadow and Grazing Behaviors of Tibetan Sheep in the Qinghai–Tibetan Plateau. Animals 2020, 10, 488. [Google Scholar] [CrossRef]
- Shah, L.; Yahya, M.; Shah, S.M.A.; Nadeem, M.; Ali, A.; Ali, A.; Wang, J.; Riaz, M.W.; Rehman, S.; Wu, W.; et al. Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. Int. J. Mol. Sci. 2019, 20, 4211. [Google Scholar] [CrossRef]
- Giunta, F.; Pruneddu, G.; Cadeddu, F.; Motzo, R. Old Tall Durum Wheat Cultivars Are Suited for Dual-Purpose Utilization. Eur. J. Agron. 2017, 90, 67–77. [Google Scholar] [CrossRef]
- Frischke, A.J.; Hunt, J.R.; McMillan, D.K.; Browne, C.J. Forage and Grain Yield of Grazed or Defoliated Spring and Winter Cereals in a Winter-Dominant, Low-Rainfall Environment. Crop Pasture Sci. 2015, 66, 308. [Google Scholar] [CrossRef]
- Sprague, S.J.; Kirkegaard, J.A.; Bell, L.W.; Seymour, M.; Graham, J.; Ryan, M. Dual-Purpose Cereals Offer Increased Productivity across Diverse Regions of Australia’s High Rainfall Zone. Field Crops Res. 2018, 227, 119–131. [Google Scholar] [CrossRef]
- Bell, L.W.; Kirkegaard, J.A.; Tian, L.; Morris, S.; Lawrence, J. Interactions of Spring Cereal Genotypic Attributes and Recovery of Grain Yield After Defoliation. Front. Plant Sci. 2020, 11, 607. [Google Scholar] [CrossRef]
- Ferrise, R.; Bindi, M.; Martre, P. Grain Filling Duration and Glutenin Polymerization under Variable Nitrogen Supply and Environmental Conditions for Durum Wheat. Field Crops Res. 2015, 171, 23–31. [Google Scholar] [CrossRef]
- Vazquez, D.; Berger, A.; Prieto-Linde, M.L.; Johansson, E. Can Nitrogen Fertilization Be Used to Modulate Yield, Protein Content and Bread-Making Quality in Uruguayan Wheat? J. Cereal Sci. 2019, 85, 153–161. [Google Scholar] [CrossRef]
- Mefleh, M.; Boukid, F.; Fadda, C. Suitability of Improved and Ancient Italian Wheat for Bread-Making: A Holistic Approach. Life 2022, 12, 1613. [Google Scholar] [CrossRef]
- Hinson, P.O.; Pinchak, B.; Adams, C.B.; Jones, D.; Rajan, N.; Kimura, E.; Somenahally, A. Forage and Cattle Production during Organic Transition in Dual-purpose Wheat Systems. Agron. J. 2023, 115, 873–886. [Google Scholar] [CrossRef]
- Triboï, E.; Martre, P.; Triboï-Blondel, A.M. Environmentally-Induced Changes in Protein Composition in Developing Grains of Wheat Are Related to Changes in Total Protein Content. J. Exp. Bot. 2003, 54, 1731–1742. [Google Scholar] [CrossRef]
- Sadreddine, B. Yield and Quality of Dual-Purpose Barley and Triticale in a Semi-Arid Environment in Tunisia. Afr. J. Agric. Res. 2016, 11, 2730–2735. [Google Scholar] [CrossRef]
- Virgona, J.M.; Gummer, F.A.J.; Angus, J.F. Effects of Grazing on Wheat Growth, Yield, Development, Water Use, and Nitrogen Use. Aust. J. Agric. Res. 2006, 57, 1307. [Google Scholar] [CrossRef]
- Holman, J.D.; Thompson, C.R.; Hale, R.L.; Schlegel, A.J. Grazing Effects on Yield and Quality of Hard Red and Hard White Winter Wheat. Agron. J. 2009, 101, 775–788. [Google Scholar] [CrossRef]
- Sissons, M.J.; Ames, N.P.; Hare, R.A.; Clarke, J.M. Relationship between Glutenin Subunit Composition and Gluten Strength Measurements in Durum Wheat. J. Sci. Food Agric. 2005, 85, 2445–2452. [Google Scholar] [CrossRef]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Prieto-Linde, M.L.; Johansson, E. Towards the Understanding of Bread-Making Quality in Organically Grown Wheat: Dough Mixing Behaviour, Protein Polymerisation and Structural Properties. J. Cereal Sci. 2012, 56, 659–666. [Google Scholar] [CrossRef]
- Edwards, N.M.; Preston, K.R.; Paulley, F.G.; Gianibelli, M.C.; McCaig, T.N.; Clarke, J.M.; Ames, N.P.; Dexter, J.E. Hearth Bread Baking Quality of Durum Wheat Varying in Protein Composition and Physical Dough Properties. J. Sci. Food Agric. 2007, 87, 2000–2011. [Google Scholar] [CrossRef]
- Aussenac, T.; Rhazi, L.; Branlard, G. Molecular Weight Distribution of Polymeric Proteins in Wheat Grains: The Rheologically Active Polymers. Foods 2020, 9, 1675. [Google Scholar] [CrossRef]
- Arzadún, M.J.; Arroquy, J.I.; Laborde, H.E.; Brevedan, R.E. Effect of Planting Date, Clipping Height, and Cultivar on Forage and Grain Yield of Winter Wheat in Argentinean Pampas. Agron. J. 2006, 98, 1274–1279. [Google Scholar] [CrossRef]
- Boukid, F.; Vittadini, E.; Prandi, B.; Mattarozzi, M.; Marchini, M.; Sforza, S.; Sayar, R.; Seo, Y.W.; Yacoubi, I.; Mejri, M. Insights into a Century of Breeding of Durum Wheat in Tunisia: The Properties of Flours and Starches Isolated from Landraces, Old and Modern Genotypes. LWT 2018, 97, 743–751. [Google Scholar] [CrossRef]
- Zhang, L.-L.; Guan, E.-Q.; Zhang, K.-G.; Zhang, T.-J.; Bian, K. The Aggregation Characteristics of Wheat Globulin Induced by Heating and Edible Salts and Its Effects on Noodle Processing Quality. LWT 2022, 154, 112803. [Google Scholar] [CrossRef]
- Des Marais, D.L.; Hernandez, K.M.; Juenger, T.E. Genotype-by-Environment Interaction and Plasticity: Exploring Genomic Responses of Plants to the Abiotic Environment. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 5–29. [Google Scholar] [CrossRef]
- Zhang, H.; Berger, J.D.; Milroy, S.P. Genotype×environment Interaction Studies Highlight the Role of Phenology in Specific Adaptation of Canola (Brassica Napus) to Contrasting Mediterranean Climates. Field Crops Res. 2013, 144, 77–88. [Google Scholar] [CrossRef]
- Plessis, A.; Ravel, C.; Bordes, J.; Balfourier, F.; Martre, P. Association Study of Wheat Grain Protein Composition Reveals That Gliadin and Glutenin Composition Are Trans-Regulated by Different Chromosome Regions. J. Exp. Bot. 2013, 64, 3627–3644. [Google Scholar] [CrossRef] [PubMed]
- Martre, P.; Porter, J.R.; Jamieson, P.D.; Triboï, E. Modeling Grain Nitrogen Accumulation and Protein Composition to Understand the Sink/Source Regulations of Nitrogen Remobilization for Wheat. Plant. Physiol. 2003, 133, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Triboi, E.; Abad, A.; Michelena, A.; Lloveras, J.; Ollier, J.L.; Daniel, C. Environmental Effects on the Quality of Two Wheat Genotypes: 1. Quantitative and Qualitative Variation of Storage Proteins. Eur. J. Agron. 2000, 13, 47–64. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cultivars | Geographic or Genetic Origin | Year of Release |
|---|---|---|---|
| Triticum durum Desf. | Senatore Cappelli (Cappelli) | North-African landrace Jean Retifah | 1920 |
| Triticumturanicum | Khorasan | Near and Central Asian landrace Jakubz | 2019 |
| Triticum monococcum ssp monococcum | Monlis | Genealogical selection of local population | 2006 |
| Triticum monococcum ssp monococcum | Norberto | Genealogical selection of local population | Known before 2018 as ID331 |
| Triticum dicoccum Schubler Improved Emmer | Giovanni Paolo | Triticum dicoccum Schubler line selected from Molise population × T. turgidum spp. durum Desf. cv Simeto | 2008 |
| Triticum dicoccum Schubler Improved Emmer | Padre Pio | Triticum dicoccum Schubler line selected from Molise population × T. turgidum spp. durum Desf. cv Simeto | 2008 |
| Cultivar | Onset of Grain Filling | Duration of Grain Filling (d) | Rainfall (mm) | Rainfall Deficit (mm) | Minimum Temperature (°C) | Maximum Temperature (°C) | N° of Days with 25 °C < T < 30 °C | N° of Days with T > 30 °C |
|---|---|---|---|---|---|---|---|---|
| S1 | ||||||||
| Giovanni Paolo | 15 March | 57 | 16 | −279 | 10 | 21 | 0 | 13 |
| Cappelli, Khorasan, Padre Pio | 15 April | 41 | 1 | −208 | 11 | 24 | 0 | 13 |
| Monlis, Norberto | 15 May | 26 | 6 | −164 | 14 | 26 | 2 | 20 |
| S2 | ||||||||
| Giovanni Paolo | 1 April | 59 | 240 | −6 | 13 | 22 | 10 | 2 |
| Cappelli, Khorasan, Padre Pio | 20 April | 49 | 238 | 17 | 15 | 24 | 15 | 4 |
| Monlis, Norberto | 20 May | 36 | 73 | −135 | 19 | 29 | 22 | 11 |
| Cultivar | Clipping | Season | Cultivar × Clipping × Season | Cultivar × Clipping | Clipping × Season | |
|---|---|---|---|---|---|---|
| EGNµg | *** | ** | *** | ns | ns | ns |
| 90.05 | 3.27 | 4.88 | 0.66 | 0.61 | 0.53 | |
| UPP% | *** | ns | ** | ns | ns | ns |
| 89.3 | 1.67 | 3.36 | 2.44 | 3.22 | 0 | |
| UP% | *** | ns | *** | ns | ns | ns |
| 84.92 | 0.54 | 9.45 | 2.86 | 1.6 | 0.63 | |
| µg of N in Albumins and Globulins | *** | * | * | ns | *** | ** |
| 56.23 | 0.88 | 3.76 | 6.5 | 12.6 | 20.04 | |
| µg of N in Gliadins | *** | ns | ns | *** | *** | *** |
| 51.35 | 0.18 | 0.4 | 16.16 | 17.15 | 14.75 | |
| µg of N in Glutenins | *** | ** | ** | *** | *** | *** |
| 42.96 | 3.54 | 3.67 | 20.86 | 13.52 | 15.44 | |
| GLI/GLU | *** | * | ns | *** | *** | *** |
| 32.06 | 5.2 | 0.31 | 20.69 | 23.04 | 18.7 | |
| HMW/LMW | *** | ns | ns | *** | *** | *** |
| 74.63 | 0.36 | 0.1 | 6.62 | 15.25 | 3.04 |
| Factor | EGNµg | UP% | UPP% |
|---|---|---|---|
| Clipping | * | ns | ns |
| NC | 893 | 8.12 | 29.3 |
| C | 814 | 8.41 | 29.8 |
| Cultivars | *** | *** | *** |
| Cappelli | 929 b | 6.37 c | 33.9 bc |
| Giovanni Paolo | 1095 a | 10.6 a | 36.6 a |
| Khorasan | 1027 a | 8.26 bc | 31.4 d |
| Monlis | 533 c | 10.7 a | 34.1 b |
| Norberto | 614 c | 9.32 ab | 32.6 cd |
| Padre Pio | 925 b | 4.37 d | 28.3 e |
| Season | *** | *** | ** |
| S1 | 806 | 9.00 | 30.0 |
| S2 | 902 | 7.54 | 29.2 |
| µg of N in Albumins and Globulins | µg of N in Gliadins | µg of N in Glutenins | GLI/GLU | HMW/LMW | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NC | C | NC | C | NC | C | NC | C | NC | C | |
| S1 | ||||||||||
| Cappelli | 241 | 411 | 540 | 149 | 199 | 369 | 2.71 | 0.40 | 0.18 | 0.20 |
| Giovanni Paolo | 182 | 275 | 634 | 329 | 218 | 352 | 2.90 | 1.24 | 0.24 | 0.30 |
| Khorasan | 232 | 261 | 685 | 632 | 178 | 173 | 3.85 | 3.65 | 0.28 | 0.24 |
| Monlis | 76.0 | 79.0 | 248 | 247 | 100 | 97.0 | 2.48 | 2.53 | 0.25 | 0.24 |
| Norberto | 89.0 | 86.0 | 344 | 314 | 114 | 98.0 | 3.04 | 3.21 | 0.25 | 0.26 |
| Padre Pio | 206 | 196 | 600 | 515 | 115 | 87.0 | 5.20 | 5.96 | 0.32 | 0.34 |
| LSD0.05 | 53.8 | 143 | 65.5 | 0.91 | 0.02 | |||||
| S2 | ||||||||||
| Cappelli | 241 | 198 | 577 | 466 | 112 | 155 | 3.75 | 4.18 | 0.20 | 0.19 |
| Giovanni Paolo | 353 | 216 | 246 | 707 | 679 | 186 | 0.37 | 3.82 | 0.30 | 0.35 |
| Khorasan | 276 | 223 | 607 | 544 | 188 | 140 | 3.23 | 3.88 | 0.27 | 0.21 |
| Monlis | 203 | 203 | 223 | 412 | 236 | 67.0 | 1.02 | 6.18 | 0.24 | 0.24 |
| Norberto | 286 | 116 | 152 | 433 | 313 | 138 | 0.48 | 3.15 | 0.25 | 0.26 |
| Padre Pio | 229 | 194 | 617 | 558 | 181 | 213 | 3.41 | 2.60 | 0.37 | 0.25 |
| LSD0.05 | 41.8 | 64.2 | 63.7 | 0.39 | 0.09 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mefleh, M.; Motzo, R.; Boukid, F.; Giunta, F. Clipping Effect on the Grain Nitrogen and Protein Fractions of Ancient and Old Wheats Grown in a Mediterranean Environment. Foods 2023, 12, 2582. https://doi.org/10.3390/foods12132582
Mefleh M, Motzo R, Boukid F, Giunta F. Clipping Effect on the Grain Nitrogen and Protein Fractions of Ancient and Old Wheats Grown in a Mediterranean Environment. Foods. 2023; 12(13):2582. https://doi.org/10.3390/foods12132582
Chicago/Turabian StyleMefleh, Marina, Rosella Motzo, Fatma Boukid, and Francesco Giunta. 2023. "Clipping Effect on the Grain Nitrogen and Protein Fractions of Ancient and Old Wheats Grown in a Mediterranean Environment" Foods 12, no. 13: 2582. https://doi.org/10.3390/foods12132582
APA StyleMefleh, M., Motzo, R., Boukid, F., & Giunta, F. (2023). Clipping Effect on the Grain Nitrogen and Protein Fractions of Ancient and Old Wheats Grown in a Mediterranean Environment. Foods, 12(13), 2582. https://doi.org/10.3390/foods12132582