Abstract
Zoonotic foodborne parasites often represent complex, multi host life cycles with parasite stages in the hosts, but also in the environment. This manuscript aims to provide an overview of important zoonotic foodborne parasites, with a focus on the different food chains in which parasite stages may occur. We have chosen some examples of meat-borne parasites occurring in livestock (Taenia spp., Trichinella spp. and Toxoplasma gondii), as well as Fasciola spp., an example of a zoonotic parasite of livestock, but transmitted to humans via contaminated vegetables or water, covering the ‘farm to fork’ food chain; and meat-borne parasites occurring in wildlife (Trichinella spp., Toxoplasma gondii), covering the ‘forest to fork’ food chain. Moreover, fish-borne parasites (Clonorchis spp., Opisthorchis spp. and Anisakidae) covering the ‘pond/ocean/freshwater to fork’ food chain are reviewed. The increased popularity of consumption of raw and ready-to-eat meat, fish and vegetables may pose a risk for consumers, since most post-harvest processing measures do not always guarantee the complete removal of parasite stages or their effective inactivation. We also highlight the impact of increasing contact between wildlife, livestock and humans on food safety. Risk based approaches, and diagnostics and control/prevention tackled from an integrated, multipathogen and multidisciplinary point of view should be considered as well.
1. Introduction
Foodborne parasites (FBPs) have been long neglected, yet are slowly obtaining more attention, as diagnostic tools are improved and increasingly used, and burden data are becoming slowly available. Efforts made by the Food and Agriculture Organisation and the World Health Organisation (FAO/WHO) into the development of a multicriteria-based ranking for risk management of foodborne parasites have further placed the FBPs in the picture. This ranking was based on a number of criteria including amongst other number of global illnesses, morbidity, mortality; leading to a top four list related to the parasite’s public health impact: Taenia solium, Echinococcus granulosus, Echinococcus multilocularis, Toxoplasma gondii; and a top four of Trichinella spiralis, T. solium, Taenia saginata, Anisakidae when assessing their trade impact [1]. In the WHO’s burden assessment of foodborne pathogens initiative, conducted by the foodborne disease burden epidemiology reference group, the lack of knowledge of the burden of FBPs was acknowledged as well. The report lists the Disability Adjusted Life Years (DALYs) caused by 31 foodborne pathogens, including 14 parasites for which a total of 7,195,014 DALYs, 90,391,678 illnesses and 51,468 deaths were estimated for 2010. The estimates were judged conservative as often data were missing [2]. Foodborne parasites are notorious for their underreporting, most of them not having an obligatory notification.
On the level of the European Union (EU), several COST Actions including CYSTINET (TD1302) and EURO-FBP (FA1408) established or enforced Networks which have contributed greatly to the knowledge and management of FBPs in the EU.
While FBPs were previously majorly linked to endemic areas in the global south, this picture is rapidly changing due to globalisation, including increased international transports, distributions of food stuffs/products, increased trade, changing culinary behaviours moving towards less cooked, raw dishes and increased travel and migration of people. This, combined with an increase in susceptibility of a higher proportion of the global population due to, for example, a higher proportion of elderly people, leads to an increased number of people at higher risk for foodborne parasitic infections [3]. With improvements in diagnostic tools, increase in research efforts, knowledge and awareness on the impact of FBPs has increased, yet a lot of gaps are remaining.
A challenge typically related to FBPs is their long incubation times in humans, whereby clinical symptoms may take even years to appear (e.g., cyst stages of Echinococcus spp., T. solium) which complicates diagnosis, as well as the establishment of the original source of infection.
The FBPs often represent complex, multi host life cycles with parasite stages in the hosts, but also in the environment, where parasite stages may survive for many months or even years. Parasite stages may also contaminate other food stuffs such as fruits and vegetables. A human infection risk therefore may occur (even for a single parasite) at several points in different food chains.
Reduction of the risk can and should be envisaged at different points as well, following a One Health approach, accompanied by an efficient detection (and monitoring) of infection in different hosts. Yet, as will be exemplified below, currently, on a global level, many FBPs are not or insufficiently controlled, diagnostic tools not used or characterised by an insufficient performance. Furthermore, generally, levels of awareness by the different stakeholders are still too low.
In this Special Issue on safety of the food chain, this manuscript aims to provide an overview of a number of important zoonotic foodborne parasites, with a focus on the different food chains in which parasite stages may occur. This paper focusses on the human infection risk, on safety of the food chain, and as such does not cover diagnosis/treatment in human patients. We have chosen some examples of meat-borne parasites occurring in livestock (Taenia spp., Trichinella spp. and Toxoplasma gondii), as well as Fasciola spp., an example of a zoonotic parasite of livestock, but transmitted to humans via contaminated vegetables or water, covering the ‘farm to fork’ food chain as well as meat-borne parasites occurring in wildlife (Trichinella spp., Toxoplasma gondii), covering the ‘forest to fork’ food chain. Moreover, fish-borne parasites (Clonorchis spp., Opisthorchis spp. and Anisakidae) covering the ‘pond/ocean/freshwater to fork’ food chain were added (Figure 1).
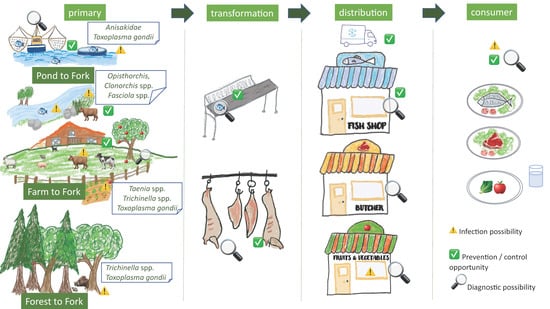
Figure 1.
Simplified presentation of the foodborne zoonotic parasites in the different food chains, and identification of sites for diagnosis and control/prevention.
2. Farm to Fork and Forest to Fork
In this chapter, four important meat-borne zoonotic parasites from livestock will be discussed, including T. solium, T. saginata, Trichinella spp. and Toxoplasma gondii, as well as Fasciola spp. in the framework of the Farm to Fork food chain, while for Trichinella spp. and Toxoplasma gondii the Forest to Fork food chain will be highlighted as well (Figure 1).
2.1. Taenia solium, Taenia saginata
2.1.1. Introduction
Taenia solium and T. saginata are two human tapeworms. Taenia saginata has cattle as the intermediate hosts and primarily represents an economic burden to the meat sector. Taenia solium has pigs as intermediate hosts, but also humans may act as accidental intermediate host, developing neurocysticercosis, a main cause of epilepsy in endemic areas. Taenia solium therefore represents not only an economic problem in the human and veterinary sector, but also a serious public health problem. Taenia saginata has a global distribution, occurring in a high number of countries, with higher prevalences described in countries with raw meat consumption practices such as Belgium [4,5].
After consumption of undercooked infected pork/beef, a tapeworm develops in the human intestine (taeniosis). Eggs are shed from the gravid proglottids, which leave the host actively (only for T. saginata) or with the stool and subsequently contaminate the environment. Taeniosis generally leads to no/limited clinical signs and symptoms, though abdominal complaints, nausea and weight loss have been reported [4].
The metacestode larvae (cysticerci) develop in the (accidental) intermediate hosts after ingestion of the eggs (cysticercosis). In the natural intermediate host, the cysticerci lodge in the muscles, but also subcutaneously and in the brain (porcine cysticercosis, bovine cysticercosis). Generally speaking, the pig/cattle host shows no clinical signs or symptoms, though seizures and changes in behaviour have been described in heavily infected pigs [6,7]. For cattle, heavily infected carcasses detected at slaughter are condemned, while lightly infected carcasses require treatment, but can be sold afterwards. Nevertheless, the freezing of carcasses also entails a value loss. In Belgium, farmers pay an insurance to cover their losses due to bovine cysticercosis, the cattle owners bear an economic cost estimated at €3,408,455/year, while on the human side, an economic cost of €795,858/year was estimated for the taeniosis cases [8]. For porcine cysticercosis, infected carcasses should be condemned at slaughter, leading to economic losses for the farmer or trader. While meat inspection is often not implemented in rural areas in the Global South where most infections occur and therefore carcasses not condemned, infected carcasses do frequently represent an economic loss, as the value of the carcasses may be reduced by 50% [9]. For example, in Tanzania, a loss of nearly three million USD was estimated due to porcine cysticercosis in 2012 [10]. In the human accidental intermediate host, T. solium cysticerci may also lodge in the muscles, subcutaneously, and in the central nervous system. The latter is responsible for most clinical signs and symptoms including seizures, epilepsy, severe progressive chronic headache, vision problems etc. [4]. Taenia solium has been recognised as the FBP with the highest burden, estimated conservatively at 2,788,426 DALYs [2].
2.1.2. Localisation of the Infection Risks for the Consumer in the Food Chain
People develop taeniosis after consumption of undercooked infected pork/beef with viable cysticerci. When the infected pork/beef reaches the consumer in the food chain, the risk of exposure depends on the culinary practices of the consumer (see Section 2.1.4).
For T. solium, causing human cysticercosis, the routes of human infection are more complex, multi causal, and do not require the pig host as the transmission is human-to-human either direct or indirect. Eggs excreted by a human tapeworm carrier are immediately infective. Eggs may be ingested by tapeworm carriers or by close contacts of the carrier via the fecal oral route due to insufficient hand hygiene [4]. Eggs may also end up in the environment and contaminate fruits, vegetables, soil, water and surfaces [11], all potential sources of human infection. In areas with poor sanitation this happens in a more direct way via open defecation, equally so human stool may be used as a fertiliser for vegetable gardens. On the other hand, Taenia spp. eggs have been detected in sewage, in water purification plants, including effluent of these plants, indicating an insufficient clearing of the eggs from the dirty water entering the plants [11,12]. The role of insects in the dispersal of eggs in the environment needs clarification, and their importance for transmission of infective eggs to the human host is unknown [12]. The importance of the different routes of transmission to humans, either direct via contact with a carrier, or indirect via fruits, vegetables, water, etc., needs further research.
Also the survival time of the eggs in different environmental matrices (e.g., soil, water, …) under different climatic conditions, and as such their potential infectivity to humans/animals needs further investigation. Studies have been conducted, though primarily for T. saginata, indicating timespans of survival of up to one year and also the capability of the eggs to survive a European winter [13] (reviewed by [12]).
2.1.3. Diagnostic Options in the Food Chain
In the food chain, different points of diagnosis are possible. For taeniosis, infective cysticerci (viable) should not enter the food chain, therefore, infected carcasses should be picked up and removed from the food chain or should be treated (for T. saginata) [4]. There are two major hurdles in this detection, (1) the implementation of meat inspection and (2) the sensitivity of meat inspection. While meat inspection is implemented on most/all carcasses in industrialised countries, this is not the case in a lot of countries in the Global South, where backyard slaughter, or slaughter in illegal slaughterslabs is routine practice. Even though there seems to be an increasing level of knowledge and awareness regarding the risk of cysticercosis [14], the knowledge is still limited, and the risk perception insufficiently high. Farmers/butchers will still allow the infected meat to reach consumers, often for economic reasons [15]. As an alternative, farmers and traders may conduct a tongue inspection on the live pig, before purchasing. While this system works reasonably well in heavily infected pigs, the sensitivity is very low for other levels of infection, allowing a high number of infected carcasses to enter the food chain [16,17].
Meat inspection has a high specificity, but a notoriously low sensitivity, especially in lightly infected carcasses. For porcine cysticercosis, a sensitivity of 22% has been determined [16], for bovine cysticercosis, this has been estimated at 0.54% in Belgium, where primarily low levels of infection were observed [18].
Serological tools, detecting circulating antigens or specific antibodies have been developed, both for porcine and bovine cysticercosis. Yet, here also, though sensitivities are usually (somewhat) higher than for meat inspection, the tests have specificity problems. For circulating antigen detection, cross reactions with Taenia hydatigena are common, which is especially a problem for porcine cysticercosis, as in a number of endemic areas there is a co-occurrence of these parasites [16].
Most available molecular test are used for confirmation of detected lesions in the meat (reviewed by [19] for porcine cysticercosis).
Diagnostic options in the food chain related to human cysticercosis (T. solium) are currently not routinely implemented. Tests to detect eggs on fruits/vegetables or in environmental matrices such as water, soil have been reviewed recently by [11], with as main conclusions a general lack of sensitivity largely due to low recovery rates of the eggs from the matrices; specificity issues when only microscopic detection is included; and mostly a complete lack of standardisation of tests.
2.1.4. Prevention and Control Options in the Food Chain
Detection and removal of infected carcasses from the food chain (or treatment of lightly infected carcasses) is one of the main preventive targets for taeniosis. As mentioned above, this target is complicated by the lack of implementation of meat inspection in certain highly endemic areas, and even if implemented, especially lightly infected carcasses may still enter the food chain.
For bovine cysticercosis, the impact of meat inspection has been estimated in Belgium, where a prevalence of 42.5% was determined [18]. Meat inspection would only pick up 408 viable cysticerci from an estimated total of 213,344 viable cysticerci, present in the infected carcasses. Important here is that the performance of meat inspection is particularly low in the Belgian setting as most carcasses have very light infections. Nevertheless, in the same study, implementing a serological tool, in this case the circulating antigen detecting ELISA, would in a ten-year period greatly reduce the occurrence of bovine cysticercosis from >40% to 0.6%. The European legislation does allow for serological testing (EC 2019/627 [20]), but in practice, this is not yet implemented, probably due to the costs related to this approach.
In the EU, a more risk-based meat inspection approach is currently implemented considering age of the animals and their production system allowing a selection of animals to undergo a more in-depth meat inspection [21].
As for other FBPs, production systems are of major importance in allowing parasite access the animal hosts. In highly confined and controlled systems (see Section 2.2.4) pigs/cattle would not have access to Taenia eggs due to controlled feed and water access, high hygiene levels avoiding direct contamination via a potential human tapeworm carrier, etc. Nevertheless, while for pigs this is the case in a lot of commercial farming systems, for cattle this is much less so, as outdoor grazing is common; and with the trend towards more biological/organic farming, (renewed) access to potentially contaminated environment/feed/water is created [3].
Vaccination of cattle and pigs has been a subject of research for many years [22]. Especially for the pig host a number of vaccines have been developed, and have been evaluated in the field. The TSOL 18 vaccine is now commercially available, and has shown a very high efficacy, in combination with a single treatment with oxfendazole [23,24]. The latter, treatment with oxfendazole, has also been proposed as an effective option for the control of porcine cysticercosis [25], yet is not implemented outside scientific studies.
At the end of the food chain, the consumer may play a major role as well. Culinary habits will greatly influence the inactivation of viable cysticerci in pork/beef. As for other FBPs, good cooking practices avoiding raw/undercooked pork/beef consumption practices play an essential role in the level of risk of exposure of the consumer, leaving an important role to health education and awareness creation [25,26].
Of course, interventions directed towards the human host such as treatment of tapeworm carriers, stopping open defecation, or management of sludge on pastures are highly relevant [25], but not the focus of this food chain-oriented manuscript.
2.2. Trichinella spp.
2.2.1. Introduction
Larvae from the Trichinella genus present in different meat and meat products have been a source of infection for humans for centuries, with outbreaks still occurring regularly today, with a global distribution.
Transmission of Trichinella spp. among non-human animals occurs by predation or carrion consumption. Transmission to humans occurs via the consumption of raw or undercooked meat [27], whereby infective larvae situated within the muscle cells are released in the stomach to further mature in the small intestine into adult worms. Subsequently, after mating, adult females produce larvae that will migrate to the muscles. Myalgia, diarrhea, fever, facial edema and headaches were mostly reported as clinical signs and symptoms in infected people. Most of these disappeared within 2–8 weeks after treatment, nevertheless myalgia and fatigue may remain present for years. Early diagnosis and treatment in the last decades probably have contributed to a decrease in mortality due to trichinellosis [27]. Trichinella spp. were detected in domestic and wild animals in 66 countries and in humans in 55 countries [27,28]. Trichinellosis represents 550 DALYs, with an estimated 4470 illnesses globally in 2010 [2]. A review conducted to assess the global incidence and clinical impact of trichinellosis identified 65,818 cases from 41 countries between 1986 and 2009. Reporting of cases varies greatly though, and is mainly linked to hospitalised cases, representing a serious underestimated from the true number of cases, including mild or asymptomatic cases. The World health Organisation European Region accounted for the majority of the cases (87%) [27] occurring primarily in adults.
Two life cycles are described for Trichinella spp., a domestic and a sylvatic cycle. The domestic cycle, primarily involving pigs and rodents as hosts and the encapsulated species Trichinella spiralis (T1), relates in this manuscript to the Farm to Fork food chain.
The sylvatic cycle including a large range of wild mammals, birds and reptiles [29], relating to the Forest to Fork food chain, includes primarily the other Trichinella species and genotypes. The encapsulated clade with species infecting mammals only, e.g., Trichinella nativa (T2), Trichinella britovi (T3), Trichinella murelli (T5), Trichinella nelsoni (T7), Trichinella patagoniensis (T12), and Trichinella chanchalensis (T13), with three genotypes Trichinella T6; Trichinella T8 and Trichinella T9, and less common T. spiralis (T1). Species from the non-encapsulated clade infecting mammals and birds include T. pseudospiralis (T4) and infecting mammals and reptiles include T. papuae (T10) and T. zimbabwensis (T11) [28,29].
A recent paper by [30] reviews the presence of Trichinella in wildlife globally, identifying the polar bear (57.58%), martens (32.39%) as the species with the highest prevalence in the palearctic region. While most studies cover terrestrial mammals with as most studied the wild boar, red fox, raccoon dog, wolf, black and polar bears, Trichinella has been detected in marine mammals and birds as well. In addition, rodents and lagomorph species were found with Trichinella. These animal species are important for the maintenance of the transmission cycle, at the same time may be hunted for food consumption, yet are often not part of the food safety control systems. While lions and hyenas have been reported as hosts, and again play a role in the maintenance of the life cycle, infection detected in crocodiles presents a higher risk for unsafe food for humans. The zoonotic potential of T. zimbabwensis is still a matter of debate, an increased use of molecular tools to identify larvae either from a patient (after a muscle biopsy) or after tracing back the meat source will bring more clarity.
2.2.2. Localisation of the Infection Risks for the Consumer in the Food Chain
People are infected via the consumption of infected, undercooked meat. While most of the human infections/outbreaks are associated with pork and pork products from pigs from outdoor breeding farms, cases of infection via consumption of insufficiently cooked other meat and meat products including game meat, especially from wild boars, have been reported regularly [27]. Examples of outbreaks related to horse meat in France and Italy [31,32], dog meat in China [33] have been described. Two large trichinellosis outbreaks in France with direct parasitological evidence indicating horse meat as the source of infection included 128 and 407 cases (reviewed by [31]).
Pork and pork products originating from pigs kept under highly confined and controlled conditions is not a source of infection if the conditions are properly implemented and maintained (see below). Uncontrolled pork/meat/game meat often consumed at the household level, the latter linked to hunting activities, has been regularly reported as source of infection. As described above, a high number of wildlife species may be infected with Trichinella spp., and may therefore be potential sources of infection. Moreover, illegal import of uncontrolled meat from endemic areas has led to outbreaks. International travellers returning home from endemic areas may also develop disease upon their return [27].
Within the limitations of the diagnostic tests, controlled pork/meat/game meat (after slaughter) should prevent infected meat reaching the consumer, or at least at infection levels leading to clinical disease.
2.2.3. Diagnostic Options in the Food chain
Direct testing of muscle samples collected from pig carcasses at the slaughterhouse based on the artificial digestion method is now the most often implemented diagnostic technique, while previously, compression of a small amount of (porcine) muscle between glass slides followed by microscopic examination was routinely applied [34]. Pooled samples of pig muscles are analysed by the magnetic stirrer digestion method (in acidified pepsin). Allowing testing of multiple samples saves a substantial amount of time, especially in low prevalence situations, even though a positive batch result has to be followed by smaller batch testing. The method has a high specificity when conducted by well trained staff. As detected larvae can be recovered, subsequent molecular identification majorly helps epidemiological investigations. Lack of a high sensitivity is often mentioned as a major disadvantage, as infection levels of 3–5 larvae in one gram of muscle would be needed to obtain a sensitivity close to 100% [34]. Nevertheless, application of the test does allow detection of carcasses at this minimum infection level, which are exactly those carcasses that present the highest risk for causing clinical disease in humans [35]. As the food safety objective is to avoid clinical trichinellosis, the performance is satisfactory.
For game, hunters are advised (or obliged depending on the applying regulations) to collect muscle samples for testing.
Indirect detection of infection by specific antibody detection in serum via Enzyme Linked Immunosorbent Assays (ELISAs) is currently not advised for the determination of the infection status of individual carcasses in a food safety control system. Antibody detection suffers from insufficient sensitivity and specificity, though this has been a matter of debate [32,36]. Cross reactions may occur with other parasites infecting pigs, though the level is strongly dependant on the antigen used in the ELISA. Use of properly prepared excretory-secretory (ES) antigens, or ES products, recombinant and synthetic antigens with the same dominant epitopes have been claimed to provide a good specificity in ELISA [36]. Nevertheless, taking into consideration potential false positive results, positive serology would have to be followed by a direct test to confirm infection. The ELISA is usually characterised by a higher sensitivity than the artificial digestion, with a reported detection of one larva per gram of tissue [35,36]. However, this performance is related to the dose of infection, which will influence the level and timing of presence of detectable levels of specific antibodies in the serum, whereby low infection doses may be detectable only as early as six weeks after infection, allowing for a rather large diagnostic window of false negativity. This particularly renders the ELISA much less suitable for individual carcass assessment, as while the test result will be negative, Trichinella larvae may already be present in the muscles. Heavy infections (large infections dose) on the contrary, can be picked up as early as seven days post infection. However, as [35] debates, these low levels of infection may be missed by the direct detection methods as well, as these methods require a presence of 3–5 larvae per gram of tissue to be detectable. Nevertheless, given the challenges related to both the ELISA’s diagnostic specificity and sensitivity, currently the artificial digestion method is still the method of choice recommended by the scientific community and described in regulatory documents.
On the contrary, ELISA can be useful for surveillance or monitoring in pigs and other animals [35] at primary level, in live animals.
Besides the diagnostic performance of the test, cost plays an important role as well. Barlow and colleagues [37] reviewed the available tests for assessment of safe pork and estimated the artificial digestion to be the most optimal choice. The possibility for analyses of pooled samples greatly contributed to a cost reduction, allowing for a system with a satisfactory diagnostic performance and an acceptable cost. ELISA was not selected for individual carcass assessment for reasons explained above. Molecular techniques, such as the conventional and real time PCR, while ideal to identify the Trichinella larva(e), are currently not cost efficient for large scale testing of carcasses. Nevertheless, with technological advances, molecular based methods may become more plausible replacements of the artificial digestion [37]. A promising example could be the lateral flow- recombinase polymerase amplification (LF-RPA) targeting the mitochondrial small subunit ribosomal RNA (rrnS) gene, which can be applied to fresh and frozen pork samples. Besides a very high sensitivity, it has a reported 100% specificity, quick time to result (less than 20 min) and relatively low equipment needs [38]. Nonetheless, a more large-scale evaluation on meat samples is needed [37].
2.2.4. Prevention and Control Options in the Food Chain
In the farm to fork food chain, direct systematic testing using the artificial digestion (see Section 2.2.3) of individual pig carcasses has been a cornerstone of clinical human trichinellosis prevention in many countries for many years. While testing removes a number of infected carcasses from the food chain, it does not detect the lightly infected carcasses and as such does not remove the parasite from the food chain, neither does it fully prevent human exposure to the parasite [35]. Still, the aim to avoid human clinical trichinellosis cases can be achieved implementing this system [39]. Moreover, detection of (heavily) infected carcasses and subsequent correct removal of these carcasses, avoids further potential transmission via animal feed for example. Of course, systematic testing comes with a high cost. In the European Union, this was estimated at an annual cost ranging from 25–400 million euro [40].
With the changes in farm management system, moving towards highly confined and highly controlled housing and farming systems (high levels of biosecurity), the risk of exposure of the pig host is removed and the occurrence of Trichinella spp. in the pig host subsequently dropped drastically. Indeed, under these conditions, the parasite may be removed from this particular farm to fork food chain. The development of international guidelines describing criteria for controlled housing and management, thereby prevented the risk of exposure of animals. Implementation of these systems actually reduces/removes the need for testing [35], as the parasite is absent from these particular commercial pork production systems. Indeed, summary data from the EU (European Centre for Disease Prevention and Control, [41]) for 2019 described that routine slaughter testing of 72.8 million pigs raised under controlled housing reported no Trichinella spp. infections. Considering the cost of testing, especially in low prevalence settings, with most animals testing negative, alternatives were searched to replace the systematic testing.
A set of criteria described by the World Organisation for Animal Health (OIE) to recognise free regions or countries was fairly rapidly replaced by a set of requirements to meet for a status of negligible risk as advised by the International Commission on trichinellosis [35,42]. The latter has as a purpose to ensure food safety (consumer health) and to obtain standardised requirements for international trade, removing the need for testing of animals originating from herds classified as negligible risk. The standards included amongst other good feed manufacturing and storage practices, rodent control, prevention of pigs accessing wildlife, removal of deceased pigs and controlled animal movement [35].
Of course, not all pigs are bred under these controlled management systems, and as such, are exposed to a higher risk of Trichinella spp. infection as exemplified by the EU summary data whereby 218 positive pigs were detected from 139.6 million pigs from non-controlled housing tested (European Centre for Disease Prevention and Control, [41]). Non-controlled housing may allow pigs access to potentially infected rodents and wildlife. The same counts for other animal species bred under non-controlled systems, such as horses. Carcasses from the latter also need to be tested systematically, like pigs. The trend towards biological/organic pig farming [3], moving away from these highly controlled/confined systems opens opportunities for the parasite to enter the farms. Whether there is a risk and how high this risk is, depends on the infection levels in the local wildlife, including rodents. In this interface area setting, both food chains, farm to fork and forest to fork merge. The expansion of wild boars into areas of free-range pig production in the Unites States represents a risk for pig infection, leading to the introduction of Trichinella spp. into the farm to fork food chain [43], at the same time increasing the availability of potentially infected wild boar meat via hunters.
For Trichinella spp., the forest to fork food chain is an important source of infection to humans, especially game meat consumers, hunters. Human behaviour is particularly important in this food chain, as the testing (or not) of game meat, and consumption practices are dependent on the hunter/consumer. Often home consumption of hunted game is not subject to regulations, therefore testing is frequently not performed. Risk of infection is subsequently dependant on the presence and level of infection as well as on the culinary practice of the household which may include cooking, freezing or curing, influencing the inactivation of the parasite. Health education on good preparation methods for meat that might contain Trichinella larvae, as described by the International Commission on Trichinellosis is encouraged, including cooking, freezing (for meat from domestic pigs), and irradiation [34].
Efforts have been ongoing in the development of vaccines targeting to reduce the larval or adult worm burdens, but to date, no sufficiently effective vaccine is available [44].
As highlighted in Section 2.2.3, ELISA detecting specific antibodies may be used for surveillance and monitoring in pigs and other animal species [36].
2.3. Toxoplasma gondii
2.3.1. Introduction
Toxoplasma gondii is an obligate intracellular protozoon that causes toxoplasmosis in man and animals. Toxoplasmosis is one of the most common parasitic zoonoses worldwide with an estimated one third of the global population being infected [45]. Toxoplasmosis is present in every country and seropositivity rates range from less than 10% to over 90% [45]. FAO and WHO have ranked T. gondii fourth out of 24 foodborne parasites of global importance [1]. The global annual incidence of congenital toxoplasmosis was estimated to be 190,100 cases. This was equivalent to a burden of 1.20 million DALYs (95% CI: 0.76–1.90). High burdens were seen in South America and in some Middle Eastern and low-income countries [46].
T. gondii uses felines as the definitive hosts, but is less specific when it comes to the intermediate hosts, which can be any warm-blooded animal, including mammals and birds. The lifecycle comprises an intestinal sexual phase that takes place only in the final hosts and results in the production of unsporulated oocysts that are faecally shed by infected felines and sporulate in the environment; and an extra-intestinal phase in intermediate hosts which comprises the formation of tachyzoites and tissue cysts that contain bradyzoites. In most hosts, T. gondii causes a lifelong latent infection in tissues such as skeletal and heart muscles, visceral organs and the central nervous system [47].
Most infections with T. gondii in humans and animals are subclinical or cause mild clinical signs and symptoms. Severe disease may occur in hosts with an immature or compromised immune system, or in case of higher pathogenicity of the parasite strain. In livestock, T. gondii is an important cause of abortion in sheep and goats. In humans, infection with T. gondii is particularly important in pregnant women and in immunocompromised people. When primary infection occurs during pregnancy T. gondii can cross the placenta and reach the foetus causing a mild to life threatening infection depending on the gestational stage. Disease- or medication-induced immunosuppression can lead to encephalitis or disseminated toxoplasmosis in adults. While toxoplasmosis in immunocompetent individuals is asymptomatic in around 80% of cases, in about 20% infection it may cause fever, mononucleosis-like symptoms, or ocular manifestations such as chorioretinitis [47].
2.3.2. Localisation of the Infection Risks for the Consumer in the Food Chain
Infection in humans may occur from the accidental ingestion of oocysts that can be present in food such as vegetables and fruits but also shellfish, or in water due to environmental contamination with cat faeces; or from the consumption of raw or undercooked meat containing tissue cysts. As mentioned above vertical transmission may occur when a parasite-naïve women gets infected during pregnancy. Less frequent ways of infection are caused by drinking goat milk containing tachyzoites during the acute phase of infection or by transplantation of tissues from a T. gondii-infected donor. Consumption of undercooked infected meat is considered a major risk factor for humans, especially in Europe, where it has been associated with 30–63% of infections [48,49]; Hill et al. [50] demonstrated the predominance of oocyst-caused infections in North America. Based on the relative proportion of the different animals in the overall meat consumption, the proportion of meat types consumed raw or undercooked, the exposure and susceptibility of different animal species to T. gondii and the subsequent establishment and survival of parasites in their tissues, meat from pigs and small ruminants are to be seen as the main sources of infection (farm to fork food chain). Meat from wildlife (forest to fork food chain), horses, poultry and cattle are less common sources [49].
Toxoplasma gondii is an example of a health issue that can be directly connected to outdoor animal husbandry [51]. Grazing ruminants mostly acquire infection by ingestion of oocysts from the environment. As a result, the seroprevalence in sheep tends to be high, e.g., between 27.8% and 87.4% in West-European countries [52]. Pigs may become infected both by ingesting oocysts and by the consumption of meat containing tissue cysts from infected rodents or kitchen leftovers. Consequently, a low prevalence (0–1%) is found in pig farms with well-managed controlled housing conditions that practise rodent control, keep cats away from the farm and the feed, and restrict the access to the farm. In contrast, a high prevalence of up to 60% is found in poorly managed or free-range farms [53,54,55]. Outdoor access of pigs considerably increases the risk for T. gondii infection such as in free-range organic farms [54,56]. Thomas et al. [57] studied the detailed anatomical distribution of T. gondii in naturally and experimentally infected lambs and found that parasite DNA could be detected in all the edible parts.
Environmental contamination with T. gondii oocysts is understudied and likely underestimated, which is partly due to the lack of suitable harmonized sampling approaches and detection methods. Oocysts can be spread in the soil by arthropods, earthworms, wind, and rain [58]. Sporulated oocysts are highly resistant and can remain infective in soil for up to two years [59]. Several waterborne infections associated with T. gondii oocysts have been described. Water in irrigation systems, rivers, lakes, beaches, and coasts, as well as wastewater and groundwater can be contaminated with the environmentally resistant oocysts. Moreover, oocysts can survive various inactivation procedures using chemical reagents, including sodium hypochlorite and chlorine [60]. Oocyst contamination of fresh vegetables may occur through cultivation in contaminated soil or using contaminated water for irrigation or washing. Consumption of raw vegetables and fruits are a risk factor. T. gondii oocysts can also enter the marine environment through disposal of sewage and water runoff, where they can cause infections in marine animals [61]. Oocysts have been detected in wild and commercial bivalve mollusks which are filter-feeders and can concentrate microorganisms. They can retain viable T. gondii oocysts for 85 days following uptake [62]. These shellfishes can pose another risk for consumers when consumed undercooked or raw [63].
2.3.3. Diagnostic Options in the Food Chain
Toxoplasma gondii infections can be detected by direct and indirect techniques. Coprological methods are used to detect oocysts in cat faeces. (Immuno-)histological and molecular methods and bioassay are used to detect the parasite in tissues, in most cases post-mortally. Recently, an improved molecular method using magnetic capture combined with RT-PCR has been developed that allows the detection of the parasite in larger portions of tissue [64]. Cat and mouse bioassays are the reference direct techniques to isolate T. gondii; however, these tests are not commonly used due to the long time it takes to obtain results, ethical issues, and high costs [47]. An alternative method is cell culture which is limited in use because of the variability of the results [65]. Many serological methods (indirect detection) have been developed and validated in humans and several animal species. Among these techniques are the immunofluorescent assay (IFAT), enzyme-linked immunosorbent assay (ELISA), latex agglutination tests (LAT) and the modified agglutination test (MAT). An important question is whether seropositivity can be linked to the infectivity of tissues. Opsteegh et al. [66] found that the correlation between antibody detection against T. gondii and direct parasite detection is high in pigs, small ruminants, and chickens. In these species, the use of serology can help determine the risk to the consumer, but it may not be as useful in other species, such as horses and cattle. In addition, a seronegative result does not necessarily mean that the meat is free of T. gondii [67].
2.3.4. Prevention and Control Options in the Food Chain
Although T. gondii is a high priority foodborne zoonotic parasite, it is not systematically controlled. At present, there are no specific regulations and no standardised methods for the detection of T. gondii in any food matrix. Because chronically infected animals are mostly asymptomatic and tissue cysts cannot be detected during routine meat inspection—the size of tissue cysts is less than 100 µm—most infected carcasses pass meat inspection and enter the food chain.
Currently, most of the control of T. gondii infection is carried out at home, especially in people that are most vulnerable to the parasite, such as pregnant women and immune-compromised persons. Primary prevention consisting of dietary recommendations, pet care measures, environmental measures, knowledge of risk factors and ways to control toxoplasmosis infection, has been found to be effective in reducing congenital toxoplasmosis [68].
At farm level preventive measures mostly apply to the pig industry where the parasite can be virtually eliminated by a set of hygienic measures, as discussed above. The serological prevalence of T. gondii in the pig population may be a useful indicator of the risk of human toxoplasmosis associated with the consumption of pork products. In the EU, the Commission Regulation No. 219/2014 modernised some specific requirements of the post-mortem inspection of pigs, favouring visual inspection instead of palpation and incisions [69]. However, it does not solve the Toxoplasma problem at slaughterhouse level. Therefore, the categorisation of risk for different types of farms (intensive systems and organic farms) would help the official veterinarian during ante-mortem and post-mortem visits [70].
Post-slaughter methods involve meat processing for human consumption and include mainly heat inactivation and freezing that are effective ways to kill the parasites if properly applied. Rani and Pradhan [71] studied the survival of T. gondii during cooking and low temperature storage and concluded that viable parasites were not found when the internal temperature of meat reached 64 °C and below −18 °C. Other meat curing procedures such as salting, smoking, or fermentation are less reliable for killing the parasites in meat because they are very much dependent of the concentration of the salt, the storage temperature, and the time of the processing [55]. The modern trend steered toward meat production from organic breeding, consumed raw or undercooked, with low concentrations of salt and additives (e.g., nitrites), may result in an increase of the zoonotic risk [54,56]. The current scientific knowledge is outdated and not sufficient for a full risk assessment. For this reason, innovative studies on T. gondii inactivation focusing on modern processing technologies may contribute to outline new preventive measures for consumers [55].
Currently, testing for parasite contamination in fresh produce is neither regulated nor mandatory. The increased popularity of consumption of raw and ready-to-eat vegetables may pose a new potential risk for consumers who could be accidentally exposed to oocysts, since most post-harvest processing measures do not guarantee the complete removal of oocysts or their effective inactivation [58].
Similar to other pathogens research is ongoing on the development of vaccines against toxoplasmosis. Currently, in animals, the only available vaccine is a live, attenuated, T. gondii S48 strain licensed for use in sheep in Europe and New Zealand for prevention of abortion [72]. However, using a live vaccine raises safety concerns for use in food-producing animals since the vaccine strain may revert to a wild type that might cause tissue cyst formation. DNA vaccination has been shown to determine long-lived humoral and cellular immune responses in vivo in animals [73]. Vaccination of cats has been proposed as the ultimate preventive measure because of the pivotal role of cats in the lifecycle of T. gondii. However, in case an effective vaccine for cats would be available, prospects on preventing oocyst-originated human toxoplasmosis by vaccination in large populations of cats are not favourable due to the large vaccination coverage needed [74].
2.4. Fasciola spp.
2.4.1. Introduction
The trematodes Fasciola hepatica and Fasciola gigantica are the causative agents of the disease fasciolosis. The life cycle of Fasciola spp. includes plant-eating mammals (mainly ruminants, but also pigs and others) as final hosts, aquatic, lymnaeid snails as intermediate hosts and aquatic plants as carriers. Humans can also act as final hosts, and may even contribute to the perpetuation of the life cycle in endemic areas, where poor sanitation occurs [75], although the extent to which this actually occurs has never been quantified.
For decades, fasciolosis was perceived to be a purely veterinary problem [76], however, it is now also seen as an important disease in humans. As a response, WHO has listed fasciolosis as one of the neglected tropical diseases to be prioritized for control [77]. Human fasciolosis is known to occur worldwide, although it mainly affects the poorest communities in rural areas across subtropical and tropical countries. No recent burden data are available for fasciolosis. In 2012, Fürst et al. [78] estimated the global burden of fasciolosis at 35,206 DALYs, whereas the 2015 Global Burden of Disease estimate amounted to 90,041 DALYs, however the latter estimate came with a wide uncertainty interval (58,050–209,097) [79]. Estimates for the number of people infected range between 2.4 and 17 million [78,80,81], whereas those for the population at risk range between 91 and 180 million people [82,83]. Based on reported infection intensities, it has been estimated that 14% of fasciolosis infections are symptomatic [78]. These numbers could underestimate the true occurrence and impact of the disease, as for many regions, such as for instance a number of countries on the African continent, few or no community-based epidemiological surveys have been performed [84], and considering that the current burden estimation does not yet account for account for the immunosuppression, neurological or ocular effects due to fasciolosis.
Overall, fasciolosis is considered an emerging disease [82,85,86]. Its emergence could be due to the increased attention it received since its designation as neglected tropical disease, combined with factors facilitating its expansion and importance. For instance, climate change is thought to cause an increased risk for its snail host survival and distribution, and thus spread of the disease [87,88]. Moreover, the increased consumption of raw vegetables and fruits as part of increasingly popular healthy life styles and the introduction of sylvatic reservoir animals can assist in the spread of the disease [89,90].
Infected individuals harbour the adult Fasciola spp. worms, and unlike in Clonorchis/Opisthorchis spp. infection, will shed unembryonated eggs with their faeces/stool [91]. Once in a favourable freshwater environment, the eggs will then embryonate after approximately two weeks. After hatching, the miracidia will actively search for a suitable freshwater snail to penetrate and infect. In the snail, the miracidia will undergo several developmental stages (i.e., sporocyst and redia stages) and various rounds of asexual multiplication [90]. Next, an exponential number of free-swimming cercariae will leave the snail and encyst into metacercariae (MC) on plants growing in the same aquatic environment. Susceptible final hosts can acquire the infection by ingesting contaminated raw water plants. Upon ingestion, the MC will excyst in the small intestine, and penetrate the intestinal wall. Next, the juvenile flukes will migrate through the liver where they will mature in the bile ducts and start producing eggs, completing the life cycle [92]. Ectopic infections can also occur, with the juvenile or immature flukes erroneously migrating to subcutaneous tissues, gastrointestinal tract, heart, lung, or rarely, brain and eye [93,94].
In humans, fasciolosis can cause fever, abdominal colic, digestive disorders, weight loss, anaemia and jaundice due to fluke migration and subsequent destruction of the liver tissue, inflammation and blockage of the bile ducts [94]. In ectopic infections, symptoms and signs are specific to the organ affected by the migration path and presence of the flukes. Recently, it has been shown that neurological, meningeal and ocular symptoms, such limb and facial paralysis, speech disorders, blindness can also occur in hepatic fasciolosis, due to leakages in blood-brain barrier [95]. Finally, for a long time, Fasciola spp. infection was thought to cause parasitic pharyngitis, called Halzoun. The proposed route for acquiring this condition was the ingestion of raw liver, contaminated with immature or young flukes, attaching to the pharyngeal mucosa and invoking pain and bleeding. However, it is now argued the condition is rather due to infection with other parasites, such as Linguatula serrata, and Dicrocoelium dendriticum [90].
2.4.2. Localisation of the Infection Risks for the Consumer in the Food Chain
Based on current knowledge, the most common pathway of infection for humans is the consumption of contaminated water plants. The best known infection source is watercress (Nasturtium spp.), an ubiquitous green leafy vegetable, with case reports around the globe mentioning the consumption of this plant in the anamnesis [96,97,98]. Furthermore, a wide range of other freshwater plants have been reported as infection sources, both wild and cultivated ones, such as dandelion (Taraxacum spp.) leaves, lamb’s lettuce (Valerianella locusta), and water spinach (Ipomoea aquatica). Contamination of freshwater plants is mostly due to the direct shedding of eggs by an infected host, being domestic livestock, humans or even sylvatic hosts, such as the nutria (Myocastor coypus), into an aquatic environment where the suitable intermediate snail host is present as well as the water plant eventually serving as carrier for the infective stage, metacercariae [90].
Next to water plants, a variety of terrestrial plants can carry the infective stage of Fasciola spp. For instance, infections due the consumption of lettuce (Lactuca sativa), and parsley (Petroselinum sativum) have been reported [99,100], while in other cases, infection was presumed to be due to consumption of wild aromatic plants, such as mint (Mentha spp.) [101]. Contamination of these plants can be due to them being submerged in water during a certain period of time, either in an environment where eggs had been shed, and thus snails were infected, or due to flooding of an adjacent field, with the runoff transporting infected snails or MC over a certain distance [102]. Other contamination pathways in terrestrial plants are washing in contaminated water bodies, and through irrigation with contaminated water.
Next to the ingestion of plants, the chewing and sucking contaminated plants are common infection routes for Fasciola spp. Well known examples are grass chewing, especially in children [103], as well as chewing on leaves of khat (Catha edulis), a popular tradition in the Horn of Africa and the Arabian Peninsula. Moreover, in certain regions, drinking beverages, such as herbal teas, and juices made from local plants are a known risk factor [104]. Drinking contaminated water is another suggested route of infection, although its importance is not well understood. Indeed, even though it is proven that unattached MC sink quite quickly to the bottom of water bodies, drinking contaminated water has been identified as the sole infection routes in a number of areas [105,106,107]. Naturally, ingestion of dishes and soups made with contaminated water, and washing of vegetables, fruits, tubercles, kitchen utensils or other objects with contaminated water can equally cause infection. Finally, in pig experiments, it was shown that the ingestion of raw liver with juvenile Fasciola spp. can lead to established liver infections [108], however to what extent this occurs in humans is not known.
2.4.3. Diagnostic Options in the Food Chain
Surprisingly little attention has been given to the development of diagnostic tools for MC detection on plants and in water. Moreover, the sensitivity and specificity of available techniques have never been assessed. Up to now, most techniques for plants entail the screening of plant surfaces by means of a stereomicroscope [109]. Not only is this is not a feasible option in a commercial context, due to the time consuming nature of the method, it also requires the necessary expertise, to recognize Fasciola spp. MC, especially on thicker leave and stem surfaces, and to differentiate them from other digenean MC (e.g., Paramphistomum spp.) [109]. In water, while originally developed to investigate the contamination of rice fields, floats or buoys could be used to catch MC present in important waterflows from irrigation channels and/or streams [110,111]. Overall, cost-effective techniques to detect metacercarial contamination of consumed plants and water are lacking.
2.4.4. Prevention and Control Options in the Food Chain
The most obvious method for fasciolosis prevention is avoiding oral contact with raw plants, whether it is the sucking/chewing or consumption of raw plants, especially those growing in an aquatic environment. However, culinary habits are often difficult to break, and therefore such recommendation might not be realistic for many communities and individuals. Another option would be the removal and/or destruction of MC attached on plant leaves. Unfortunately, washing vegetables using running water alone has proven to be only moderately successful in detaching MC [112]. On the contrary, briefly (5–10 min) soaking plants in vinegar (e.g., 120 mL/L) or liquid soap (e.g., 12 mL/L) solutions, seems effective in detaching and killing the MC [112,113], however these studies should be repeated to confirm the application of these methods in a commercial context. Some studies have investigated the effects of other chemical agents, of which potassium permanganate, and sodium hydroxide treatments seemed successful detachment/destruction methods [101,112,113], however, such agents have a considerable impact on plant palatability and appearance, and therefore their application seems unfeasible in a commercial context [113]. Cooking vegetables seems a more effective method to kill MC, however culinary traditions in raw plant consumption and in some regions, inadequate means to ensure sufficiently high temperatures while cooking, might hamper the application of this preventive measures. Similarly, boiling potentially contaminated water might not be feasible due to lack of means, in such regions filtration with an appropriate mesh size might be more effective [90].
Currently there are few preventive measures for fasciolosis implemented by governments. In the European Union, all bovines and small ruminants processed through abattoirs will be inspected for Fasciola spp., by means of visual inspection and incisions of the liver, in line with the European meat inspection legislation (EC 2019/627, [20]), however this inspection will not stop infected livestock from contaminating fields and therefore indirectly causing human fasciolosis cases. In a wider preventive context, plants commonly consumed by humans, should be grown under controlled conditions, inaccessible to snails, ruminants and other animals. Appropriate legislation has been implemented by a number of countries, e.g., France and Australia [90,114]. Moreover, the risk of contamination of these fields, due to run-off or irrigation with water from adjacent areas where Fasciola spp. can be present, should be excluded. A One Health approach is needed to ensure that all stakeholders are involved and informed about the disease, the risks and responsibilities. In any given region, appropriate fasciolosis prevention and control may only be achieved once the local transmission dynamics of Fasciola spp. are fully understood.
3. Pond/Ocean/River to Fork: Fish-Borne Parasites
In this chapter, three important fish-borne zoonotic parasites will be discussed, including Clonorchis spp. and Opisthorchis spp., related to freshwater fish, as well as the group of Anisakidae, related to marine fish (Figure 1).
3.1. Fish Borne Trematodes: Clonorchis, Opisthorchis spp.
3.1.1. Introduction
Liver flukes of the Opisthorchiidae family, of which Opisthorchis viverrini, O. felineus and Clonorchis sinensis are the most important, cause opisthorchiasis and clonorchiasis in humans. The parasites have a complex lifecycle, requiring Bithynia spp. freshwater snails and cyprinid fish as primary and secondary intermediate hosts, respectively, and a fish-consuming mammal, such as humans, cats and dogs, as the final hosts [115]. Although cat and dogs are known to contribute to the life cycle of for instance O. viverrini, it has equally been shown for the same parasite, cats and dogs cannot sustain transmission in absence of humans as a final host [116]. It is not known, if the same is true for O. felineus and C. sinensis. The infected final host will discharge embryonated eggs in the biliary ducts and shed them via the stool/faeces. If shed in a suitable environment, the eggs will be ingested by the snail intermediate host. In the snail, the eggs will release miracidia that will subsequently develop into sporocysts, rediae and cercariae [117]. The latter developmental stage will be released from the snail host into the aquatic environment, where it will actively seek for a suitable secondary intermediate host, a freshwater cyprinid fish, penetrate its flesh or skin and encyst as metacercaria (MC) [118]. The final host acquires the infection by ingesting the raw or undercooked contaminated fish. Upon ingestion, the MC excysts in the small intestine and travels to the biliary tract via the ampulla of Vater. In the biliary ducts, the flukes will mature and start producing eggs, thereby restarting the life cycle [119].
Opistorchiosis and clonorchiosis can cause cholangitis, jaundice, cholecystitis, hepatomegaly and cholelithiasis [117]. Moreover, the International Agency for Research on Cancer has classified both O. viverrini and C. sinensis as Type I carcinogens. Indeed, in chronic infections, the (i) repeated mechanical damage due to the feeding and migrating flukes, combined with (ii) the secretion and excretion of metabolic products by the flukes as well as by Helicobacter spp. often co-infecting the final hosts, and (iii) the immunopathological response by the host, causing fibrosis and blockage of the bile ducts, may over time lead to oxidative damage to the epithelial cell DNA. Normal repair mechanisms and apoptosis are inhibited by certain fluke excretory/secretory products, and oncogenic mutations can occur as a consequence [117,120,121]. Eventually, the malignant transformations will lead to the development of cholangiocarcinoma (CCA), a highly lethal cancer of the bile duct, with an estimated median survival time of 4.3 months after diagnosis [121,122]. Up to now, while O. felineus has not been listed officially as a Type I carcinogen, there are indications both from animal experiments and reviews of the occurrence of cholangiocarcinoma in regions where the parasite is prevalent, that this fluke too has carcinogenic potential [123,124]. Opistorchiosis and clonorchiosis are two neglected yet emerging zoonotic diseases [82,125]. In the 1990s the total number of clonorchiosis and opistorchiosis cases was estimated at 7 and 10 million, respectively [83]. Nowadays, the total global number of infected people is estimated at about 20 million for C. sinensis, and at 10 million for O. viverinni, with most infections occurring in East Asia. Between 1.2 and 1.6 million people; mainly in Eastern Europe, are estimated to be infected with O. felineus [126]. Another 601 million people are thought to be at risk for C. sinensis infection, while 80 million for Opisthorchis spp. infection. The estimates for global burden due to clonorchiosis ranges between 275,370 and 522,863 Disability Adjusted Life Years (DALYS), whereas the estimates for opistorchiosis burden vary between 74,367 and 188,346 [78,79]. Based on reported infection intensities, it has been estimated that 8·2% of clonorchiosis, and 4·9% of opistorchiosis cases are symptomatic [78]. Increased importation of potentially contaminated fish, and newly acquired raw-fish eating habits might cause a future expansion in the distribution of the diseases [26].
3.1.2. Localisation of the Infection Risks for the Consumer in the Food Chain
Humans principally acquire opistorchiosis and clonorchiosis through the consumption of undercooked or raw infected freshwater cyprinid fish [91]. Moreover, other preparation styles, such as fermentation, pickling, inadequately freezing, or smoking of fish can pose a risk to the consumer [127]. In addition to cyprinid fish, some other fish types such eleotrids, cichlids, and osmerids are known to harbour MC [128]. Finally, some freshwater shrimp species have been reported to carry MC, and their consumption could thus pose the consumer at risk for infections [128].
While it is clear that the MC only encyst in fish/shrimp tissue, and thus the consumption of these tissues is the main infection source for acquiring the diseases, it is not well understood whether the preparation of contaminated fish can lead to contamination of the cooking environment, and thus subsequent human infection via contact with this environment. Indeed, Opisthorchis and Clonorchis spp. MC are distributed over different parts of the fish body, being the muscles, fins, heads and organs [129]. Therefore, it is not unimaginable that cutting boards might become contaminated with MC during the preparation of the fish dish. For instance, in their study performed in Laos, Araki et al. [130] reported on the observation that household cooks clean their chopping boards by scratching them using a knife and running water, and at times fish scales could still be found on the chopping boards after cleaning. Likewise, utensils and hands might become contaminated during food preparation or while eating [130]. Such contamination could explain infections in people reporting to never consume raw fish [131]. Finally, it has been hypothesized that people could become infected by drinking water contaminated by MC released from dead fish tissues, although this has never been proven to occur under field conditions [128].
3.1.3. Diagnostic Options in the Food Chain
All techniques to detect Opisthorchis spp. and C. sinensis MC in fish are based on postmortem investigations. Most consist of rather labour-intensive, traditional parasitological techniques. The most basic method is the direct compression method: fins, muscle, scales and subcutaneous tissues are collected, compressed between glass slides and examined for MC under a stereomicroscope [83]. In a second commonly used method, the artificial digestion method, the fish is divided in five parts (head, anterior and posterior trunk, tail and subcutaneous tissue). The tissues are subsequently ground, then digested using an artificial gastric juice (mostly a pepsin-HCl solution), and after sedimentation, MC are sought using a stereomicroscope [83]. A systematic review has shown that studies applying the compression method find higher prevalence estimates than those using the digestion method, however, their performance has not been compared directly [132]. Either way, both techniques require expert knowledge on the morphology of the different MC present in fish to allow for their correct differentiation (e.g., of Opisthorchiidae, Heterophyidae and Lecithodendriidae) [133].
Molecular techniques are however available to ensure correct differentiation of MC. For instance, a multiplex PCR was developed, targeting mitochondrial DNA, to allow differentiation of Clonorchis and Opisthorchis MC, particularly useful for regions where their distribution overlaps [134]. Moreover, a loop-mediated isothermal amplification (LAMP) was also developed for C. sinensis detection in fish, with a markedly higher sensitivity as compared to PCR [135]. Nevertheless, such molecular techniques remain expensive and not available in a commercial aquaculture context. Up to now, quick, easy, cheap diagnostic tools with adequate test performance are still lacking for Clonorchis and Opisthorchis detection in fish.
3.1.4. Prevention and Control Options in the Food Chain
Prevention of clonorchiosis and opisthorchis entails preventing the consumption of raw or undercooked contaminated fish/shrimps and avoiding contact with a contaminated environment. In many communities, the consumption of raw or undercooked fish is however deeply engrained in the culinary and social culture. For instance, the consumption of raw fish is often part of a social drinking events for males in many Asian countries [136,137,138]. Simply halting its consumption or changing the preparation style might therefore not be feasible. Moreover, MC are moderately tolerant to several preparation and preservation styles, which would usually be considered as effective in killing off germs. For instance, freezing at −12 °C had to be continued for 480 h to inactivate Clonorchis MC, whereas cooking at 50 °C required five hours to inactivate unspecified MC [127]. The only effective method is thoroughly heating the fish at a high temperature (e.g., 65 °C for 1 min) [127]. Another route for prevention in the intermediate host, namely a vaccine for freshwater fish, is currently being explored. Indeed, an oral vaccine based on Bacillus subtilis expressing enolase, was developed and is being tested [139,140]. In a wider preventive perspective, the life cycle can be broken by disconnecting human and animal faeces from the aquatic environment. At the moment, in some countries, the use of toilet types draining stool directly to ponds, still persist [141], and animal and human faeces continue being used as fish feed [139], therefore the transmission perpetuates. In Thailand, an opistorchiosis control program, called the “Lawa model” was developed. This EcoHealth/One Health-inspired approach combining human treatment with novel community-based and school health education, ecosystem monitoring and community participation, was implemented in the opistorchiosis endemic area at Lawa Lake, Khon Kaen province, Thailand [142]. The program successfully cut back infection rates in both humans, fish and snails, and will now be scaled up to other regions in Thailand and beyond [143].
3.2. Anisakidae
3.2.1. Introduction
Nematodes from the family Anisakidae are by far the most prevalent macroparasites in fish implicated in human disease. In their adult stage, anisakids are mostly found in the stomach of marine mammals as their definitive hosts, whereas the larval stages are found in smaller invertebrates, such as crustaceans as their first intermediate host, and in fish as their second intermediate host. Many commonly exploited marine fish species are infected, with the main zoonotic anisakid species being, although not limited to these, Anisakis simplex sensu stricto, A. pegreffii, Pseudoterranova decipiens, and Contracaecum osculatum [144]. Data on their prevalence in fish are known for many fish species with varying prevalences (e.g., up to 100% in herring) dependent on the geographical fishing grounds and seasons [145,146]. Yet ultimately, almost all species of teleost fish throughout the oceans may act as hosts where larvae can be found in the gastrointestinal tract or musculature of the fish [147,148].
Human infection, collectively named anisakidosis, takes place after consumption of undercooked fish containing a viable third-stage larva (L3), and may lead to several gastro-intestinal symptoms depending on the localisation of the larvae (acute, chronic, and ectopic) [149]. In addition to abdominal symptoms, a series of thermostable allergens present in A. simplex and A. pegreffii may also compromise human health with acute allergic manifestations ranging from urticaria, angioedema, asthma, conjunctivitis, to even a potential lethal anaphylactic shock [150]. The number of human health problems related to Anisakidae has not been fully quantified as global awareness of this foodborne parasite is still in its upsurge. Collection of more epidemiological data on the disease is therefore encouraged to provide better insights into the impact of Anisakidae on human disease. Nevertheless, specific country case studies from Spain and Norway have estimated 10,383–20,978 annual anisakidosis cases, as well as a 22% prevalence value of Anisakis-sensitization in certain regions [151,152]. Furthermore, it was stated that the most frequent cause of an anaphylactic episode due to a hidden allergen, is fish infected with A. simplex [153]. Finally, sufficient proof-of-principle has been provided in the past that demonstrates the transmissibility of anisakid allergenic peptides from fishmeal, as a feed component, to aquacultured fish and chicken meat [154,155,156,157]. These findings may significantly change the importance of these zoonotic nematodes from originally a purely fish borne food risk, to a much wider risk from several food sources (pork meat, chicken meat, aquacultured fish, etc.).
3.2.2. Localisation of the Infection Risks for the Consumer in the Food Chain
As abovementioned, consumers obtain an anisakid infection via consumption of infected fish that has not been sufficiently frozen or cooked [149]. Different preventive measurements can be taken to avoid this (see below), though these are primarily related to the infection risk with viable larvae. By adequate cooking or freezing, Anisakis-sensitized consumers remain at risk given the thermoresistant characteristics of the allergens [158]. Moreover, removal of the larva does not at all guarantee freedom of allergens since some of these anisakid allergens are excretory-secretory products excreted/secreted by the larval body during its migration through the fish flesh. As such, patients may be exposed to them when consuming fish, even though the larvae might have been removed during the quality control of the fish [159]. Finally, and different to other food allergies, occupational allergies in aquaculture and fishery workers after inhalation and/or skin contact with anisakid allergens have been reported [160].
3.2.3. Diagnostic Options in the Food Chain
Fish and fishery products can be examined for the presence of anisakid larvae and traces by a variety of detection methods. In the industrial setting (i.e., on the boat, in processing plants), candling is the most routinely used method for the detection of anisakids in commercial fish fillets. It entails a brief visual inspection of fish fillets on a light table to spot and manually remove parasites [161]. While this technique has the major advantage of not affecting the fish quality and thus allowing consumption afterwards, it is labour intensive and as such highly costly. Moreover, studies report a poor sensitivity with up to 76% of the larvae not being recovered, although this is dependent on the fish (colour, thickness, skin), the larvae (size, colour), and the skills of the inspector [161,162,163,164]. In laboratory settings on the other hand, a range of highly accurate alternatives such as UV press method, enzymatic digestion, and immunoassays are available and implemented, though the complete destruction of the fish tissue, renders these methods unfit for the application in the industry [165,166,167]. As a result, candling is still the standard method for the detection and removal of anisakids from fish fillets on an industrial scale. Future research should thus look into the development of more accurate, fast, non-destructive scanning methods to replace candling.
As dead larvae may still be responsible for allergic reactions in sensitized consumers, other tools directly targeting anisakid proteins have been developed [168]. However, the presence of anisakid proteins does not necessarily correlate with an allergic reaction since not all proteins are allergens. To deal with this, liquid chromatography tandem mass spectrometry methods and allergen specific enzyme-linked immunosorbent assays are available [169,170], but high equipment costs and their destructive entity still hamper their use in an industrial setting.
3.2.4. Prevention and Control Options in the Food Chain
Prevention of gastro-intestinal anisakidosis is simply based on avoiding the ingestion of a live L3 in raw/undercooked fish. In the food chain, this can be accomplished by maintaining a cold chain from boat to plate and by immediate gutting of the fish on the boat in order to avoid post-mortem migration from the fish gut to musculature. Care must be taken to destroy these viscera rather than disposing them in the sea water as this practice results in, once again, dissemination of the parasites [171]. Freezing of the fish has also been recommended by public health agencies and included in the current legislation of the European Union and Japan [172,173]. Specifically, food industry business that sell fish intended for raw, marinated, or salted consumption, must first freeze the fish in its entirety at −20 °C for >24 h, or −35 °C for >15 h to ensure killing of the larvae [174]. It is to be expected though, that in a big container of fish, not all parts of the fish will reach a temperature to kill all larvae. Ideally, freezing should therefore be followed by a period of storage in the frozen state to ensure complete elimination of the parasite. In addition to freezing/cooking, a visual check for larvae by the fish industry can be conducted by ways of candling (see above). The abovementioned limitations of this tool, however, make this tool insufficient for full clearance of the parasite and emphasizes the importance of adequate deep-freezing. Finally, at the stage of not only the consumer, but also the relevant governmental institutes and medical/veterinary staff, raising awareness regarding the presence of these parasites and possible preventive measures (e.g., cooking, consumption behaviour) is a principle preventive measure that should be taken.
While a variety of measures can be taken to reduce the incidence of human gastro-intestinal anisakidosis, it is important to consider that an Anisakis-sensitized individual may still develop an allergic reaction if the larva is dead/removed or if its traces are present [158]. So far, no allergen destruction process has been discovered, and standard testing for anisakid allergens is up to date not conducted on any food type. A suggestion, however, could be to label high-risk products for the possible presence of Anisakis allergens to warn sensitized patients who do not tolerate even properly cooked or canned fish.
4. Discussion and Conclusions
In this paper we have attempted to highlight the presence of a number of FBPs in different food chains, the risk of infection of humans by parasites, and their diagnostic and control challenges. We have also highlighted the complexity of their life cycles whereby different parasite stages, infective to the human host, may occur at different places within a food chain, for example, in the case of T. solium whereby the meat-borne component will place humans at risk when consuming pig carcasses infected with viable cysticerci, but humans may also be at risk when consuming vegetables or fruits contaminated with the parasite’s eggs from the farm, or via contaminated water. Toxoplasma gondii is also a good example of multiple complex ways of transmission to humans.
As mentioned, the increased popularity of consumption of raw and ready-to-eat vegetables may pose a new potential risk for consumers, since most post-harvest processing measures do not guarantee the complete removal of oocysts/eggs or their effective inactivation [58]. The same can be said for raw fish/meat consumption, considering the generally low sensitivities or simply non implementation of detection tools.
We have also highlighted how interactions in interface areas may lead to the introduction of parasites from the forest to fork food chain into the farm to fork food chain. The impact of increasing contact between wildlife, livestock and humans on food safety needs to be considered carefully, especially bearing in mind the trend towards outdoor farming [3] and the increase in relevant wildlife species such as the wild boar [27].
The multidisciplinary One Health approach will be needed to deal with the impact of globalisation and climate change on the transmission of foodborne parasites to humans [26]. The One Health approach is increasingly applied by governments, yet mostly considering human and animal components. The environment is the most understudied component of parasite transmission, yet is very important. There is an urgent need for researchers to take up this work, e.g., to develop highly performing diagnostic tools to detect environmental stages, to assess their viability, etc.
The availability of cheap, easy to use, highly performing diagnostic tools fit for purpose is another challenge. While an increased amount of effort has been put into developments of new tests in different formats, and target product profiles are being developed (e.g., for T. solium [175]), their large-scale evaluation in relevant matrices is often lacking. Integrated systems looking into more integrated sample collections where relevant (e.g., environmental samples, meat samples from pig farms with outdoor access), and test systems allowing multipathogen detection are other aspects that need to be considered.
Monitoring and surveillance, and improved reporting based on proper diagnostics would be useful to implement for a number of FBPs. As resources are always limited, risk-based surveillance might help in the prioritisation, and lead to an efficient and effective allocation of, resources [21]. For Trichinella spp. and T. saginata this system has been put in place in the EU (see Section 2.1.4 and Section 2.2.4).
Integration may be considered not only on a detection level, but also on a prevention/control level, where chosen interventions may impact on several pathogens. The implementation of highly controlled and confined farming systems has led to commercial pork originating from these farms to be free of Trichinella spp. These high levels of biosecurity not only impact the presence of Trichinella spp., but also Taenia spp. and T. gondii are rarely detected. Moreover, a focus on herd health and management, biosecurity at the farm level would tackle not only a number of FBPs, but also other pathogens such as Salmonella and Campylobacter, and this at a primary, pre-harvest level [176]. Nevertheless, the trend towards farming systems with more outdoor access, encouraged by animal welfare expectations, increase infection risk [3]. The latter should be compensated with either pre-harvest (e.g., improved detection) or post-harvest (e.g., inactivation via cooking) measures.
To finalise, the consumer also plays an essential role in risk of exposure, via human behaviour in choice of food to consume but also in the culinary practices in how to process food for consumption. While cooking at a sufficient temperature would deal with all parasites described above, often, this is not done. Other practices such as marinating, salting, drying, smoking are more often than not, insufficient to inactivate the parasite. From a different perspective, human migration and import of infected animals from endemic to non-endemic areas may lead to a (re)introduction of pathogens [3]. Consumer education regarding the risks and awareness creation would be highly beneficial.
Author Contributions
Conceptualization, S.G., P.D., V.D. and G.S.; writing S.G., P.D., G.S. and V.D. Original Draft Preparation, Writing—Review & Editing, S.G.; Visualization, S.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO/WHO (Food and Agriculture Organization of the United Nations/World Health Organization). Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites; Microbiological Risk Assessment Series No. 23, Rome; World Health Organization: Geneva, Switzerland, 2014; p. 302. [Google Scholar]
- WHO. World Health Organisation, WHO Estimates of the Global Burden of Foodborne Diseases Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Gabriël, S.; Johansen, M.V.; Pozio, E.; Smit, G.S.; Devleesschauwer, B.; Allepuz, A.; Papadopoulos, E.; van der Giessen, J.; Dorny, P. Human migration and pig/pork import in the European Union: What are the implications for Taenia solium infections? Vet. Parasitol. 2015, 213, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Murrell, K.D.; Dorny, P.; Flisser, A.; Geerts, S.; Kyvsgaard, N.C.; McManus, D.; Nash, T.E.; Pawlowski, Z.S. WHO/FAO/OIE Guidelines for the Surveillance, Prevention & Control of Taeniosis/Cysticercosis; Murrell, K.D., Ed.; OIE: Paris, France, 2005. [Google Scholar]
- Jansen, F.; Dorny, P.; Berkvens, D.; Van Hul, A.; Van den Broeck, N.; Makay, C.; Praet, N.; Eichenberger, R.M.; Deplazes, P.; Gabriël, S. High prevalence of bovine cysticercosis found during evaluation of different post-mortem detection techniques in Belgian slaughterhouses. Vet. Parasitol. 2017, 244, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, C.; Mkupasi, E.M.; Ngowi, H.A.; Forkman, B.; Johansen, M.V. Severe seizures in pigs naturally infected with Taenia solium in Tanzania. Vet. Parasitol. 2016, 220, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, C.; Johansen, M.V.; Mkupasi, E.M.; Ngowi, H.A.; Forkman, B. Disease behaviours of sows naturally infected with Taenia solium in Tanzania. Vet. Parasitol. 2017, 235, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Dorny, P.; Trevisan, C.; Dermauw, V.; Laranjo-González, M.; Allepuz, A.; Dupuy, C.; Krit, M.; Gabriël, S.; Devleesschauwer, B. Economic impact of bovine cysticercosis and taeniosis caused by Taenia saginata in Belgium. Parasit. Vectors 2018, 11, 241. [Google Scholar] [CrossRef]
- Hobbs, E.C.; Mwape, K.E.; Devleesschauwer, B.; Gabriël, S.; Chembensofu, M.; Mambwe, M.; Phiri, I.K.; Masuku, M.; Zulu, G.; Colston, A.; et al. Taenia solium from a community perspective: Preliminary costing data in the Katete and Sinda districts in Eastern Zambia. Vet. Parasitol. 2018, 251, 63–67. [Google Scholar] [CrossRef]
- Trevisan, C.; Devleesschauwer, B.; Schmidt, V.; Winkler, A.S.; Harrison, W.; Johansen, M.V. The societal cost of Taenia solium cysticercosis in Tanzania. Acta Trop. 2017, 165, 141–154. [Google Scholar] [CrossRef]
- Saelens, G.; Robertson, L.; Gabriël, S. Diagnostic tools for the detection of taeniid eggs in different environmental matrices: A systematic review. Food Waterborne Parasitol. 2022, 26, e00145. [Google Scholar] [CrossRef]
- Jansen, F.; Dorny, P.; Gabriël, S.; Dermauw, V.; Johansen, M.V.; Trevisan, C. The survival and dispersal of Taenia eggs in the environment: What are the implications for transmission? A systematic review. Parasit. Vectors 2021, 14, 88. [Google Scholar] [CrossRef]
- Bucur, I.; Gabriël, S.; Van Damme, I.; Dorny, P.; Vang Johansen, M. Survival of Taenia saginata eggs under different environmental conditions. Vet. Parasitol. 2019, 266, 88–95. [Google Scholar] [CrossRef]
- Gabriël, S.; Mwape, K.E.; Phiri, I.K.; Devleesschauwer, B.; Dorny, P. Taenia solium control in Zambia: The potholed road to success. Parasite Epidemiol. Control 2018, 4, e00082. [Google Scholar] [CrossRef]
- Thys, S.; Mwape, K.E.; Lefèvre, P.; Dorny, P.; Phiri, A.M.; Marcotty, T.; Phiri, I.K.; Gabriël, S. Why pigs are free-roaming: Communities’ perceptions, knowledge and practices regarding pig management and taeniosis/cysticercosis in a Taenia solium endemic rural area in Eastern Zambia. Vet. Parasitol. 2016, 225, 33–42. [Google Scholar] [CrossRef]
- Dorny, P.; Phiri, I.K.; Vercruysse, J.; Gabriel, S.; Willingham, A.L., 3rd; Brandt, J.; Victor, B.; Speybroeck, N.; Berkvens, D. A Bayesian approach for estimating values for prevalence and diagnostic test characteristics of porcine cysticercosis. Int. J. Parasitol. 2004, 34, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Chembensofu, M.; Mwape, K.E.; Van Damme, I.; Hobbs, E.; Phiri, I.K.; Masuku, M.; Zulu, G.; Colston, A.; Willingham, A.L.; Devleesschauwer, B.; et al. Re-visiting the detection of porcine cysticercosis based on full carcass dissections of naturally Taenia solium infected pigs. Parasit. Vectors 2017, 10, 572. [Google Scholar] [CrossRef]
- Jansen, F.; Dorny, P.; Berkvens, D.; Gabriël, S. Bovine cysticercosis and taeniosis: The effect of an alternative post-mortem detection method on prevalence and economic impact. Prev. Vet. Med. 2018, 161, 1–8. [Google Scholar] [CrossRef]
- Waema, M.W.; Misinzo, G.; Kagira, J.M.; Agola, E.L.; Ngowi, H.A. DNA-Detection Based Diagnostics for Taenia solium Cysticercosis in Porcine. J. Parasitol. Res. 2020, 2020, 5706981. [Google Scholar] [CrossRef]
- EU. Commission implementing regulation (EU) 2019/627 of 15 March 2019 laying down uniform practical arrangements for the performance of official controls on products of animal origin intended for human consumption in accordance with Regulation (EU) 2017/625 of the European Parliament and of the Council and amending Commission Regulation (EC) No 2074/2005 as regards official controls. Off. J. Eur. Union 2019, L 131, 72. [Google Scholar]
- Alban, L.; Häsler, B.; van Schaik, G.; Ruegg, S. Risk-based surveillance for meat-borne parasites. Exp. Parasitol. 2020, 208, 107808. [Google Scholar] [CrossRef]
- Lightowlers, M.W. Vaccines for prevention of cysticercosis. Acta Trop. 2003, 87, 129–135. [Google Scholar] [CrossRef]
- Lightowlers, M.W. Eradication of Taenia solium cysticercosis: A role for vaccination of pigs. Int. J. Parasitol. 2010, 40, 1183–1192. [Google Scholar] [CrossRef]
- Gabriël, S.; Mwape, K.E.; Hobbs, E.C.; Devleesschauwer, B.; Van Damme, I.; Zulu, G.; Mwelwa, C.; Mubanga, C.; Masuku, M.; Mambwe, M.; et al. Evidence for potential elimination of active Taenia solium transmission in Africa? N. Engl. J. Med. 2020, 383, 396–397. [Google Scholar] [CrossRef] [PubMed]
- De Coster, T.; Van Damme, I.; Baauw, J.; Gabriël, S. Recent advancements in the control of Taenia solium: A systematic review. Food Waterborne Parasitol. 2018, 13, e00030. [Google Scholar] [CrossRef] [PubMed]
- Pozio, E. How globalization and climate change could affect foodborne parasites. Exp. Parasitol. 2020, 208, 107807. [Google Scholar] [CrossRef] [PubMed]
- Murrell, K.D.; Pozio, E. Worldwide Occurrence and Impact of Human Trichinellosis, 1986–2009. Emerg. Infect. Dis. 2011, 17, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Pozio, E. World distribution of Trichinella spp. infections in animals and humans. Vet. Parasitol. 2007, 149, 3–21. [Google Scholar] [CrossRef]
- Pozio, E.; Hoberg, E.; La Rosa, G.; Zarlenga, D.S. Molecular taxonomy, phylogeny and biogeography of nematodes belonging to the Trichinella genus. Infect Genet. Evol. 2009, 9, 606–616. [Google Scholar] [CrossRef]
- Crisóstomo-Jorquera, V.; Landaeta-Aqueveque, C. The genus Trichinella and its presence in wildlife worldwide: A review. Transbound. Emerg. Dis. 2022, 69, e1269–e1279. [Google Scholar] [CrossRef]
- Boireau, P.; Vallée, I.; Roman, T.; Perret, C.; Mingyuan, L.; Gamble, H.R.; Gajadhar, A. Trichinella in horses: A low frequency infection with high human risk. Vet. Parasitol. 2000, 93, 309–320. [Google Scholar] [CrossRef]
- Gottstein, B.; Pozio, E.; Nöckler, K. Epidemiology, diagnosis, treatment, and control of trichinellosis. Clin. Microbiol. Rev. 2009, 22, 127–145. [Google Scholar] [CrossRef]
- Liu, M.; Boireau, P. Trichinellosis in China: Epidemiology and control. Trends Parasitol. 2002, 18, 553–556. [Google Scholar] [CrossRef]
- Noeckler, K.; Pozio, E.; van der Giessen, J.; Hill, D.E.; Gamble, H.R. International Commission on Trichinellosis: Recommendations on post-harvest control of Trichinella in food animals. Food Waterborne Parasitol. 2019, 21, e00041. [Google Scholar] [CrossRef]
- Gamble, H.R. Trichinella spp. control in modern pork production systems. Food Waterborne Parasitol. 2022, 28, e00172. [Google Scholar] [CrossRef]
- Bruschi, F.; Gómez-Morales, M.A.; Hill, D.E. International Commission on Trichinellosis: Recommendations on the use of serological tests for the detection of Trichinella infection in animals and humans. Food Waterborne Parasitol. 2019, 5, e00032. [Google Scholar] [CrossRef]
- Barlow, A.; Roy, K.; Hawkins, K.; Ankarah, A.A.; Rosenthal, B. A review of testing and assurance methods for Trichinella surveillance programs. Food Waterborne Parasitol 2021, 9, e00129. [Google Scholar] [CrossRef]
- Li, X.; Liu, W.; Wang, J.; Zou, D.; Wang, X.; Yang, Z.; Yin, Z.; Cui, Q.; Shang, W.; Li, H.; et al. Rapid detection of Trichinella spiralis larvae in muscles by loop-mediated isothermal amplification. Int. J. Parasitol 2012, 42, 1119–1126. [Google Scholar] [CrossRef]
- Van Knapen, F. Control of trichinellosis by inspection and farm management practices. Vet. Parasitol. 2000, 93, 385–392. [Google Scholar] [CrossRef]
- Kapel, C.M. Changes in the EU legislation on Trichinella inspection—New challenges in the epidemiology. Vet. Parasitol. 2005, 132, 189–194. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Trichinellosis. In ECDC. Annual Epidemiological Report for 2019; ECDC: Stockholm, Sweden, 2021; Available online: https://www.ecdc.europa.eu/en/publications-data/trichinellosis-annual-epidemiological-report-2019 (accessed on 23 November 2022).
- Dupouy-Camet, J.; Murrell, K.D. FAO/WHO/OIE Guidelines for the Surveillance, Management. Prevention and Control of Trichinellosis; OIE: Paris, France, 2007; pp. 101–110. [Google Scholar]
- Burke, R.; Masuoka, P.; Murrell, K.D. Swine Trichinella infection and geographic information system tools. Emerg. Infect Dis. 2008, 14, 1109–1111. [Google Scholar] [CrossRef]
- Tang, B.; Li, J.; Li, T.; Xie, Y.; Guan, W.; Zhao, Y.; Yang, S.; Liu, M.; Xu, D. Vaccines as a Strategy to Control Trichinellosis. Front. Microbiol. 2022, 23, 857786. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.; Weiss, L. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Mastroiacovo, P. The global burden of congenital toxoplasmosis: A systematic review. Bull. WHO 2013, 91, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasmosis of Animals and Humans; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Cook, A.J.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of toxoplasma infection in pregnant women: European multi- centre case-control study. European Research Network on Congenital Toxoplasmosis. British Med. J. 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Tenter, A.M. Toxoplasma gondii in animals used for human consumption. Mem. Inst. Oswaldo Cruz. 2009, 104, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.; Coss, C.; Dubey, J.P.; Wroblewski, K.; Sautter, M.; Hosten, T.; Munoz-Zanzi, C.; Mui, E.; Withers, S.; Boyer, K.; et al. Identification of a sporozoite-specific antigen from Toxoplasma gondii. J. Parasitol. 2011, 97, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Kijlstra, A.; Meerburg, B.G.; Bos, A.P. Food safety in free-range and organic livestock systems: Risk management and responsibility. J. Food Prot. 2009, 72, 2629–2637. [Google Scholar] [CrossRef]
- Verhelst, D.; De Craeye, S.; Vanrobaeys, M.; Czaplicki, G.; Dorny, P.; Cox, E. Seroprevalence of Toxoplasma gondii in domestic sheep in Belgium. Vet. Parasitol. 2014, 205, 57–61. [Google Scholar] [CrossRef]
- Dubey, J.P.; Jones, J.L. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol. 2008, 38, 1257–1278. [Google Scholar] [CrossRef]
- Bacci, C.; Vismarra, A.; Mangia, C.; Bonardi, S.; Bruini, I.; Genchi, M.; Kramer, L.; Brindani, F. Detection of Toxoplasma gondii in free-range, organic pigs in Italy using serological and molecular methods. Int. J. Food Microbiol. 2015, 202, 54–56. [Google Scholar] [CrossRef]
- De Berardinis, A.; Paludi, D.; Pennisi, L.; Vergara, A. Toxoplasma gondii, a foodborne pathogen in the swine production chain from a European perspective. Foodborne Pathogens Dis. 2017, 14, 637–648. [Google Scholar] [CrossRef]
- Kijlstra, A.; Eissen, O.A.; Cornelissen, J.; Munniksma, K.; Eijck, I.; Kortbeek, T. Toxoplasma gondii infection in animal-friendly pig production systems. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3165–3169. [Google Scholar] [CrossRef]
- Thomas, M.; Aubert, D.; Escotte-Binet, S.; Durand, B.; Robert, C.; Geers, R.; Alliot, A.; Belbis, G.; Villena, I.; Blaga, R. Anatomical distribution of Toxoplasma gondii in naturally and experimentally infected lambs. Parasite 2022, 29, 3. [Google Scholar] [CrossRef]
- Shapiro, K.; Kim, M.; Rajal, V.B.; Arrowood, M.J.; Packham, A.; Aguilar, B.; Wuertz, S. Simultaneous detection of four protozoan parasites on leafy greens using a novel multiplex PCR assay. Food Microbiol. 2019, 84, 103252. [Google Scholar] [CrossRef]
- Dumètre, A.; Dardé, M.L. How to detect Toxoplasma gondii oocysts in environmental samples? FEMS Microbiol. Rev. 2003, 27, 651–661. [Google Scholar] [CrossRef]
- Mirza Alizadeh, A.; Jazaeri, S.; Shemshadi, B.; Hashempour-Baltork, F.; Sarlak, Z.; Pilevar, Z.; Hosseini, H.A. Review on inactivation methods of Toxoplasma gondii in foods. Pathog. Glob. Health 2018, 112, 306–319. [Google Scholar] [CrossRef]
- Shapiro, K.; Vanwormer, E.; Aguilar, B.; Conrad, P.A. Surveillance for Toxoplasma gondii in California mussels (Mytilus californianus) reveals transmission of atypical genotypes from land to sea. Environ. Microbiol. 2015, 17, 4177–4188. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Collins, M.V.; Mitchell, S.M.; Wetch, C.N.; Rosypal, A.C.; Flick, G.J.; Zajac, A.M.; Lindquist, A.; Dubey, J.P. Survival of Toxoplasma gondii oocysts in eastern oysters (Crassostrea virginica). J. Parasitol. 2004, 90, 1054–1057. [Google Scholar] [CrossRef]
- Hohweyer, J.; Dumètre, A.; Aubert, D.; Azas, N.; Villena, I. Tools and methods for detecting and characterizing Giardia, Cryptosporidium, and Toxoplasma parasites in marine mollusks. J. Food Prot. 2013, 76, 1649–1657. [Google Scholar] [CrossRef]
- Gisbert Algaba, I.; Geerts, M.; Jennes, M.; Coucke, W.; Opsteegh, M.; Cox, E.; Dorny, P.; Dierick, K.; De Craeye, S. A more sensitive, efficient and ISO 17025 validated Magnetic Capture real time PCR method for the detection of archetypal Toxoplasma gondii strains in meat. Int. J. Parasitol. 2017, 47, 875–884. [Google Scholar] [CrossRef]
- Opsteegh, M.; Dam-Deisz, C.; de Boer, P.; De Craeye, S.; Faré, A.; Hengeveld, P.; Luiten, R.; Schares, G.; van Solt-Smits, C.; Verhaegen, B.; et al. Methods to assess the effect of meat processing on viability of Toxoplasma gondii: Towards replacement of mouse bioassay by in vitro testing. Int. J. Parasitol. 2020, 50, 357–369. [Google Scholar] [CrossRef]
- Opsteegh, M.; Maas, M.; Schares, G.; van der Giessen, J. Relationship between seroprevalence in the main livestock species and presence of Toxoplasma gondii in meat (GP/EFSA/BIOHAZ/2013/01) An extensive literature review, Final report. EFSA Support Publ. 2016, 13, 1–294. [Google Scholar] [CrossRef]
- Zdolec, N.; Kiš, M. Meat safety from farm to slaughter—Risk-based control of Yersinia enterocolitica and Toxoplasma gondii. Processes 2021, 9, 815. [Google Scholar] [CrossRef]
- Wehbe, K.; Pencole, L.; Lhuaire, M.; Sibiude, J.; Mandelbrot, L.; Villena, I.; Picone, O. Hygiene measures as primary prevention of toxoplasmosis during pregnancy: A systematic review. J. Gynecol. Obstet. Hum. Reprod. 2022, 51, 102300. [Google Scholar] [CrossRef] [PubMed]
- EU. Commission Regulation No. 219/2014 Commission Regulation (EU) No 219/2014 of 7 March 2014 amending Annex I to Regulation (EC) No 854/2004 of the European Parliament and of the Council as regards the specific requirements for post-mortem inspection of domestic swine. Off. J. Eur. Union 2014, L 69, 99–100. [Google Scholar]
- EFSA (European Food Safety Agency). Scientific Opinion on the public health hazard to be covered by inspection of meat (swine). EFSA J. 2011, 9, 2351. [Google Scholar] [CrossRef]
- Rani, S.; Pradhan, A.K. Evaluating uncertainty and variability associated with Toxoplasma gondii survival during cooking and low temperature storage of fresh cut meats. Int. J. Food Microbiol. 2021, 341, 109031. [Google Scholar] [CrossRef]
- Buxton, D.; Thomson, K.; Maley, S.; Wright, S.; Bos, H.J. Vaccination of sheep with a live incomplete strain (S48) of Toxoplasma gondii and their immunity to challenge when pregnant. Vet. Rec. 1991, 129, 89–93. [Google Scholar] [CrossRef]
- Yuan, Z.G.; Zhang, X.X.; Lin, R.Q.; Petersen, E.; He, S.; Yu, M.; He, X.H.; Zhou, D.H.; He, Y.; Li, H.X.; et al. Protective effect against toxoplasmosis in mice induced by DNA immunization with gene encoding Toxoplasma gondii ROP18. Vaccine 2011, 29, 6614–6619. [Google Scholar] [CrossRef]
- Bonačić Marinović, A.A.; Opsteegh, M.; Deng, H.; Suijkerbuijk, A.W.M.; van Gils, P.F.; van der Giessen, J. Prospects of toxoplasmosis control by cat vaccination. Epidemics 2020, 30, 100380. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Esteban, J.; Bargues, M. Epidemiology of human fascioliasis: A review and proposed new classification. Bull. WHO 1999, 77, 340–346. [Google Scholar]
- Chen, M.G.; Mott, K.E. Progress in assessment of morbidity due to Fasciola hepatica infection: A review of recent literature. Trop Dis. Bull 1990, 87, 1–38. [Google Scholar]
- WHO. Working to Overcome the Global Impact of Neglected Tropical Diseases. First WHO Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar] [CrossRef]
- Fürst, T.; Keiser, J.; Utzinger, J. Global burden of human food-borne trematodiasis: A systematic review and meta-analysis. Lancet Inf. Dis. 2012, 12, 210–221. [Google Scholar] [CrossRef]
- Havelaar, A.; Kirk, M.; Torgerson, P.; Gibb, H.; Hald, T.; Lake, R.; Praet, N.; Bellinger, D.; de Silva, N.; Gargouri, N.; et al. Disease Burden Epidemiology Reference Group. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef]
- Hopkins, D. Homing in on helminths. Am. J. Trop. 1992, 46, 626–634. [Google Scholar] [CrossRef]
- Rim, H.; Madsen, H.; Cross, J.H.; Rim, H.; Farag, H.F.; Sornmani, S. Food-borne trematodes: Ignored or emerging? Food-borne Ignored or Emerging? Parasitol. Today 1994, 10, 207–209. [Google Scholar] [CrossRef]
- Keiser, J.; Utzinger, J. Emerging foodborne trematodiasis. Emerg. Inf. Dis. 2005, 11, 1507–1514. [Google Scholar] [CrossRef]
- WHO. Control of Foodborne Trematode infections: Report of a WHO Study Group; 1995 WHO Technical Report Series 894; WHO: Geneva, Switzerland, 1995. [Google Scholar]
- Dermauw, V.; Muchai, J.; Kappany, Y.; Al Fajardo, A.L.; Dorny, P. Human fascioliasis in Africa: A systematic review. PLoS ONE 2021, 16, e0261166. [Google Scholar] [CrossRef] [PubMed]
- Dorny, P.; Praet, N.; Deckers, N.; Gabriel, S. Emerging food-borne parasites. Vet. Parasitol. 2009, 163, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Mas-Coma, S.; Valero, M.; Bargues, M. Chapter 2—Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission, epidemiology, evolutionary genetics, molecular epidemiology and control. In Advances in Parasitology, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar] [CrossRef]
- Fox, N.J.; White, P.C.L.; McClean, C.J.; Marion, G.; Evans, A.; Hutchings, M.R. Predicting impacts of climate change on fasciola hepatica risk. PLoS ONE 2011, 6, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Climate change effects on trematodiases, with emphasis on zoonotic fascioliasis and schistosomiasis. Vet. Parasitol. 2009, 163, 264–280. [Google Scholar] [CrossRef]
- Mas-Coma, S. Human fascioliasis emergence risks in developed countries: From individual patients and small epidemics to climate and global change impacts. Enferm. Infec. y Microbiol. Clinic. 2020, 38, 253–256. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Bargues, M.D.; Valero, M.A. Human fascioliasis infection sources, their diversity, incidence factors, analytical methods and prevention measures. Parasitology 2018, 145, 1665–1699. [Google Scholar] [CrossRef]
- Keiser, J.; Utzinger, J. Food-borne trematodiases. Clin. Microbiol. Rev. 2009, 22, 466–483. [Google Scholar] [CrossRef]
- Mas-Coma, S. Human Fascoliasis: Epidemiological patterns in human endemic areas of South America, Africa and Asia. Southeast Asian J. Trop. Med. Public Health 2004, 35, 1–11. [Google Scholar]
- Taghipour, A.; Zaki, L.; Rostami, A.; Foroutan, M.; Ghaffarifar, F.; Fathi, A.; Abdoli, A. Highlights of human ectopic fascioliasis: A systematic review. Inf. Dis. 2019, 51, 785–792. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Agramunt, V.; Valero, M. Chapter 2. Neurological and Ocular Fascioliasis in Humans. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2014; Volume 84. [Google Scholar] [CrossRef]
- González-Miguel, J.; Valero, M.A.; Reguera-Gomez, M.; Mas-Bargues, C.; Bargues, M.D.; Simón, F.; Mas-Coma, S. Numerous Fasciola plasminogen-binding proteins may underlie blood-brain barrier leakage and explain neurological disorder complexity and heterogeneity in the acute and chronic phases of human fascioliasis. Parasitol 2019, 146, 284–298. [Google Scholar] [CrossRef]
- Mailles, A.; Capek, A.; Ajana, F.; Schepens, C.; Ilef, D.; Vaillant, V. Commercial watercress as an emerging source of fascioliasis in Northern France in 2002: Results from an outbreak investigation. Epidemiol. Inf. 2006, 134, 942–945. [Google Scholar] [CrossRef]
- Micic, D.; Oto, A.; Charlton, M.R.; Benoit, J.-L.; Siegler, M. Hiding in the Water. N. Engl. J. Med. 2020, 382, 1844–1849. [Google Scholar] [CrossRef]
- Sah, R.; Khadka, S.; Khadka, M.; Gurubacharya, D.; Sherchand, J.B.; Parajuli, K.; Shah, N.P.; Kattel, H.P.; Pokharel, B.M.; Rijal, B. Human fascioliasis by Fasciola hepatica: The first case report in Nepal. BMC Res. Notes 2017, 10, 10–13. [Google Scholar] [CrossRef]
- Goral, V.; Senturk, S.; Mete, O.; Cicek, M.; Ebik, B.; Kaya, B. A Case of Biliary Fascioliasis by Fasciola gigantica in Turkey. Korean J. Parasitol. 2011, 49, 65–68. [Google Scholar] [CrossRef]
- Nino Incani, R.; Vieira, J.; Pacheco, M.; Planchart, S.; Amarista, M.; Lazdins, J. Human infection by Fasciola hepatica in Venezuela: Report of a geriatric case. Investig. Clin. 2003, 44, 255–260. [Google Scholar]
- Ashrafi, K.; Valero, M.A.; Massoud, J.; Sobhani, A.; Solaymani-Mohammadi, S.; Conde, P.; Khoubbane, M.; Bargues, M.D.; Mas-Coma, S. Plant-borne human contamination by fascioliasis. Am. J. Trop. Med. Hyg. 2006, 75, 295–302. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Milas, S.; Rossi, C.; Philippart, I.; Dorny, P.; Bottieau, E. Autochthonous human Fascioliasis, Belgium. Em. Infect. Dis. 2020, 26, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ulloa, C.; Rivera-jacinto, M.; del Valle-Mendoza, J.; Cerna, C.; Hoban, C.; Ortiz, P. Risk factors for human fascioliasis in schoolchildren in Baños del Inca. Trans. Royal Soc. Trop. Med. Hyg. 2018, 112, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Park, S.; Hong, S.; Shin, H.; Jung, B.K.; Kim, M.J. Green vegetable juice as a potential source of human fascioliasis in Korea. One Health 2022, 15, 100441. [Google Scholar] [CrossRef] [PubMed]
- Curtale, F.; Mas-Coma, S.; Hassanein, Y.A.W.; Barduagni, P.; Pezzotti, P.; Savioli, L. Clinical signs and household characteristics associated with human fascioliasis among rural population in Egypt: A case-control study. Parassitologia 2003, 45, 5–11. [Google Scholar]
- Esteban, J.G.; Gonzalez, C.; Curtale, F.; Muñoz-Antoli, C.; Valero, M.; Bargues, M.; El Sayed, M.; El Wakeel, A.; Abdel-Wahab, Y.; Montresor, A.; et al. Hyperendemic fascioliasis associated with schistosomiasis in villages in the Nile Delta of Egypt. Am J Trop. Med. Hyg. 2003, 69, 429–437. [Google Scholar] [CrossRef]
- Mas-Coma, S. Human fascioliasis. In Waterborne Zoonoses: Identification, Causes and Control; IWA Publish: London, UK, 2004. [Google Scholar]
- Taira, N.; Yoshifuji, H.; Boray, J.C. Zoonotic potential of infection with Fasciola spp. by consumption of freshly prepared raw liver containing immature flukes. Int. J. Parasitol. 1997, 27, 775–779. [Google Scholar] [CrossRef]
- Dreyfuss, G.; Vignoles, P.; Rondelaud, D. Fasciola hepatica: Epidemiological surveillance of natural watercress beds in central France. Parasitol. Res. 2005, 95, 278–282. [Google Scholar] [CrossRef]
- Gaasenbeek, C.P.H.; Over, H.J.; Noorman, N.; Leeuw, W.A.; An, W.L. An epidemiological study of Fasciola hepatica in the Netherlands. Vet. Q. 2011, 2176, 140–144. [Google Scholar] [CrossRef]
- Ueno, H. Metacercaria detecting buoy (MDB) method for estimating contamination of rice fields with Fasciola metacercariae. Jarq 1976, 10, 149–152. [Google Scholar]
- El-Sayad, M.; Allam, A.; Osman, M. Prevention of human fascioliasis: A study on the role of acids detergents and potassium permenganate in clearing salads from metacercariae. J. Egyptical. Soc. Parasitol. 1997, 27, 163–169. [Google Scholar]
- Hassan, A.; Shoukary, N.; El-Motayam, M.; Morsy, A. Efficacy of five chemicals on Fasciola gigantica encysted metacercariae infectivity. J. Egyptical. Soc. Parasitol. 2008, 38, 919–928. [Google Scholar]
- Fennell, J. Potential for Watercress Production in AUSTRALIA. Rural Industries Research and Development Corporation, Australian Government, Report; RIRDC Publication No. 06/105, Project No. DAS-49A (Issue 06); RIRDC Publication: Canberra, Australia.
- King, S.; Scholz, T. Trematodes of the family Opisthorchiidae: A minireview. Korean J. Parasitol. 2001, 39, 209–221. [Google Scholar] [CrossRef][Green Version]
- Bürli, C.; Harbrecht, H.; Odermatt, P.; Sayasone, S.; Chitnis, N. Mathematical analysis of the transmission dynamics of the liver fluke, Opisthorchis viverrini. J. Theoretical. Biol. 2018, 439, 181–194. [Google Scholar] [CrossRef]
- Sripa, B.; Kaewkes, S.; Sithithaworn, P.; Mairiang, E.; Laha, T.; Smout, M.; Pairojkul, C.; Bhudhisawasdi, V.; Tesana, S.; Thinkamrop, B.; et al. Liver fluke induces cholangiocarcinoma. PLoS Med. 2007, 4, 1148–1155. [Google Scholar] [CrossRef]
- Sithithaworn, P.; Andrews, R.H.; Van De, N.; Wongsaroj, T.; Sinuon, M.; Odermatt, P.; Nawa, Y.; Liang, S.; Brindley, P.J.; Sripa, B. The current status of opisthorchiasis and clonorchiasis in the Mekong Basin. Parasitol. Int. 2012, 61, 10–16. [Google Scholar] [CrossRef]
- Kaewkes, S. Taxonomy and biology of liver flukes. Acta Trop. 2003, 88, 177–186. [Google Scholar] [CrossRef]
- Na, B.K.; Pak, J.H.; Hong, S.J. Clonorchis sinensis and clonorchiasis. Acta Trop. 2020, 203, 105309. [Google Scholar] [CrossRef]
- Sripa, B.; Tangkawattana, S.; Brindley, P.J. Update on Pathogenesis of Opisthorchiasis and Cholangiocarcinoma. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2018; Volume 12. [Google Scholar] [CrossRef]
- Woradet, S.; Promthet, S.; Songserm, N.; Parkin, D.M. Factors affecting survival time of cholangiocarcinoma patients: A prospective study in Northeast Thailand. Asian Pacific. J. Cancer Prev. 2013, 14, 1623–1627. [Google Scholar] [CrossRef]
- Gouveia, M.J.; Pakharukova, M.Y.; Laha, T.; Sripa, B.; Maksimova, G.A.; Rinaldi, G.; Brindley, P.J.; Mordvinov, V.A.; Amaro, T.; Santos, L.L.; et al. Infection with Opisthorchis felineus induces intraepithelial neoplasia of the biliary tract in a rodent model. Carcinogenesis 2017, 38, 929–937. [Google Scholar] [CrossRef]
- Pakharukova, M.Y.; Mordvinov, V.A. The liver fluke Opisthorchis felineus: Biology, epidemiology and carcinogenic potential. Trans. Royal Soc. Trop. Med. Hyg. 2015, 110, 28–36. [Google Scholar] [CrossRef] [PubMed]
- WHO. Accelerating Work to Overcome the Global Impact of Neglected Tropical Diseases—A Roadmap for Implementation; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Chai, J.; Jung, B.K. General overview of the current status of human foodborne trematodiasis. Parasitol 2022, 149, 1262–1285. [Google Scholar] [CrossRef] [PubMed]
- Murrell, K.; Pozio, E. The liver flukes: Clonorchis sinensis, Opisthorchis spp., and Metorchis spp. In Global Water Pathogen Project. Part Three. Specific Excreted Pathogens: Environmental and Epidemiology Aspects; Michigan State University: East Lansing, MI, USA, 2017; pp. 3–17. [Google Scholar]
- Rim, H. The current pathobiology and chemotherapy of clonorchiasis. Korean J. Parasitol. 1986, 24, 1–141. [Google Scholar] [CrossRef] [PubMed]
- Manivong, K.; Komalamisra, C.; Waikagul, J.; Radomyos, P. Opisthorchis viverrini Metacercariae in Cyprinoid fish from three rivers in Khammouane Province, Lao PDR. J. Trop. Med. Parasitol. 2009, 32, 23–29. [Google Scholar] [CrossRef]
- Araki, H.; Ong, K.I.C.; Lorphachan, L.; Soundala, P.; Iwagami, M.; Shibanuma, A.; Hongvanthong, B.; Brey, P.T.; Kano, S.; Jimba, M. Mothers’ Opisthorchis viverrini infection status and raw fish dish consumption in Lao People’s Democratic Republic: Determinants of child infection status. Trop. Med. Health 2018, 46, 1–7. [Google Scholar] [CrossRef]
- Nguyen, T.T.B.; Dermauw, V.; Dahma, H.; Bui, D.T.; Le, T.T.H.; Phi, N.T.T.; Lempereur, L.; Losson, B.; Vandenberg, O.; Do, D.T.; et al. Prevalence and risk factors associated with Clonorchis sinensis infections in rural communities in northern Vietnam. PLoS Negl. Trop. Dis. 2020, 14, e0008483. [Google Scholar] [CrossRef]
- Zhang, Y.; Gong, Q.L.; Lv, Q.B.; Qiu, Y.Y.; Wang, Y.C.; Qiu, H.Y.; Guo, X.R.; Gao, J.F.; Chang, Q.C.; Wang, C.R. Prevalence of Clonorchis sinensis infection in fish in South-East Asia: A systematic review and meta-analysis. J. Fish Dis. 2020, 43, 1409–1418. [Google Scholar] [CrossRef]
- Sun, J.; Xin, H.; Jiang, Z.; Qian, M.; Duan, K.; Li, W.; Huang, S.; Gan, X.; Yang, Y.; Li, Z. High endemicity of Clonorchis sinensis infection in Binyang County, Southern China. PLoS Negl. Trop. Dis. 2020, 14, e0008540. [Google Scholar] [CrossRef]
- Le, T.H.; Van De, N.; Blair, D.; Sithithaworn, P.; McManus, D.P. Clonorchis sinensis and Opisthorchis viverrini: Development of a mitochondrial-based multiplex PCR for their identification and discrimination. Exp. Parasitol. 2006, 112, 109–114. [Google Scholar] [CrossRef]
- Cai, X.Q.; Xu, M.; Wang, W.H.; Qiu, D.Y.; Liu, G.X.; Lin, A.; Tang, J.D.; Zhang, R.L.; Zhu, X.Q. Sensitive and rapid detection of Clonorchis sinensis infection in fish by loop-mediated isothermal amplification (LAMP). Parasitol. Res. 2010, 106, 1379–1383. [Google Scholar] [CrossRef]
- Lee, S.E.; Shin, H.E.; Lee, M.R.; Kim, Y.H.; Cho, S.H.; Ju, J.W. Risk factors of clonorchis sinensis human infections in endemic areas, Haman-gun, Republic of Korea: A case-control study. Korean J. Parasitol. 2020, 58, 647–652. [Google Scholar] [CrossRef]
- Qian, M.B.; Li, H.M.; Jiang, Z.H.; Yang, Y.C.; Lu, M.F.; Wei, K.; Wei, S.L.; Chen, Y.; Zhou, C.H.; Chen, Y.D.; et al. Severe hepatobiliary morbidity is associated with clonorchis sinensis infection: The evidence from a cross-sectional community study. PLoS Negl. Trop. Dis. 1 2021, 5, e0009116. [Google Scholar] [CrossRef]
- Steele, J.A.; Richter, C.H.; Echaubard, P.; Saenna, P.; Stout, V.; Sithithaworn, P.; Wilcox, B.A. Thinking beyond Opisthorchis viverrini for risk of cholangiocarcinoma in the lower Mekong region: A systematic review and meta-analysis. Inf. Dis. Poverty 2018, 7, 44. [Google Scholar] [CrossRef]
- Qian, M.B.; Utzinger, J.; Keiser, J.; Zhou, X.N. Clonorchiasis. Lancet 2016, 387, 800–810. [Google Scholar] [CrossRef]
- Wang, X.; Chen, W.; Tian, Y.; Mao, Q.; Lv, X.; Shang, M.; Li, X.; Yu, X.; Huang, Y. Surface display of Clonorchis sinensis enolase on Bacillus subtilis spores potentializes an oral vaccine candidate. Vaccine 2014, 32, 1338–1345. [Google Scholar] [CrossRef]
- Huyen, T.; Thi, D.B.; Trung, D.D.; Phi, N.; Thuy, T. Knowledge, attitude and practices related to small liver fluke infection in rural communities of Ha Trung District, Thanh Hoa Province, Vietnam. J. Kasetsart Vet. 2021, 31, 15–32. [Google Scholar]
- Sripa, B.; Tangkawattana, S.; Laha, T.; Kaewkes, S.; Mallory, F.F.; Smith, J.F.; Wilcox, B.A. Toward integrated opistorchiasis control in Northeast Thailand: The Lawa Project. Acta Trop. 2015, 141, 361–367. [Google Scholar] [CrossRef]
- Sripa, B.; Tangkawattana, S.; Sangnikul, T. The Lawa model: A sustainable, integrated opisthorchiasis control program using the EcoHealth approach in the Lawa Lake region of Thailand. Parasitol. Int. 2017, 66, 346–354. [Google Scholar] [CrossRef]
- Aibinu, I.E.; Smooker, P.M.; Lopata, A.L. Anisakis Nematodes in Fish and Shellfish- from infection to allergies. Int. J. Parasitol. Parasites Wildl. 2019, 6, 384–393. [Google Scholar] [CrossRef]
- Baird, F.J.; Gasser, R.B.; Jabbar, A.; Lopata, A.L. Foodborne anisakiasis and allergy. Mol. Cell Probes. 2014, 28, 167–174. [Google Scholar] [CrossRef]
- Mercken, E.; Van Damme, I.; Vangeenberghe, S.; Serradell, A.; De Sterck, T.; Lumain, J.P.L.; Gabriël, S. Ascaridoids in commercial fish: Occurrence, intensity and localization in whole fish and fillets destined for the Belgian market. Int. J. Food Microbiol. 2020, 16, 108657. [Google Scholar] [CrossRef] [PubMed]
- Adroher-Auroux, F.J.; Benítez-Rodríguez, R. Anisakiasis and Anisakis: An underdiagnosed emerging disease and its main etiological agents. Res. Vet. Sci. 2020, 132, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, P.; Acerra, V.; Bellisario, B.; Sbaraglia, G.L.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Larval migration of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy, Engraulis encrasicolus: Implications to seafood safety. Food Control 2016, 519, 148–157. [Google Scholar] [CrossRef]
- Bucci, C.; Gallotta, S.; Morra, I.; Fortunato, A.; Ciacci, C.; Iovino, P. Anisakis, just think about it in an emergency! Int J Infect Dis 2013, 17, e1071–e1072. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuizen, N.E. Anisakis-immunology of a foodborne parasitosis. Parasite Immunol. 2016, 38, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Mazzucco, W.; Raia, D.D.; Marotta, C.; Costa, A.; Ferrantelli, V.; Vitale, F.; Casuccio, A. Anisakis sensitization in different population groups and public health impact: A systematic review. PLoS ONE 2018, 20, e0203671. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, A.R.; Kiani, B.; Afshari, A.; Moghaddas, E.; Williams, M.; Shamsi, S. World-wide prevalence of Anisakis larvae in fish and its relationship to human allergic anisakiasis: A systematic review. Parasitol. Res. 2020, 119, 3585–3594. [Google Scholar] [CrossRef]
- Añíbarro, B.; Seoane, F.J.; Múgica, M.V. Involvement of hidden allergens in food allergic reactions. J. Investig. Allergol. Clin. Immunol. 2007, 17, 168–172. [Google Scholar]
- Armentia, A.; Martin-Gil, F.J.; Pascual, C.; Martín-Esteban, M.; Callejo, A.; Martínez, C. Anisakis simplex allergy after eating chicken meat. J. Investig. Allergol. Clin. Immunol. 2006, 16, 258–263. [Google Scholar]
- Fæste, C.; Levsen, A.; Lin, A.H.; Larsen, N.; Plassen, C.; Moen, A.; Van Do, T.; Egaas, E. Fish feed as source of potentially allergenic peptides from the fish parasite Anisakis simplex (s.l.). Anim. Feed Sci. Technol. 2015, 202, 52–61. [Google Scholar] [CrossRef]
- Polimeno, L.; Lisanti, M.T.; Rossini, M.; Giacovazzo, E.; Polimeno, L.; Debellis, L.; Ballini, A.; Topi, S.; Santacroce, L. Anisakis Allergy: Is aquacultured fish a safe and alternative food to wild-capture fisheries for Anisakis simplex-sensitized patients? Biology 2021, 10, 106. [Google Scholar] [CrossRef]
- Saelens, G.; Planckaert, S.; Devreese, B.; Gabriël, S. Transmissibility of anisakid allergenic peptides from animal feed to chicken meat: Proof of concept. J. Food Comp. Anal. 2023, 115, 104939. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Lopata, A.L. Anisakis-A food-borne parasite that triggers allergic host defences. Int. J. Parasitol. 2013, 43, 1047–1057. [Google Scholar] [CrossRef]
- Ivanović, J.; Baltić, M.Ž.; Bošković, M.; Kilibarda, N.; Dokmanović, M.; Marković, R.; Janjić, J.; Baltić, B. Anisakis allergy in human. Trends Food Sci. Technol. 2017, 59, 25–29. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Lopata, A. Allergic reactions to Anisakis found in fish. Curr. Allergy Asthma Rep. 2014, 14, 455. [Google Scholar] [CrossRef]
- Levsen, A.; Lunestad, B.T.; Berland, B. Low detection dfficiency of candling as a commonly recommended inspection method for nematode larvae in the flesh of Pelagic fish. J. Food Protect 2005, 68, 828–832. [Google Scholar] [CrossRef]
- Karl, H.; Leinemann, M. A fast and quantitative detection method for nematodes in fish fillets and fishery products. Arch. Lebensmittelhyg. 1993, 44, 124–125. [Google Scholar]
- Petrie, A.; Wootten, R.; Bruno, D.; MacKenzie, K.; Bron, J. A Survey of Anisakis and Pseudoterranova in Scottish Fisheries and the Efficacy of Current Detection Methods FSAS Project S14008; Food Standards Scotland: Aberdeen, UK, 2007. [Google Scholar]
- Mercken, E.; Van Damme, I.; Šoba Šparl, B.; Vangeenberghe, S.; Serradell, A.; De Sterck, T. Sensitivity of candling as routine method for the detection and recovery of ascaridoids in commercial fish fillets. Sci. Rep. 2022, 12, 1358. [Google Scholar] [CrossRef]
- Pippy, J. Use of ultraviolet light to find parasitic nematodes in situ. J. Fish Res. Board. Can. 1970, 27, 963–965. [Google Scholar] [CrossRef]
- Llarena-Reino, M.; Piñeiro, C.; Antonio, J.; Outeriño, L.; Vello, C.; González, Á.F.; Pascual, S. Optimization of the pepsin digestion method for anisakids inspection in the fishing industry. Vet. Parasitol. 2013, 191, 276–283. [Google Scholar] [CrossRef]
- Kochanowski, M.; Różycki, M.; Dąbrowska, J.; Karamon, J.; Sroka, J.; Antolak, E.; Bełcik, A.; Cencek, T. Development and application of novel chemiluminescence immunoassays for highly sensitive detection of Anisakis simplex proteins in thermally processed seafood. Pathogens 2020, 23, 777. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.T.; Fæste, C.K.; Levsen, A.; Egaas, E. A quantitative sandwich ELISA for the detection of Anisakis simplex protein in seafood. Eur. Food Res. Technol. 2011, 232, 967–973. [Google Scholar] [CrossRef]
- Fæste, C.K.; Moen, A.; Schniedewind, B.; Haug Anonsen, J.; Klawitter, J.; Christians, U. Development of liquid chromatography-tandem mass spectrometry methods for the quantitation of Anisakis simplex proteins in fish. J. Chromatogr. A 2016, 5, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mahillo, A.I.; González-Muñoz, M.; de las Heras, C.; Tejada, M.; Moneo, I. Quantification of Anisakis simplex allergens in fresh, long-term frozen, and cooked fish muscle. Foodborne Pathog. Dis. 2010, 7, 967–973. [Google Scholar] [CrossRef]
- McClelland, G. Larval anisakine nematodes in various fish species from Sable Island Bank and vicinity. Can. Bull Fish Aquat. Sci. 1990, 222, 83–118. [Google Scholar]
- EU. Commission regulation no 1276/2011 of 8 December 2011 amending annex III to regulation (EC) no 853/2004 of the European Parliament and of the council as regards the treatment to kill viable parasites in fishery products for human consumption. Off. J. Eur. Union 2011, L 327, 39–41. [Google Scholar]
- FDA. Parasites. In Fish and Fishery Products Hazards and Controls Guidance, 4th ed.; Food and Drug Administration, U.S. Department of Health and Human Services: Silver Spring, MD, USA, 2019; pp. 91–98. [Google Scholar]
- Chen, H.Y.; Cheng, Y.S.; Grabner, D.S.; Chang, S.H.; Shih, H.H. Effect of different temperatures on the expression of the newly characterized heat shock protein 90 (Hsp90) in L3 of Anisakis spp. isolated from Scomber australasicus. Vet. Parasitol. 2014, 205, 540–550. [Google Scholar] [CrossRef]
- Donadeu, M.; Fahrion, A.S.; Olliaro, P.L.; Abela-Ridder, B. Target product profiles for the diagnosis of Taenia solium taeniasis, neurocysticercosis and porcine cysticercosis. PLoS Negl. Trop. Dis. 2017, 11, e0005875. [Google Scholar] [CrossRef]
- Rodrigues da Costa, M.; Pessoa, J.; Meemken, D.; Nesbakken, T. A systematic review on the effectiveness of pre-harvest meat safety interventions in pig herds to control Salmonella and other foodborne pathogens. Microorganisms 2021, 9, 1825. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).