
A Comparison Study on the Therapeutic Effect of High Protein Diets Based on Pork Protein versus Soybean Protein on Obese Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Samples and Animal Diets
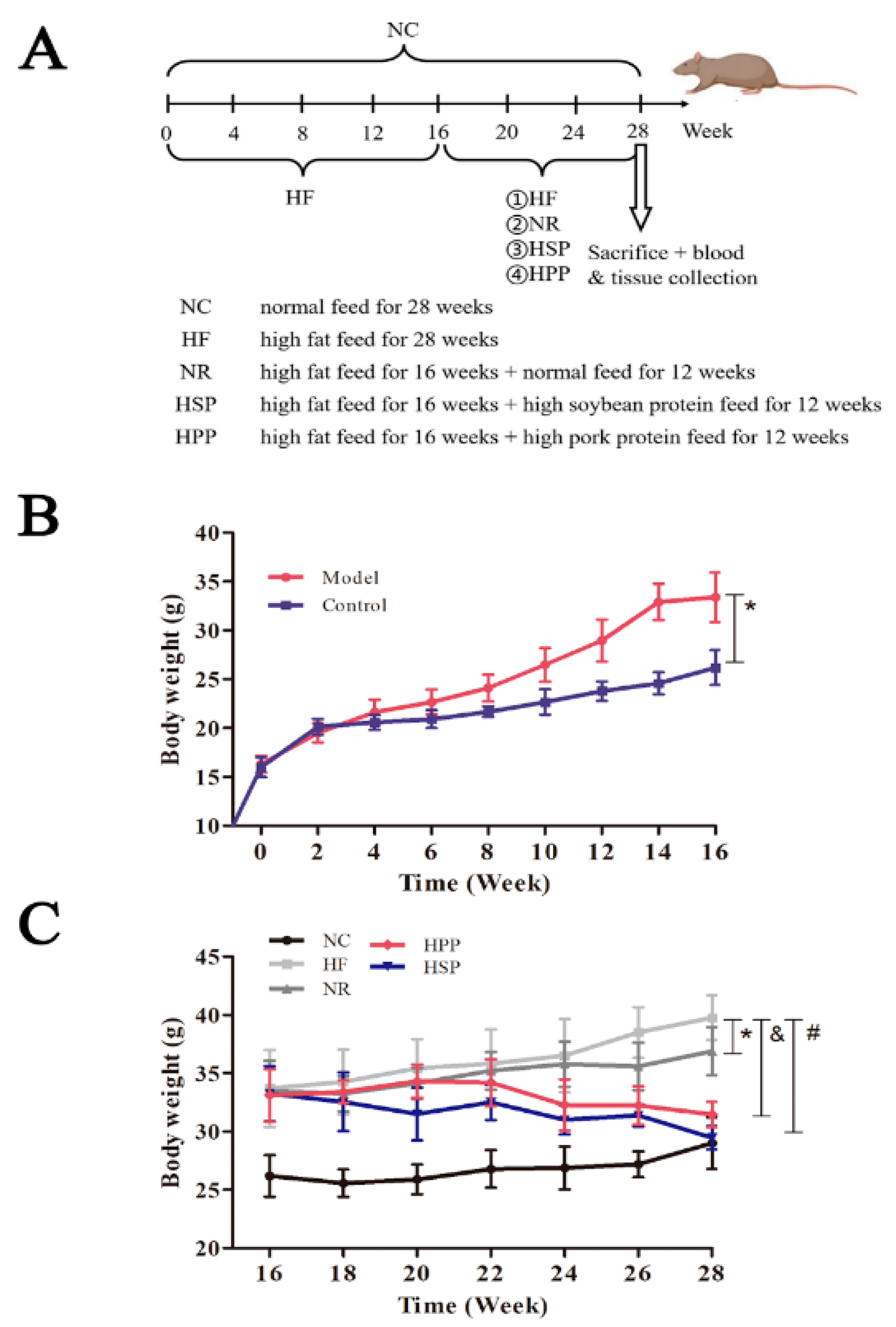
2.2. Experimental Protocol
2.3. Determination of Body Weight and Fat Coefficients
2.4. Biochemical Analysis
2.5. Hematoxylin-Eosin (HE) Staining and Oil Red O (ORO) Staining
2.6. Determination of Amino Acid Levels
2.7. RNA Extraction and Quantitative Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-qPCR)
2.8. Western Blot
2.9. Statistical Analysis
3. Results
3.1. Changes in Body Weight and Physiological Status of Mice
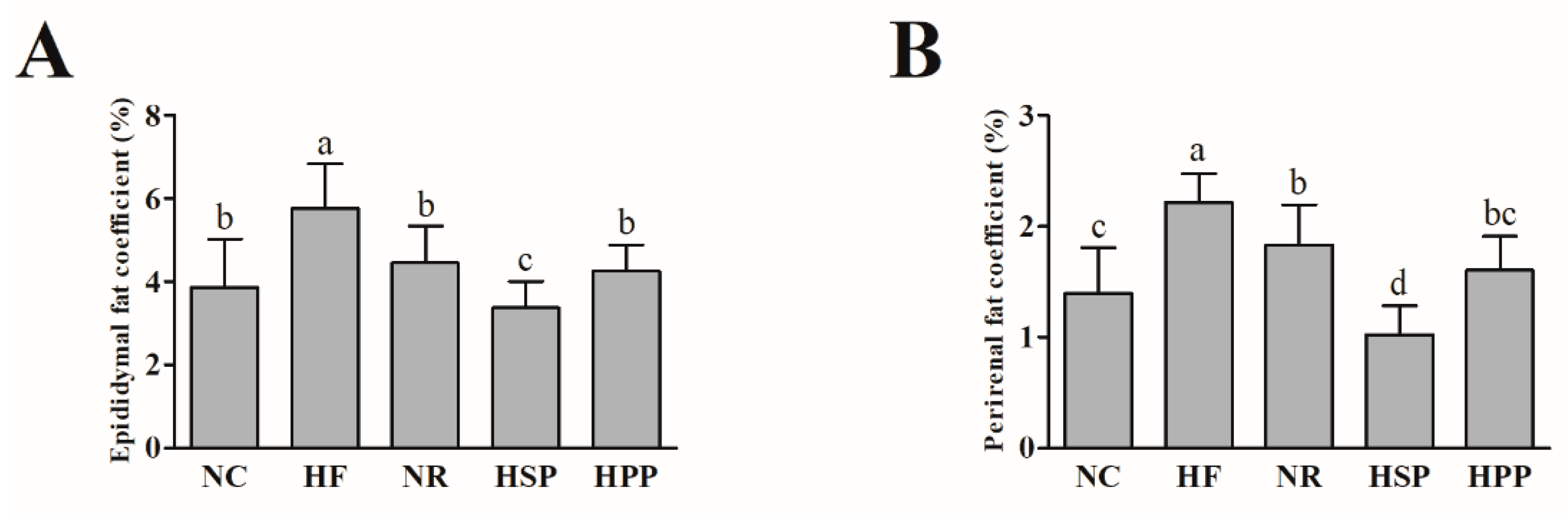
3.2. Effect of Different Proteins on Epididymis and Perirenal Fat Coefficients in Obese Mice
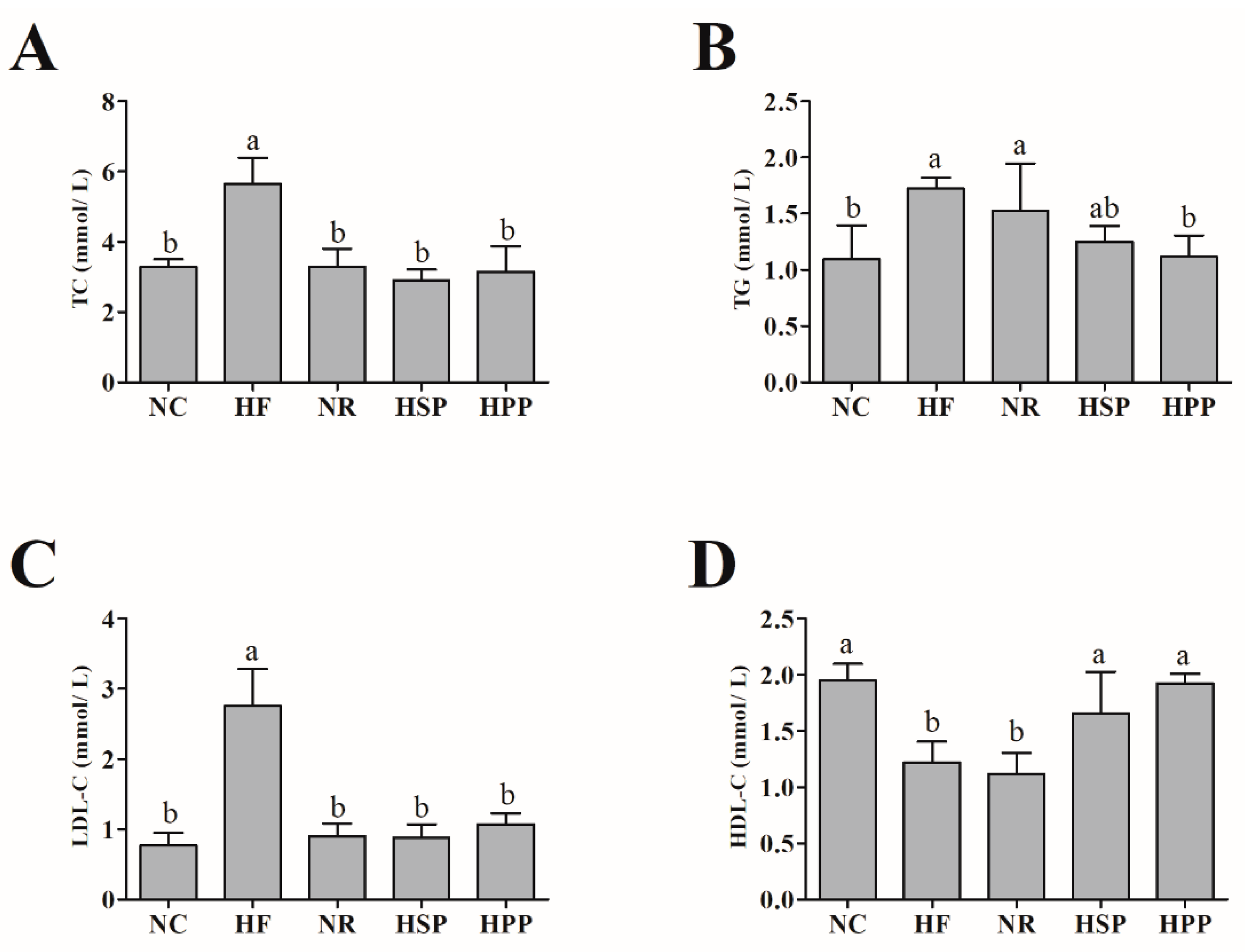
3.3. Effect of Different Proteins on Serum TC, TG, LDL-C, and HDL-C in Obese Mice
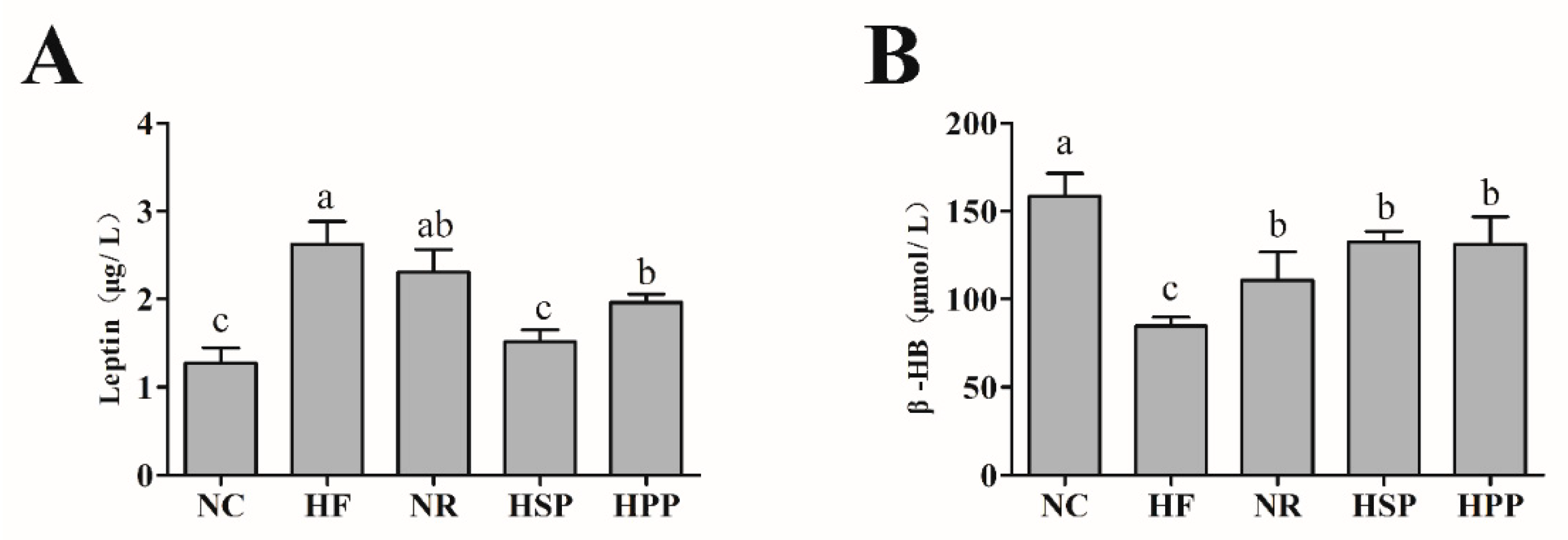
3.4. Effect of Different Proteins on Serum Leptin and β-HB in Obese Mice
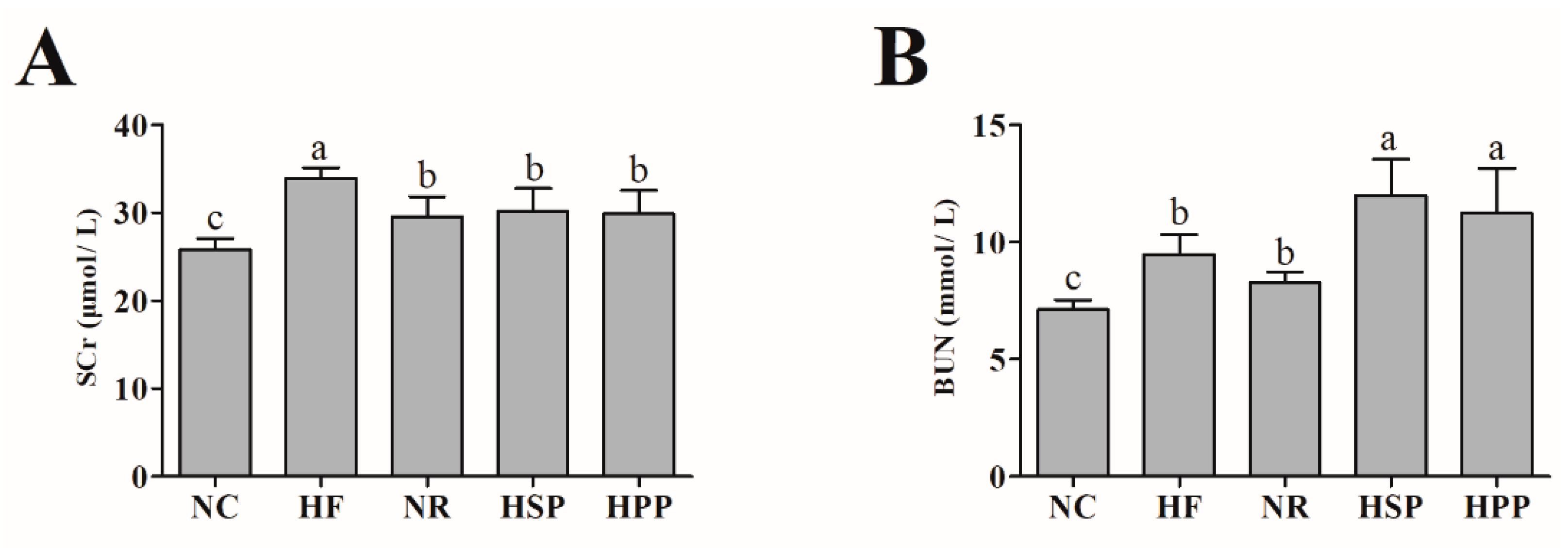
3.5. Effect of Different Proteins on SCr and BUN in Obese Mice
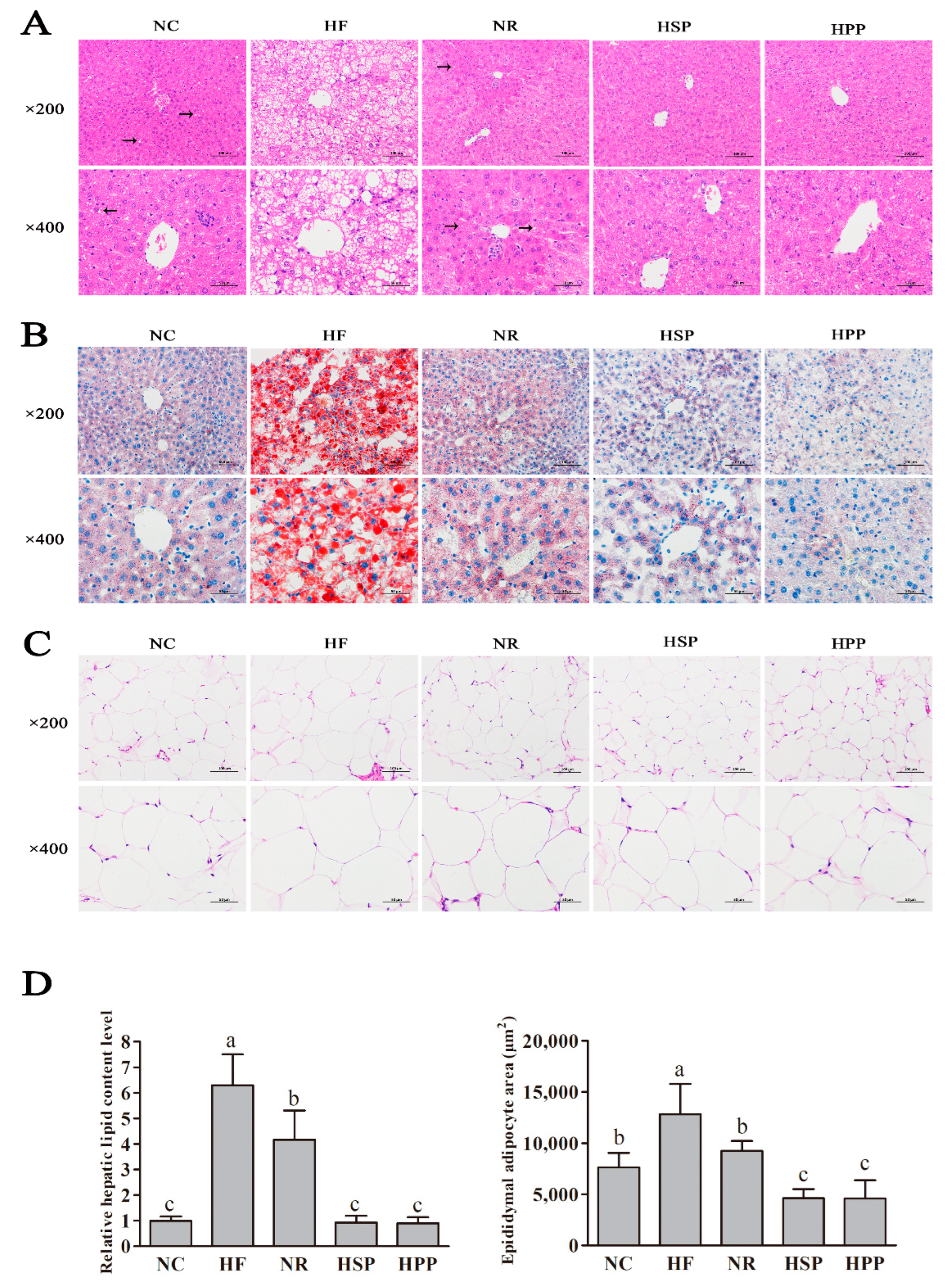
3.6. Effect of Different Proteins on Liver and Fat Histomorphology in Obese Mice
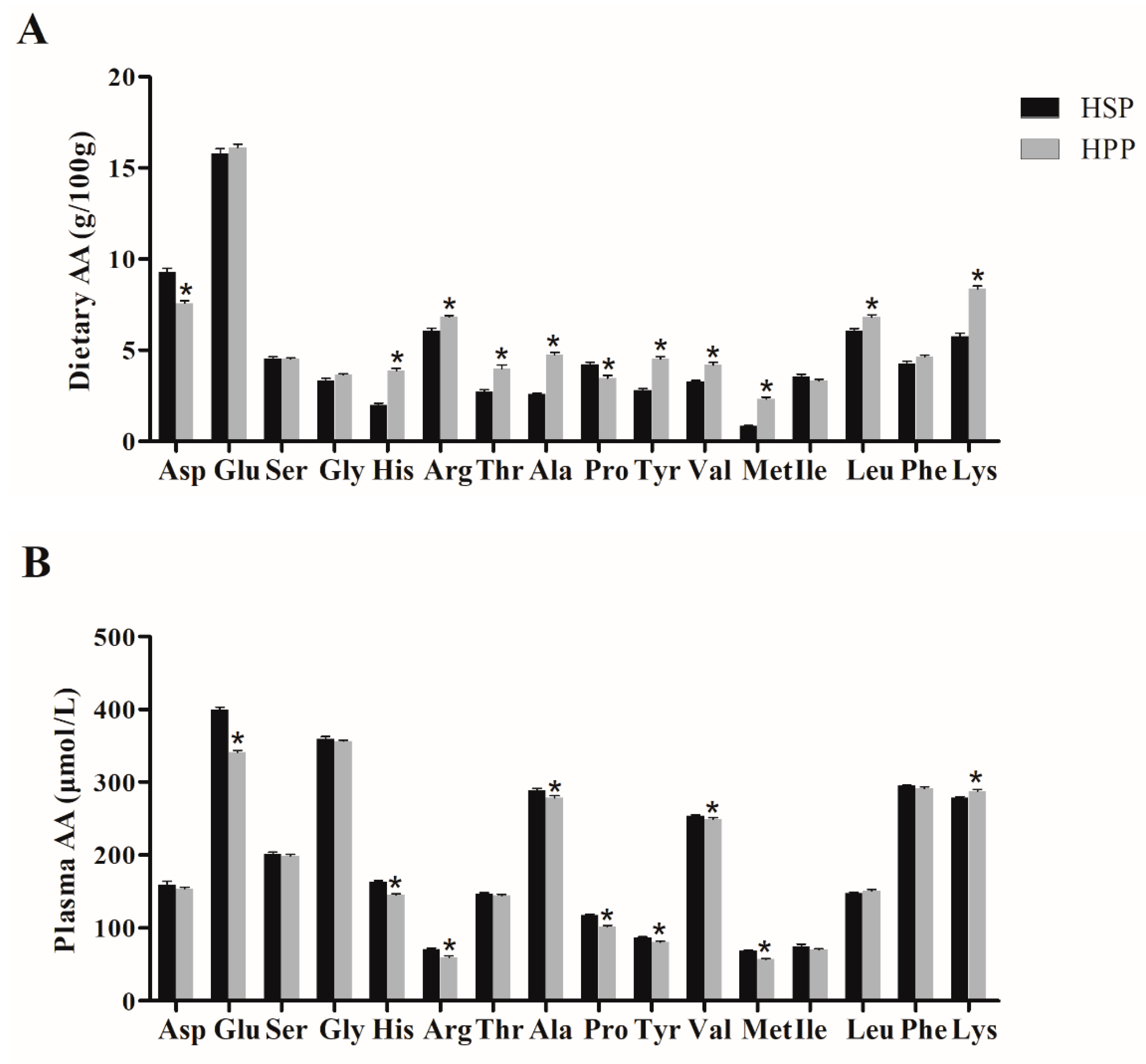
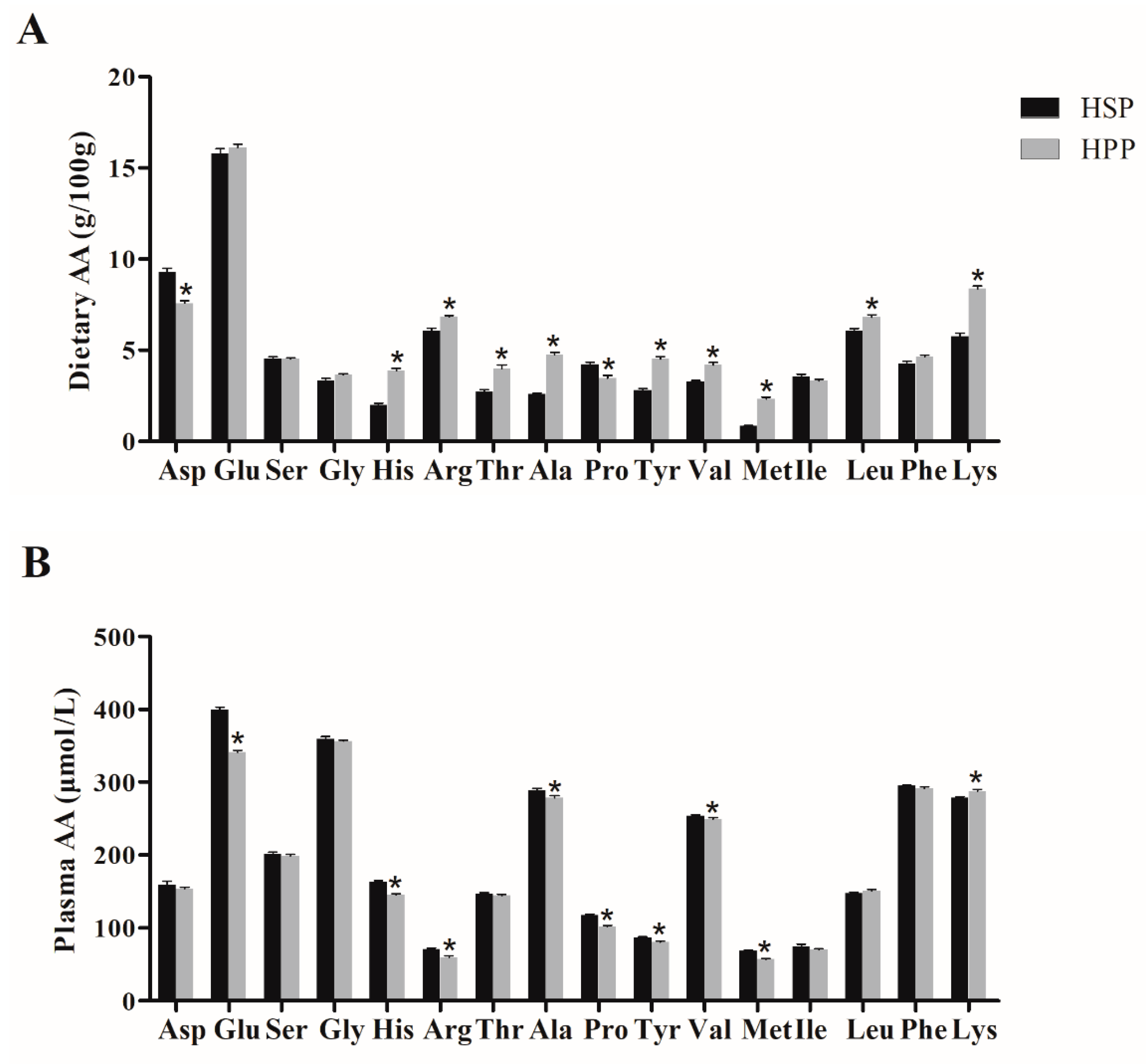
3.7. Dietary Amino Acid (AA) Components of Different Proteins and Effect on Plasma Amino Acids in Obese Mice
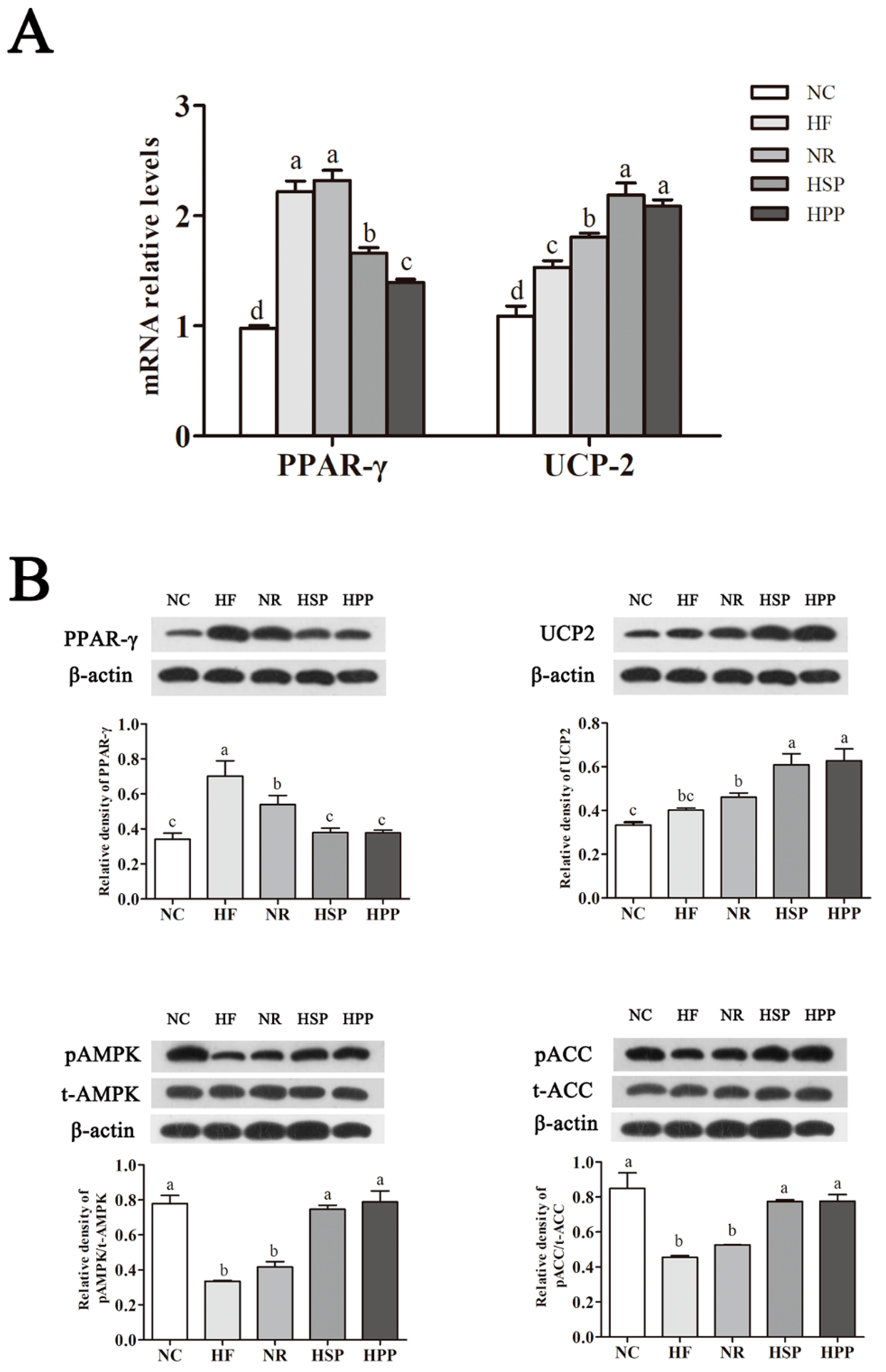
3.8. Effect of Different Protein Sources on the Expression of Hepatic PPAR-γ, UCP2, pAMPK, and pACC in Obese Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghanemi, A.; St-Amand, J. Redefining obesity toward classifying as a disease. Eur. J. Intern. Med. 2018, 55, 20–22. [Google Scholar] [CrossRef]
- 8th Fribourg Obesity Research Conference (FORC-2015) Abstracts: ‘Nutrition, Movement & Sleep Behaviors: Their interactions in pathways to Obesity and Cardiometabolic diseases’. Obes. Rev. 2017, 18 (Suppl. 1), 88–93.
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; McGowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The obesity transition: Stages of the global epidemic. Lancet Diabetes Endo. 2019, 7, 231–240. [Google Scholar] [CrossRef]
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 9 February 2022).
- Wang, Y.; Sun, M.; Xue, H.; Zhao, W.; Yang, X.; Zhu, X.; Zhao, L.; Yang, Y. Understanding the China Blue Paper on Obesity Prevention and Control and policy implications and recommendations for obesity prevention and control in China. Chin. J. Prev. Med. 2019, 53, 875–884. [Google Scholar]
- Bing, G.; Si-Qi, L.; Shu-Feng, L. The obesity indices mediate the relationships of blood lipids and bone mineral density in Chinese elders. Mol. Cell. Probes 2021, 56, 101705–102709. [Google Scholar]
- Jeong, N.I.; Sun, P.J.; Byul, P.S. Association between Abdominal Obesity and Oxidative Stress in Korean Adults. Korean J. Fam. Med. 2019, 40, 395–398. [Google Scholar]
- Musaad, S.; Haynes, E.N. Biomarkers of obesity and subsequent cardiovascular events. Epidemiol. Rev. 2007, 29, 98–114. [Google Scholar] [CrossRef] [Green Version]
- Bramante, C.T.; Lee, C.J.; Gudzune, K.A. Treatment of Obesity in Patients With Diabetes. Diabetes Spectr. 2017, 30, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popkin, B.M.; Du, S.; Green, W.D.; Beck, M.A.; Algaith, T.; Herbst, C.H.; Alsukait, R.F.; Alluhidan, M.; Alazemi, N.; Shekar, M. Individuals with obesity and COVID-19: A global perspective on the epidemiology and biological relationships. Obes. Rev. 2020, 21, 13128–13144. [Google Scholar] [CrossRef]
- van Baak, M.A.; Larsen, T.M.; Jebb, S.A.; Martinez, A.; Saris, W.H.M.; Handjieva-Darlenska, T.; Kafatos, A.; Pfeiffer, A.F.H.; Kunešová, M.; Astrup, A. Dietary Intake of Protein from Different Sources and Weight Regain, Changes in Body Composition and Cardiometabolic Risk Factors after Weight Loss: The DIOGenes Study. Nutrients 2017, 9, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, E.; Choi, S.K.; Joung, H. Impact of dietary protein intake and obesity on lean mass in middle-aged individuals after a 12-year follow-up: The Korean Genome and Epidemiology Study (KoGES). Br. J. Nutr. 2019, 122, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Yue, L.; Ci, H.; Ci, X.; Wang, H. Effect of high protein diet intervention on obesity and chronic diseases. Chin. Bull. Life Sci. 2020, 32, 170–178. [Google Scholar]
- Speaker, K.J.; Sayer, R.D.; Peters, J.C.; Foley, H.N.; Pan, Z.; Wyatt, H.R.; Flock, M.R.; Mukherjea, R.; Hill, J.O. Effects of consuming a high-protein diet with or without soy protein during weight loss and maintenance: A non-inferiority, randomized clinical efficacy trial. Obes. Sci. Pract. 2018, 4, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Shumei, X.; Ying, X. Protein intake and obesity in young adolescents. Exp. Ther. Med. 2016, 11, 1545–1549. [Google Scholar]
- Pereira, P.M.d.C.C.; Vicente, A.F.d.R.B. Meat nutritional composition and nutritive role in the human diet. Meat Sci. 2013, 93, 586–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, L.; Liu, R.; Cao, S.; Zhang, W.; Guanghong, Z. Meat protein based bioactive peptides and their potential functional activity: A review. Int. J. Food Sci. Technol. 2019, 54, 1956–1966. [Google Scholar] [CrossRef] [Green Version]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Regulators of the transsulfuration pathway. Br. J. Pharmacol. 2019, 176, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.-A.; Bendsen, N.T.; Tremblay, A.; Astrup, A. Effect of proteins from different sources on body composition. Nutr. Metab. Cardiovasc. Dis. 2010, 21, B16–B31. [Google Scholar] [CrossRef] [PubMed]
- Millward, D.J.; Layman, D.K.; Tomé, D.; Schaafsma, G. Protein quality assessment: Impact of expanding understanding of protein and amino acid needs for optimal health. Am. J. Clin. Nutr. 2008, 87, 1576S–1581S. [Google Scholar] [CrossRef] [PubMed]
- Song, S. Effects of Different Dietary Protein Sources on Growth, Blood Metabolic Items, Liver Transcriptomics and Proteomics of Rats. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besten, G.D.; Bleeker, A.; Gerding, A.; Eunen, K.V.; Havinga, R.; Dijk, T.V.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Chen, F.; Ye, H.; Yin, Y.; Sun, J.; Wu, Y. Effect of different treatment on expression of tumor necrosis factor-alpha in adipose tissues of high fat diet-induced obese mice. Shanghai Med. J. 2010, 33, 81–84. [Google Scholar]
- Yang, Q.; Wang, J.-H.; Huang, D.-D.; Li, D.-G.; Chen, B.; Zhang, L.-M.; Yuan, C.-L.; Cai, L.-J. Clinical significance of analysis of the level of blood fat, CRP and hemorheological indicators in the diagnosis of elder coronary heart disease. Saudi J. Biol. Sci. 2018, 25, 1812–1816. [Google Scholar] [CrossRef]
- Yuanyuan, S.; Qigui, Y.; Jun, X. Investigating the changes in the levels of HbA1c, blood fat and insulin sensitivity in elder patients with type II diabetes mellitus due to combined medication of pioglitazone and melbine and single-use of pioglitazone. Cell. Mol. Biol. (Noisy-Le-Grand) 2020, 66, 21–28. [Google Scholar]
- Tovar, A.R.; Torre-Villalvazo, I.; Ochoa, M.; Elias, A.L.; Ortiz, V.; Aguilar-Salina, C.A.; Torres, N. Soy protein reduces hepatic lipotoxicity in hyperinsulinemic obese Zucker fa/fa rats. J. Lipid. Res. 2005, 46, 1823–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuela, M.; Barbara, L.; Francesca, A. Sarcopenic obesity in fatty liver. Curr. Opin. Clin. Nutr. 2019, 22, 185–190. [Google Scholar]
- Zhang, Y.; Wan, J.; Xu, Z.; Hua, T.; Sun, Q. Exercise ameliorates insulin resistance via regulating TGFβ-activated kinase 1 (TAK1)-mediated insulin signaling in liver of high-fat diet-induced obese rats. J. Cell. Physiol. 2019, 234, 7467–7474. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.Q.; Yang, X.F.; Ma, X.Y.; Xiong, Y.X.; Tian, Z.M.; Fan, Q.L.; Wang, L.; Jiang, Z.Y. CIDE gene expression in adipose tissue, liver, and skeletal muscle from obese and lean pigs. J. Zhejiang Univ. Sci. B 2017, 18, 492–500. [Google Scholar]
- Zhang, L.; Piao, X. Different dietary protein sources influence growth performance, antioxidant capacity, immunity, fecal microbiota and metabolites in weaned piglets. Anim. Nutr. 2022, 8, 71–81. [Google Scholar]
- Parisa, H.; Razieh, H.; Fahimeh, H.; Noushin, M.; Masoumeh, S.; Hamidreza, R.; Firoozeh, S.; Nizal, S. The long-term association of different dietary protein sources with metabolic syndrome. Sci. Rep. 2021, 11, 19394. [Google Scholar]
- Song, S.; Hooiveld, G.; Li, M.; Zhao, F.; Zhang, W.; Xu, X.; Muller, M.; Li, C.; Zhou, G. Dietary soy and meat proteins induce distinct physiological and gene expression changes in rats. Sci. Rep. 2016, 6, 20036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Hooiveld, G.; Li, M.; Zhao, F.; Zhang, W.; Xu, X.; Muller, M.; Li, C.; Zhou, G. Distinct physiological, plasma amino acid, and liver transcriptome responses to purified dietary beef, chicken, fish, and pork proteins in young rats. Mol. Nutr. Food Res. 2016, 60, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Yuki, F.; Katsuyasu, K.; Kumiko, O.; Harunobu, N.; Masayuki, I. Leptin mediates the relationship between fat mass and blood pressure: The Hamamatsu School-based health study. Medicine 2019, 98, e14934. [Google Scholar]
- Zhou, J.; Lu, Y.; Jia, Y.; Lu, J.; Jiang, Z.; Chen, K. Ketogenic diet ameliorates lipid dysregulation in type 2 diabetic mice by downregulating hepatic pescadillo 1. Mol. Med. 2022, 28, 1. [Google Scholar] [CrossRef] [PubMed]
- Liisberg, U.; Myrmel, L.S.; Fjære, E.; Rønnevik, A.K.; Bjelland, S.; Fauske, K.R.; Nolm, J.B.; Basse, A.L.; Hansen, J.B.; Liaset, B.; et al. The protein source determines the potential of high protein diets to attenuate obesity development in C57BL/6J mice. Adipocyte 2016, 5, 196–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myrmel, L.S.; Fauske, K.R.; Fjære, E.; Bernhard, A.; Liisberg, U.; Hasselberg, A.E.; Øyen, J.; Kristiansen, K.; Madsen, L. The Impact of Different Animal-Derived Protein Sources on Adiposity and Glucose Homeostasis during Ad Libitum Feeding and Energy Restriction in Already Obese Mice. Nutrients 2019, 11, 1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Huang, Z.; Zhou, G.; Li, C. Dietary Protein From Different Sources Exerted a Great Impact on Lipid Metabolism and Mitochondrial Oxidative Phosphorylation in Rat Liver. Front. Nutr. 2021, 8, 719144. [Google Scholar] [CrossRef] [PubMed]
- Brandsch, C.; Shukla, A.; Hirche, F.; Stangl, G.I.; Eder, K. Effect of proteins from beef, pork, and turkey meat on plasma and liver lipids of rats compared with casein and soy protein. Nutrition 2006, 22, 1162–1170. [Google Scholar] [CrossRef]
- Shi, X. Different Meat Protein Long-Term Feeding Effect on Rat Liver Metabolism. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2018. [Google Scholar]
- Takahashi, Y.; Ide, T. Effects of soy protein and isoflavone on hepatic fatty acid synthesis and oxidation and mRNA expression of uncoupling proteins and peroxisome proliferator-activated receptor γ in adipose tissues of rats. J. Nutr. Biochem. 2007, 19, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F.; Hojhabrimanesh, A.; Helli, B.; Seyedian, S.-S.; Ahmadi-Angali, K. An energy-restricted high-protein diet supplemented with β-cryptoxanthin alleviated oxidative stress and inflammation in nonalcoholic fatty liver disease: A randomized controlled trial. Nutr. Res. 2020, 73, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yan, H.; Geng, Y.; Shi, H.; Li, H.; Wang, S.; Wang, Y.; Xu, J.; Zhao, G.; Lu, X. Target genes associated with lipid and glucose metabolism in non-alcoholic fatty liver disease. Lipids Health Dis. 2019, 18, 211–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gettys, T.W. Dietary methionine restriction: An approach to treating obesity that does not involve food restriction. J. Anim. Sci. 2020, 98, 66. [Google Scholar] [CrossRef]
- Wang, L.; Ren, B.; Zhang, Q.; Chu, C.; Zhao, Z.; Wu, J.; Zhao, W.; Liu, Z.; Liu, X. Methionine restriction alleviates high-fat diet-induced obesity: Involvement of diurnal metabolism of lipids and bile acids. BBA-Mol. Basis Dis. 2020, 1866, 165908. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, A.; Petzke, K.J.; Klaus, S. Dietary L-leucine and L-alanine supplementation have similar acute effects in the prevention of high-fat diet-induced obesity. Amino Acids 2013, 44, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Stone, K.P.; Forney, L.A.; Sims, L.C.; Gutierrez, G.C.; Ghosh, S.; Gettys, T.W. Implementation of dietary methionine restriction using casein after selective, oxidative deletion of methionine. iScience 2021, 24, 102470–102491. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.S.; Kang, K.H.; Kim, S.; Lee, S.; Lee, J.-H.; Kim, J.W.; Byun, B.; Meadows, G.G.; Joe, C.O. Amino acid-dependent NPRL2 interaction with Raptor determines mTOR Complex 1 activation. Cell Signal. 2016, 28, 32–41. [Google Scholar] [CrossRef]
- Abbasi, A.; Moghadam, A.A.; Kahrarian, Z.; Abbsavaran, R.; Yari, K.; Alizadeh, E. Molecular effects of leptin on peroxisome proliferator activated receptor gamma (PPAR-γ) mRNA expression in rat’s adipose and liver tissue. Cell. Mol. Biol. 2017, 63, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Yang, R.; Munir, K.M.; Horenstein, R.B.; Shuldiner, A.R. New progress in adipocytokine research. Curr. Opin. Endocrinol. 2003, 10, 115–121. [Google Scholar] [CrossRef]
- Aquila, H.; Link, T.A.; Klingenberg, M. The uncoupling protein from brown fat mitochondria is related to the mitochondrial ADP/ATP carrier. Analysis of sequence homologies and of folding of the protein in the membrane. EMBO J. 1985, 4, 2369–2376. [Google Scholar] [CrossRef]
- Song, S.; Hooiveld, G.J.; Zhang, W.; Li, M.; Zhao, F.; Zhu, J.; Xu, X.; Muller, M.; Li, C.; Zhou, G. Comparative Proteomics Provides Insights into Metabolic Responses in Rat Liver to Isolated Soy and Meat Proteins. J. Proteome Res. 2016, 15, 1135–1142. [Google Scholar] [CrossRef]
- Carling, D.; Mayer, F.V.; Sanders, M.J.; Gamblin, S.J. AMP-activated protein kinase: Nature’s energy sensor. Nat. Chem. Biol. 2011, 7, 512–518. [Google Scholar] [CrossRef]
- Zhang, B.B.; Zhou, G.; Li, C. AMPK: An Emerging Drug Target for Diabetes and the Metabolic Syndrome. Cell Metab. 2009, 9, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Foretz, M.; Even, P.; Andreelli, F.; Kahn, A.; Vaulont, S.; Viollet, B. Regulation of the oxydation of the fatty acids by the AMP-activated protein kinase (AMPK) in the liver during fast. Diabetes Metab. 2006, 32, S14. [Google Scholar]
- Yuan, E.; Duan, X.; Xiang, L.; Ren, J.; Lai, X.; Li, Q.; Sun, L.; Sun, S. Aged Oolong Tea Reduces High-Fat Diet-Induced Fat Accumulation and Dyslipidemia by Regulating the AMPK/ACC Signaling Pathway. Nutrients 2018, 10, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, F.; Fan, W.; Guang, C.; Hui, D.; Jingbin, L.; Yan, Z.; Lijun, X.; Xin, Z.; Fuer, L. Diosgenin ameliorates palmitic acid-induced lipid accumulation via AMPK/ACC/CPT-1A and SREBP-1c/FAS signaling pathways in LO2 cells. BMC Complement. Altern. Med. 2019, 19, 255. [Google Scholar]
- Bugge, A.; Siersbæk, M.; Madsen, M.S.; Göndör, A.; Rougier, C.; Mandrup, S. A novel intronic peroxisome proliferator-activated receptor gamma enhancer in the uncoupling protein (UCP) 3 gene as a regulator of both UCP2 and -3 expression in adipocytes. J. Biol. Chem. 2010, 285, 17310–17317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, N.; Yuan, K.; Chen, G.; Sun, J.; Chen, H. Research Progress in High Protein Diet Regulating Gut Microbes to Influence Obesity. Med. Recapitul. 2019, 25, 1960–1964. [Google Scholar]
- Pia, K.; Secher, M.L.; Even, F.; Qin, H.; Floor, H.; Brask, S.S.; Muriel, D.; Møller, P.L.; Koefoed, P.R.; Alicja, M.; et al. Effect of a long-term high-protein diet on survival, obesity development, and gut microbiota in mice. Am. J. Physiol.-Endocrinol. Metab. 2016, 310, E886–E899. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NC | HF | NR | HSP | HPP | ||||||
| Ingredient | gm | kcal | gm | kcal | gm | kcal | gm | kcal | gm | kcal |
| Pork protein | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 368.42 | 1400 |
| Soy protein | 0 | 0 | 0 | 0 | 0 | 0 | 388.89 | 1400 | 0 | 0 |
| Casein | 200 | 800 | 200 | 800 | 200 | 800 | 0 | 0 | 0 | 0 |
| L-cystine | 3 | 12 | 3 | 12 | 3 | 12 | 3 | 12 | 3 | 12 |
| Corn starch | 506.2 | 2024.8 | 0 | 0 | 506.2 | 2024.8 | 304.61 | 1218.44 | 325.08 | 1300.32 |
| Maltodextrin | 125 | 500 | 125 | 500 | 125 | 500 | 116 | 464 | 116 | 464 |
| Sucrose | 68.8 | 275.2 | 68.8 | 275 | 68.8 | 275.2 | 50 | 200 | 50 | 200 |
| Cellulose | 50 | 0 | 50 | 0 | 50 | 0 | 50 | 0 | 50 | 0 |
| Soybean oil | 25 | 225 | 25 | 225 | 25 | 225 | 40 | 360 | 40 | 360 |
| Lard | 20 | 180 | 245 | 2205 | 20 | 180 | 0 | 0 | 0 | 0 |
| Vitamin mix V10037 | 10 | 40 | 10 | 40 | 10 | 40 | 10 | 40 | 10 | 40 |
| Mineral mix S10022G | 35 | 0 | 35 | 0 | 35 | 0 | 35 | 0 | 35 | 0 |
| Line bitartrate | 2.5 | 0 | 2.5 | 0 | 2.5 | 0 | 2.5 | 0 | 2.5 | 0 |
| Total | 1045.5 | 4057 | 764.3 | 4057 | 1045.5 | 4057 | 1000 | 3694.44 | 1000 | 3776.32 |
| Gene | Upstream Primer (5′-3′) | Downstream Primer (3′-5′) |
|---|---|---|
| PPAR-γ | TCGCTGATGCACTGCCTATG | GAGAGGTCCACAGAGCTGATT |
| UCP2 | ATGGTTGGTTTCAAGGCCACA | CGGTATCCAGAGGGAAAGTGAT |
| β-actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Ji, S.; Tang, X.; Wang, T.; Wang, H.; Meng, X. A Comparison Study on the Therapeutic Effect of High Protein Diets Based on Pork Protein versus Soybean Protein on Obese Mice. Foods 2022, 11, 1227. https://doi.org/10.3390/foods11091227
Jiang S, Ji S, Tang X, Wang T, Wang H, Meng X. A Comparison Study on the Therapeutic Effect of High Protein Diets Based on Pork Protein versus Soybean Protein on Obese Mice. Foods. 2022; 11(9):1227. https://doi.org/10.3390/foods11091227
Chicago/Turabian StyleJiang, Songsong, Shanshan Ji, Xinlei Tang, Tao Wang, Hengpeng Wang, and Xiangren Meng. 2022. "A Comparison Study on the Therapeutic Effect of High Protein Diets Based on Pork Protein versus Soybean Protein on Obese Mice" Foods 11, no. 9: 1227. https://doi.org/10.3390/foods11091227
APA StyleJiang, S., Ji, S., Tang, X., Wang, T., Wang, H., & Meng, X. (2022). A Comparison Study on the Therapeutic Effect of High Protein Diets Based on Pork Protein versus Soybean Protein on Obese Mice. Foods, 11(9), 1227. https://doi.org/10.3390/foods11091227