
Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Preparation of Fermented Sausage Samples
2.3. Lipid Composition Analysis
2.3.1. Lipid Extraction
2.3.2. Lipid Separation
2.4. Lipid Oxidation Analysis
2.4.1. Lipoxygenase (LOX) Activity Determination
2.4.2. Peroxide Value Determination
2.4.3. Thiobarbituric Acid Reactive Substances (TBARS) Determination
2.5. Bacterial Analysis
2.5.1. Bacterial Counting
2.5.2. High-Throughput Sequencing of Bacteria
2.6. Flavor Compound Determination
2.7. Statistics Analysis
3. Results and Discussion
3.1. Lipid Composition
3.2. pH
3.3. POV, TBAR Value, and LOX Activity
3.4. Bacterial Counting
3.5. Bacterial Community
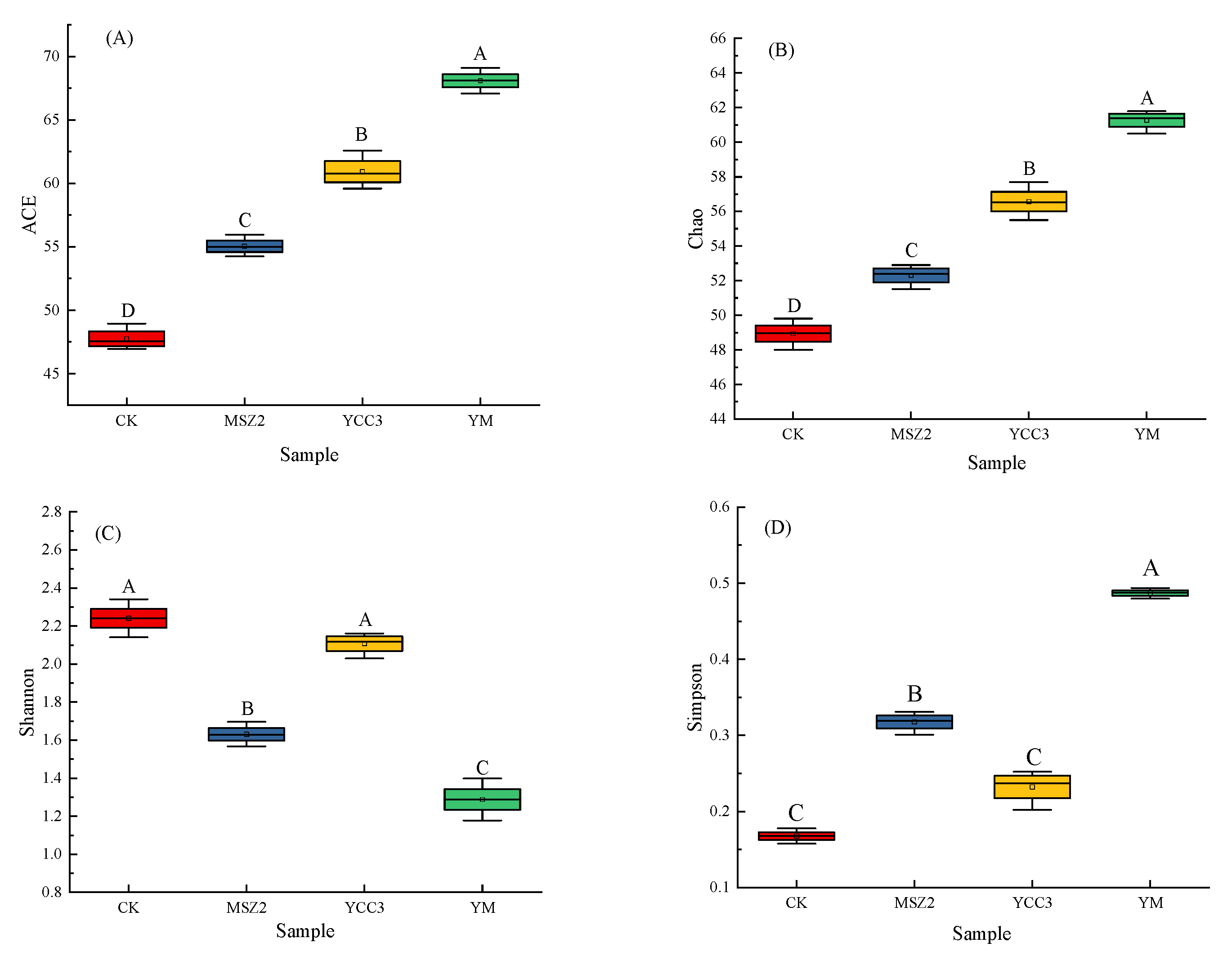
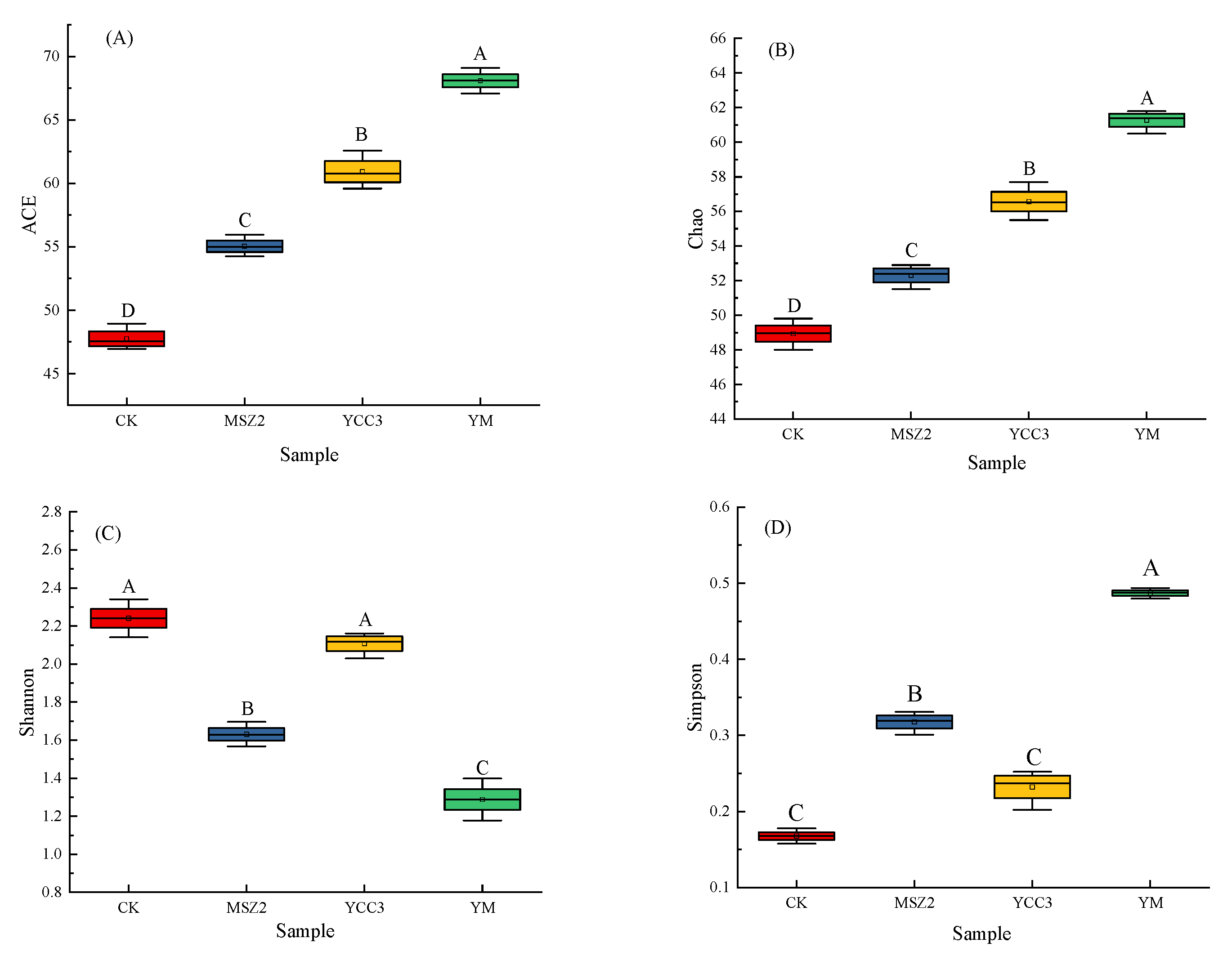
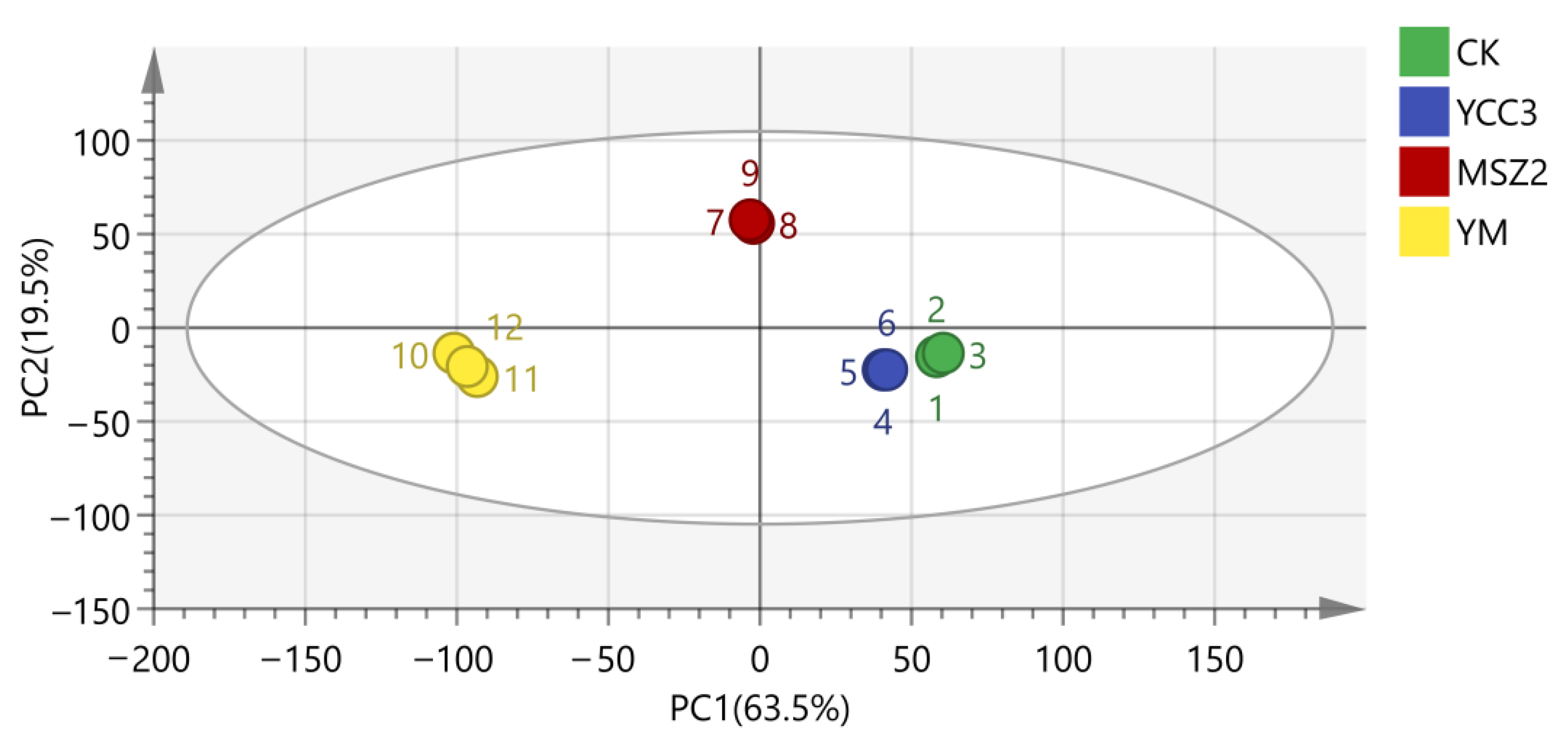
3.5.1. Bacterial Richness and Diversity
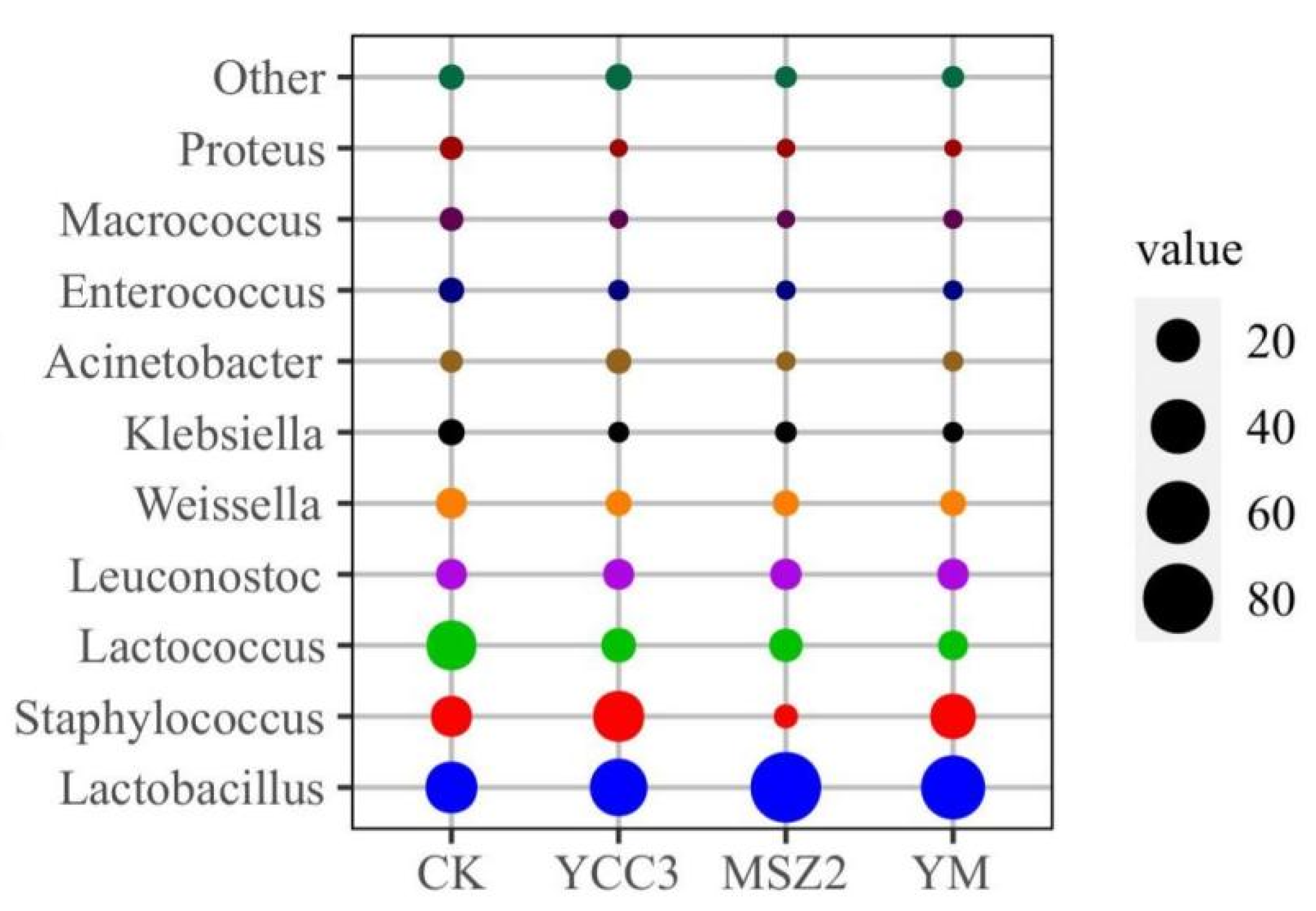
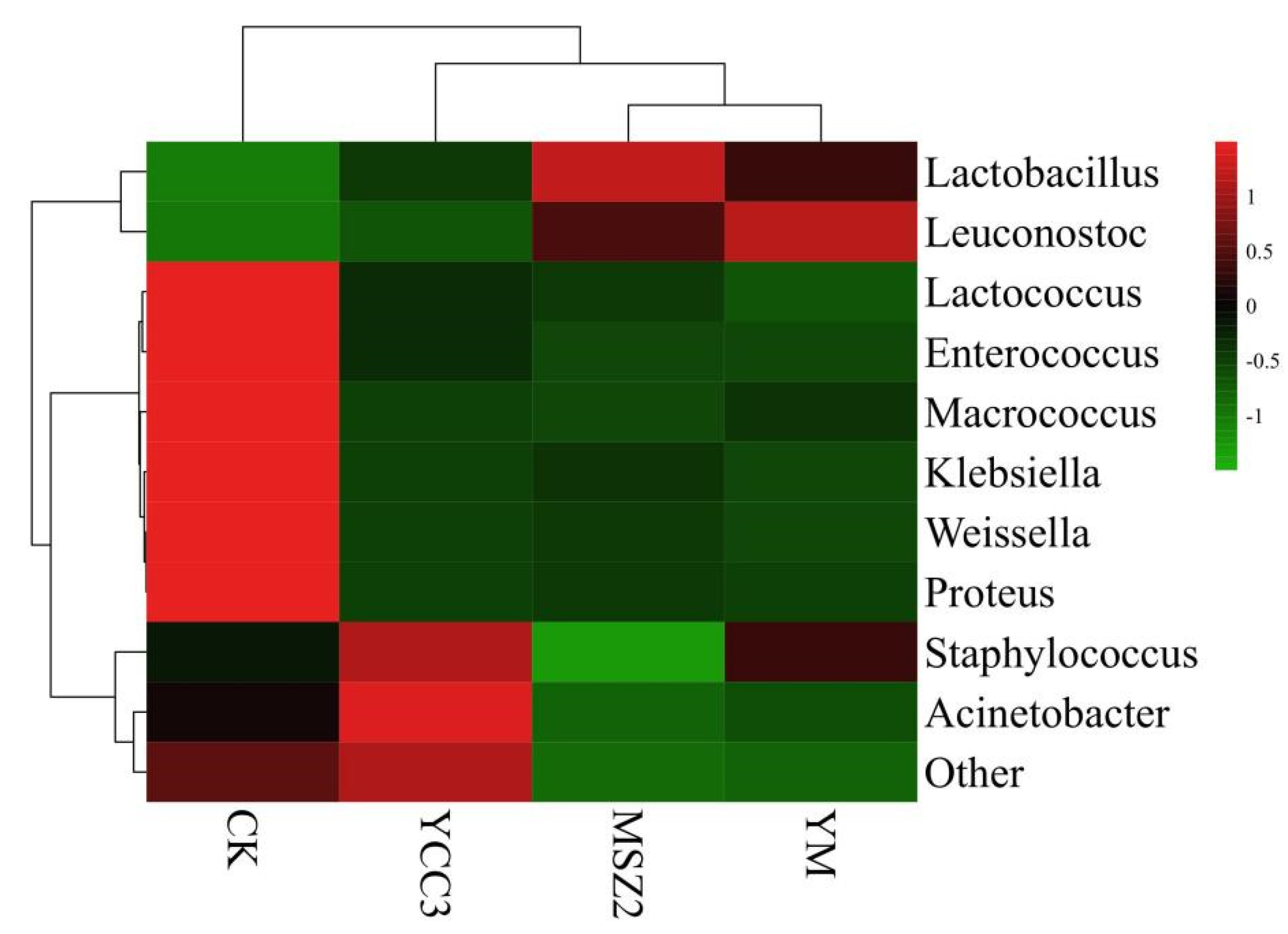
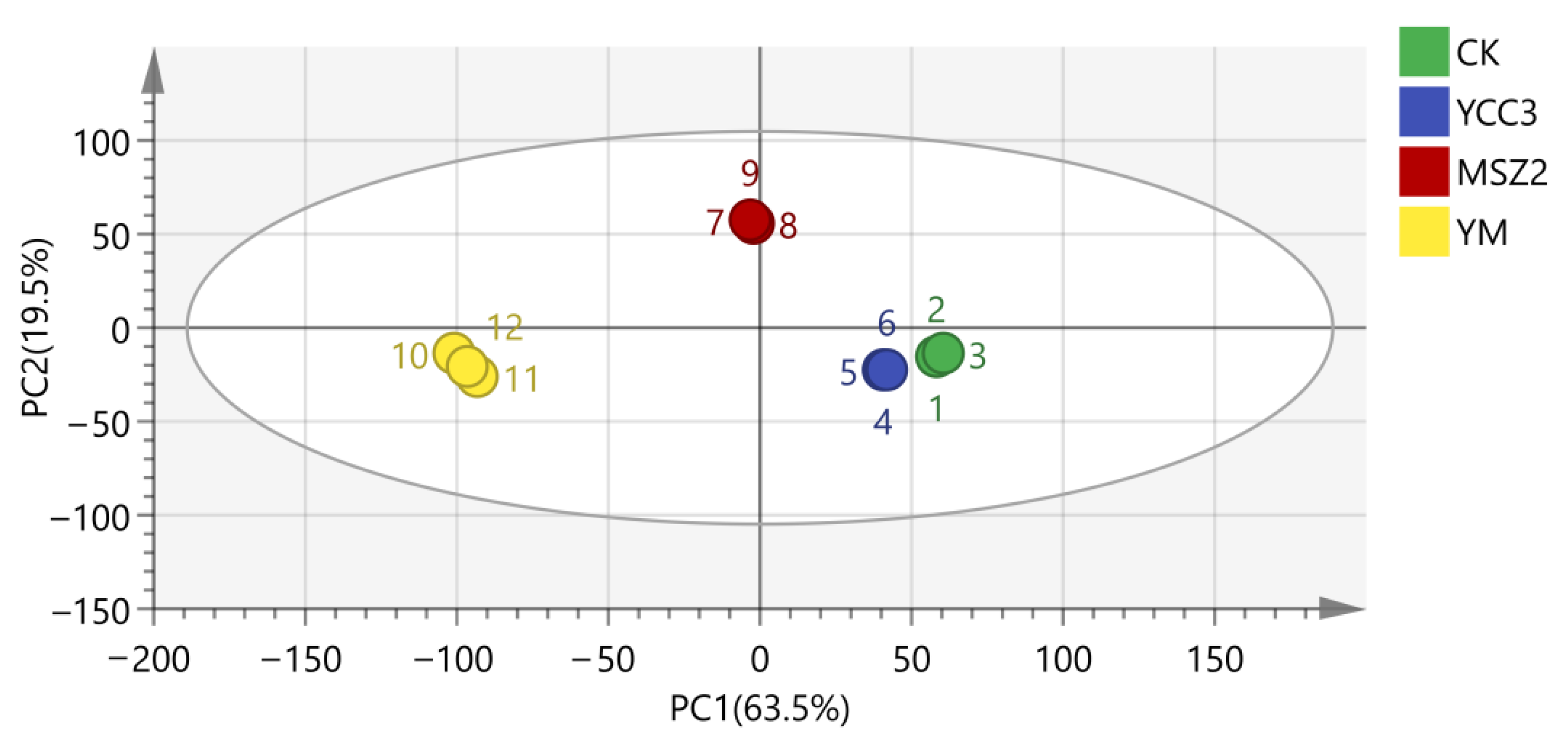
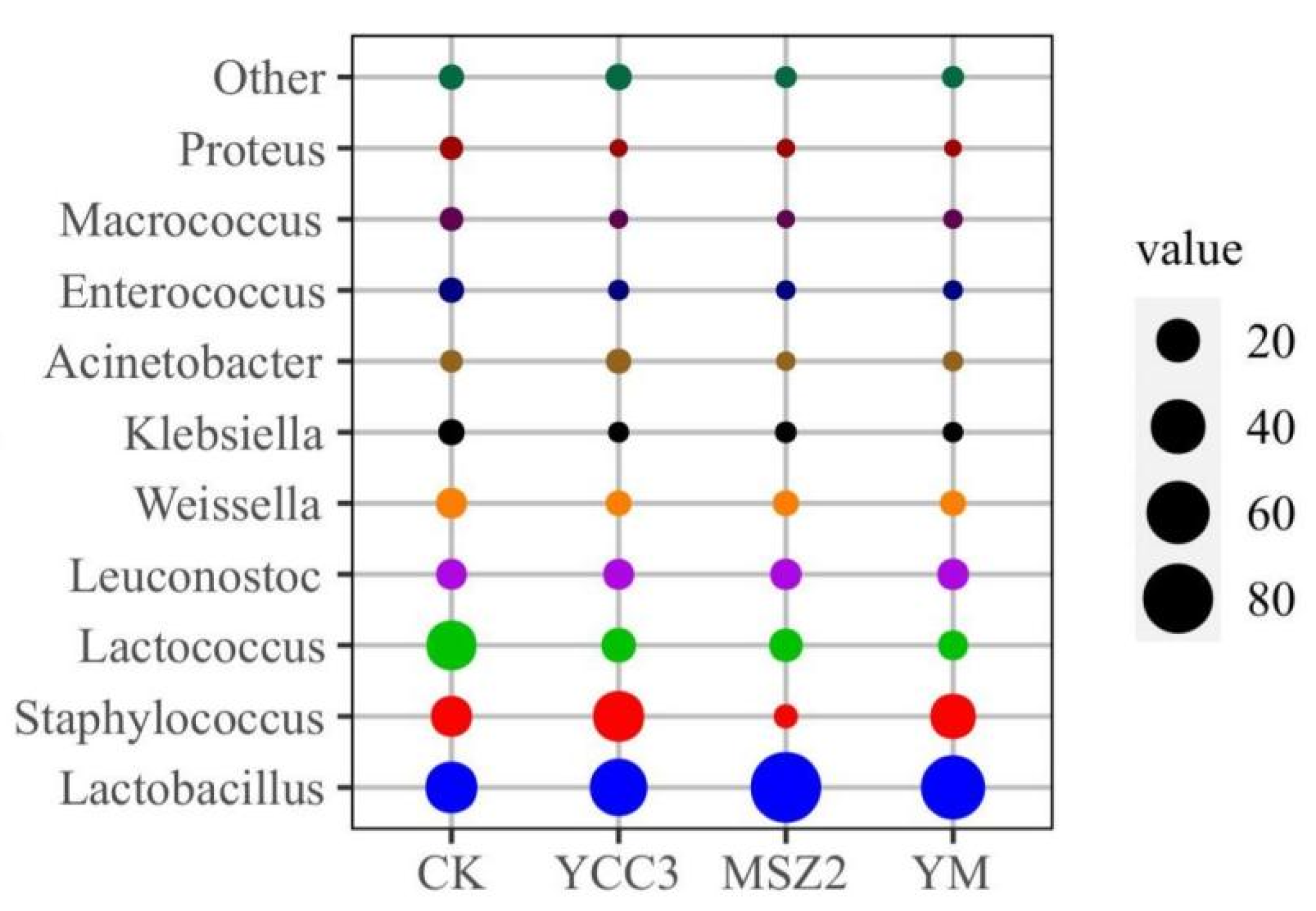
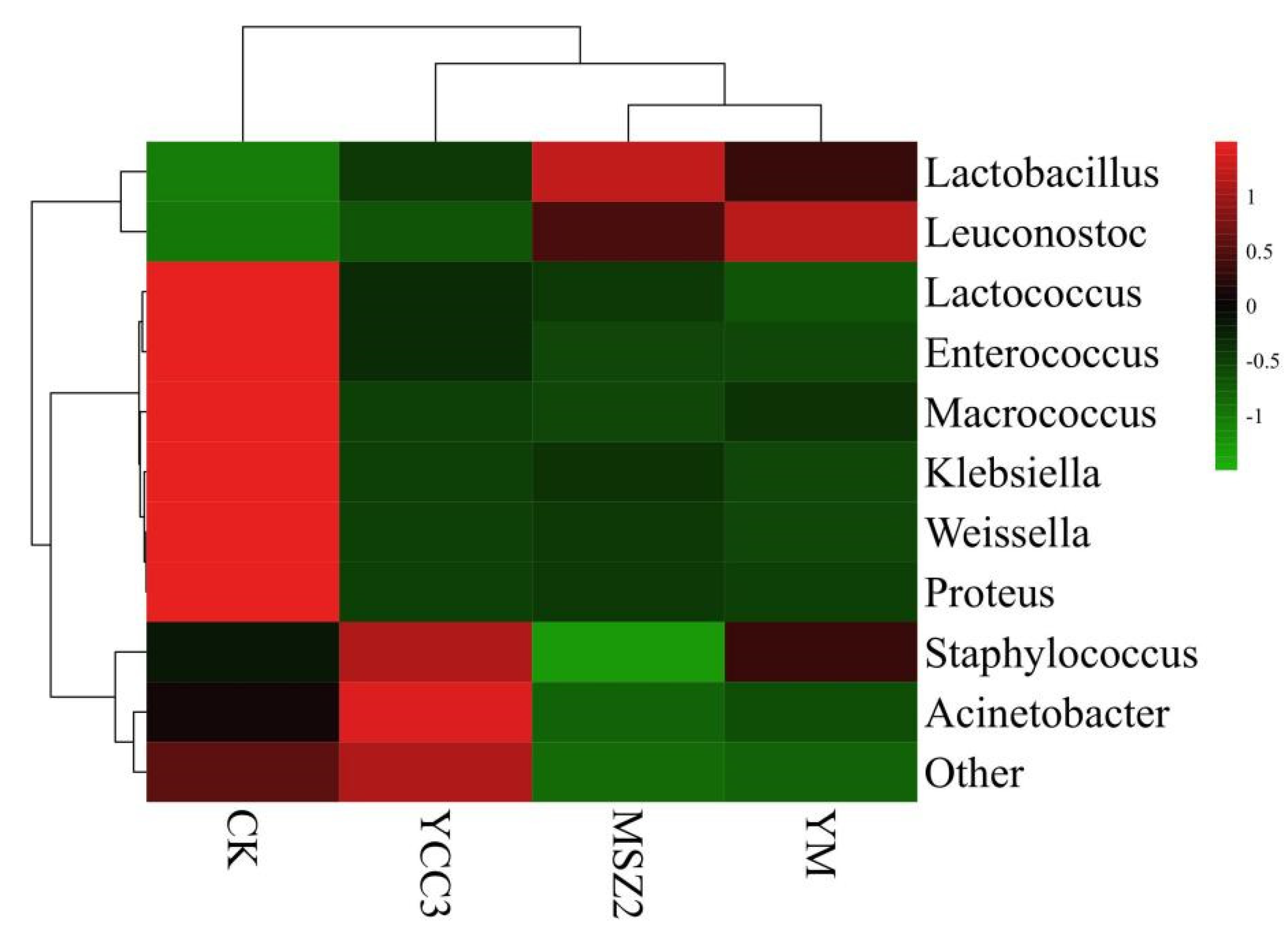
3.5.2. Bacterial Composition
3.6. Volatile Compounds
3.6.1. Volatile Compound Composition
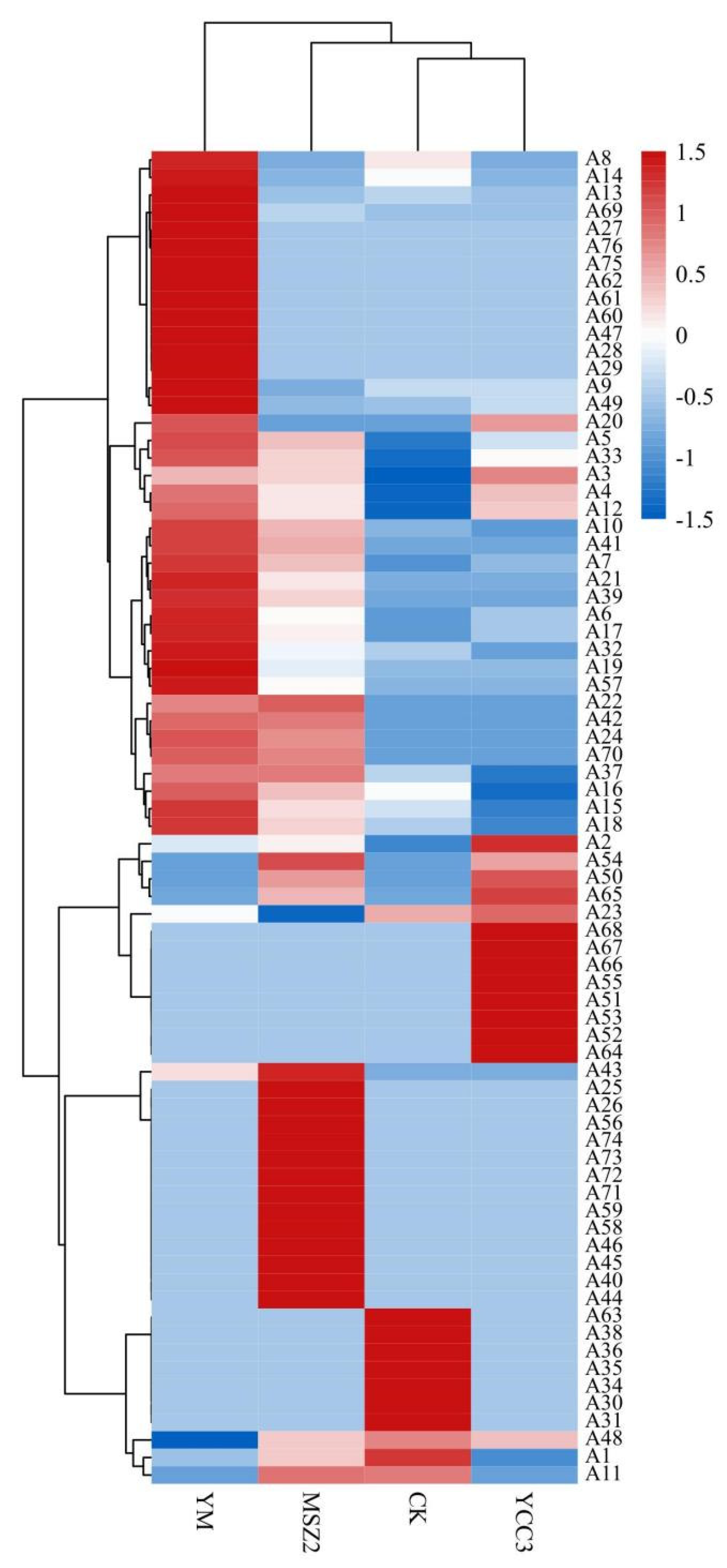
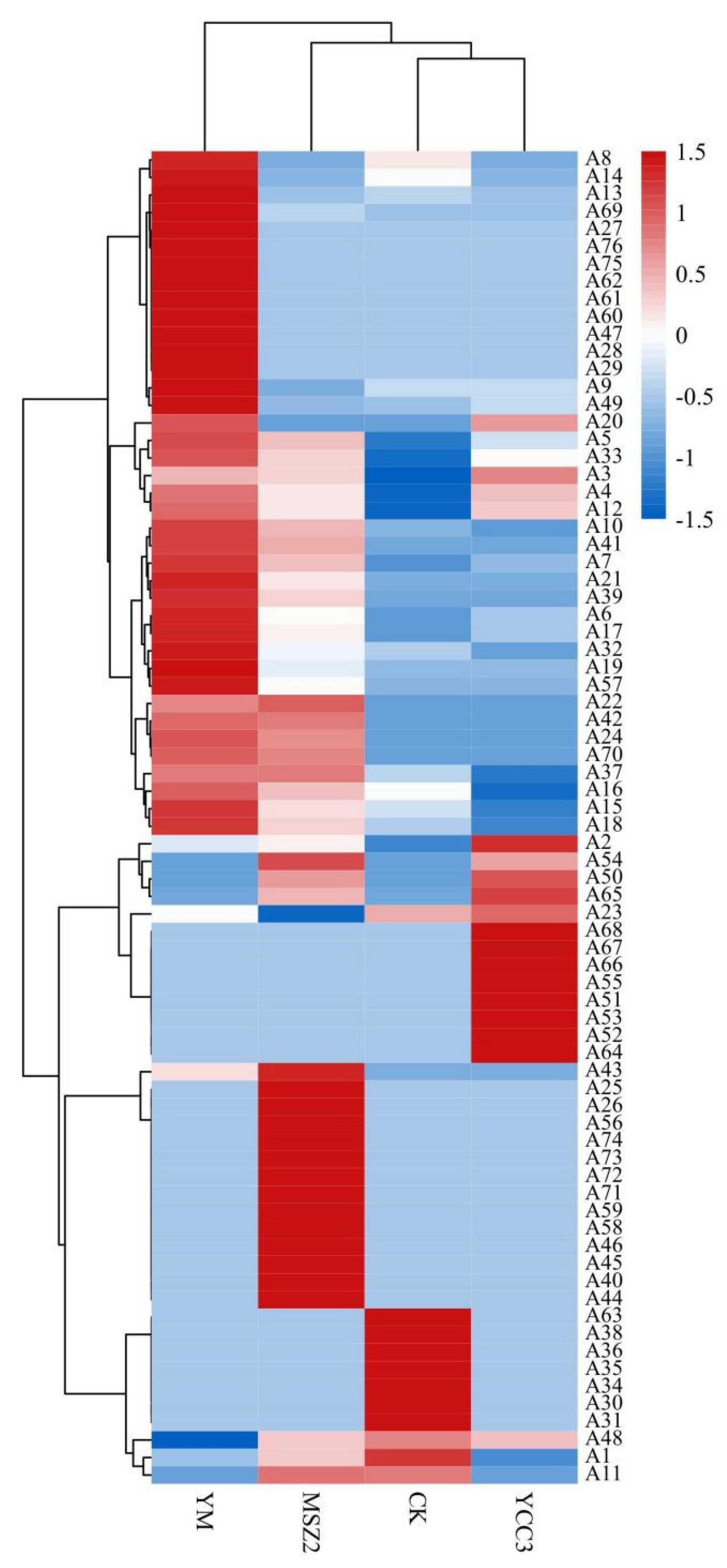
3.6.2. Difference in the Volatile Composition
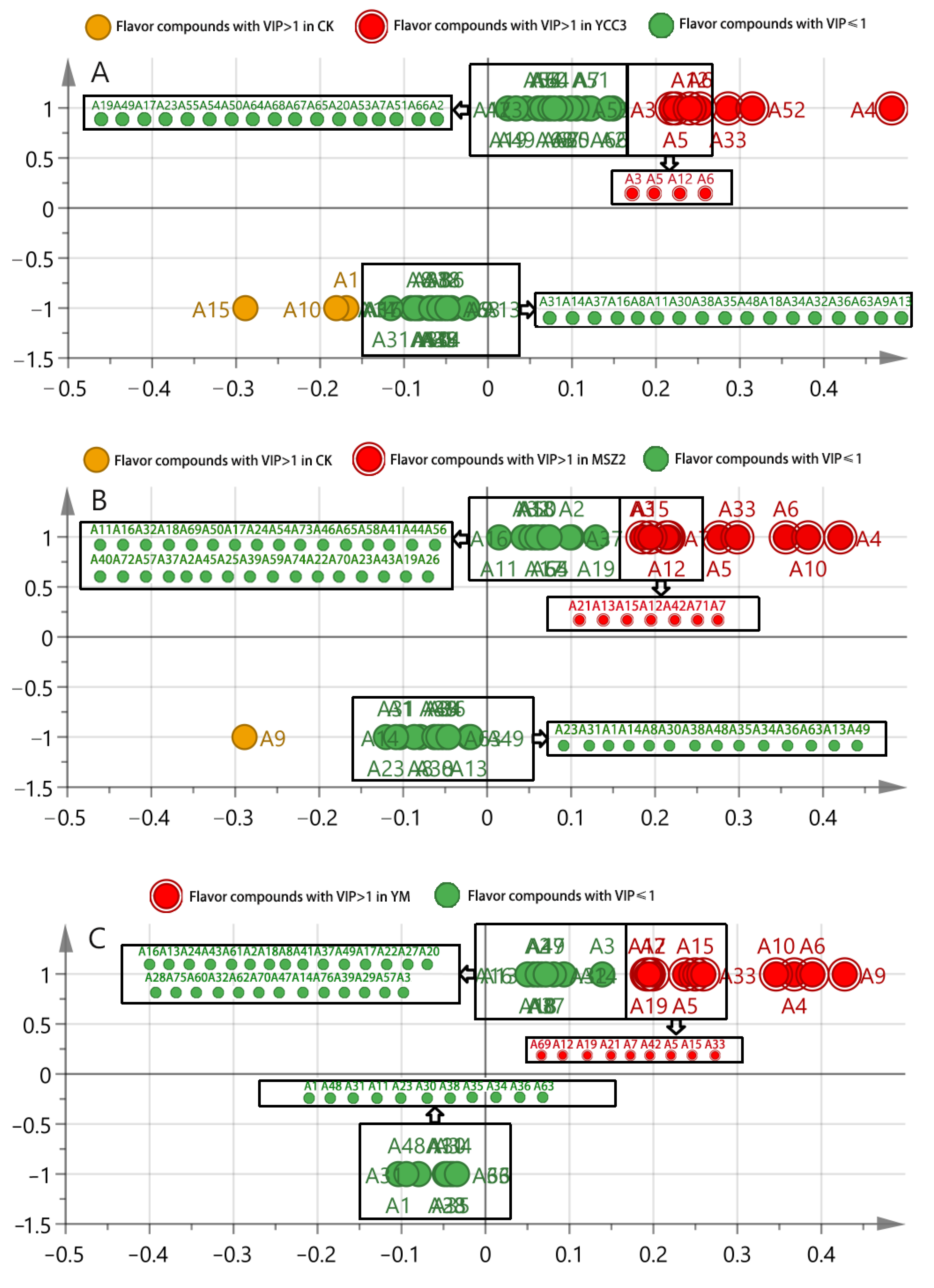
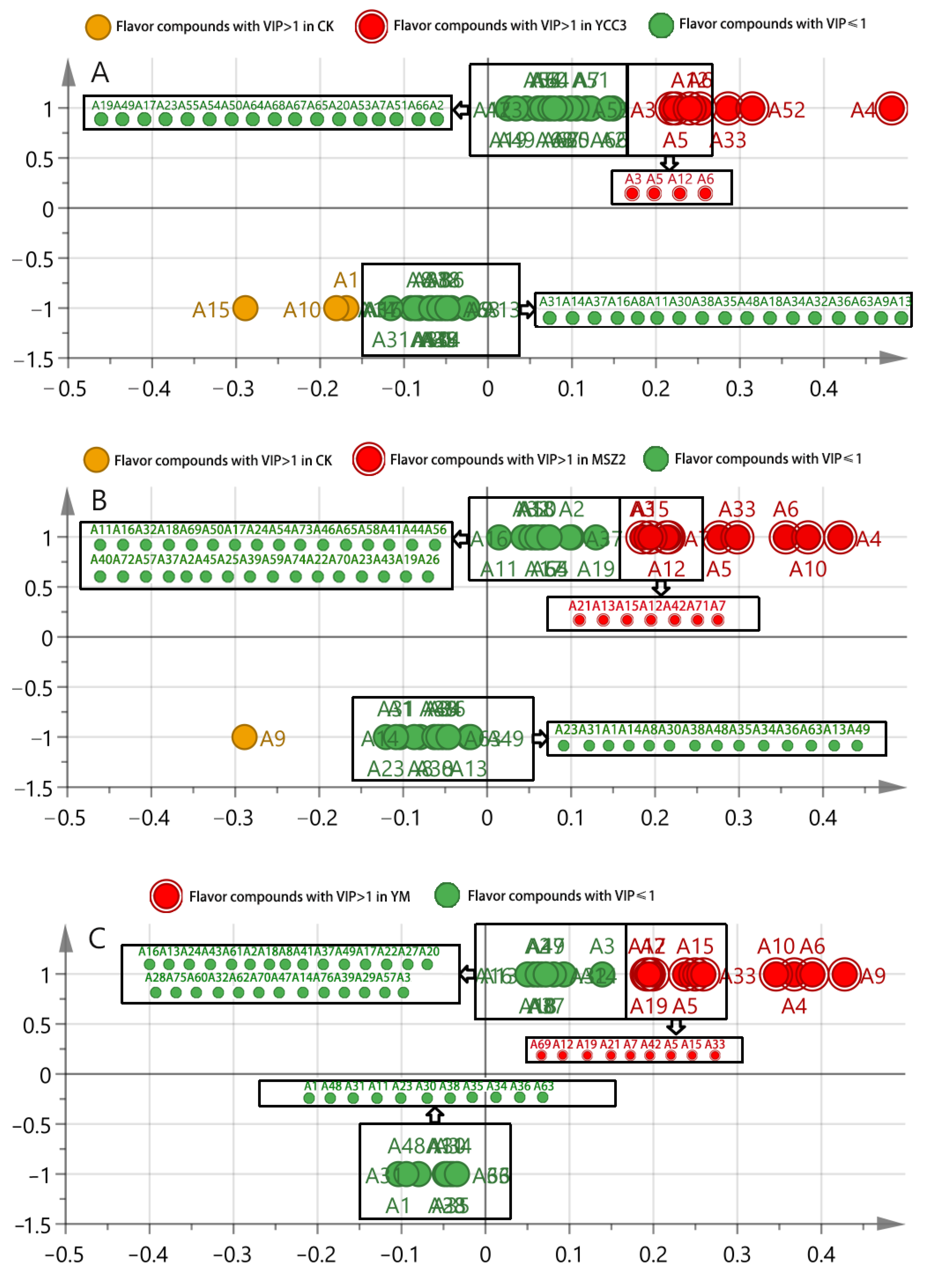
3.6.3. Identification of Differential Flavor Compounds from Fermented Sausages
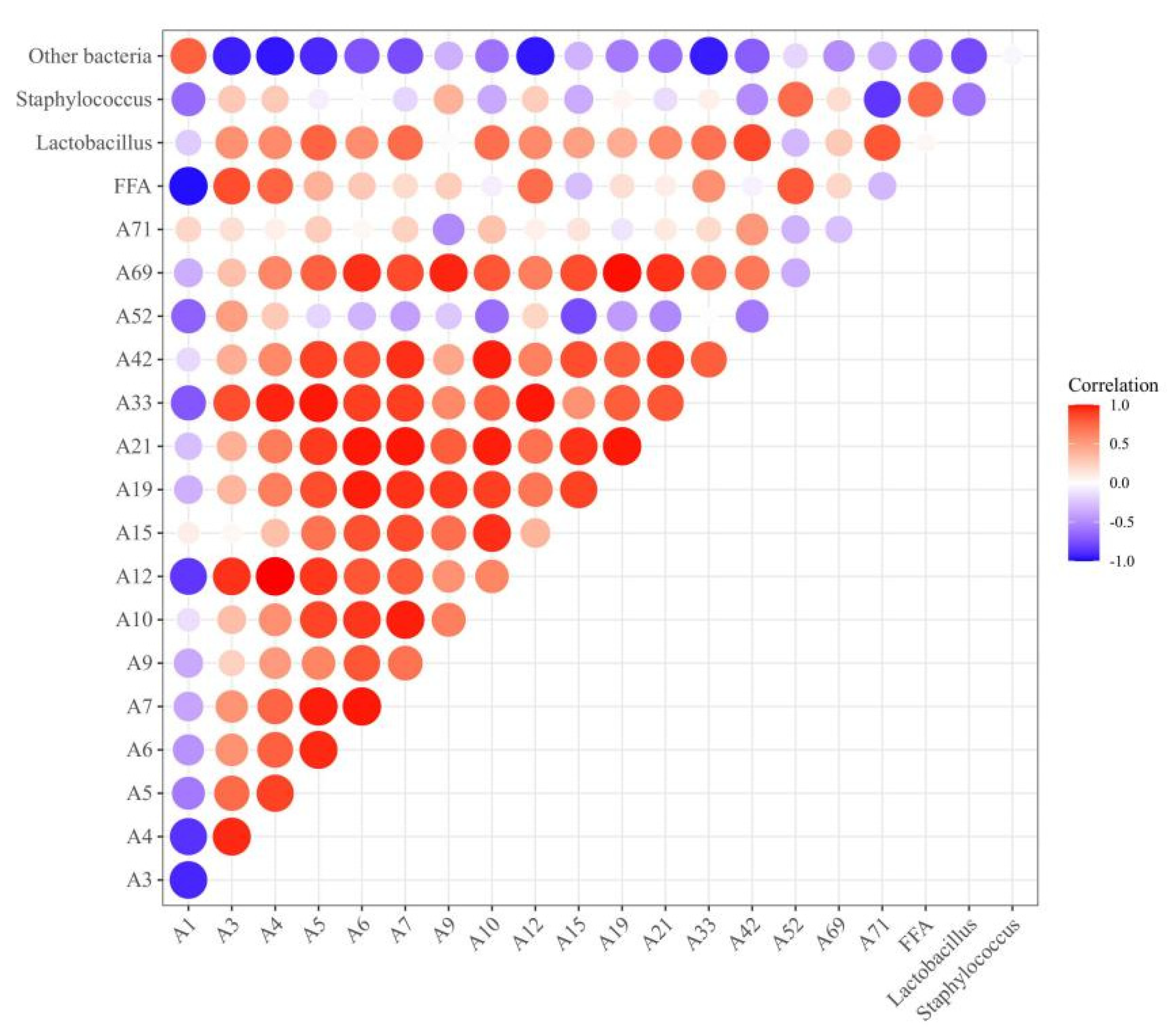
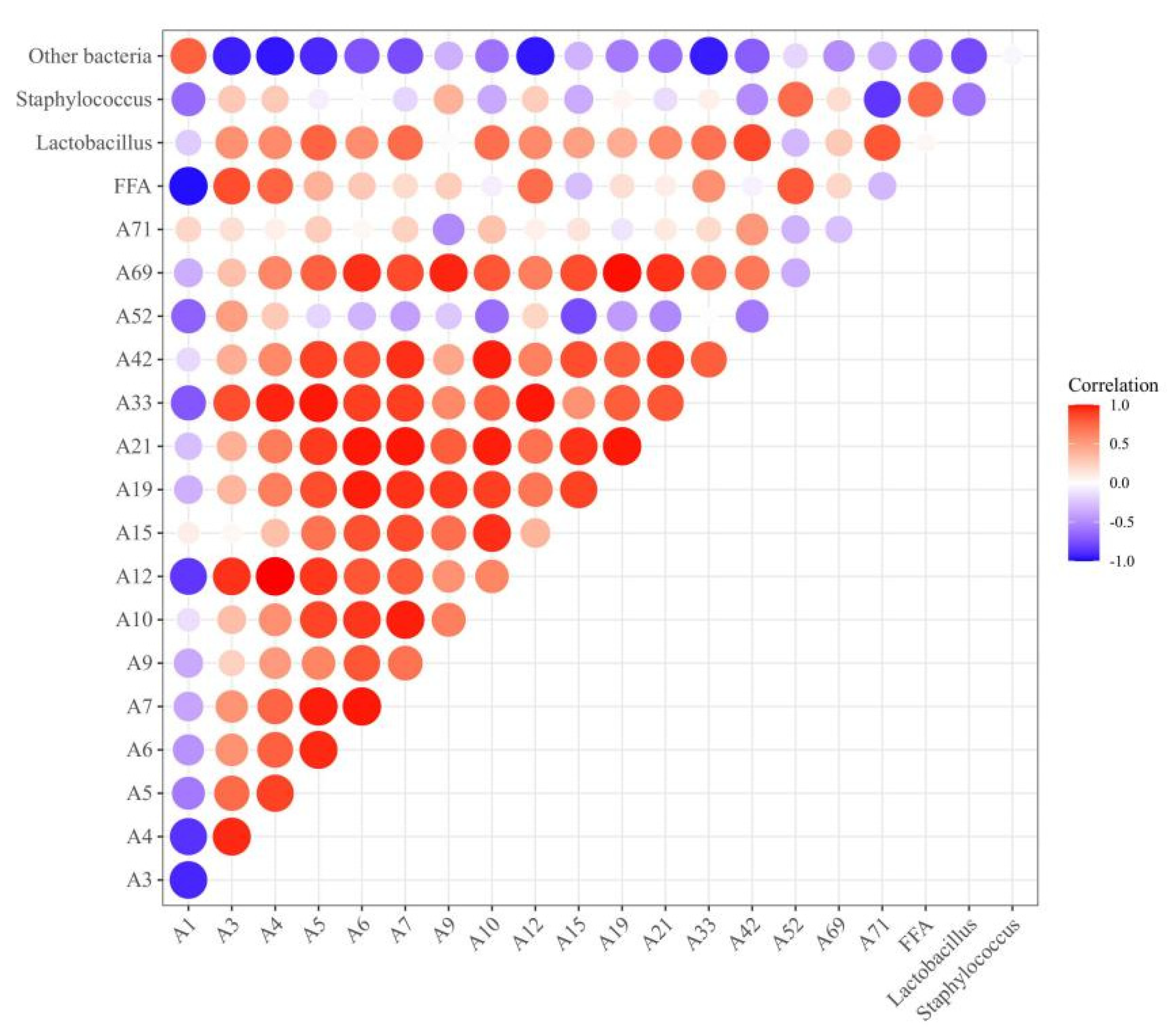
3.7. Correlation Analysis of Bacterial Relative Abundance, FFA Content, and Differential Flavor Compounds
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarter, S.; Ho, P.H.; To, K. Current context of food safety in Vietnam: A glance at food of animal origin. Qual. Assur. Saf. Crop. 2015, 7, 57–62. [Google Scholar] [CrossRef]
- Bao, R.; Liu, S.; Ji, C.; Liang, H.; Yang, S.; Yan, X.; Zhou, Y.; Lin, X.; Zhu, B. Shortening fermentation period and quality improvement of fermented fish, Chouguiyu, by co-inoculation of lactococcus lactis, M10 and Weissella cibariaM3. Front. Microbiol. 2018, 9, 3003. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Gómez, M.; Fonseca, S. Effect of commercial starter cultures on physicochemical characteristics, microbial counts and free fatty acid composition of dry-cured foal sausage. Food Control 2014, 46, 382–389. [Google Scholar] [CrossRef]
- Mejri, L.; Ziadi, A.; Adab, S.E.; Boulares, M.; Essid, I.; Hassouna, M. Effect of commercial starter cultures on physicochemical, microbiological and textural characteristics of a traditional dry fermented sausage reformulated with camel meat and hump fat. J. Food Meas. Charact. 2016, 2, 758–767. [Google Scholar] [CrossRef]
- Ammor, M.S.; Mayo, B. Selection criteria for lactic acid bacteria to be used as functional starter cultures in dry sausage production: An update. Meat Sci. 2007, 76, 138–146. [Google Scholar] [CrossRef]
- Wang, Y.; Li, F.; Chen, J.; Sun, Z.; Fu, L. High-throughput sequencing-based characterization of the predominant microbial community associated with characteristic flavor formation in Jinhua Ham. Food Microbiol. 2021, 94, 103643. [Google Scholar] [CrossRef]
- Cruxen, C.E.D.S.; Funck, G.D.; Dannenberg, G.D.S.; Haubert, L.; Marques, J.D.L.; Kroning, I.S.; Chaves, F.C.; Da Silva, W.P.; Fiorentini, Â.M. Characterization of Staphylococcus xylosus LQ3 and its application in dried cured sausage. LWT-Food Sci. Technol. 2017, 86, 538–543. [Google Scholar] [CrossRef]
- Li, P.; Luo, H.; Kong, B.; Liu, Q.; Chen, C. Formation of red myoglobin derivatives and inhibition of spoilage bacteria in raw meat batters by lactic acid bacteria and Staphylococcus xylosus. LWT-Food Sci. Technol. 2016, 68, 251–257. [Google Scholar] [CrossRef]
- Ge, Q.; Pei, H.; Liu, R.; Chen, L.; Gao, X.; Gu, Y.; Hou, Q.; Yin, Y.; Yu, H.; Wu, M.; et al. Effects of Lactobacillus plantarum NJAU-01 from Jinhua ham on the quality of dry-cured fermented sausage. LWT-Food Sci. Technol. 2019, 101, 513–518. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.; Han, Q.; Xia, X.; Xu, L. The role of bacterial fermentation in lipolysis and lipid oxidation in Harbin dry sausages and its flavor development. LWT-Food Sci. Technol. 2017, 77, 389–396. [Google Scholar] [CrossRef]
- Toldra, F. Proteolysis and lipolysis in flavor development of dry-cured meat products. Meat Sci. 1998, 49, S101–S110. [Google Scholar] [CrossRef]
- Huang, Y.; Li, H.; Huang, T.; Li, F.; Sun, J. Lipolysis and lipid oxidation during processing of Chinese traditional smoke-cured bacon. Food Chem. 2014, 149, 31–39. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Ji, L.; Zhang, J.; Chen, L. A Review: Microbial Diversity and Function of Fermented Meat Products in China. Front. Microbiol. 2021, 12, 645435. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, Y.; Chen, C.; Xie, T.; Li, P. Effect of Lactobacillus plantarum and Staphylococcus xylosus on flavor development and bacterial communities in Chinese dry fermented sausages. Food Res. Int. 2020, 135, 109247. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Toldrá, F. Microbial enzymatic activities for improved fermented meats. Trends Food Sci. Technol. 2011, 22, 81–90. [Google Scholar] [CrossRef]
- Navarro, J.; Nadal, M.; Izquierdo, L.; Flores, J. Lipolysis in dry cured sausages as affected by processing conditions. Meat Sci. 1997, 45, 161–168. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Gómez, M.; Purriños, L.; Fonseca, S. Effect of commercial starter cultures on volatile compound profile and sensory characteristics of dry-cured foal sausage. J. Sci. Food Agric. 2016, 96, 1194–1201. [Google Scholar] [CrossRef]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipidprotein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef]
- Flores, M.; Dura, M.A.; Marco, A.; Toldra, F. Effect of Debaryomyces spp. on aroma formation and sensory quality of dry-fermented sausages. Meat Sci. 2004, 68, 439–446. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.; Sun, Q.; Dong, F.; Liu, Q. Antioxidant potential of a unique LAB culture isolated from Harbin dry sausage: In vitro and in a sausage model. Meat Sci. 2015, 110, 180–188. [Google Scholar] [CrossRef]
- Bozkurt, H.; Erkmen, O. Effects of starter cultures and additives on the quality of Turkish style sausage (sucuk). Meat Sci. 2002, 61, 149–156. [Google Scholar] [CrossRef]
- Li, Y.; Yu, Z.; Zhu, Y.; Cao, Z. Selection of nitrite-degrading and biogenic amine-degrading strains and its involved genes. Food Qual. Saf. 2020, 4, 225–235. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Jiang, M.; Rui, X.; Li, W.; Chen, X.H.; Dong, M.S. Effect of rose polyphenols on oxidation, biogenic amines and microbial diversity in naturally dry fermented sausages. Food Control 2017, 78, 324–330. [Google Scholar] [CrossRef]
- Folch, J. A simple method for the isolation and purification of total lipides from animal tissues. J. Bid. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Kaluzny, M.A.; Duncan, L.A. Rapid separation of lipid classes in high yield and purity using bonded phase columns. J. Lipid Res. 1985, 26, 135–140. [Google Scholar] [CrossRef]
- Xu, Y.; Li, L.; Regenstein, J.M.; Gao, P.; Zang, J.; Xia, W.; Jiang, Q. The contribution of autochthonous microflora on free fatty acids release and flavor development in low-salt fermented fish. Food Chem. 2018, 256, 259–267. [Google Scholar] [CrossRef]
- Li, F.F.; Zhong, Q.; Kong, B.H.; Wang, B.; Pan, N.; Xia, X.F. Deterioration in quality of quick-frozen pork patties induced by changes in protein structure and lipid and protein oxidation during frozen storage. Food Res. Int. 2020, 133, 109142. [Google Scholar] [CrossRef]
- Lu, S.; Ji, H.; Wang, Q.; Li, B.; Li, K.; Xu, C.; Jiang, C. The effects of starter cultures and plant extracts on the biogenic amine accumulation in traditional Chinese smoked horsemeat sausages. Food Control 2015, 50, 869–875. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, F.; Shi, X.; Wang, B.; Li, K.; Li, B.; Bin, Z. Dynamic correlations between microbiota succession and flavor development involved in the ripening of Kazak artisanal cheese. Food Res. Int. 2018, 105, 733–742. [Google Scholar] [CrossRef]
- De Lima Alves, L.; Donadel, J.Z.; Athayde, D.R.; Da Silva, M.S.; Klein, B.; Fagundes, M.B.; Cichoski, A.J. Effect of ultrasound on proteolysis and the formation of volatile compounds in dry fermented sausages. Ultrason. Sonochem. 2020, 67, 105161. [Google Scholar] [CrossRef]
- Iacumin, L.; Cocolin, L.; Cantoni, C.; Comi, G. Preliminary analysis of the lipase gene (gehM) expression of Staphylococcus xylosus in vitro and during fermentation of naturally fermented sausages. J. Food Protect. 2007, 70, 2665. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Metaxopoulos, J.; Vlassi, M.; Pappa, A. Stability and safety of traditional Greek salami—A microbiological ecology study. Int. J. Food Microbiol. 1998, 44, 69–82. [Google Scholar] [CrossRef]
- Wang, J.; Lu, S.; Wang, Q.; Guo, X.; He, X. Effects of starter cultures on lipid oxidation and accumulation of biogenic amines in traditional Chinese smoked horsemeat sausage. J. Food Process. Pres. 2020, 44, 14439. [Google Scholar] [CrossRef]
- Barrière, C.; Centeno, D.; Lebert, A.; Leroy-Sétrin, S.; Berdagué, J.L.; Talon, R. Roles of superoxide dismutase and catalase of Staphylococcus xylosus in the inhibition of linoleic acid oxidation. FEMS Microbiol. Lett. 2001, 201, 181–185. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, W.; Ge, C. Characterization of fermented silver carp sausages inoculated with mixed starter culture. LWT-Food Sci. Technol. 2008, 41, 730–738. [Google Scholar] [CrossRef]
- Mejri, L.; Vasquez-Villanueva, R.; Hassouna, M.; Marina, L.M.; Marina, C.G. Identification of peptides with antioxidant and antihypertensive capacities by RP-HPLC-Q-TOF-MS in dry fermented camel sausages inoculated with different starter cultures and ripening times. Food Res. Int. 2017, 100, 708–716. [Google Scholar] [CrossRef]
- Fu, X.; Xu, S.; Wang, Z. Kinetics of lipid oxidation and off-odor formation in silver carp mince: The effect of lipoxygenase and hemoglobin. Food Res. Int. 2009, 42, 85–90. [Google Scholar] [CrossRef]
- Gata, J.L.; Pinto, M.C.; Macías, P. Lipoxygenase activity in pig muscle: Purification and partial characterization. J. Agric. Food chem. 1996, 44, 2573–2577. [Google Scholar] [CrossRef]
- Spaziani, M.; Del Torre, M.; Stecchini, M.L. Changes of physicochemical, microbiological, and textural properties during ripening of Italian low-acid sausages. Proteolysis, sensory and volatile profiles. Meat Sci. 2009, 81, 77–85. [Google Scholar] [CrossRef]
- Essid, I.; Hassouna, M. Effect of inoculation of selected Staphylococcus xylosus and Lactobacillus plantarum strains on biochemical, microbiological and textural characteristics of a Tunisian dry fermented sausage. Food Control 2013, 32, 707–714. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Ren, H.; Zhan, Y. Comparison of bacterial diversity profiles and microbial safety assessment of salami, Chinese dry-cured sausage and Chinese smoked-cured sausage by high-throughput sequencing. LWT-Food Sci. Technol. 2018, 90, 108–115. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Liu, Q.; Wang, Y.; Chen, Q.; Kong, B. The potential correlation between bacterial diversity and the characteristic volatile flavor of traditional dry sausages from Northeast China. Food Microbiol. 2020, 91, 103505. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Tang, N.; Liu, R.; Gong, M.; Wang, Z.; Guo, Y.; Wang, Y.; Zhang, Y.; Chang, M. The relationship between flavor formation, lipid metabolism, and microorganisms in fermented fish products. Food Funct. 2021, 12, 5686. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, F.; Milanovi, V.; Osimani, A.; Aquilanti, L.; Taccari, M.; Garofalo, C.; Polverigiani, S.; Clementi, F.; Franciosi, E.; Tuohy, K. Microbial dynamics of model Fabriano-like fermented sausages as affected by starter cultures, nitrates and nitrites. Int. J. Food Microbiol. 2018, 278, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, J.; Pei, Z.; Wei, P.; Xiang, D.; Cao, X.; Shen, X.; Li, C. Volatile flavor components and the mechanisms underlying their production in golden pompano (Trachinotus blochii) fillets subjected to different drying methods: A comparative study using an electronic nose, an electronic tongue and SDE-GC-MS. Food Res. Int. 2019, 123, 217–225. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Li, C.; Li, L.; Yang, X.; Wu, Y.; Chen, S.; Zhao, Y. Novel insight into physicochemical and flavor formation in naturally fermented tilapia sausage based on microbial metabolic network. Food Res. Int. 2021, 141, 110122. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Z.; Sun, L.; Dong, L.; Du, M. Dynamics of Microbial Communities, Texture and flavor in Suan zuo yu during Fermentation. Food Chem. 2020, 332, 127364. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Zhao, Y.; Li, L.; Yang, X.; Wu, Y.; Chen, S.; Cen, J.; Yang, S.; Yang, D. Novel insight into the formation mechanism of volatile flavor in Chinese fish sauce (Yu-lu) based on molecular sensory and metagenomics analyses. Food Chem. 2020, 323, 126839. [Google Scholar] [CrossRef]
- Stahnke, L.H. Aroma components from dried sausages fermented with Staphylococcus xylosus. Meat Sci. 1994, 38, 39–53. [Google Scholar] [CrossRef]
- da Silva, G.S.; Canuto, K.M.; Ribeiro, P.R.V.; de Brito, E.S.; Nascimento, M.M.; Zocolo, G.J.; de Jesus, R.M. Chemical profiling of guarana seeds (Paullinia cupana) from different geographical origins using UPLC-QTOF-MS combined with chemometrics. Food Res. Int. 2017, 102, 700–709. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Y.; Li, X.; Zhang, H.; Chen, Q.; Kong, B. Application of lactic acid bacteria for improving the quality of reduced-salt dry fermented sausage: Texture, color, and flavor profiles. LWT-Food Sci. Technol. 2022, 154, 112723. [Google Scholar] [CrossRef]
- Ravyts, F.; Steen, L.; Goemaere, O.; Paelinck, H.; De Vuyst, L.; Leroy, F. The application of staphylococci with flavor-generating potential is affected by acidification in fermented dry sausages. Food Microbiol. 2010, 27, 945–954. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Meat | End of Ripening | ||||
|---|---|---|---|---|---|
| CK | YCC3 | MSZ2 | YM | ||
| Neutral lipids | 77.13 ± 0.78 a | 75.30 ± 0.99 a | 74.59 ± 0.60 a | 75.29 ± 0.75 a | 74.92 ± 0.10 a |
| Phospholipids | 18.16 ± 0.21 a | 12.15 ± 0.21 b | 7.01 ± 0.85 e | 10.58 ± 0.95 c | 8.92 ± 0.22 d |
| Free fatty acids | 4.71 ± 0.83 e | 12.55 ± 0.99 d | 18.40 ± 0.59 a | 14.13 ± 0.68 c | 16.16 ± 0.32 b |
| Days of Ripening (d) | |||||
|---|---|---|---|---|---|
| Batch | Raw Meat | 0 | 6 | 12 | |
| pH | CK | 5.78 ± 0.02 ax | 5.41 ± 0.02cx | 5.41 ± 0.01 cx | 5.51 ± 0.01 bx |
| YCC3 | 5.78 ± 0.02 ax | 5.30 ± 0.02 cy | 5.23 ± 0.01 dy | 5.36 ± 0.01 by | |
| MSZ2 | 5.78 ± 0.02 ax | 4.80 ± 0.01 bt | 4.70 ± 0.01 ct | 4.78 ± 0.01 bt | |
| YM | 5.78 ± 0.02 ax | 4.96 ± 0.02 cz | 4.90 ± 0.01 cz | 4.95 ± 0.01 bz | |
| POV (mmol/kg) | CK | 0.83 ± 0.06 dx | 1.23 ± 0.01 cx | 2.87 ± 0.05 bx | 3.67 ± 0.04 ax |
| YCC3 | 0.83 ± 0.06 dx | 0.98 ± 0.01 cz | 1.99 ± 0.01 bt | 2.40 ± 0.03 at | |
| MSZ2 | 0.83 ± 0.06 dx | 1.08 ± 0.01 cy | 2.61 ± 0.06 by | 2.83 ± 0.03 ay | |
| YM | 0.83 ± 0.06 dx | 0.91 ± 0.01 ct | 2.23 ± 0.04 bz | 2.66 ± 0.07 az | |
| TBARS value (mg/kg) | CK | 0.11 ± 0.00 dx | 0.12 ± 0.00 cx | 0.31 ± 0.07 bx | 0.35 ± 0.02 ax |
| YCC3 | 0.11 ± 0.00 cx | 0.10 ± 0.00 cy | 0.13 ± 0.00 bt | 0.23 ± 0.02 az | |
| MSZ2 | 0.11 ± 0.00 bx | 0.11 ± 0.01 by | 0.28 ± 0.07 ay | 0.27 ± 0.06 ay | |
| YM | 0.11 ± 0.00 cx | 0.10 ± 0.00 cy | 0.20 ± 0.01 bz | 0.25 ± 0.04 az | |
| LOX activity (U/g sample) | CK | 74.00 ± 4.46 ax | 42.95 ± 1.44 bx | 27.51 ± 2.00 cx | 18.42 ± 0.36 dx |
| YCC3 | 74.00 ± 4.46 ax | 34.96 ± 0.21 by | 26.67 ± 0.71 cx | 15.99 ± 0.07 dy | |
| MSZ2 | 74.00 ± 4.46 ax | 25.95 ± 0.37 bz | 17.23 ± 0.07 cy | 8.26 ± 0.11 dt | |
| YM | 74.00 ± 4.46 ax | 26.87 ± 0.91 bz | 18.26 ± 0.38 cy | 9.38 ± 0.25 dz |
| Days of Ripening (d) | |||||
|---|---|---|---|---|---|
| Bacterial | Batch | Raw Meat | 0 | 6 | 12 |
| LAB | CK | 4.61 ± 0.02 dx | 7.08 ± 0.08 ct | 7.80 ± 0.04 az | 7.53 ± 0.05 by |
| YCC3 | 4.61 ± 0.02 dx | 7.42 ± 0.05 bz | 7.57 ± 0.05 ay | 7.17 ± 0.03 cz | |
| MSZ2 | 4.61 ± 0.02 cx | 8.94 ± 0.04 sx | 8.88 ± 0.04 ax | 8.60 ± 0.05 bx | |
| YM | 4.61 ± 0.02 cx | 8.59 ± 0.04 by | 8.73 ± 0.05 ay | 8.69 ± 0.04 ax | |
| Staphylococcus | CK | 4.21 ± 0.02 dx | 7.42 ± 0.05 az | 6.89 ± 0.03 bz | 6.72 ± 0.05 cz |
| YCC3 | 4.21 ± 0.02 dx | 8.04 ± 0.05 ay | 8.96 ± 0.01 bx | 8.82 ± 0.02 cx | |
| MSZ2 | 4.21 ± 0.02 cx | 6.79 ± 0.03 at | 6.76 ± 0.03 at | 6.55 ± 0.09 bt | |
| YM | 4.21 ± 0.02 dx | 7.91 ± 0.02 ay | 7.84 ± 0.03 by | 7.64 ± 0.04 cy | |
| Enterobacteriaceae | CK | 4.04 ± 0.05 dx | 5.67 ± 0.03 bx | 5.98 ± 0.01 ax | 5.52 ± 0.06 cx |
| YCC3 | 4.04 ± 0.05 cx | 5.24 ± 0.04 ay | 4.63 ± 0.07 byz | 4.11 ± 0.09 cz | |
| MSZ2 | 4.04 ± 0.05 cx | 5.18 ± 0.02 ay | 4.71 ± 0.05 by | 4.58 ± 0.07 by | |
| YM | 4.04 ± 0.05 dx | 4.73 ± 0.09 az | 4.55 ± 0.07 bz | 4.24 ± 0.01 cz |
| Volatile Compound | RT | CK | YCC3 | MSZ2 | YM |
|---|---|---|---|---|---|
| Hexanal (A1) | 5.72 | 304.81 ± 11.40 a | 181.89 ± 3.91 d | 253.75 ± 7.07 b | 207.76 ± 6.17 c |
| trans-2-Hexenal (A2) | 7.42 | 38.92 ± 5.68 c | 134.28 ± 2.87 a | 86.90 ± 3.66 b | 74.33 ± 5.04 b |
| Heptaldehyde (A3) | 9.13 | 74.04 ± 1.93 c | 279.59 ± 10.38 a | 234.82 ± 8.18 a | 259.69 ± 17.02 ab |
| (Z)-Hept-2-enal (A4) | 11.22 | 212.80 ± 9.74 d | 1210.64 ± 16.77 b | 1052.60 ± 24.56 c | 1463.59 ± 29.35 a |
| (E)-2-Octenal (A5) | 14.69 | 111.68 ± 3.22 d | 326.70 ± 8.81 c | 474.92 ± 15.69 b | 637.82 ± 20.71 a |
| 1-Nonanal (A6) | 16.92 | 916.26 ± 14.34 d | 1192.00 ± 26.48 c | 1518.29 ± 33.28 b | 2311.95 ± 40.11 a |
| 2-Nonenal (A7) | 19.00 | 150.87 ± 6.97 d | 211.47 ± 8.78 c | 372.84 ± 9.33 b | 516.98 ± 10.36 a |
| (Z)-4-Decenal (A8) | 20.37 | 29.46 ± 0.88 b | n.d. | n.d. | 71.90 ± 1.01 a |
| 2-Decenal (A9) | 22.66 | 557.00 ± 34.12 b | 548.82 ± 20.64 b | 159.92 ± 7.68 c | 2249.98 ± 40.66 a |
| 2-Undecenal (A10) | 26.14 | 794.71 ± 17.68 c | 653.05 ± 10.12 d | 1488.17 ± 53.11 b | 1900.83 ± 48.42 a |
| Benzylcarboxaldehyde (A11) | 14.59 | 21.14 ± 0.12 a | n.d. | 21.99 ± 0.08 a | n.d. |
| Octanal (A12) | 13.02 | 168.38 ± 10.84 c | 417.43 ± 15.66 b | 387.40 ± 14.51 b | 506.59 ± 17.33 a |
| Decanal (A13) | 20.69 | 45.54 ± 2.52 b | 42.91 ± 2.13 b | 43.17 ± 1.84 b | 75.10 ± 3.28 a |
| 2,4-Nonadienal (A14) | 21.05 | 36.25 ± 1.25 b | n.d. | n.d. | 115.13 ± 2.36 a |
| (E,E)-2,4-decadienal (A15) | 23.79 | 358.31 ± 5.68 c | n.d. | 537.94 ± 10.33 b | 935.40 ± 12.56 a |
| Undecanal (A16) | 24.24 | 30.51 ± 0.64 c | n.d. | 39.13 ± 1.72 b | 52.66 ± 2.34 a |
| Dodecyl aldehyde (A17) | 27.59 | 80.59 ± 1.38 d | 89.12 ± 2.75 c | 102.51 ± 3.52 b | 128.60 ± 1.88 a |
| Lily aldehyde (A18) | 31.22 | 13.59 ± 0.22 c | n.d. | 29.34 ± 1.39 b | 49.39 ± 3.15 a |
| Tetradecanal (A19) | 39.39 | 25.10 ± 0.17 c | 27.59 ± 1.01 b | 106.09 ± 3.66 b | 375.24 ± 5.84 a |
| 2,4-Decadienal (A20) | 23.90 | n.d. | 42.22 ± 2.41 b | n.d. | 54.88 ± 1.28 a |
| Pentanal (A21) | 3.41 | n.d. | n.d. | 157.44 ± 5.38 b | 357.70 ± 6.32 a |
| cis-9-Tetradecenal (A22) | 30.88 | n.d. | n.d. | 57.32 ± 2.27 a | 49.54 ± 1.64 b |
| Nerylacetone (A23) | 28.78 | 69.42 ± 3.62 b | 84.42 ± 2.31 a | n.d. | 49.54 ± 2.36 c |
| 2-Heptanone (A24) | 8.69 | n.d. | n.d. | 26.26 ± 2.22 b | 31.91 ± 1.34 a |
| 5-Decanone (A25) | 22.06 | n.d. | n.d. | 53.20 ± 1.61 a | n.d. |
| Geranylacetone (A26) | 28.76 | n.d. | n.d. | 88.90 ± 4.39 a | n.d. |
| 1-Octen-3-one (A27) | 12.00 | n.d. | n.d. | n.d. | 50.14 ± 1.16 a |
| 2-Undecanone (A28) | 23.70 | n.d. | n.d. | n.d. | 55.76 ± 1.27 a |
| 2-Tridecanone (A29) | 30.25 | n.d. | n.d. | n.d. | 117.02 ± 2.10 a |
| 2,6-Octadien-1-ol (A30) | 26.53 | 19.39 ± 1.00 a | n.d. | n.d. | n.d. |
| 2,3-Butanediol (A31) | 5.52 | 57.04 ± 2.02 a | n.d. | n.d. | n.d. |
| 1-Hexanol (A32) | 8.07 | 13.28 ± 0.39 c | n.d. | 26.45 ± 2.34 b | 75.09 ± 1.79 a |
| 1-Octen-3-ol (A33) | 12.17 | 94.43 ± 1.88 d | 449.92 ± 11.97 c | 518.80 ± 15.12 b | 719.48 ± 20.39 a |
| (Z)-5-Octen-1-ol (A34) | 13.39 | 13.57 ± 0.61 a | n.d. | n.d. | n.d. |
| 1-Octyn-3-ol (A35) | 14.69 | 15.85 ± 0.54 a | n.d. | n.d. | n.d. |
| (2Z)-2-Octene-1-ol (A36) | 17.30 | 10.74 ± 0.17 a | n.d. | n.d. | n.d. |
| 2-Pentadecyn-1-ol (A37) | 25.62 | 31.87 ± 0.35 b | n.d. | 76.79 ± 1.49 a | 78.11 ± 0.92 a |
| 2,6-Octadien-1-ol (A38) | 26.53 | 19.39 ± 0.08 a | n.d. | n.d. | n.d. |
| 1-Pentanol (A39) | 4.91 | n.d. | n.d. | 56.68 ± 2.37 b | 110.41 ± 3.16 a |
| 2-Methyl-1-butanol (A40) | 5.15 | n.d. | n.d. | 32.62 ± 1.69 a | n.d. |
| n-Heptanol (A41) | 11.84 | n.d. | n.d. | 28.96 ± 0.54 b | 43.72 ± 1.58 a |
| 2,4-Dimethylpent-1-en-3-ol (A42) | 16.55 | n.d. | n.d. | 374.42 ± 9.34 b | 407.25 ± 10.47 a |
| Phenylethyl Alcohol (A43) | 17.39 | n.d. | n.d. | 71.63 ± 2.94 a | 32.78 ± 2.96 b |
| 2,4-Decadien-1-ol (A44) | 20.36 | n.d. | n.d. | 29.14 ± 1.94 a | n.d. |
| trans-2-Octen-1-ol (A45) | 26.46 | n.d. | n.d. | 52.76 ± 1.83 a | n.d. |
| 2-Hexadecanol (A46) | 23.38 | n.d. | n.d. | 26.74 ± 0.28 a | n.d. |
| 2-Octyldecanol (A47) | 32.11 | n.d. | n.d. | n.d. | 77.23 ± 1.06 a |
| 4-tert-Butylcyclohexyl acetate (A48) | 24.99 | 81.36 ± 2.05 a | 66.98 ± 3.04 b | 65.19 ± 1.75 b | n.d. |
| Propanoic acid, 2-methyl-, 1-(1,1-dimethylethyl) -2-methyl-1,3-propanediyl ester (A49) | 33.18 | 26.25 ± 1.41 c | 31.33 ± 1.06 c | 24.55 ± 1.32 c | 72.95 ± 1.85 a |
| 3,6-Octadecadiynoic acid, methyl ester (A50) | 13.40 | n.d. | 22.15 ± 0.64 a | 17.19 ± 0.85 b | n.d. |
| Octaethylene glycol monododecyl ether (A51) | 6.20 | n.d. | 64.77 ± 2.53 a | n.d. | n.d. |
| Benzoic acid, 2-[(trimethylsilyl)oxy]-, trimethylsilyl ester (A52) | 18.13 | n.d. | 428.85 ± 12.35 a | n.d. | n.d. |
| 4-Hexenoic acid, 6-hydroxy-4-methyl-, methyl ester, (E)-(A53) | 13.40 | n.d. | 47.72 ± 1.24 a | n.d. | n.d. |
| Cyclopropanetetradecanoic acid, 2-octyl-, methyl ester (A54) | 28.32 | n.d. | 18.61 ± 0.52 b | 26.36 ± 1.07 a | n.d. |
| Formic acid, hexyl ester (A55) | 8.07 | n.d. | 18.12 ± 0.96 a | n.d. | n.d. |
| 2′-Hexyl-1,1′-bicyclopropane-2-octanoic acid methyl ester (A56) | 22.17 | n.d. | n.d. | 30.71 ± 1.04 a | n.d. |
| 4-Hydroxynonanoic acid gamma-lactone (A57) | 25.95 | n.d. | n.d. | 43.24 ± 2.11 b | 129.20 ± 1.97 a |
| Phenoxyethyl isobutyrate (A58) | 30.89 | n.d. | n.d. | 28.19 ± 0.38 a | n.d. |
| Carbonic acid, ethyl hexadecyl ester (A59) | 31.41 | n.d. | n.d. | 56.82 ± 1.29 a | n.d. |
| (E)-9-Tetradecen-1-olacetate (A60) | 24.99 | n.d. | n.d. | n.d. | 56.13 ± 1.05 a |
| Ethyl caprate (A61) | 27.11 | n.d. | n.d. | n.d. | 35.18 ± 1.11 a |
| Crotonic acid, methyl ester (A62) | 29.66 | n.d. | n.d. | n.d. | 63.32 ± 2.76 a |
| 3-Hydroxydodecanoic acid (A63) | 23.54 | 10.33 ± 0.56 a | n.d. | n.d. | n.d. |
| Allantoic acid (A64) | 1.51 | n.d. | 23.07 ± 0.31 a | n.d. | n.d. |
| Undec-10-ynoic acid (A65) | 20.28 | n.d. | 42.14 ± 0.64 a | 26.95 ± 1.31 b | n.d. |
| trans-2-Hexenoic acid (A66) | 26.72 | n.d. | 89.67 ± 1.36 a | n.d. | n.d. |
| 2-Hexenoic acid (A67) | 26.88 | n.d. | 31.89 ± 1.02 a | n.d. | n.d. |
| trans-2-undecenoic acid (A68) | 27.12 | n.d. | 27.23 ± 0.09 a | n.d. | n.d. |
| Nonanoic acid (A69) | 23.29 | n.d. | n.d. | 16.99 ± 0.39 b | 242.59 ± 9.46 a |
| 7-Nonynoic acid (A70) | 12.82 | n.d. | n.d. | 62.36 ± 2.66 b | 72.87 ± 1.94 a |
| Malonamic acid (A71) | 1.96 | n.d. | n.d. | 825.88 ± 13.58 a | n.d. |
| 3-Decenoic acid (A72) | 23.37 | n.d. | n.d. | 36.37 ± 0.22 a | n.d. |
| 7-Oxooctanoic acid (A73) | 30.26 | n.d. | n.d. | 26.74 ± 1.23 a | n.d. |
| Erucic acid (A74) | 33.41 | n.d. | n.d. | 57.32 ± 2.36 a | n.d. |
| Octanoic acid (A75) | 19.81 | n.d. | n.d. | n.d. | 55.81 ± 0.99 a |
| n-Decanoic acid (A76) | 23.36 | n.d. | n.d. | n.d. | 109.79 ± 2.37 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Hou, J.; Zhang, X.; Hu, J.; Yu, Z.; Zhu, Y. Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3. Foods 2022, 11, 736. https://doi.org/10.3390/foods11050736
Wang J, Hou J, Zhang X, Hu J, Yu Z, Zhu Y. Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3. Foods. 2022; 11(5):736. https://doi.org/10.3390/foods11050736
Chicago/Turabian StyleWang, Ji, Jinna Hou, Xin Zhang, Jingrong Hu, Zhihui Yu, and Yingchun Zhu. 2022. "Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3" Foods 11, no. 5: 736. https://doi.org/10.3390/foods11050736
APA StyleWang, J., Hou, J., Zhang, X., Hu, J., Yu, Z., & Zhu, Y. (2022). Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3. Foods, 11(5), 736. https://doi.org/10.3390/foods11050736