
Detection of Balenine in Mouse Plasma after Administration of Opah-Derived Balenine by HPLC with PITC Pre-Column Derivatization

 ,
,
Abstract

1. Introduction
2. Materials and Methods
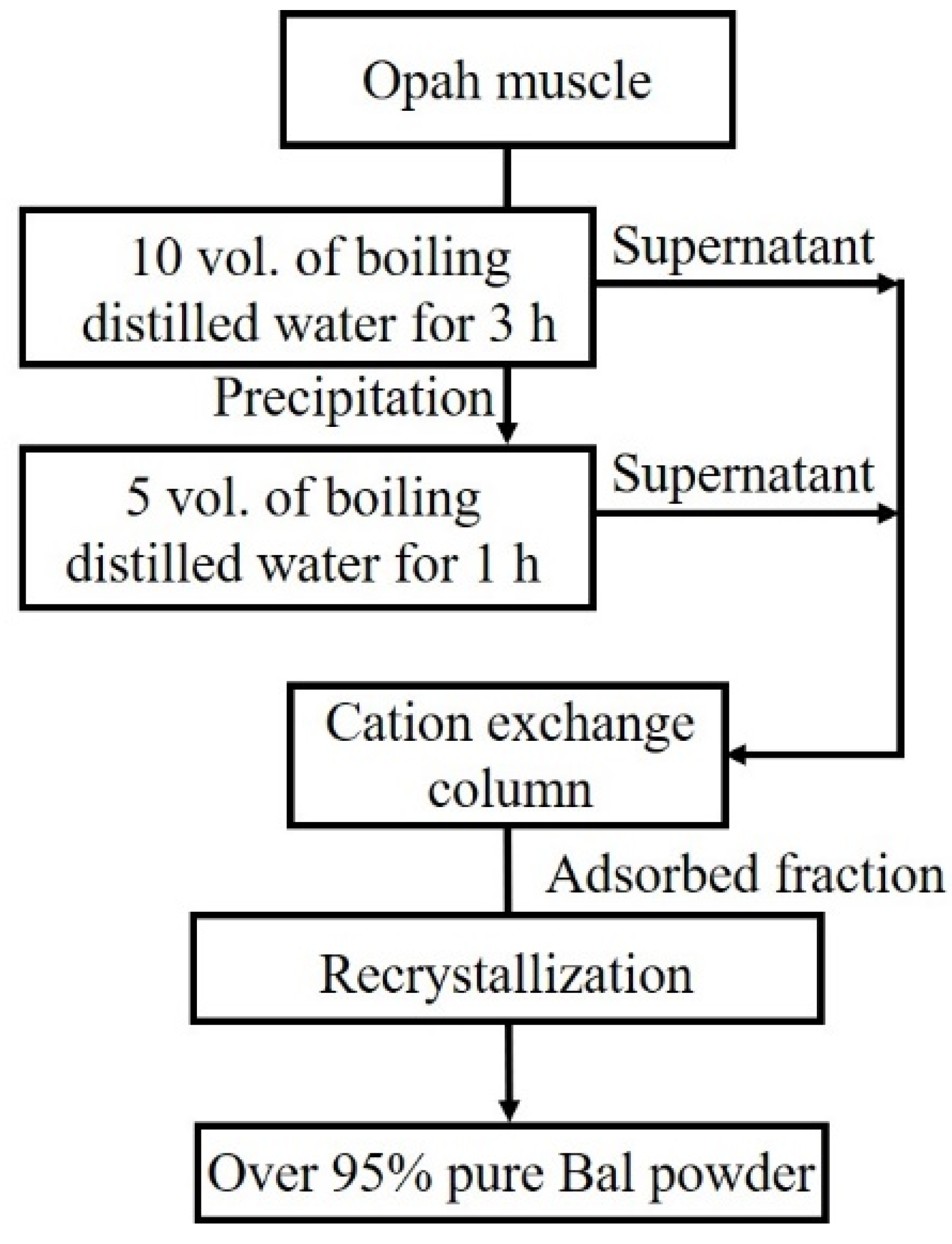
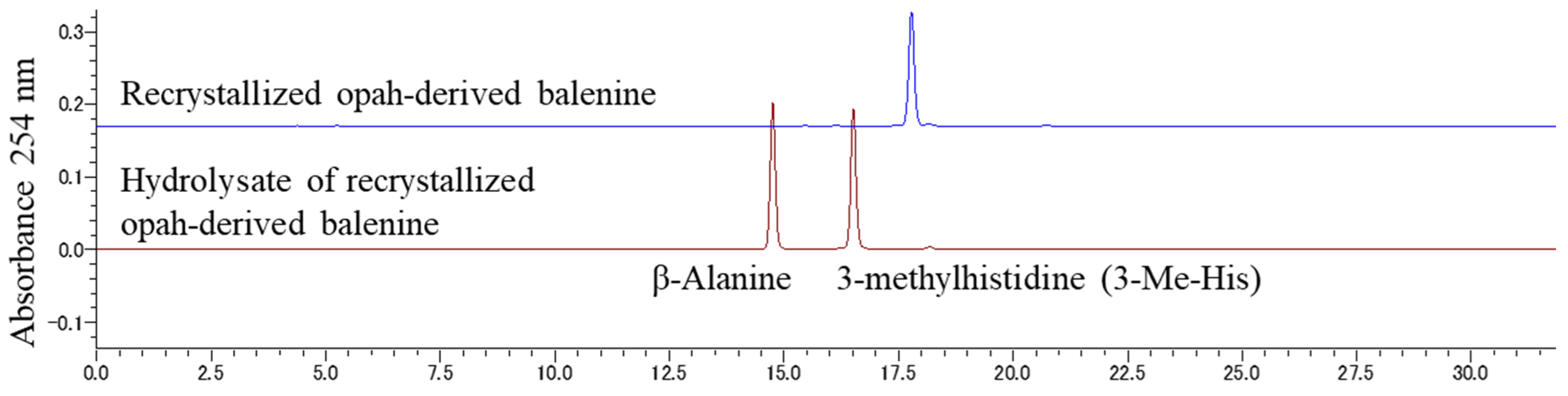
2.1. Balenine Samples
2.2. Chemicals
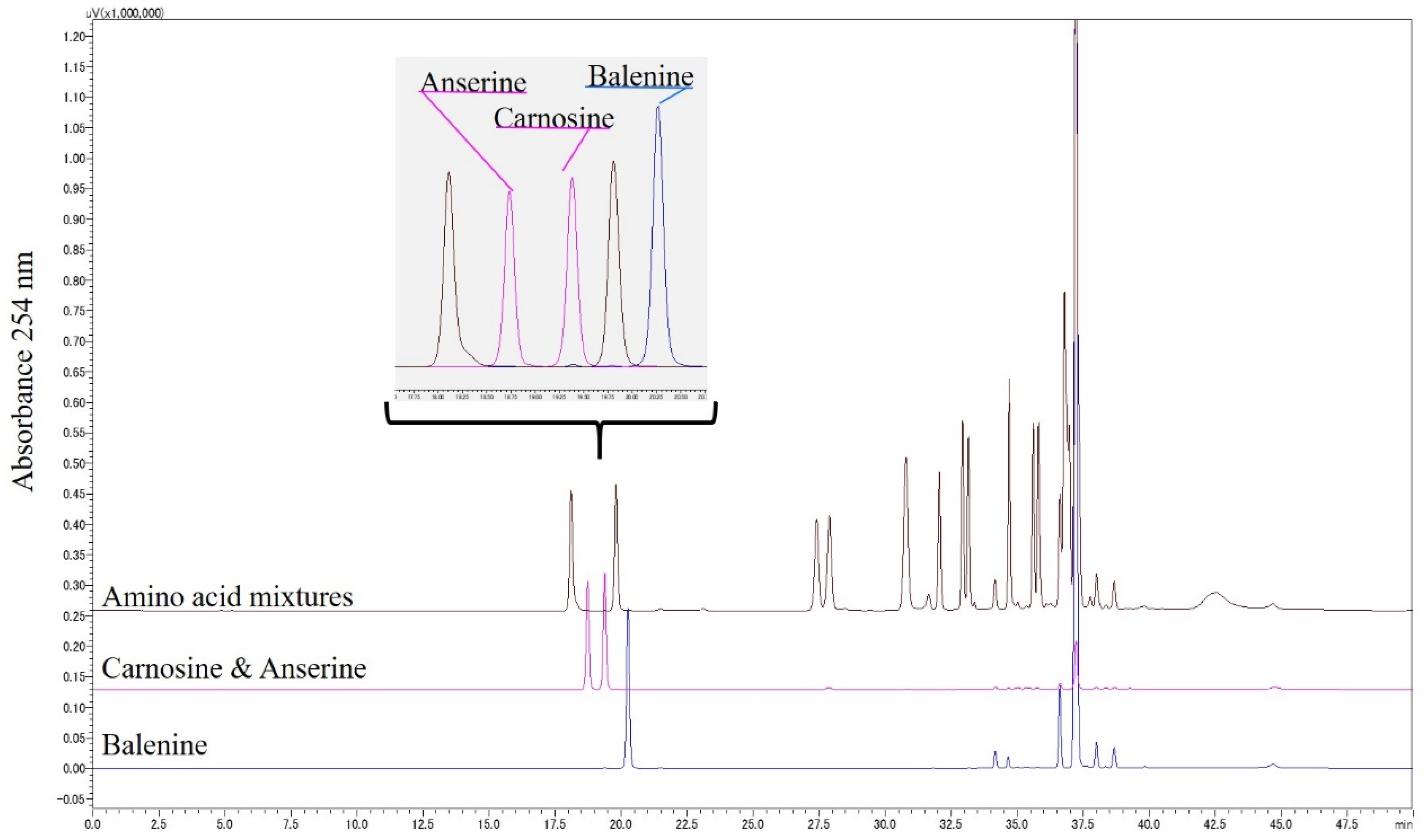
2.3. Resolution of Balenine and Related Amino Acids by HPLC
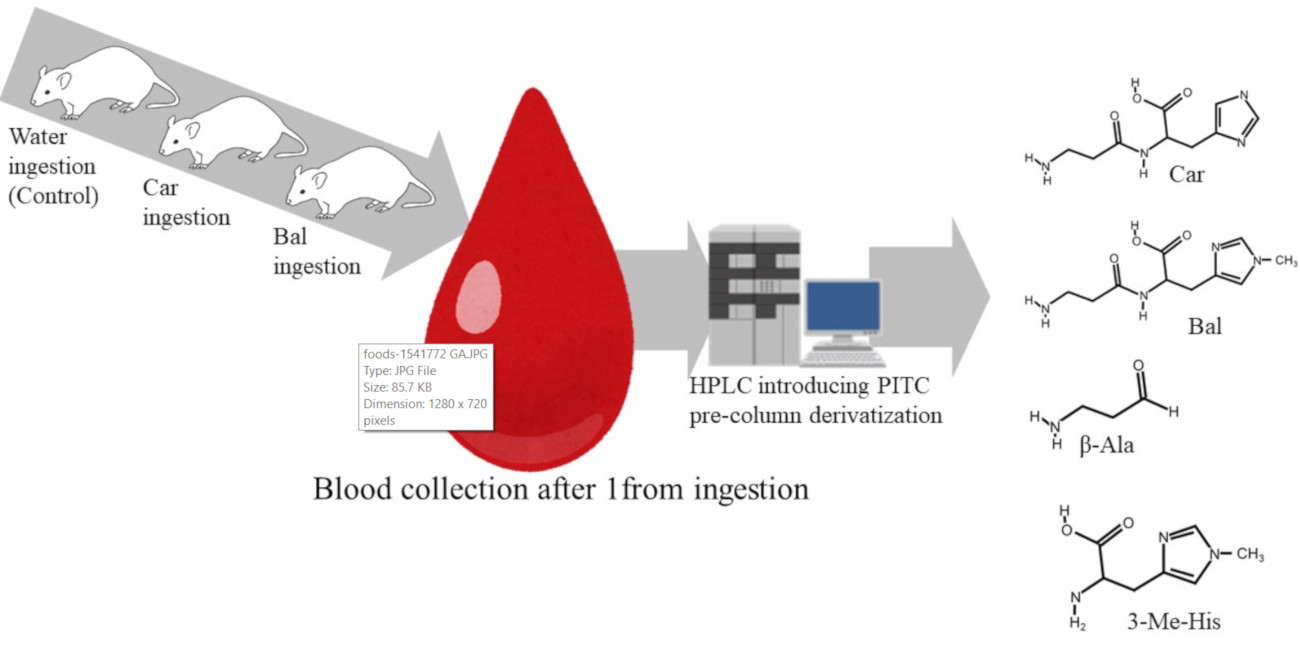
2.4. Administration of Imidazole Peptides and Blood Collection from Mice
2.5. Amino Acid Analysis of Mouse Plasma
2.6. Statistical Analysis
3. Results
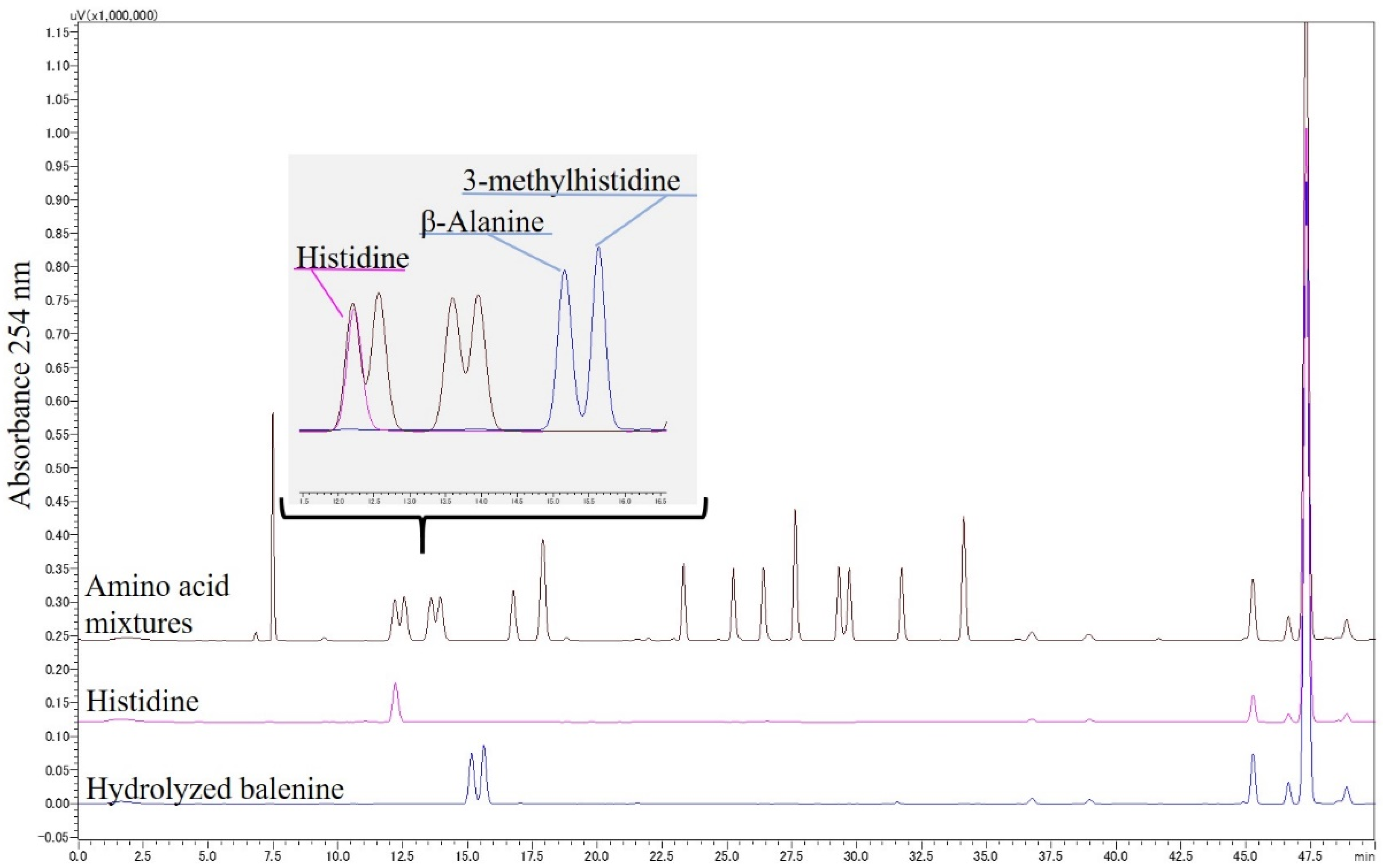
3.1. HPLC Conditions for the Resolution of Bal, β-Ala, and 3-Me-His
3.2. HPLC Detection of Food-Derived Bal, Car, and Constituent Amino Acids from Mouse Plasma
3.3. Quantitative Comparison of Imidazole Peptides and Amino Acids in Mouse Plasma
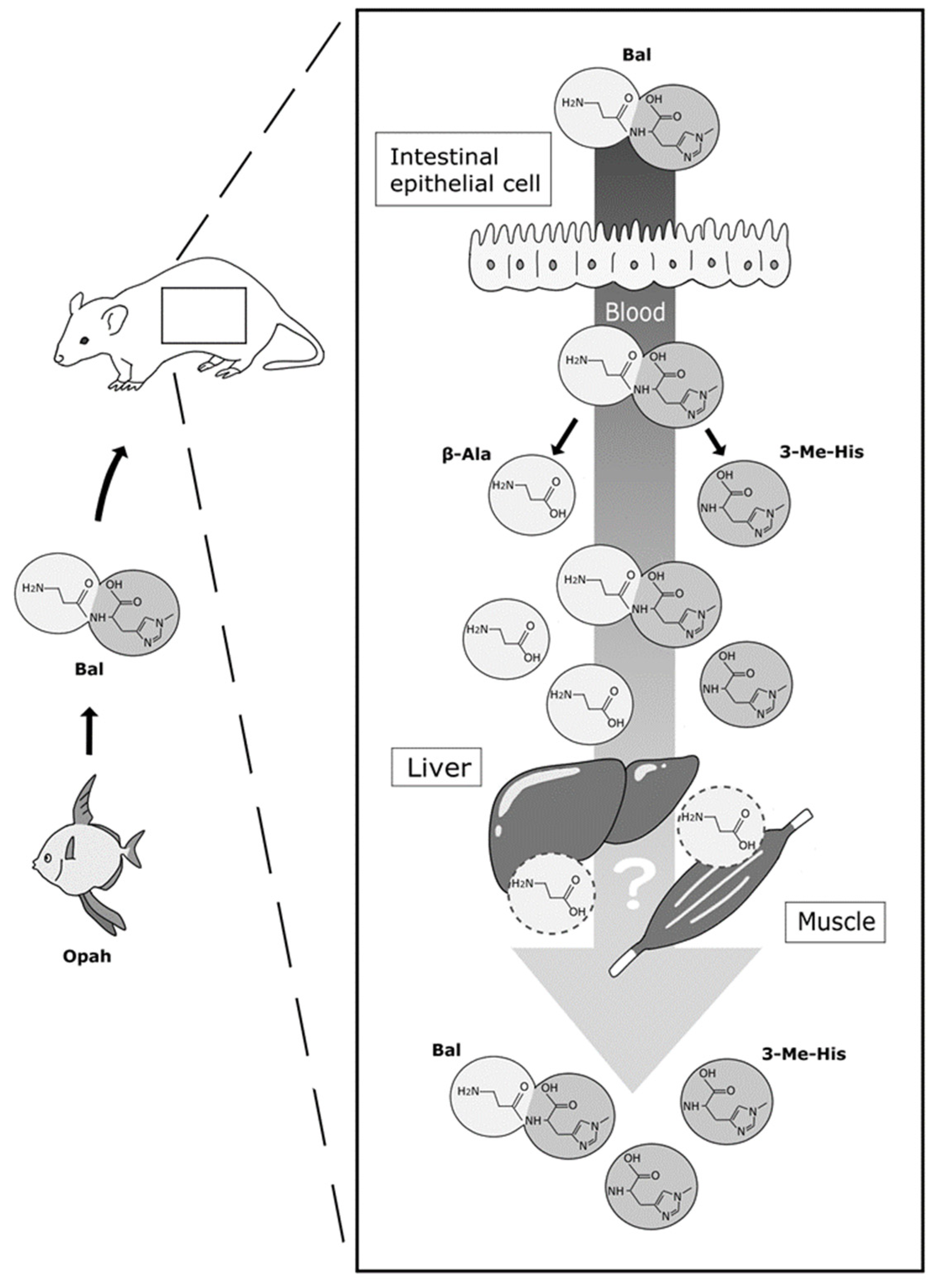
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Derave, W.; De Courten, B.; Baba, S.P. An update on carnosine and anserine research. Amino Acids 2019, 51, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A.; Crum, A.D.; Calvert, J.T. Differences in the antioxidant mechanism of carnosine in the presence of copper and iron. J. Agric. Food Chem. 1992, 40, 756–759. [Google Scholar] [CrossRef]
- Pavlov, A.R.; Revina, A.A.; Dupin, A.M.; Boldyrev, A.A.; Yaropolov, A.I. The mechanism of interaction of carnosine with superoxide radicals in water solutions. Biochim. Biophys. Acta 1993, 1157, 304–312. [Google Scholar] [CrossRef]
- Decker, E.A.; Livisay, S.A.; Zhou, S. A re-evaluation of the antioxidant activity of purified carnosine. Biochemistry 2000, 65, 766–770. [Google Scholar]
- Ihara, H.; Kakihana, Y.; Yamakage, A.; Kai, K.; Shibata, T.; Nishida, M.; Yamada, K.I.; Uchida, K. 2-Oxo-histidine-containing dipeptides are functional oxidation products. J. Biol. Chem. 2019, 294, 1279–1289. [Google Scholar] [CrossRef]
- Rezzani, R.; Favero, G.; Ferroni, M.; Lonati, C.; Moghadasian, M.H. A carnosine analog with therapeutic potentials in the treatment of disorders related to oxidative stress. PLoS ONE 2019, 14, e0215170. [Google Scholar] [CrossRef]
- Charoensin, S.; Laopaiboon, B.; Boonkum, W.; Phetcharaburanin, J.; Villareal, M.O.; Isoda, H.; Duangjinda, M. Thai Native Chicken as a Potential Functional Meat Source Rich in Anserine, Anserine/Carnosine, and Antioxidant Substances. Animals 2021, 11, 902. [Google Scholar] [CrossRef]
- Baye, E.; Ukropec, J.; de Courten, M.; Mousa, A.; Kurdiova, T.; Johnson, J.; Wilson, K.; Plebanski, M.; Aldini, G.; Ukropcova, B.; et al. Carnosine Supplementation Improves Serum Resistin Concentrations in Overweight or Obese Otherwise Healthy Adults: A Pilot Randomized Trial. Nutrients 2018, 10, 1258. [Google Scholar] [CrossRef]
- Baye, E.; Ukropec, J.; de Courten, M.; Kurdiova, T.; Krumpolec, P.; Fernández-Real, J.M.; Aldini, G.; Ukropcova, B.; de Courten, B. Carnosine supplementation reduces plasma soluble transferrin receptor in healthy overweight or obese individuals: A pilot randomised trial. Amino Acids 2019, 51, 73–81. [Google Scholar] [CrossRef]
- Elbarbary, N.S.; Ismail, E.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Pediatric Diabetes 2018, 19, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Katakura, Y.; Totsuka, M.; Imabayashi, E.; Matsuda, H.; Hisatsune, T. Anserine/Carnosine Supplementation Suppresses the Expression of the Inflammatory Chemokine CCL24 in Peripheral Blood Mononuclear Cells from Elderly People. Nutrients 2017, 9, 1199. [Google Scholar] [CrossRef] [PubMed]
- Hisatsune, T.; Kaneko, J.; Kurashige, H.; Cao, Y.; Satsu, H.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H. Effect of Anserine/Carnosine Supplementation on Verbal Episodic Memory in Elderly People. J. Alzheimer’s Dis. 2016, 50, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Rokicki, J.; Li, L.; Imabayashi, E.; Kaneko, J.; Hisatsune, T.; Matsuda, H. Daily Carnosine and Anserine Supplementation Alters Verbal Episodic Memory and Resting State Network Connectivity in Healthy Elderly Adults. Front. Aging Neurosci. 2015, 7, 219. [Google Scholar] [CrossRef]
- Szcześniak, D.; Budzeń, S.; Kopeć, W.; Rymaszewska, J. Anserine and carnosine supplementation in the elderly: Effects on cognitive functioning and physical capacity. Arch. Gerontol. Geriatr. 2014, 59, 485–490. [Google Scholar] [CrossRef]
- Cornelli, U. Treatment of Alzheimer’s disease with a cholinesterase inhibitor combined with antioxidants. Neuro-Degener. Dis. 2010, 7, 193–202. [Google Scholar] [CrossRef]
- Masuoka, N.; Yoshimine, C.; Hori, M.; Tanaka, M.; Asada, T.; Abe, K.; Hisatsune, T. Effects of Anserine/Carnosine Supplementation on Mild Cognitive Impairment with APOE4. Nutrients 2019, 11, 1626. [Google Scholar] [CrossRef]
- Wada, N.; Yamanaka, S.; Shibato, J.; Rakwal, R.; Hirako, S.; Iizuka, Y.; Kim, H.; Matsumoto, A.; Kimura, A.; Takenoya, F.; et al. Behavioral and omics analyses study on potential involvement of dipeptide balenine through supplementation in diet of senescence-accelerated mouse prone 8. Genom. Data 2016, 10, 38–50. [Google Scholar] [CrossRef][Green Version]
- Yang, M.; Sun, L.; Kawabata, Y.; Maegawa, T.; Taniyama, S.; Tachibana, K.; Hirasaka, K. Balenine, imidazole dipeptide, induces activation of superoxide dismutase in myotubes. Fish. Sci. 2021, 87, 403–409. [Google Scholar] [CrossRef]
- Yang, M.; Sun, L.; Jiang, T.; Kawabata, Y.; Murayama, F.; Maegawa, T.; Taniyama, S.; Tachibana, K.; Hirasaka, K. Safety evaluation and physiological function of dietary balenine derived from opah Lampris guttatus on skeletal muscle of mice. Int. J. Pept. Res. Ther. 2021, 27, 1–7. [Google Scholar] [CrossRef]
- Takahashi, K.; Uchida, T.; Higashi, N.; Kamei, J. L-balenine inhibits the catalytic activity of Pin1, a peptidyl prolyl cis/trans -isomerase. Pers. Med. Universe 2020, 9, 51–58. [Google Scholar] [CrossRef]
- Omura, Y.; Kimiya, T.; Matsuda, T.; Kuniyoshi, M.; Maegawa, T.; Kawabata, Y.; Adachi, S.; Amitani, Y. Analysis of balenine in muscle extract of opah Lampris guttatus with automatic amino acid analyzer. Nippon. Suisan Gakkaishi 2018, 84, 1025–1033. [Google Scholar] [CrossRef]
- Ishihara, K.; Watanabe, R.; Kato, T.; Seko, T.; Matsuda, T.; Omura, Y.; Shigemura, Y.; Kawabata, Y.; Maegawa, T. Isolation of balenine from opah (Lampris megalopsis) muscle and comparison of antioxidant and iron-chelating activities with other major imidazole dipeptides. Food Chem. 2021, 364, 130343. [Google Scholar] [CrossRef]
- Kubomura, D.; Matahira, Y.; Masui, A.; Matsuda, H. Intestinal absorption and blood clearance of L-histidine-related compounds after ingestion of anserine in humans and comparison to anserine-containing diets. J. Agric. Food Chem. 2009, 57, 1781–1785. [Google Scholar] [CrossRef] [PubMed]
- Everaert, I.; Baron, G.; Barbaresi, S.; Gilardoni, E.; Coppa, C.; Carini, M.; Vistoli, G.; Bex, T.; Stautemas, J.; Blancquaert, L.; et al. Development and validation of a sensitive LC-MS/MS assay for the quantification of anserine in human plasma and urine and its application to pharmacokinetic study. Amino Acids 2019, 51, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Volpe, S.L.; Decker, E.A. Quantitation of carnosine in humans plasma after dietary consumption of beef. J. Agric. Food Chem. 2005, 53, 4736–4739. [Google Scholar] [CrossRef] [PubMed]
- Bauchart, C.; Savary-Auzeloux, I.; Mirand, P.P.; Thomas, E.; Morzel, M.; Rémond, D. Carnosine concentration of ingested meat affects carnosine net release into the portal vein of minipigs. J. Nutr. 2007, 137, 589–593. [Google Scholar] [CrossRef][Green Version]
- Geissler, S.; Zwarg, M.; Knütter, I.; Markwardt, F.; Brandsch, M. The bioactive dipeptide anserine is transported by human proton-coupled peptide transporters. FEBS J. 2010, 277, 790–795. [Google Scholar] [CrossRef]
- Oppermann, H.; Heinrich, M.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. The proton-coupled oligopeptide transporters PEPT2, PHT1 and PHT2 mediate the uptake of carnosine in glioblastoma cells. Amino Acids 2019, 51, 999–1008. [Google Scholar] [CrossRef]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L. Rapid analysis of amino acids using pre-column derivatization. J. Chromatogr. 1984, 336, 93–104. [Google Scholar] [CrossRef]
- Iwai, K.; Hasegawa, T.; Taguchi, Y.; Morimatsu, F.; Sato, K.; Nakamura, Y.; Higashi, A.; Kido, Y.; Nakabo, Y.; Ohtsuki, K. Identification of food-derived collagen peptides in human blood after oral ingestion of gelatin hydrolysates. J. Agric. Food Chem. 2005, 53, 6531–6536. [Google Scholar] [CrossRef] [PubMed]
- Shigemura, Y.; Akaba, S.; Kawashima, E.; Park, E.Y.; Nakamura, Y.; Sato, K. Identification of a novel food-derived collagen peptide, hydroxyprolyl-glycine, in human peripheral blood by pre-column derivatisation with phenyl isothiocyanate. Food Chem. 2011, 129, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Schönherr, J. Analysis of products of animal origin in feeds by determination of carnosine and related dipeptides by high-performance liquid chromatography. J. Agric. Food Chem. 2002, 50, 1945–1950. [Google Scholar] [CrossRef] [PubMed]
- Stautemas, J.; Everaert, I.; Lefevere, F.; Derave, W. Pharmacokinetics of β-Alanine Using Different Dosing Strategies. Front. Nutr. 2018, 5, 70. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| nmol/mL Plasma | Control | Car-Administrated Mouse | Bal-Administrated Mouse |
|---|---|---|---|
| Asp | 30.46 ± 2.15 | 30.56 ± 9.69 | 34.84 ± 1.89 |
| Glu | 64.13 ± 7.80 | 59.20 ± 24.31 | 67.62 ± 10.77 |
| Asn | 16.94 ± 3.79 | 14.19 ± 4.28 | 18.64 ± 3.45 |
| Ser | 84.95 ± 15.22 | 70.14 ± 6.08 | 95.44 ± 4.15 |
| Gln | 224.71 ± 6.42 | 205.01 ± 33.75 | 276.79 ± 6.16 |
| Gly | 179.38 ± 8.94 | 154.14 ± 18.80 | 193.63 ± 36.79 |
| His | 39.20 ± 1.36 | 47.02 ± 5.75 | 44.08 ± 6.36 |
| Arg | 53.21 ± 25.01 | 43.13 ± 22.16 | 31.66 ± 3.48 |
| Thr | 94.71 ± 12.98 | 73.47 ± 9.67 | 90.78 ± 4.61 |
| Ala | 282.50 ± 46.88 | 235.93 ± 38.22 | 303.85 ± 22.53 |
| Pro | 101.35 ± 10.30 | 96.48 ± 12.92 | 111.30 ± 11.03 |
| Tyr | 92.53 ± 10.24 | 81.38 ± 17.60 | 90.61 ± 9.93 |
| Val | 218.21 ± 9.05 | 193.44 ± 36.32 | 228.41 ± 22.89 |
| Met | 41.92 ± 3.43 | 35.91 ± 3.97 | 42.74 ± 2.64 |
| Cys | 15.78 ± 3.54 | 22.80 ± 7.87 | 22.71 ± 3.65 |
| Ile | 90.94 ± 5.93 | 76.78 ± 18.73 | 94.37 ± 12.55 |
| Leu | 120.52 ± 15.26 | 106.71 ± 22.03 | 129.08 ± 14.32 |
| Phe | 56.86 ± 4.36 | 54.01 ± 7.51 | 64.03 ± 5.93 |
| Lys | 100.91 ± 4.09 | 93.03 ± 5.14 | 96.67 ± 4.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shigemura, Y.; Iwasaki, Y.; Sato, Y.; Kato, T.; Seko, T.; Ishihara, K. Detection of Balenine in Mouse Plasma after Administration of Opah-Derived Balenine by HPLC with PITC Pre-Column Derivatization. Foods 2022, 11, 590. https://doi.org/10.3390/foods11040590
Shigemura Y, Iwasaki Y, Sato Y, Kato T, Seko T, Ishihara K. Detection of Balenine in Mouse Plasma after Administration of Opah-Derived Balenine by HPLC with PITC Pre-Column Derivatization. Foods. 2022; 11(4):590. https://doi.org/10.3390/foods11040590
Chicago/Turabian StyleShigemura, Yasutaka, Yu Iwasaki, Yoshio Sato, Tomomi Kato, Takuya Seko, and Kenji Ishihara. 2022. "Detection of Balenine in Mouse Plasma after Administration of Opah-Derived Balenine by HPLC with PITC Pre-Column Derivatization" Foods 11, no. 4: 590. https://doi.org/10.3390/foods11040590
APA StyleShigemura, Y., Iwasaki, Y., Sato, Y., Kato, T., Seko, T., & Ishihara, K. (2022). Detection of Balenine in Mouse Plasma after Administration of Opah-Derived Balenine by HPLC with PITC Pre-Column Derivatization. Foods, 11(4), 590. https://doi.org/10.3390/foods11040590