
Structural Transitions of Alpha-Amylase Treated with Pulsed Electric Fields: Effect of Coexisting Carrageenan

Abstract
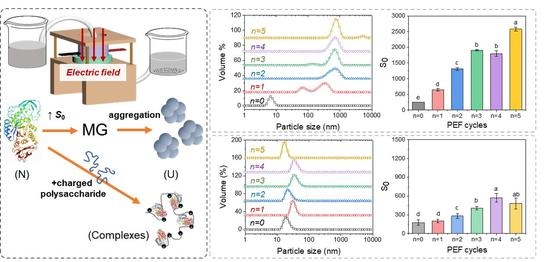
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Pulsed Electric Field (PEF) Treatment
2.4. Measurement of Enzyme Activity
2.5. Intrinsic Fluorescence
2.6. Circular Dichroism (CD) Studies
2.7. Surface Hydrophobicity (S0)
2.8. Dynamic Light Scattering and Surface Charges
2.9. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.10. Statistics Analyses
3. Results and Discussion
3.1. MG State of α-Amylase Induced by PEF during Unfolding Process
3.2. Effect of Carrageenan on the Unfolding of α-Amylase Induced by PEF
3.2.1. Effect of Carrageenan on Enzyme Activity
3.2.2. Effect of λ-Carrageenan on the Intrinsic Fluorescence
3.2.3. Effect of λ-Carrageenan on the Aggregation Behavior
3.2.4. Formation of α-Amylase/λ-Carrageenan Complexes
3.3. Proposed Mechanism of λ-Carrageenan Affecting the Unfolding of α-Amylase
3.3.1. Secondary Structure of α-Amylase and Complexes after PEF Treatment
3.3.2. Surface Hydrophobicity of α-Amylase and Complexes after PEF Treatment
3.3.3. SDS-PAGE of α-Amylase and Complexes after PEF Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Ye, Y.; Deng, J.; Zhao, P.; Mao, Y.; Chen, Z. Rapid unfolding of pig pancreas α-amylase: Kinetics, activity and structure evolution. Food Chem. 2022, 368, 130795. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, Y.; Endo, S.; Nakamura, H. How A Novel Scientific Concept Was Coined the "Molten Globule State". Biomolecules 2020, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.Q.; Liao, K.F.; Shih, O.; Shiu, Y.J.; Wu, W.R.; Su, C.J.; Lin, P.C.; Jeng, U.S. Probing the Acid-Induced Packing Structure Changes of the Molten Globule Domains of a Protein near Equilibrium Unfolding. J. Phys. Chem. Lett. 2017, 8, 470. [Google Scholar] [CrossRef]
- Zhu, P.; Yang, Y.; Zou, L.; Gao, J.; Zhang, N. The effect of trehalose on the thermodynamic stability and emulsification of soybean 11S globulin in the molten globule state. Food Hydrocoll. 2021, 118, 106811. [Google Scholar]
- Shi, Y.J.; Chiou, J.T.; Huang, C.H.; Lee, Y.C.; Chang, L.S. Modification of carboxyl groups converts α-lactalbumin into an active molten globule state with membrane-perturbing activity and cytotoxicity. Int. J. Biol. Macromol. 2020, 163, 1697–1706. [Google Scholar] [CrossRef]
- Tadashi, K.; Tomokadu, M.; Dai, K.; Takuya, H.; Keiichi, K. Thermodynamic and kinetic properties of sorbitol-induced molten globule of myoglobin. J. Therm. Anal. Calorim. 2016, 123, 1861–1869. [Google Scholar]
- Wijayanti, H.B.; Bansal, N.; Deeth, H.C. Stability of Whey Proteins during Thermal Processing: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1235–1251. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Chen, F.; Piekoszewski, W.; Yang, Y.; Wang, B.; Liu, L.; Guan, H.; Shi, Y.; Zhang, N. The effect of extreme acid-induced soybean glycinin molten globules state on foaming ability. Food Hydrocoll. 2020, 105, 105819. [Google Scholar] [CrossRef]
- Zhu, P.; Ma, C.; Fan, J.; Yang, Y.; Liu, X.; Bian, X.; Ren, L.; Xu, Y.; Yu, D.; Liu, L. The interaction of trehalose and molten globule state soybean 11S globulin and its impact on foaming capacities. J. Sci. Food Agric. 2022, 118, 106811. [Google Scholar] [CrossRef]
- Asghari, S.M.; Khajeh, K.; Moradian, F.; Ranjbar, B.; Naderi-Manesh, H. Acid-induced conformational changes in Bacillus amyloliquefaciens α-amylase: Appearance of a molten globule like state. Enzym. Microb. Technol. 2004, 35, 51–57. [Google Scholar] [CrossRef]
- Shokri, M.M.; Khajeh, K.; Alikhajeh, J.; Asoodeh, A.; Ranjbar, B.; Hosseinkhani, S.; Sadeghi, M. Comparison of the molten globule states of thermophilic and mesophilic α-amylases. Biophys. Chem. 2006, 122, 58–65. [Google Scholar] [CrossRef]
- Jin, W.; Wang, Z.; Peng, D.; Shen, W.; Huang, Q. Effect of pulsed electric field on assembly structure of α-amylase and pectin electrostatic complexes. Food Hydrocoll. 2020, 101, 105547. [Google Scholar] [CrossRef]
- Antonov, Y.A.; Zhuravleva, I.L. Complexation of lysozyme with lambda carrageenan: Complex characterization and protein stability. Food Hydrocoll. 2019, 87, 519–529. [Google Scholar] [CrossRef]
- Wu, S.; Lai, M.; Luo, J.; Pan, J.; Zhang, L.M.; Yang, L. Interactions between α-amylase and an acidic branched polysaccharide from green tea. Int. J. Biol. Macromol. 2017, 94, 669–678. [Google Scholar] [CrossRef]
- María, M.; Pokhrel, P.R.; René, R.; Barbosa-Canovas, G.V.; Hernández-Sánchez, H. Effect of pulsed electric fields on the activity of food-grade papain in a continuous system. Lebensm. Wiss. Und Technol. 2019, 109, 336–341. [Google Scholar]
- Giteru, S.G.; Oey, I.; Ali, M.A. Feasibility of using pulsed electric fields to modify biomacromolecules: A review. Trends Food Sci. Technol. 2018, 72, 91–113. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, L.; Ju, H.; Bao, Z.; Zeng, X.-a.; Lin, S. Research advances and application of pulsed electric field on proteins and peptides in food. Food Res. Int. 2021, 139, 109914. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, R.; Hu, J.; Luan, Y.; Liu, R.; Ge, Q.; Yu, H.; Wu, M. Moderate pulsed electric field-induced structural unfolding ameliorated the gelling properties of porcine muscle myofibrillar protein. Innov. Food Sci. Emerg. Technol. 2022, 81, 103145. [Google Scholar] [CrossRef]
- Wei, J.; Zeng, X.; Tang, T.; Jiang, Z.; Liu, Y. Unfolding and nanotube formation of ovalbumin induced by pulsed electric field. Innov. Food Sci. Emerg. Technol. 2018, 45, 249–254. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, Z.; Gao, Y.; Bao, Z.; Sun, N.; Lin, S. Mechanism of trypsin activation by pulsed electric field treatment revealed based on chemical experiments and molecular dynamics simulations. Food Chem. 2022, 394, 133477. [Google Scholar] [CrossRef]
- Roldan-Cruz, C.; Garcia-Hernandez, A.; Alvarez-Ramirez, J.; Vernon-Carter, E.J. Effect of the stirring speed in the in vitro activity of α-amylase. Food Hydrocoll. 2020, 110, 106127. [Google Scholar] [CrossRef]
- Jin, W.; Wang, Z.; Peng, D.; Shen, W.; Zhu, Z.; Cheng, S.; Bin, L.; Huang, Q. Effect of linear charge density of polysaccharides on interactions with α-amylase: Self-Assembling behavior and application in enzyme immobilization–ScienceDirect. Food Chem. 2020, 331, 127320. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, M.; Chang, R.; Xiong, L.; Sun, Q. In vitro inhibition of pancreatic α-amylase by spherical and polygonal starch nanoparticles. Food Funct. 2018, 9, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Zou, Y.; Huang, S.; Liu, Y.; Lin, J. Study on the physicochemical and emulsifying property of proteins extracted from Pleurotus tuoliensis. LWT-Food Sci. Technol. 2021, 151, 112185. [Google Scholar] [CrossRef]
- Dar, M.A.; Haque, M.A.; Islam, A.; Hassan, M.I.; Ahmad, F. Characterisation of molten globule-like state of sheep serum albumin at physiological pH. Int. J. Biol. Macromol. 2016, 89, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhao, W.; Yang, R.; Yan, W.; Sun, Q. Aggregation of egg white proteins with pulsed electric fields and thermal processes. J. Sci. Food Agric. 2016, 96, 3334–3341. [Google Scholar] [CrossRef] [PubMed]
- Kuwajima, K. The Molten Globule, and Two-State vs. Non-Two-State Folding of Globular Proteins. Biomolecules 2020, 10, 407. [Google Scholar] [CrossRef]
- Mishra, P.; Jha, S.K.; Acharya, N. Evidence for dry molten globule-like domains in the pH-induced equilibrium folding intermediate of a multidomain protein. J. Phys. Chem. Lett. 2016, 7, 173–179. [Google Scholar]
- Parray, Z.A.; Ahmad, F.; Alajmi, M.F.; Hussain, A.; Hassan, M.I.; Islam, A. Formation of molten globule state in horse heart cytochrome c under physiological conditions: Importance of soft interactions and spectroscopic approach in crowded milieu. Int. J. Biol. Macromol. 2020, 148, 192–200. [Google Scholar] [CrossRef]
- Xiao, J.; Nian, S.; Huang, Q. Assembly of kafirin/carboxymethyl chitosan nanoparticles to enhance the cellular uptake of curcumin. Food Hydrocoll. 2015, 51, 166–175. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Xu, X.; Meng, Y.; Zhang, L.; Chen, J.; Qiu, R. Short communication: Changes in fluorescence intensity induced by soybean soluble polysaccharide–milk protein interactions during acidification. J. Dairy Sci. 2015, 98, 8577–8580. [Google Scholar] [CrossRef]
- Wu, L.; Zhao, W.; Yang, R.; Yan, W. Pulsed electric field (PEF)-induced aggregation between lysozyme, ovalbumin and ovotransferrin in multi-protein system. Food Chem. 2015, 175, 115–120. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; Jiang, W.; Qian, J. Effect of pulsed electric field on functional and structural properties of canola protein by pretreating seeds to elevate oil yield. LWT 2017, 84, 73–81. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, T. The modification of ovalbumin surface properties treated by pulsed electric field combined with divalent metal ions. Food Chem. 2019, 293, 455–462. [Google Scholar] [CrossRef]
- Li, J. Effect of Charge Density of Polysaccharide on Self-assembly Behaviors of Ovalbumin and Sodium Alginate. International J. Biol. Macromol. 2019, 154, 1245–1254. [Google Scholar] [CrossRef]
- Sohail, S.K.; Anne, P.; Davide, D.F.; Gea, G.; Oliver, P.; Dominic, B.; Raftery, M.J.; Parkin, D.M.; Jill, T.; Ricardo, C. A chemically modified alpha-amylase with a molten-globule state has entropically driven enhanced thermal stability. Protein Eng. Des. Sel. 2010, 23, 769–780. [Google Scholar]
- Kikani, B.A.; Singh, S.P. Enzyme stability, thermodynamics and secondary structures of α-amylase as probed by the CD spectroscopy. Int. J. Biol. Macromol. 2015, 81, 450–460. [Google Scholar] [CrossRef]
- Dong, M.; Xu, Y.; Zhang, Y.; Wang, P.; Zhou, G. Physicochemical and structural properties of myofibrillar proteins isolated from pale, soft, exudative (PSE)-like chicken breast meat: Effects of pulsed electric field (PEF). Innov. Food Sci. Emerg. Technol. 2020, 59, 102277. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhang, J.; Li, C.; Huang, W.; Guo, C.; Jin, W.; Shen, W. Structural Transitions of Alpha-Amylase Treated with Pulsed Electric Fields: Effect of Coexisting Carrageenan. Foods 2022, 11, 4112. https://doi.org/10.3390/foods11244112
Li J, Zhang J, Li C, Huang W, Guo C, Jin W, Shen W. Structural Transitions of Alpha-Amylase Treated with Pulsed Electric Fields: Effect of Coexisting Carrageenan. Foods. 2022; 11(24):4112. https://doi.org/10.3390/foods11244112
Chicago/Turabian StyleLi, Junzhu, Jiayu Zhang, Chen Li, Wenjing Huang, Cheng Guo, Weiping Jin, and Wangyang Shen. 2022. "Structural Transitions of Alpha-Amylase Treated with Pulsed Electric Fields: Effect of Coexisting Carrageenan" Foods 11, no. 24: 4112. https://doi.org/10.3390/foods11244112
APA StyleLi, J., Zhang, J., Li, C., Huang, W., Guo, C., Jin, W., & Shen, W. (2022). Structural Transitions of Alpha-Amylase Treated with Pulsed Electric Fields: Effect of Coexisting Carrageenan. Foods, 11(24), 4112. https://doi.org/10.3390/foods11244112