
Anti-Colon Cancer Activity of Novel Peptides Isolated from In Vitro Digestion of Quinoa Protein in Caco-2 Cells


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
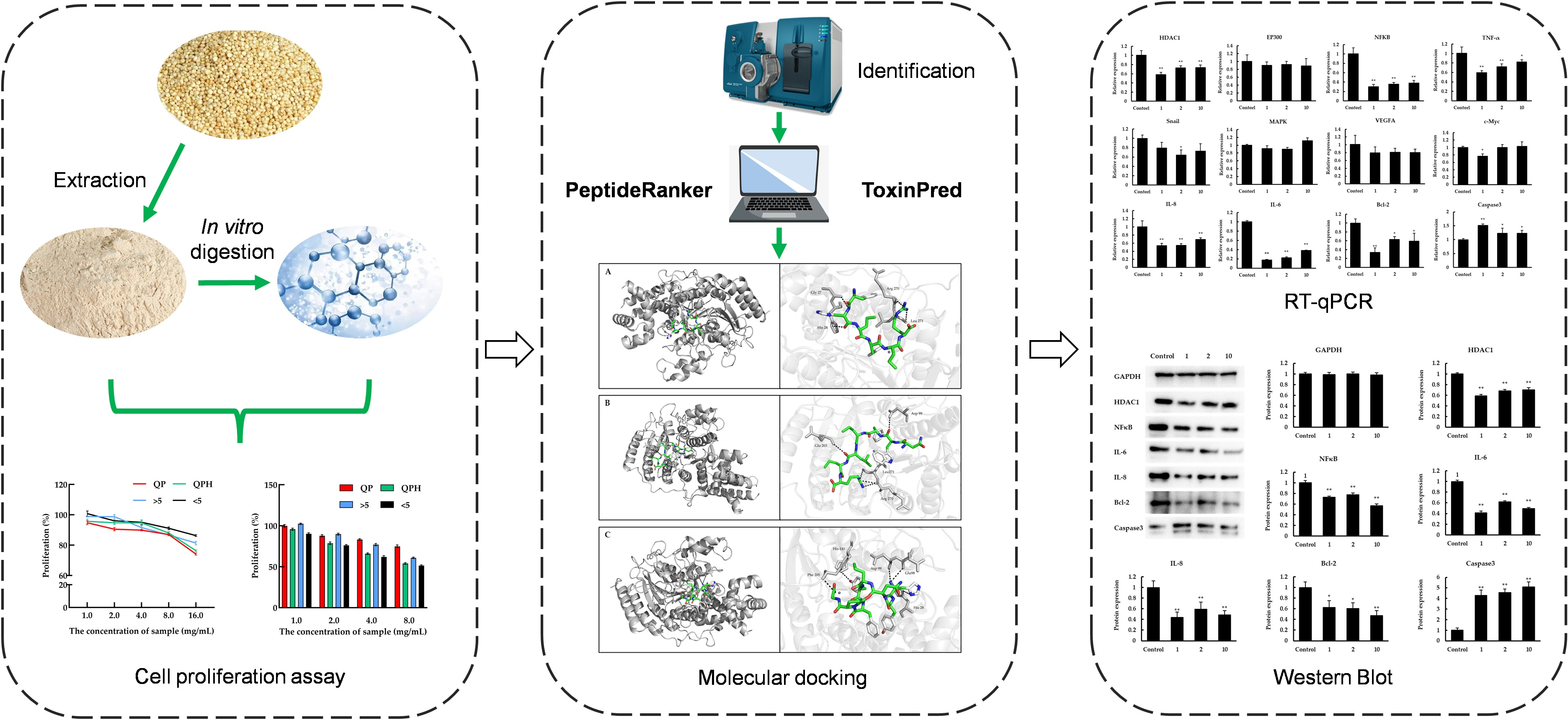
2.2. Preparation of Quinoa Protein Concentrate, Quinoa Protein Hydrolysate and Quinoa Peptides
2.3. Peptide Screening and Synthesis
2.4. Cell Proliferation Assay
2.5. HDAC1 Inhibitory Activity Assay
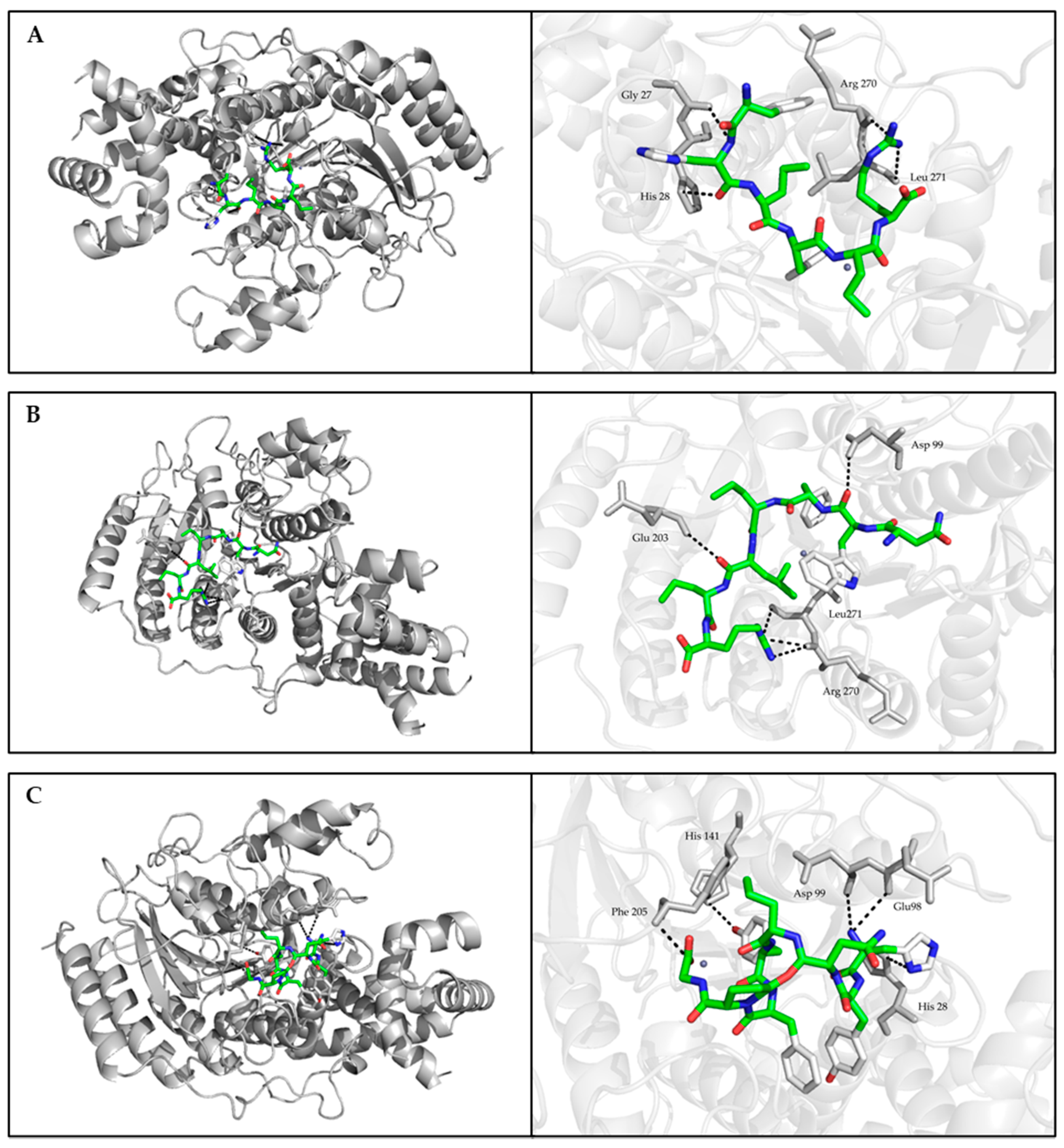
2.6. Molecular Docking
2.7. Quantitive Real Time-Polymerase Chain Reaction (RT-qPCR)
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Antiproliferative Activity of QPH against Caco-2 Cells
3.2. In Silico Analysis and Antiproliferative Activity of Quinoa Peptides
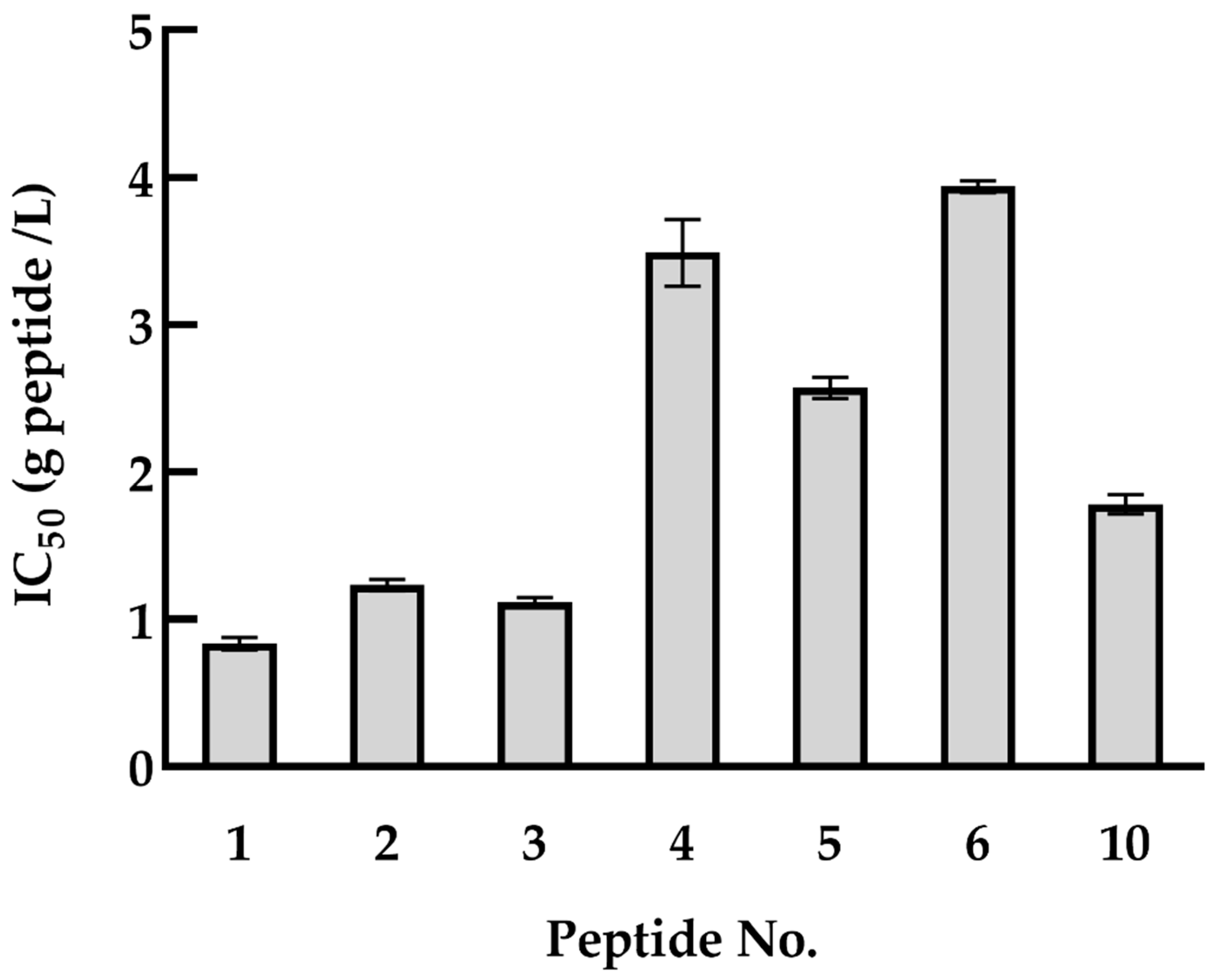
3.3. HDAC1 Inhibitory Activity of Quinoa Peptides
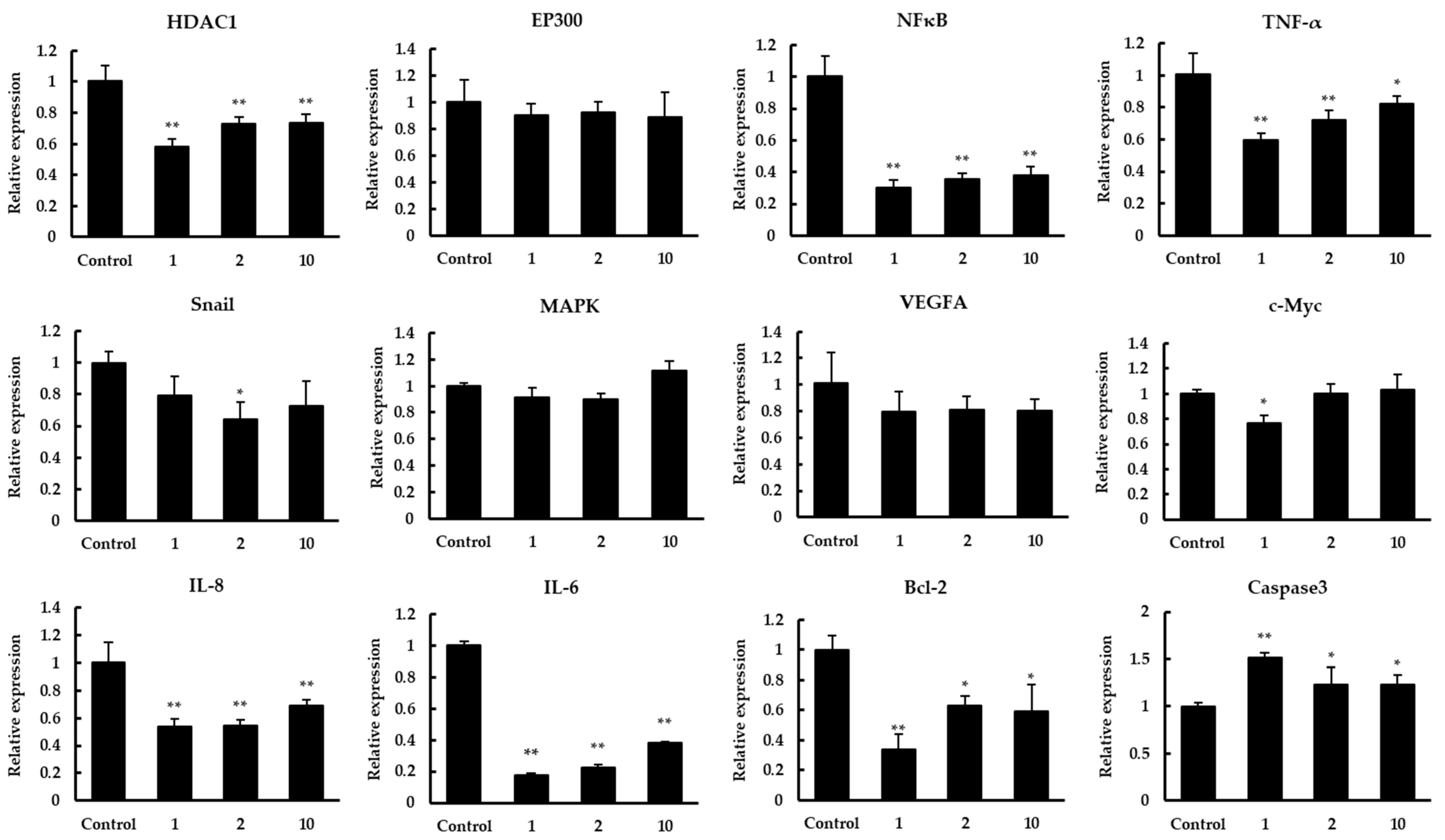
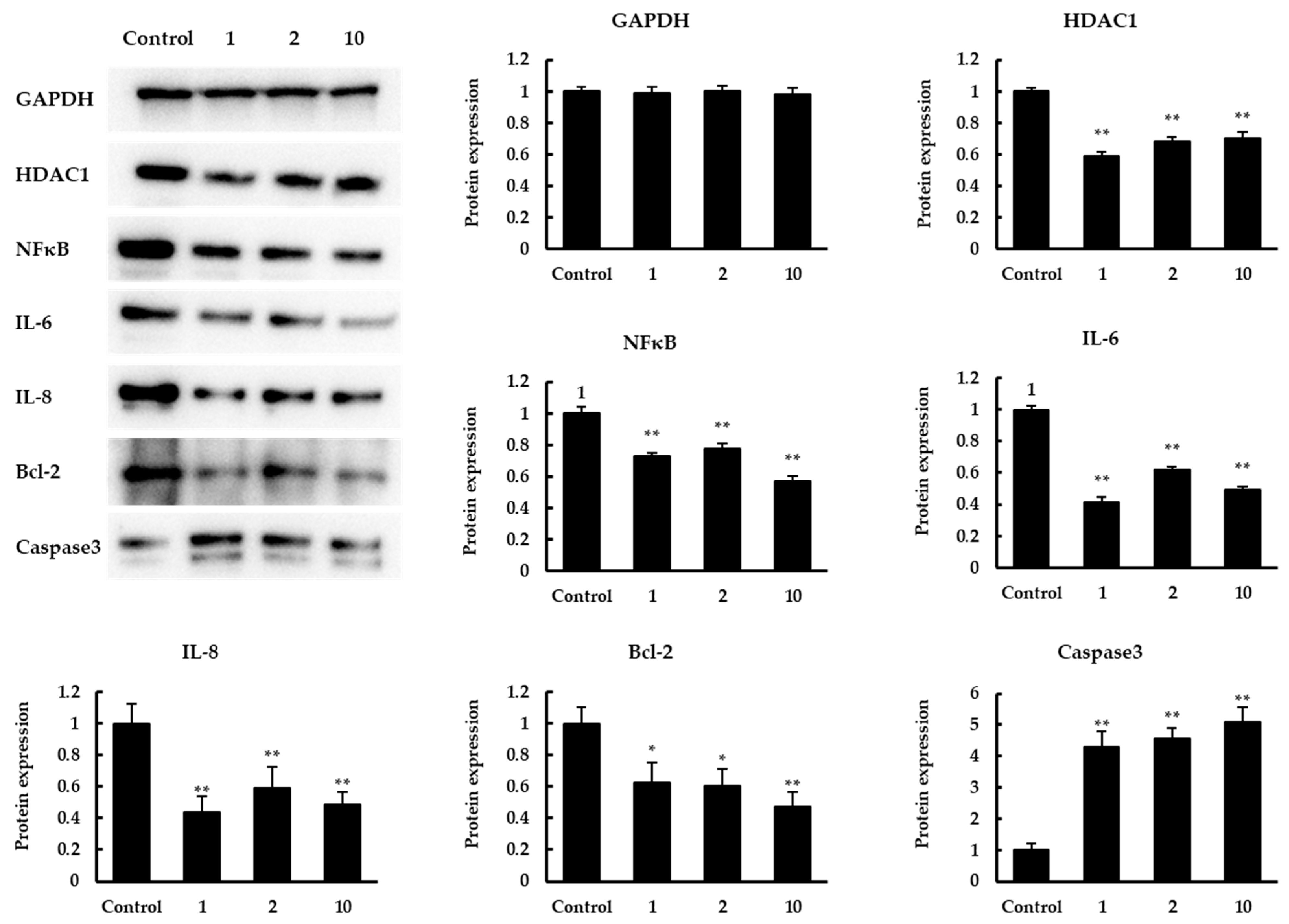
3.4. The Effect of Quinoa Peptides on HDAC1-Induced Cancer Progression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yan, W.; Sun, C.J.; Yuan, J.W.; Yang, N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017, 7, 45308. [Google Scholar] [CrossRef]
- Muenchau, S.; Deutsch, R.; de Castro, I.J.; Hielscher, T.; Heber, N.; Niesler, B.; Lusic, M.; Stanifer, M.L.; Boulant, S. Hypoxic Environment Promotes Barrier Formation in Human Intestinal Epithelial Cells through Regulation of MicroRNA 320a Expression. Mol. Cell. Biol. 2019, 39, e00553-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.M.; He, R.Z.; Tan, T.; Li, J.; Hu, Z.; Luo, W.H.; Duan, L.L.; Luo, W.N.; Luo, D.X. A novel long non-coding RNA-KAT7 is low expressed in colorectal cancer and acts as a tumor suppressor. Cancer Cell Int. 2019, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.R.; Abar, L.; Chan, D.S.M.; Vingeliene, S.; Polemiti, E.; Stevens, C.; Greenwood, D.; Norat, T. Foods and beverages and colorectal cancer risk: A systematic review and meta-analysis of cohort studies, an update of the evidence of the WCRF-AICR Continuous Update Project. Ann. Oncol. 2017, 28, 1788–1802. [Google Scholar] [CrossRef] [PubMed]
- Neri-Numa, I.A.; Carvalho-Silva, L.B.; Morales, J.P.; Malta, L.G.; Muramoto, M.T.; Ferreira, J.E.M.; de Carvalho, J.E.; Ruiz, A.L.T.G.; Marostica, M.R.; Pastore, G.M. Evaluation of the antioxidant, antiproliferative and antimutagenic potential of araca-boi fruit (Eugenia stipitata Mc Vaugh—Myrtaceae) of the Brazilian Amazon Forest. Food Res. Int. 2013, 50, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Jahandideh, F.; Wu, J.P. Food-Derived Bioactive Peptides on Inflammation and Oxidative Stress. Biomed Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruales, J.; Nair, B.M. Nutritional quality of the protein in quinoa (Chenopodium quinoa, Willd) seeds. Plant Foods Hum. Nutr. 1992, 42, 1–11. [Google Scholar] [CrossRef]
- Swieca, M.; Seczyk, L.; Gawlik-Dziki, U.; Dziki, D. Bread enriched with quinoa leaves—The influence of protein-phenolics interactions on the nutritional and antioxidant quality. Food Chem. 2014, 162, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Miralles, B.; Carrillo, W.; Hernandez-Ledesma, B. In vitro chemopreventive properties of peptides released from quinoa (Chenopodium quinoa Willd.) protein under simulated gastrointestinal digestion. Food Res. Int. 2018, 105, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Srdic, M.; Ovcina, I.; Fotschki, B.; Haros, C.M.; Llopis, J.M.L. C. quinoa and S. hispanica L. Seeds Provide Immunonutritional Agonists to Selectively Polarize Macrophages. Cells 2020, 9, 593. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.M.; Hao, Y.Q.; Richel, A.; Everaert, N.; Chen, Y.N.; Liu, M.J.; Yang, X.S.; Ren, G.X. Antihypertensive effect of quinoa protein under simulated gastrointestinal digestion and peptide characterization. J. Sci. Food Agric. 2020, 100, 5569–5576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.B.; Bu, L.; Hu, J.; Xu, Z.Y.; Ruan, L.B.; Fang, Y.; Wang, P. HDAC1 knockdown inhibits invasion and induces apoptosis in non-small cell lung cancer cells. Biol. Chem. 2018, 399, 603–610. [Google Scholar] [CrossRef]
- Weichert, W. HDAC expression and clinical prognosis in human malignancies. Cancer Lett. 2009, 280, 168–176. [Google Scholar] [CrossRef]
- Eshelman, M.A.; Shah, M.; Raup-Konsavage, W.M.; Rennoll, S.A.; Yochum, G.S. TCF7L1 recruits CtBP and HDAC1 to repress DICKKOPF4 gene expression in human colorectal cancer cells. Biochem. Biophys. Res. Commun. 2017, 487, 716–722. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, Y.; Chen, W.; Wang, X.; Miao, Z.; Cao, L.; Li, F.; Wang, G. By recruiting HDAC1, MORC2 suppresses p21 Waf1/Cip1 in gastric cancer. Oncotarget 2015, 6, 16461–16470. [Google Scholar] [CrossRef] [Green Version]
- Selokar, N.L.; St John, L.; Revay, T.; King, W.A.; Singla, S.K.; Madan, P. Effect of Histone Deacetylase Inhibitor Valproic Acid Treatment on Donor Cell Growth Characteristics, Cell Cycle Arrest, Apoptosis, and Handmade Cloned Bovine Embryo Production Efficiency. Cell. Reprogram. 2013, 15, 531–542. [Google Scholar] [CrossRef]
- Mariadason, J.M. HDACs and HDAC inhibitors in colon cancer. Epigenetics 2008, 3, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.C.; Martinez-Villaluenga, C.; de Lumen, B.O.; Hernandez-Ledesma, B. Updating the research on the chemopreventive and therapeutic role of the peptide lunasin. J. Sci. Food Agric. 2018, 98, 2070–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvez, A.F.; Chen, N.; Macasieb, J.; de Lumen, B.O. Chemopreventive property of a soybean peptide (lunasin) that binds to deacetylated histones and inhibits acetylation. Cancer Res. 2001, 61, 7473–7478. [Google Scholar]
- Dia, V.P.; de Mejia, E.G. Lunasin promotes apoptosis in human colon cancer cells by mitochondrial pathway activation and induction of nuclear clusterin expression. Cancer Lett. 2010, 295, 44–53. [Google Scholar] [CrossRef]
- Tang, Z.H.; Ding, S.J.; Huang, H.L.; Luo, P.F.; Qing, B.H.; Zhang, S.Y.; Tang, R.T. HDAC1 triggers the proliferation and migration of breast cancer cells via upregulation of interleukin-8. Biol. Chem. 2017, 398, 1347–1356. [Google Scholar] [CrossRef]
- Hartman, Z.C.; Poage, G.M.; den Hollander, P.; Tsimelzon, A.; Hill, J.; Panupinthu, N.; Zhang, Y.; Mazumdar, A.; Hilsenbeck, S.G.; Mills, G.B.; et al. Growth of Triple-Negative Breast Cancer Cells Relies upon Coordinate Autocrine Expression of the Proinflammatory Cytokines IL-6 and IL-8. Cancer Res. 2013, 73, 3470–3480. [Google Scholar] [CrossRef] [Green Version]
- Phuagkhaopong, S.; Ospondpant, D.; Kasemsuk, T.; Sibmooh, N.; Soodvilai, S.; Power, C.; Vivithanaporn, P. Cadmium-induced IL-6 and IL-8 expression and release from astrocytes are mediated by MAPK and NF-kappa B. Neurotoxicology 2017, 60, 82–91. [Google Scholar] [CrossRef]
- Zhu, H.Q.; Shan, L.M.; Schiller, P.W.; Mai, A.; Peng, T.Q. Histone Deacetylase-3 Activation Promotes Tumor Necrosis Factor-alpha (TNF-alpha) Expression in Cardiomyocytes during Lipopolysaccharide Stimulation. J. Biol. Chem. 2010, 285, 9429–9436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wei, M.; Wang, C.; Sun, D.P.; Liu, P.; Zhong, X.; He, Q.S.; Yu, W.B. The histone deacetylase HDAC1 activates HIF1 alpha/VEGFA signal pathway in colorectal cancer. Gene 2020, 754, 144851. [Google Scholar] [CrossRef]
- Liu, X.F.; Yu, Y.; Zhang, J.N.; Lu, C.X.; Wang, L.M.; Liu, P.; Song, H. HDAC1 Silencing in Ovarian Cancer Enhances the Chemotherapy Response. Cell. Physiol. Biochem. 2018, 48, 1505–1518. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.K.; Simoes, B.M.; Howell, S.J.; Farnie, G.; Clarke, R.B. Recent advances reveal IL-8 signaling as a potential key to targeting breast cancer stem cells. Breast Cancer Res. 2013, 15, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.L.; Yang, M.H.; Tsai, M.L.; Lan, H.Y.; Su, S.H.; Chang, S.C.; Teng, H.W.; Yang, S.H.; Lan, Y.T.; Chiou, S.H.; et al. SNAIL Regulates Interleukin-8 Expression, Stem Cell-Like Activity, and Tumorigenicity of Human Colorectal Carcinoma Cells. Gastroenterology 2011, 141, 279–291.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, M.; Greathouse, K.L. Targeting Dietary and Microbial Tryptophan-Indole Metabolism as Therapeutic Approaches to Colon Cancer. Nutrients 2021, 13, 1189. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Martinez-Lopez, A.; Villanueva-Lazo, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.). Antioxidants 2021, 10, 645. [Google Scholar] [CrossRef]
- Jumeri; Kim, S.M. Antioxidant and Anticancer Activities of Enzymatic Hydrolysates of Solitary Tunicate (Styela clava). Food Sci. Biotechnol. 2011, 20, 1075. [Google Scholar] [CrossRef]
- Kannan, A.; Hettiarachchy, N.; Johnson, M.G.; Nannapaneni, R. Human Colon and Liver Cancer Cell Proliferation Inhibition by Peptide Hydrolysates Derived from Heat-Stabilized Defatted Rice Bran. J. Agric. Food Chem. 2008, 56, 11643–11647. [Google Scholar] [CrossRef]
- Gonzalez-Montoya, M.; Hernandez-Ledesma, B.; Silvan, J.M.; Mora-Escobedo, R.; Martinez-Villaluenga, C. Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem. 2018, 242, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Rayaprolu, S.J.; Hettiarachchy, N.S.; Chen, P.Y.; Kannan, A.; Mauromostakos, A. Peptides derived from high oleic acid soybean meals inhibit colon, liver and lung cancer cell growth. Food Res. Int. 2013, 50, 282–288. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and Antioxidant Activities of the Peptide Fraction from Algae Protein Waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Davalos, A.; Bartolome, B.; Amigo, L. Preparation of antioxidant enzymatic hydrolysates from (alpha-lactalbumin and beta-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef]
- Cheung, K.S.; Chan, E.W.; Seto, W.K.; Wong, I.C.K.; Leung, W.K. ACE (Angiotensin-Converting Enzyme) Inhibitors/Angiotensin Receptor Blockers Are Associated With Lower Colorectal Cancer Risk A Territory-Wide Study With Propensity Score Analysis. Hypertension 2020, 76, 968–975. [Google Scholar] [CrossRef]
- Krishna, S.; Lakra, A.D.; Shukla, N.; Khan, S.; Mishra, D.P.; Ahmed, S.; Siddidi, M.I. Identification of potential histone deacetylase1 (HDAC1) inhibitors using multistep virtual screening approach including SVM model, pharmacophore modeling, molecular docking and biological evaluation. J. Biomol. Struct. Dyn. 2020, 38, 3280–3295. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, T.T.; Ren, Y.X.; Li, F.C.; Zheng, G.X.; Hong, J.J.; Yao, X.J.; Xue, W.W.; Zhu, F. Selective Inhibition of HDAC1 by Macrocyclic Polypeptide for the Treatment of Glioblastoma: A Binding Mechanistic Analysis Based on Molecular Dynamics. Front. Mol. Biosci. 2020, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Sun, L.P.; Zhuang, Y.L. Preparation and identification of novel inhibitory angiotensin-I-converting enzyme peptides from tilapia skin gelatin hydrolysates: Inhibition kinetics and molecular docking. Food Funct. 2018, 9, 5251–5259. [Google Scholar] [CrossRef]
- West, A.C.; Johnstone, R.W. New and emerging HDAC inhibitors for cancer treatment. J. Clin. Investig. 2014, 124, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.L.; Zhang, D.; Li, Z.P.; Jiang, H.Q.; Li, J.N.; Ren, R.R.; Gao, X.F.; Li, J.F.; Wang, X.; Wang, W.F.; et al. The fecal microbiota of patients with pancreatic ductal adenocarcinoma and autoimmune pancreatitis characterized by metagenomic sequencing. J. Transl. Med. 2021, 19, 215. [Google Scholar] [CrossRef] [PubMed]
- Hoshimoto, A.; Suzuki, Y.; Katsuno, T.; Nakajima, H.; Saito, Y. Caprylic acid and medium-chain triglycerides inhibit IL-8 gene transcription in Caco-2 cells: Comparison with the potent histone deacetylase inhibitor trichostatin A. Br. J. Pharmacol. 2002, 136, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.W.; Gatza, E.; Hou, G.Q.; Sun, Y.P.; Whitfield, J.; Song, Y.H.; Oravecz-Wilson, K.; Tawara, I.; Dinarello, C.A.; Reddy, P. Histone deacetylase inhibition regulates inflammation and enhances Tregs after allogeneic hematopoietic cell transplantation in humans. Blood 2015, 125, 815–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Qin, P.Y.; Hao, Y.Q.; Guo, H.M.; Blecker, C.; Everaert, N.; Ren, G.X. Overexpression of Soybean-Derived Lunasin in Wheat and Assessment of Its Anti-Proliferative Activity in Colorectal Cancer HT-29 Cells. Int. J. Mol. Sci. 2020, 21, 9594. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Peptide | Ion (m/z) | Calculated Mass | Peptides Score | Toxicity |
|---|---|---|---|---|---|
| 1 | FHPFPR | 799.90 | 799.92 | 0.9745 | Non |
| 2 | NWFPLPR | 929.00 | 929.08 | 0.9722 | Non |
| 3 | PNFHPFPR | 1011.15 | 1011.14 | 0.9631 | Non |
| 4 | NIFRPF | 793.00 | 792.92 | 0.9402 | Non |
| 5 | SLPNFHPFPR | 1211.40 | 1211.38 | 0.9388 | Non |
| 6 | INNIFRPF | 1020.20 | 1020.19 | 0.8666 | Non |
| 7 | SVENWFPLPR | 1244.50 | 1244.40 | 0.8594 | Non |
| 8 | NSLSLPNFHPFPR | 1525.80 | 1525.72 | 0.8383 | Non |
| 9 | NSWGPNWGDHG | 1226.10 | 1226.22 | 0.8118 | Non |
| 10 | HYNPYFPGGA | 1122.20 | 1122.19 | 0.8021 | Non |
| 11 | FGGGTLGHPW | 1028.10 | 1028.12 | 0.7855 | Non |
| 12 | GLESPNYPWPH | 1296.40 | 1296.39 | 0.7791 | Non |
| 13 | HGSLGFLPR | 983.10 | 983.13 | 0.7758 | Non |
| Peptide | Viable Cells (%) at Different Concentrations (g/L) | ||||
|---|---|---|---|---|---|
| 0 | 1 × 10−1 | 5 × 10−1 | 1 | 2 | |
| FHPFPR | 99.96 ± 1.53 | 90.04 ± 1.26 e | 75.46 ± 0.86 f | 51.41 ± 0.80 k | 50.82 ± 0.76 j |
| NWFPLPR | 101.01 ± 1.12 | 92.86 ± 1.10 d | 79.99 ± 0.86 e | 64.02 ± 1.39 j | 63.18 ± 0.83 i |
| PNFHPFPR | 100.97 ± 2.29 | 90.46 ± 1.43 e | 77.26 ± 1.65 f | 65.32 ± 1.45 j | 61.67 ± 1.64 i |
| NIFRPF | 100.75 ± 1.69 | 100.10 ± 1.03 a | 85.11 ± 0.66 d | 70.29 ± 0.56 h | 70.55 ± 2.01 g |
| SLPNFHPFPR | 99.55 ± 0.95 | 97.79 ± 1.14 bc | 87.21 ± 0.32 d | 78.51 ± 0.26 g | 78.00 ± 0.80 f |
| INNIFRPF | 100.40 ± 0.39 | 100.40 ± 1.29 a | 92.05 ± 1.07 c | 80.44 ± 0.68 f | 80.61 ± 0.41 e |
| SVENWFPLPR | 100.42 ± 0.75 | 99.43 ± 1.39 ab | 95.50 ± 3.60 b | 92.14 ± 0.69 c | 91.04 ± 1.53 c |
| NSLSLPNFHPFPR | 99.21 ± 1.60 | 100.79 ± 0.52 a | 92.78 ± 2.20 c | 86.55 ± 0.58 e | 83.56 ± 0.71 d |
| NSWGPNWGDHG | 101.06 ± 1.21 | 98.87 ± 0.60 abc | 98.60 ± 2.28 a | 99.58 ± 1.08 a | 97.92 ± 1.41 a |
| HYNPYFPGGA | 101.54 ± 0.74 | 93.69 ± 0.68 d | 82.34 ± 1.36 e | 67.44 ± 1.26 i | 67.10 ± 1.87 h |
| FGGGTLGHPW | 98.81 ± 1.76 | 97.09 ± 0.74 c | 91.65 ± 0.43 c | 90.41 ± 0.69 d | 89.72 ± 0.60 c |
| GLESPNYPWPH | 100.33 ± 1.34 | 99.75 ± 1.06 a | 99.32 ± 1.05 a | 97.40 ± 0.73 b | 97.70 ± 0.33 ab |
| HGSLGFLPR | 100.26 ± 1.66 | 97.35 ± 1.03 c | 96.91 ± 0.77 ab | 96.81 ± 0.88 b | 95.80 ± 0.62 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Guo, H.; Teng, C.; Zhang, B.; Blecker, C.; Ren, G. Anti-Colon Cancer Activity of Novel Peptides Isolated from In Vitro Digestion of Quinoa Protein in Caco-2 Cells. Foods 2022, 11, 194. https://doi.org/10.3390/foods11020194
Fan X, Guo H, Teng C, Zhang B, Blecker C, Ren G. Anti-Colon Cancer Activity of Novel Peptides Isolated from In Vitro Digestion of Quinoa Protein in Caco-2 Cells. Foods. 2022; 11(2):194. https://doi.org/10.3390/foods11020194
Chicago/Turabian StyleFan, Xin, Huimin Guo, Cong Teng, Biao Zhang, Christophe Blecker, and Guixing Ren. 2022. "Anti-Colon Cancer Activity of Novel Peptides Isolated from In Vitro Digestion of Quinoa Protein in Caco-2 Cells" Foods 11, no. 2: 194. https://doi.org/10.3390/foods11020194
APA StyleFan, X., Guo, H., Teng, C., Zhang, B., Blecker, C., & Ren, G. (2022). Anti-Colon Cancer Activity of Novel Peptides Isolated from In Vitro Digestion of Quinoa Protein in Caco-2 Cells. Foods, 11(2), 194. https://doi.org/10.3390/foods11020194