

Can Bioactive Compounds in Beetroot/Carrot Juice Have a Neuroprotective Effect? Morphological Studies of Neurons Immunoreactive to Calretinin of the Rat Hippocampus after Exposure to Cadmium

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Beetroot/Carrot Juice (BCJ)
2.2. Qualitative Characteristics of Juice
2.3. Animals
2.4. Experimental Procedures and Analyses
2.5. Statistical Analysis
3. Results
3.1. Anticholinesterase Activity of the Studied Juice
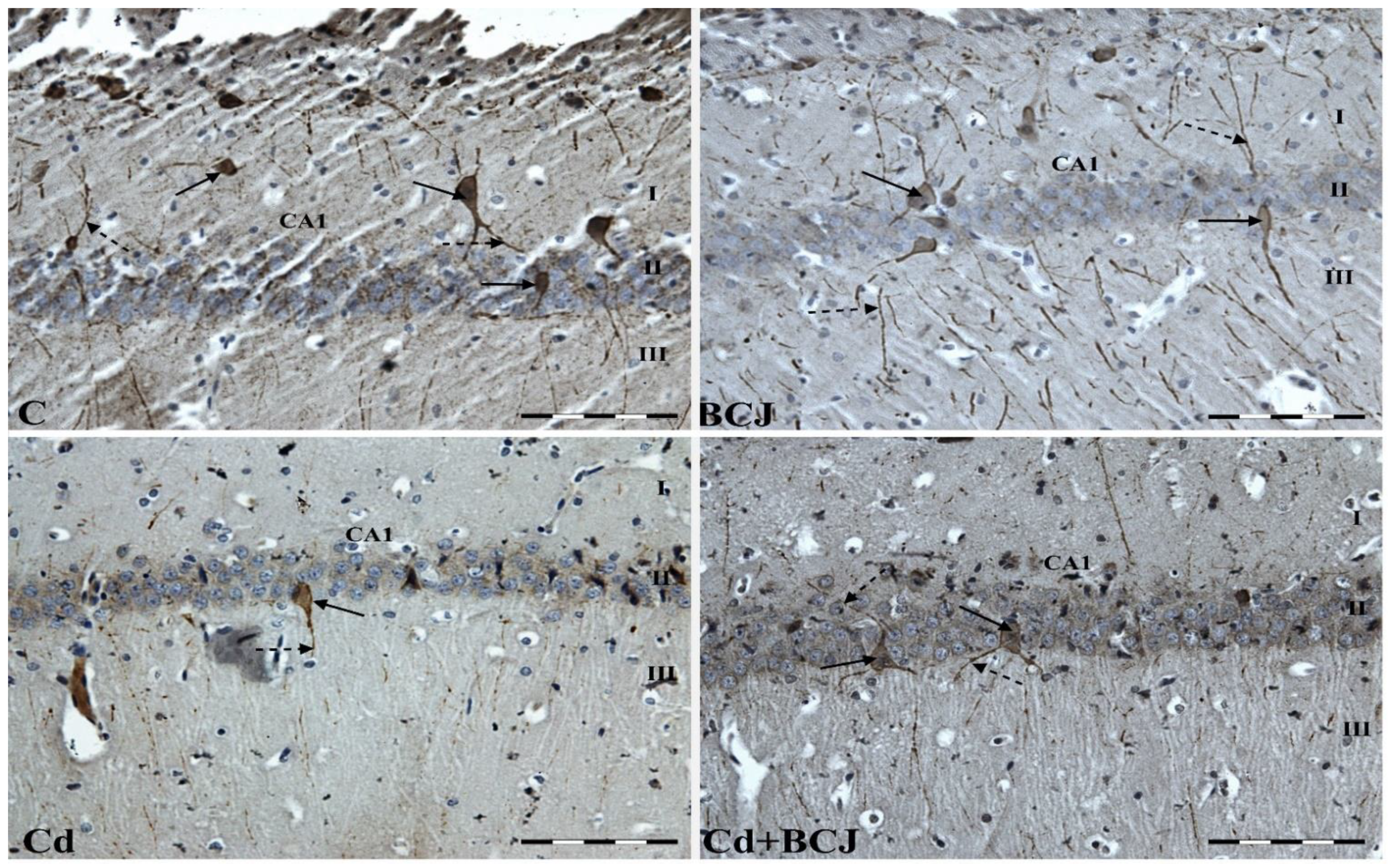
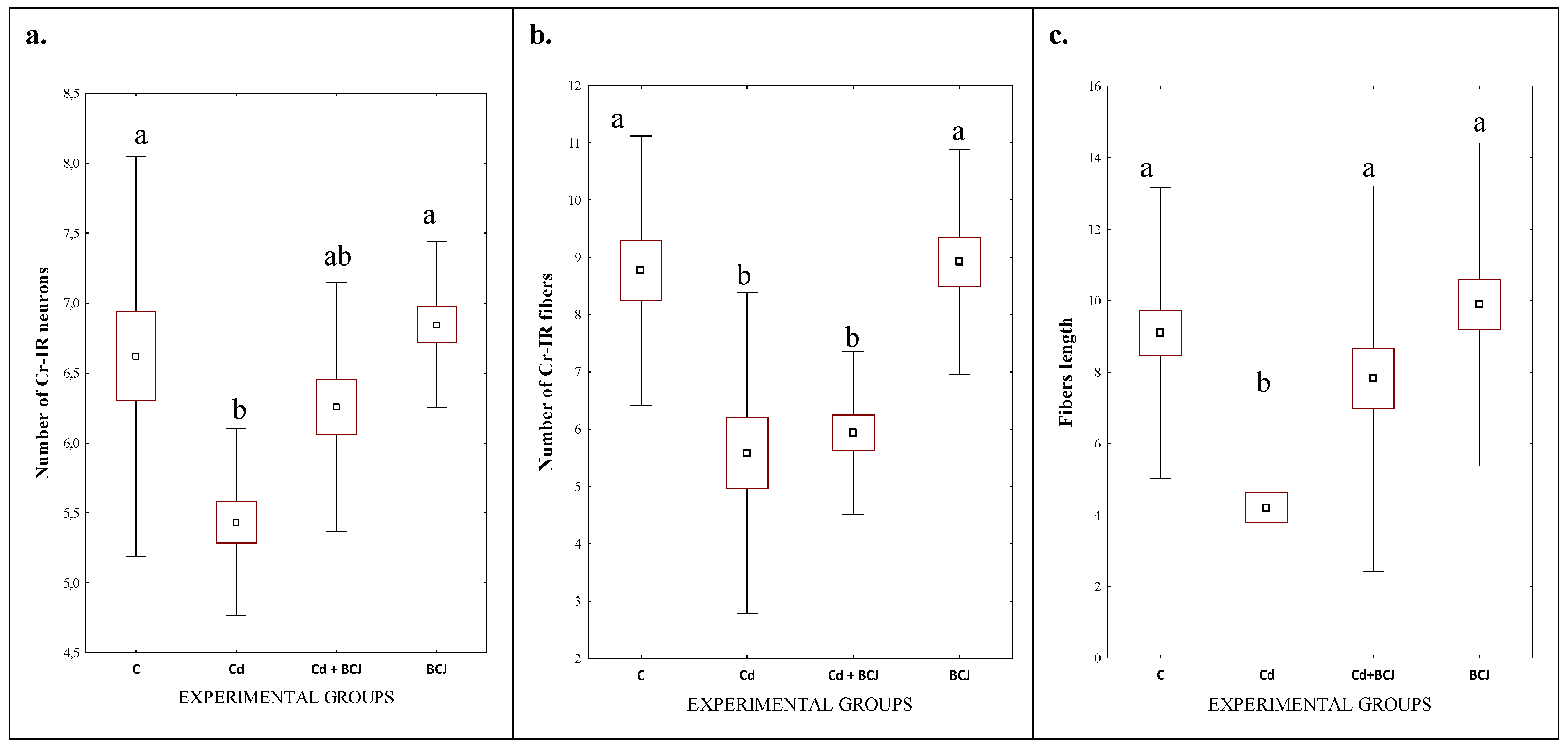
3.2. Cr Expression in the CA1 Field of the Hippocampus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andjelkovic, M.; Buha Djordjevic, A.; Antonijevic, E.; Antonijevic, B.; Stanic, M.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; Jovanovic, M.; Boricic, N.; Wallace, D.; et al. Toxic Effect of Acute Cadmium and Lead Exposure in Rat Blood, Liver, and Kidney. Int. J. Environ. Res. Public. Health 2019, 16, 274. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S. Dietary Cadmium Intake and Its Effects on Kidneys. Toxics 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Huat, T.J.; Camats-Perna, J.; Newcombe, E.A.; Valmas, N.; Kitazawa, M.; Medeiros, R. Metal Toxicity Links to Alzheimer’s Disease and Neuroinflammation. J. Mol. Biol. 2019, 431, 1843–1868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, X.; Zhao, R.; Zhang, R.; Xu, C.; Wang, X.; Liu, C.; Hu, X.; Huang, S.; Chen, L. Cadmium Results in Accumulation of Autophagosomes-Dependent Apoptosis through Activating Akt-Impaired Autophagic Flux in Neuronal Cells. Cell. Signal. 2019, 55, 26–39. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, Z.; Wen, L.; Lei, D.; Li, S.; Wang, J.; Huang, J.; Wang, N.; Durkan, C.; Liao, X.; et al. Cadmium-Induced Dysfunction of the Blood-Brain Barrier Depends on ROS-Mediated Inhibition of PTPase Activity in Zebrafish. J. Hazard. Mater. 2021, 412, 125198. [Google Scholar] [CrossRef]
- Feng, J.; Chen, S.; Wang, Y.; Liu, Q.; Yang, M.; Li, X.; Nie, C.; Qin, J.; Chen, H.; Yuan, X.; et al. Maternal Exposure to Cadmium Impairs Cognitive Development of Male Offspring by Targeting the Coronin-1a Signaling Pathway. Chemosphere 2019, 225, 765–774. [Google Scholar] [CrossRef]
- Forcella, M.; Lau, P.; Oldani, M.; Melchioretto, P.; Bogni, A.; Gribaldo, L.; Fusi, P.; Urani, C. Neuronal Specific and Non-Specific Responses to Cadmium Possibly Involved in Neurodegeneration: A Toxicogenomics Study in a Human Neuronal Cell Model. Neurotoxicology 2020, 76, 162–173. [Google Scholar] [CrossRef]
- Marchetti, C. Role of Calcium Channels in Heavy Metal Toxicity. ISRN Toxicol. 2013, 2013, 184360. [Google Scholar] [CrossRef]
- Kovács-Öller, T.; Szarka, G.; Ganczer, A.; Tengölics, Á.; Balogh, B.; Völgyi, B. Expression of Ca2+-Binding Buffer Proteins in the Human and Mouse Retinal Neurons. Int. J. Mol. Sci. 2019, 20, 2229. [Google Scholar] [CrossRef]
- Saffari, R.; Grotefeld, K.; Kravchenko, M.; Zhang, M.; Zhang, W. Calretinin+-Neurons-Mediated GABAergic Inhibition in Mouse Prefrontal Cortex. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 94, 109658. [Google Scholar] [CrossRef]
- Deseva, I.; Stoyanova, M.; Petkova, N.; Mihaylova, D. Red Beetroot Juice Phytochemicals Bioaccessibility: An In Vitro Approach. Pol. J. Food Nutr. Sci. 2020, 70, 45–53. [Google Scholar] [CrossRef]
- Shaban, N.Z.; Abd El-Kader, S.E.; Mogahed, F.A.K.; El-Kersh, M.A.L.; Habashy, N.H. Synergistic Protective Effect of Beta Vulgaris with Meso-2,3-Dimercaptosuccinic Acid against Lead-Induced Neurotoxicity in Male Rats. Sci. Rep. 2021, 11, 252. [Google Scholar] [CrossRef]
- Wulandari, D.D.; Santoso, A.P.R.; Wulansari, D.D. The Effect of Beetroot (Beta vulgaris L.) Juiceon Cholinesterase Activity in Farmers Exposed to Ogranophosphate Pesticides. Indones. J. Med. Lab. Sci. Technol. 2019, 1, 80–87. [Google Scholar] [CrossRef]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive Compounds of Beetroot and Utilization in Food Processing Industry: A Critical Review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Liliana, C.; Oana-Viorela, N. Red Beetroot: Composition and Health Effects—A Review. J. Nutr. Med. Diet Care 2020, 6, 43. [Google Scholar] [CrossRef]
- Wightman, E.L.; Haskell-Ramsay, C.F.; Thompson, K.G.; Blackwell, J.R.; Winyard, P.G.; Forster, J.; Jones, A.M.; Kennedy, D.O. Dietary Nitrate Modulates Cerebral Blood Flow Parameters and Cognitive Performance in Humans: A Double-Blind, Placebo-Controlled, Crossover Investigation. Physiol. Behav. 2015, 149, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Sulakhiya, K.; Patel, V.; Saxena, R.; Dashore, J.; Srivastava, A.; Rathore, M. Effect of Beta Vulgaris Linn. Leaves Extract on Anxiety- and Depressive-like Behavior and Oxidative Stress in Mice after Acute Restraint Stress. Pharmacogn. Res. 2016, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Cawood, M.; Iqbal, Q.; Ariño, A.; Batool, A.; Tariq, R.M.S.; Azam, M.; Akhtar, S. Phytochemicals in Daucus carota and Their Health Benefits—Review Article. Foods 2019, 8, 424. [Google Scholar] [CrossRef]
- Aćimović, M.G. Nutraceutical potential of Apiaceae. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Reference Series in Phytochemistry; Springer International Publishing: Cham, Swizerland, 2019; pp. 1311–1341. [Google Scholar] [CrossRef]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of Many Colors Provide Basic Nutrition and Bioavailable Phytochemicals Acting as a Functional Food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Mazewski, C.; Liang, K.; Gonzalez de Mejia, E. Comparison of the Effect of Chemical Composition of Anthocyanin-Rich Plant Extracts on Colon Cancer Cell Proliferation and Their Potential Mechanism of Action Using in Vitro, in Silico, and Biochemical Assays. Food Chem. 2018, 242, 378–388. [Google Scholar] [CrossRef]
- Szwajgier, D.; Baranowska-Wójcik, E.; Sosnowska, B.; Kukula-Koch, W.; Paduch, R.; Sokołowska, B.; Waśko, A.; Solarska, E. High-Pressure Processing at Increased Temperatures Provides Enhanced Removal of Indigenous Microbial Contamination in Beet/Carrot Juice without Damaging the Bioactive Components. Int. J. Food Sci. Technol. 2022, 57, 276–289. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Żarowski, M.; Plech, T.; Cielecka-Piontek, J. Permeability of Hypogymnia Physodes Extract Component—Physodic Acid through the Blood–Brain Barrier as an Important Argument for Its Anticancer and Neuroprotective Activity within the Central Nervous System. Cancers 2021, 13, 1717. [Google Scholar] [CrossRef] [PubMed]
- Buha, A.; Bulat, Z.; Đukić-Ćosić, D.; Matović, V. Effects of Oral and Intraperitoneal Magnesium Treatment against Cadmium-Induced Oxidative Stress in Plasma of Rats. Arch. Ind. Hyg. Toxicol. 2012, 63, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Elkhadragy, M.F.; Kassab, R.B.; Metwally, D.; Almeer, R.S.; Abdel-Gaber, R.; Al-Olayan, E.M.; Essawy, E.A.; Amin, H.K.; Abdel Moneim, A.E. Protective Effects of Fragaria Ananassa Methanolic Extract in a Rat Model of Cadmium Chloride-Induced Neurotoxicity. Biosci. Rep. 2018, 38, BSR20180861. [Google Scholar] [CrossRef]
- Yuan, G.; Dai, S.; Yin, Z.; Lu, H.; Jia, R.; Xu, J.; Song, X.; Li, L.; Shu, Y.; Zhao, X. Toxicological Assessment of Combined Lead and Cadmium: Acute and Sub-Chronic Toxicity Study in Rats. Food Chem. Toxicol. 2014, 65, 260–268. [Google Scholar] [CrossRef]
- Matysek, M.; Borowiec, K.; Szwajgier, D.; Szalak, R.; Arciszewski, M. Insulin Receptors in the CA1 Field of Hippocampus and Selected Blood Parameters in Diabetic Rats Fed with Bilberry Fruit. Ann. Agric. Environ. Med. 2021, 28, 430–436. [Google Scholar] [CrossRef]
- Cordner, Z.A.; Khambadkone, S.G.; Boersma, G.J.; Song, L.; Summers, T.N.; Moran, T.H.; Tamashiro, K.L.K. Maternal High-Fat Diet Results in Cognitive Impairment and Hippocampal Gene Expression Changes in Rat Offspring. Exp. Neurol. 2019, 318, 92–100. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L.; Abel, G.M.; Storm, D.R.; Xia, Z. Cadmium Exposure Impairs Cognition and Olfactory Memory in Male C57BL/6 Mice. Toxicol. Sci. 2018, 161, 87–102. [Google Scholar] [CrossRef]
- Almazan, G.; Liu, H.-N.; Khorchid, A.; Sundararajan, S.; Martinez-Bermudez, A.K.; Chemtob, S. Exposure of Developing Oligodendrocytes to Cadmium Causes HSP72 Induction, Free Radical Generation, Reduction in Glutathione Levels, and Cell Death. Free Radic. Biol. Med. 2000, 29, 858–869. [Google Scholar] [CrossRef]
- López, E.; Figueroa, S.; Oset-Gasque, M.J.; González, M.P. Apoptosis and Necrosis: Two Distinct Events Induced by Cadmium in Cortical Neurons in Culture: Cadmium Induces Apoptosis and Necrosis in Neurons. Br. J. Pharmacol. 2003, 138, 901–911. [Google Scholar] [CrossRef]
- Presley, T.D.; Morgan, A.R.; Bechtold, E.; Clodfelter, W.; Dove, R.W.; Jennings, J.M.; Kraft, R.A.; Bruce King, S.; Laurienti, P.J.; Jack Rejeski, W.; et al. Acute Effect of a High Nitrate Diet on Brain Perfusion in Older Adults. Nitric Oxide 2011, 24, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Oyeleye, S.I.; Ibeji, C.U.; Oboh, G. Beetroot Supplemented Diet Exhibit Anti-Amnesic Effect via Modulation of Cholinesterases, Purinergic Enzymes, Monoamine Oxidase and Attenuation of Redox Imbalance in the Brain of Scopolamine Treated Male Rats. Nutr. Neurosci. 2022, 25, 1011–1025. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Shah, S.; Mehmood Butt, A.; Masood Shah, S.; Jabeen, Z.; Nadeem, A. Biochemical Profiling and Elucidation of Biological Activities of Beta Vulgaris L. Leaves and Roots Extracts. Saudi J. Biol. Sci. 2021, 28, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Aliahmadi, M.; Amiri, F.; Bahrami, L.S.; Hosseini, A.F.; Abiri, B.; Vafa, M. Effects of Raw Red Beetroot Consumption on Metabolic Markers and Cognitive Function in Type 2 Diabetes Patients. J. Diabetes Metab. Disord. 2021, 20, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Schwaller, B.; Meyer, M.; Schiffmann, S. “New” Functions for “Old” Proteins: The Role of the Calcium-Binding Proteins Calbindin D-28k, Calretinin and Parvalbumin, in Cerebellar Physiology. Studies with Knockout Mice. Cerebellum 2002, 1, 241–258. [Google Scholar] [CrossRef]
- Xu, B.; Chen, S.; Luo, Y.; Chen, Z.; Liu, L.; Zhou, H.; Chen, W.; Shen, T.; Han, X.; Chen, L.; et al. Calcium Signaling Is Involved in Cadmium-Induced Neuronal Apoptosis via Induction of Reactive Oxygen Species and Activation of MAPK/MTOR Network. PLoS ONE 2011, 6, e19052. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, C.; Xu, H.; Sun, Y.; Hu, F.; Bian, J.; Liu, X.; Gu, J.; Liu, Z. Cadmium-Induced Apoptosis in Primary Rat Cerebral Cortical Neurons Culture Is Mediated by a Calcium Signaling Pathway. PLoS ONE 2013, 8, e64330. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | Cd Exposition | BCJ Supplementation | Two-Way ANOVA | ||||
|---|---|---|---|---|---|---|---|
| Impact of | |||||||
| − | + | − | + | Cd Exposition | BCJ Treatment | Cd × BCJ | |
| No of Cr-IR neurons | 6.73 a ± 0.53 | 5.85 b ± 0.57 | 6.03 B ± 0.82 | 6.55 A ± 0.47 | <0.001 | 0.025 | 0.178 |
| No of Cr-IR fibers | 8.85 a ± 1.02 | 5.76 b ± 1.06 | 7.18 ± 2.08 | 7.43 ± 1.77 | <0.00001 | 0.620 | 0.835 |
| Length of nerve fibers | 9.50 a ± 2.13 | 6.01 b ± 2.78 | 6.65 b ± 3.02 | 8.86 a ± 2.64 | <0.0001 | 0.002 | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matysek, M.; Kowalczuk-Vasilev, E.; Szalak, R.; Baranowska-Wójcik, E.; Arciszewski, M.B.; Szwajgier, D. Can Bioactive Compounds in Beetroot/Carrot Juice Have a Neuroprotective Effect? Morphological Studies of Neurons Immunoreactive to Calretinin of the Rat Hippocampus after Exposure to Cadmium. Foods 2022, 11, 2794. https://doi.org/10.3390/foods11182794
Matysek M, Kowalczuk-Vasilev E, Szalak R, Baranowska-Wójcik E, Arciszewski MB, Szwajgier D. Can Bioactive Compounds in Beetroot/Carrot Juice Have a Neuroprotective Effect? Morphological Studies of Neurons Immunoreactive to Calretinin of the Rat Hippocampus after Exposure to Cadmium. Foods. 2022; 11(18):2794. https://doi.org/10.3390/foods11182794
Chicago/Turabian StyleMatysek, Małgorzata, Edyta Kowalczuk-Vasilev, Radosław Szalak, Ewa Baranowska-Wójcik, Marcin B. Arciszewski, and Dominik Szwajgier. 2022. "Can Bioactive Compounds in Beetroot/Carrot Juice Have a Neuroprotective Effect? Morphological Studies of Neurons Immunoreactive to Calretinin of the Rat Hippocampus after Exposure to Cadmium" Foods 11, no. 18: 2794. https://doi.org/10.3390/foods11182794
APA StyleMatysek, M., Kowalczuk-Vasilev, E., Szalak, R., Baranowska-Wójcik, E., Arciszewski, M. B., & Szwajgier, D. (2022). Can Bioactive Compounds in Beetroot/Carrot Juice Have a Neuroprotective Effect? Morphological Studies of Neurons Immunoreactive to Calretinin of the Rat Hippocampus after Exposure to Cadmium. Foods, 11(18), 2794. https://doi.org/10.3390/foods11182794