
Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Fucoidan from the A. nodosum Preparation
2.3. Chemical Analysis
2.4. Monosaccharide Composition Analysis
2.5. Fourier-Transform Infrared (FT-IR) Spectroscopic Analysis of ANP-6 and ANP-7
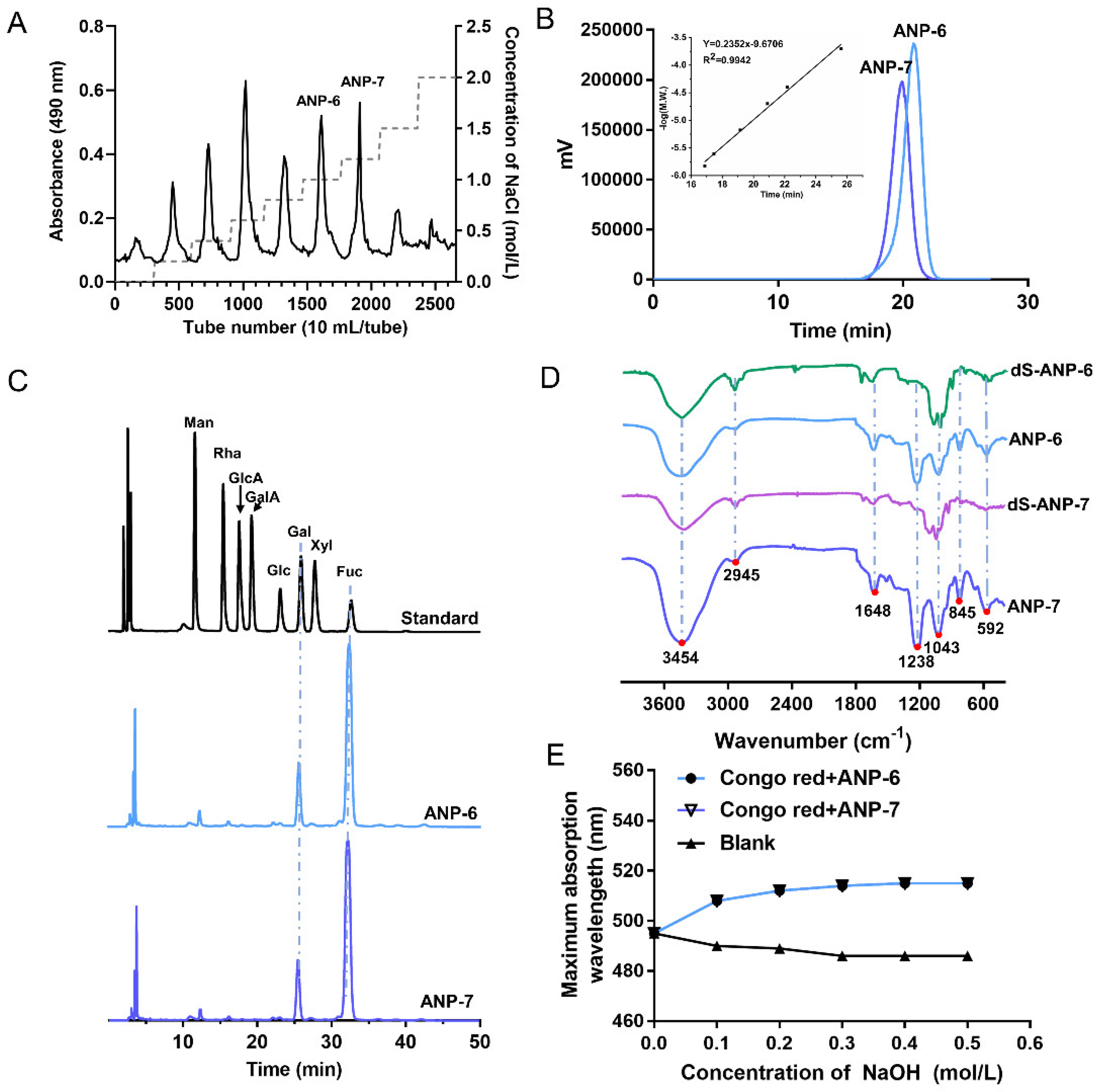
2.6. Congo Red Test of ANP-6 and ANP-7
2.7. Desulfation, Methylation, and GC-MS Analysis
2.8. NMR Spectroscopy Analysis
2.9. Cell Culture
2.10. Cell Viability Assay
2.11. Nitric Oxide (NO) Production Determination
2.12. Real-Time PCR Analysis
2.13. Statistical Analysis
3. Results and Discussions
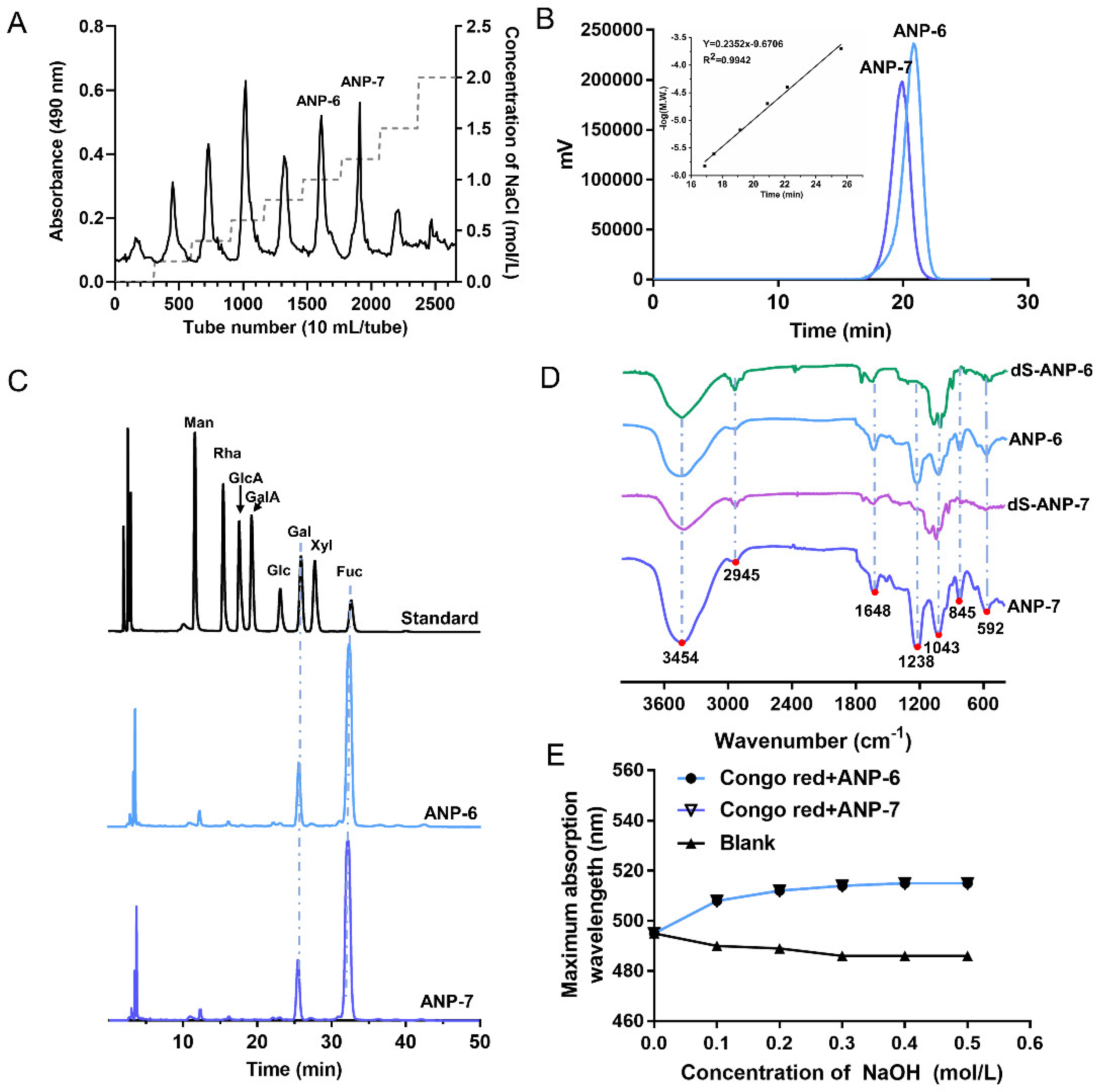
3.1. Physicochemical Properties and the Monosaccharide Composition of ANP-6 and ANP-7
3.2. FT-IR Spectroscopy Analysis of ANP-6 and ANP-7
3.3. The Helix-Coil Transition of ANP-6 and ANP-7
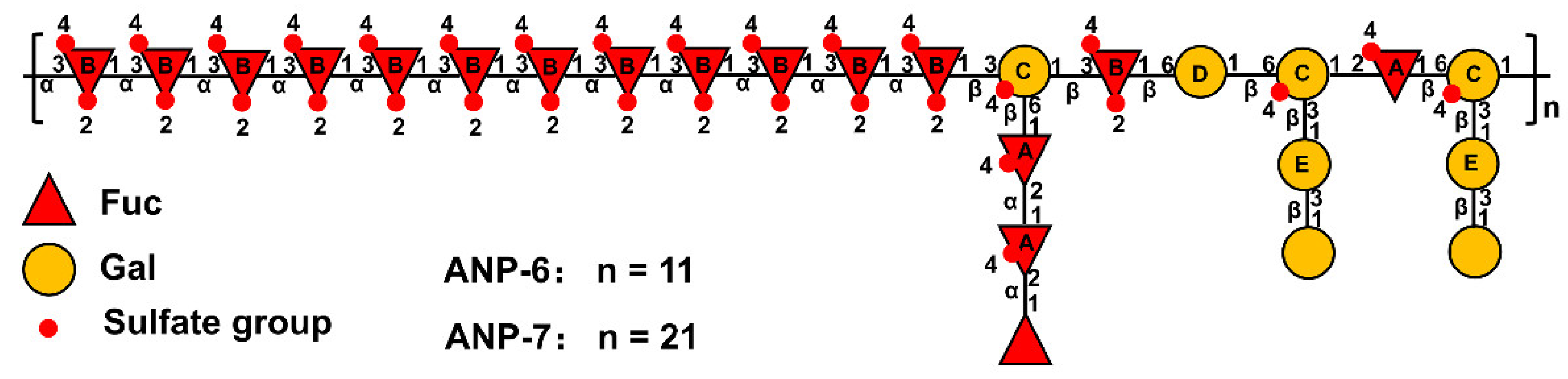
3.4. Methylation Analysis of dS-ANP-6 and dS-ANP-7
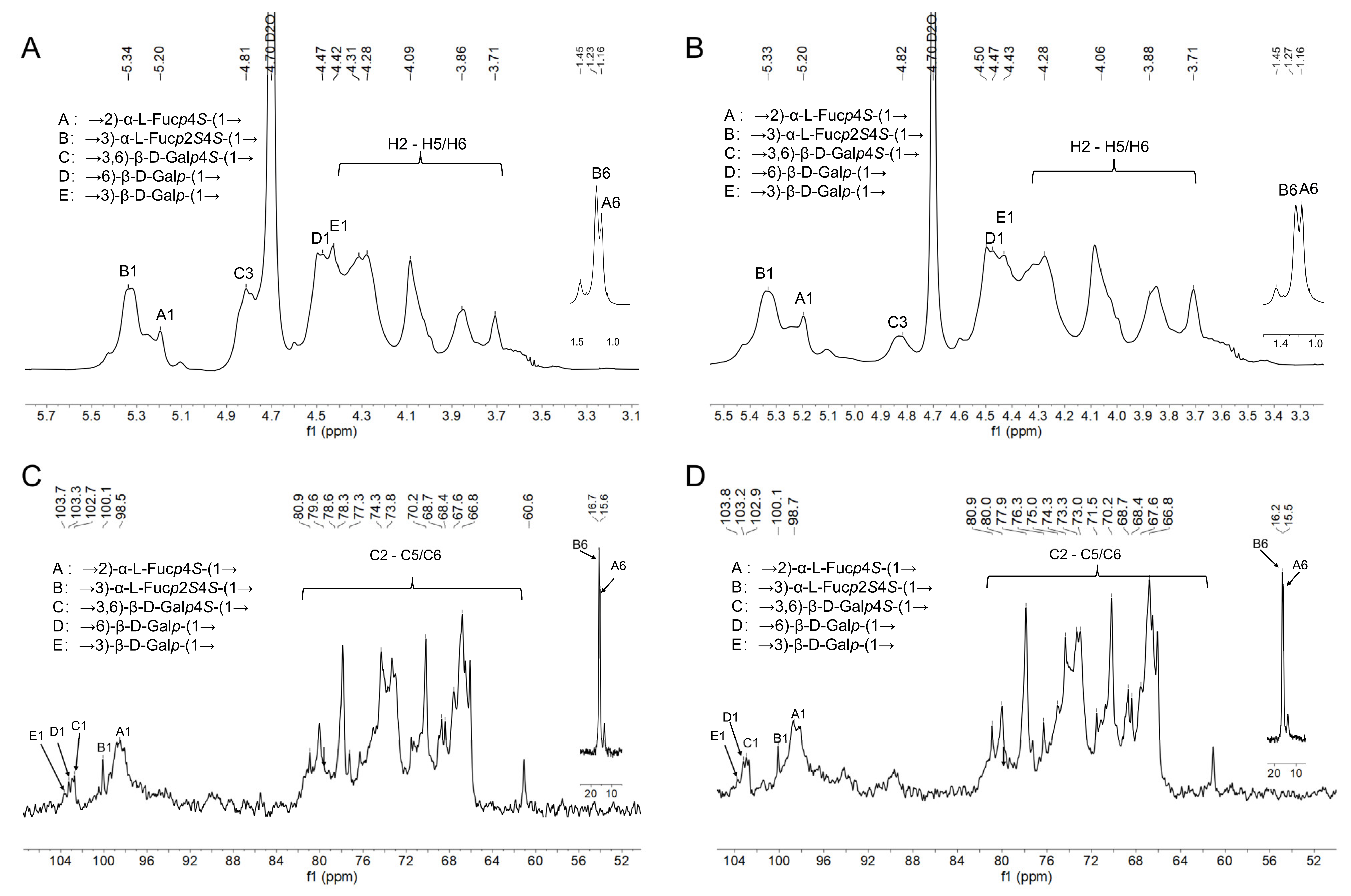
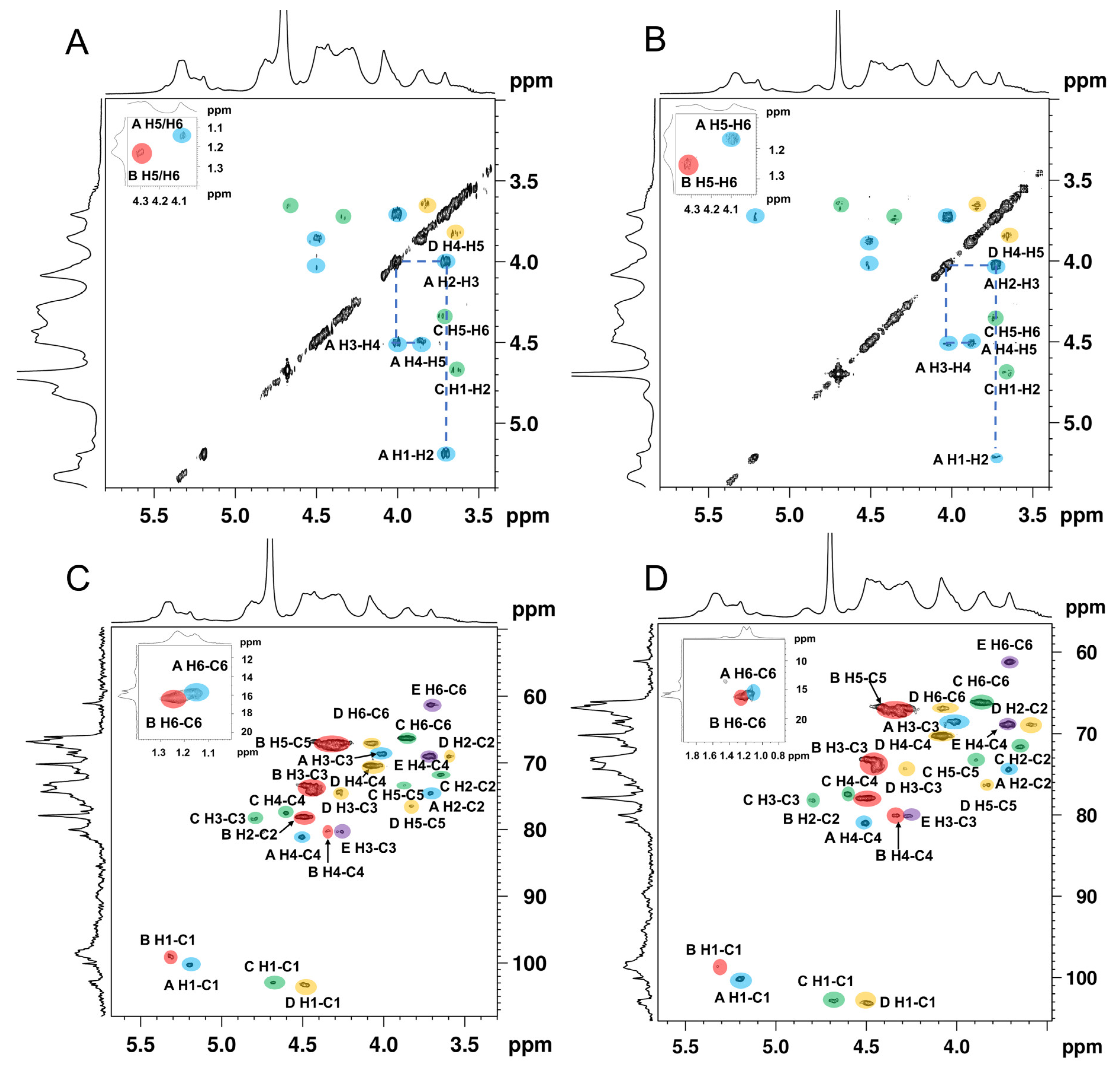
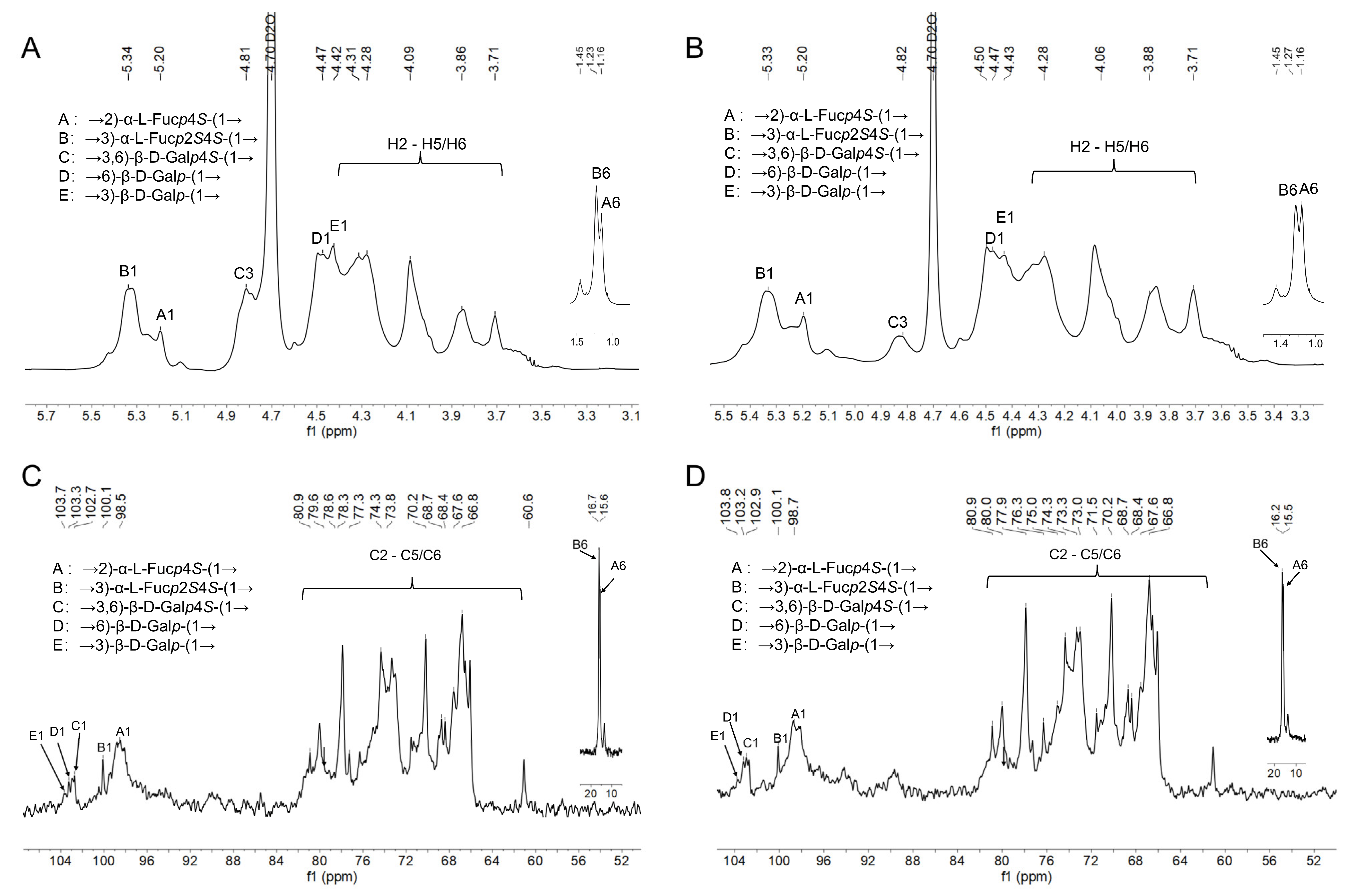
3.5. NMR Spectroscopy Analysis of ANP-6 and ANP-7
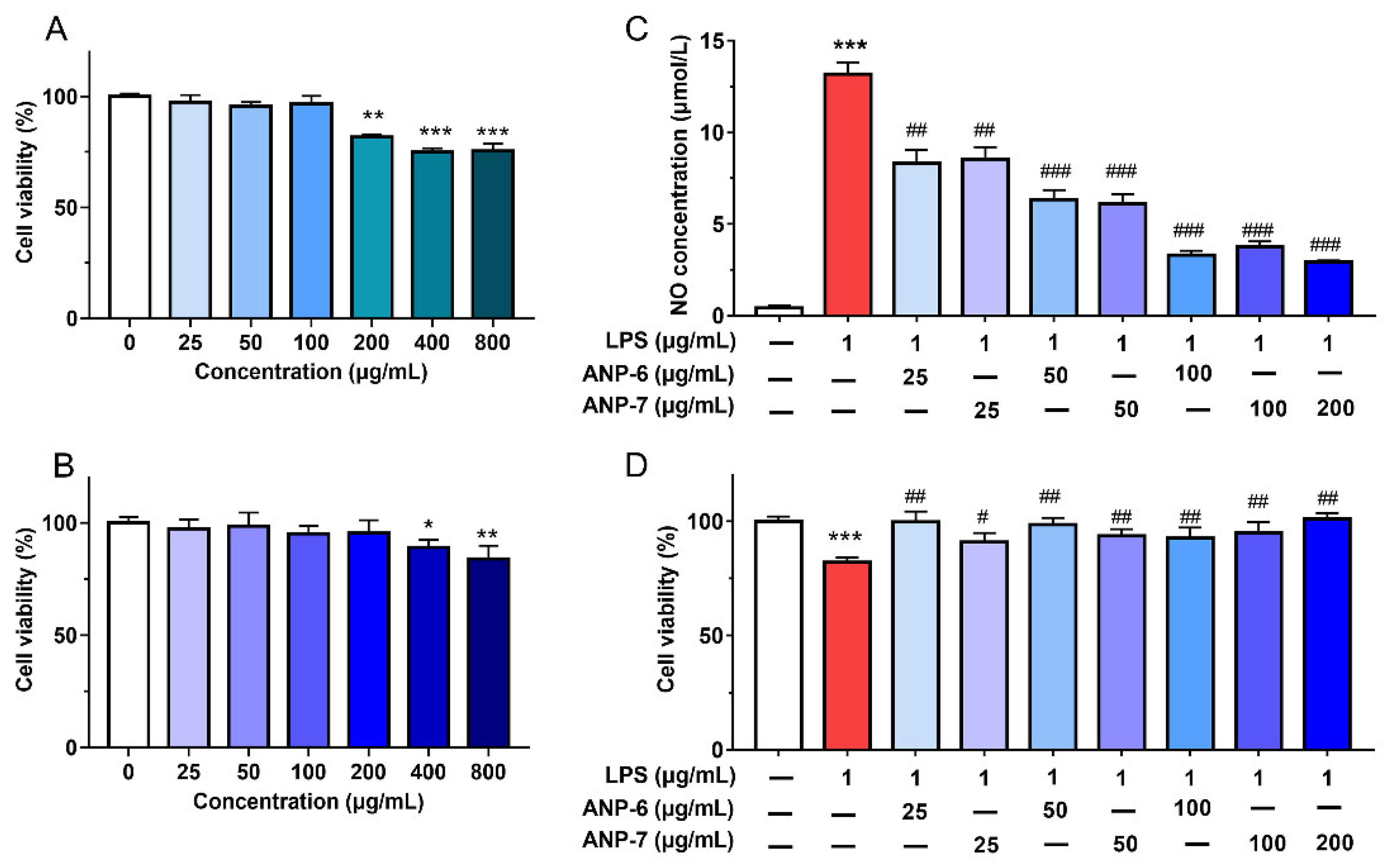
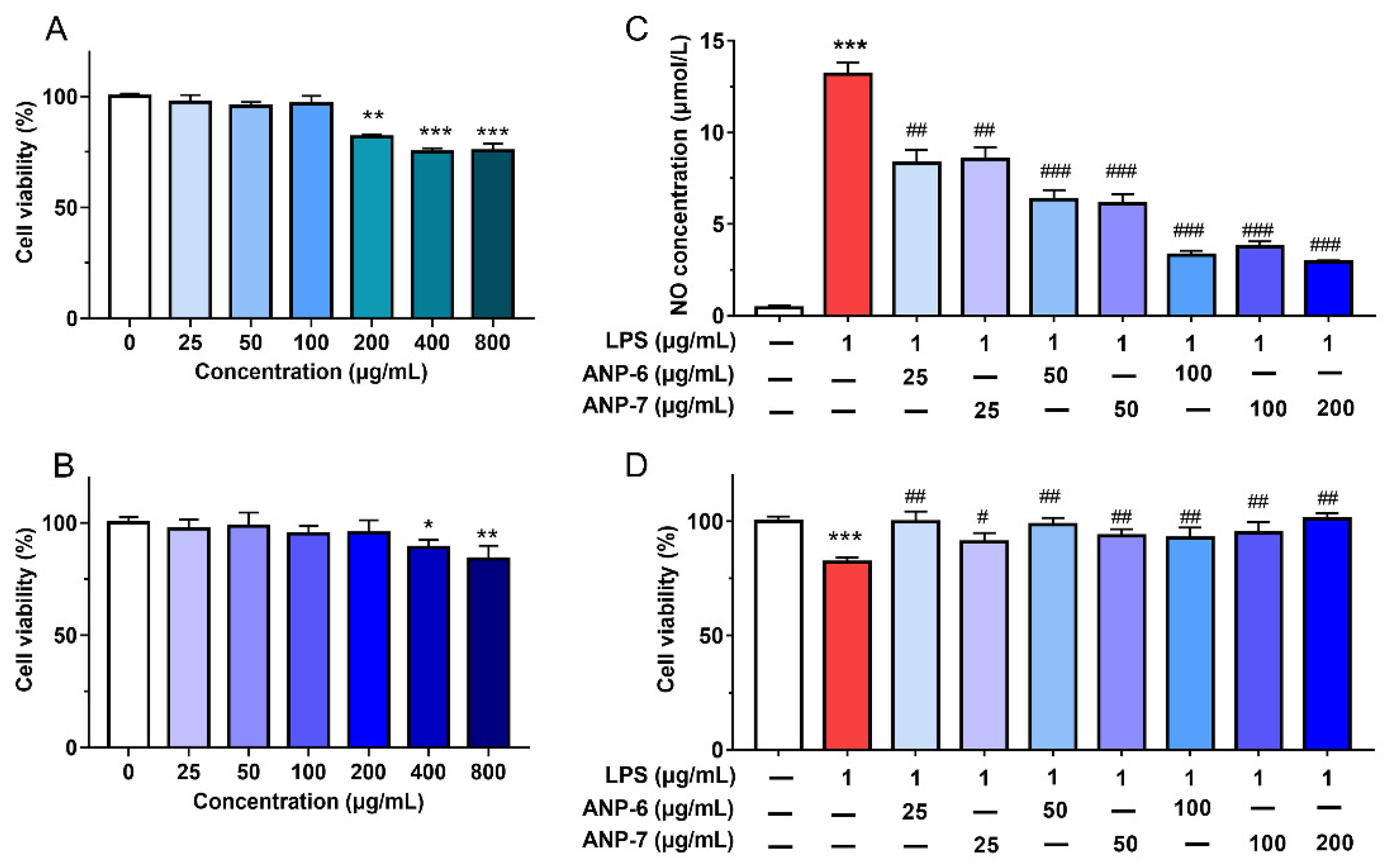
3.6. Effects of ANP-6 and ANP-7 on Cell Viability and NO Production
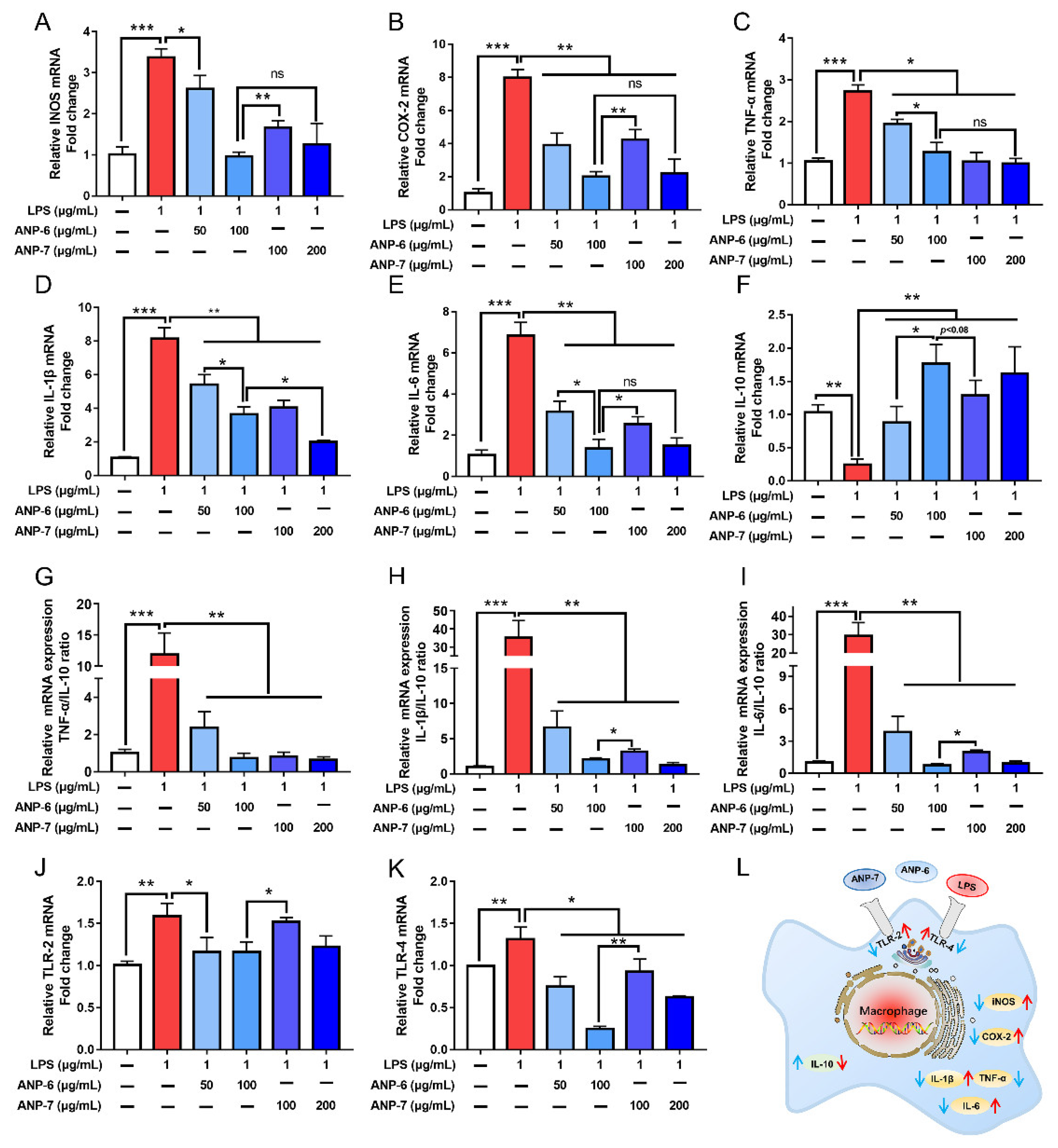
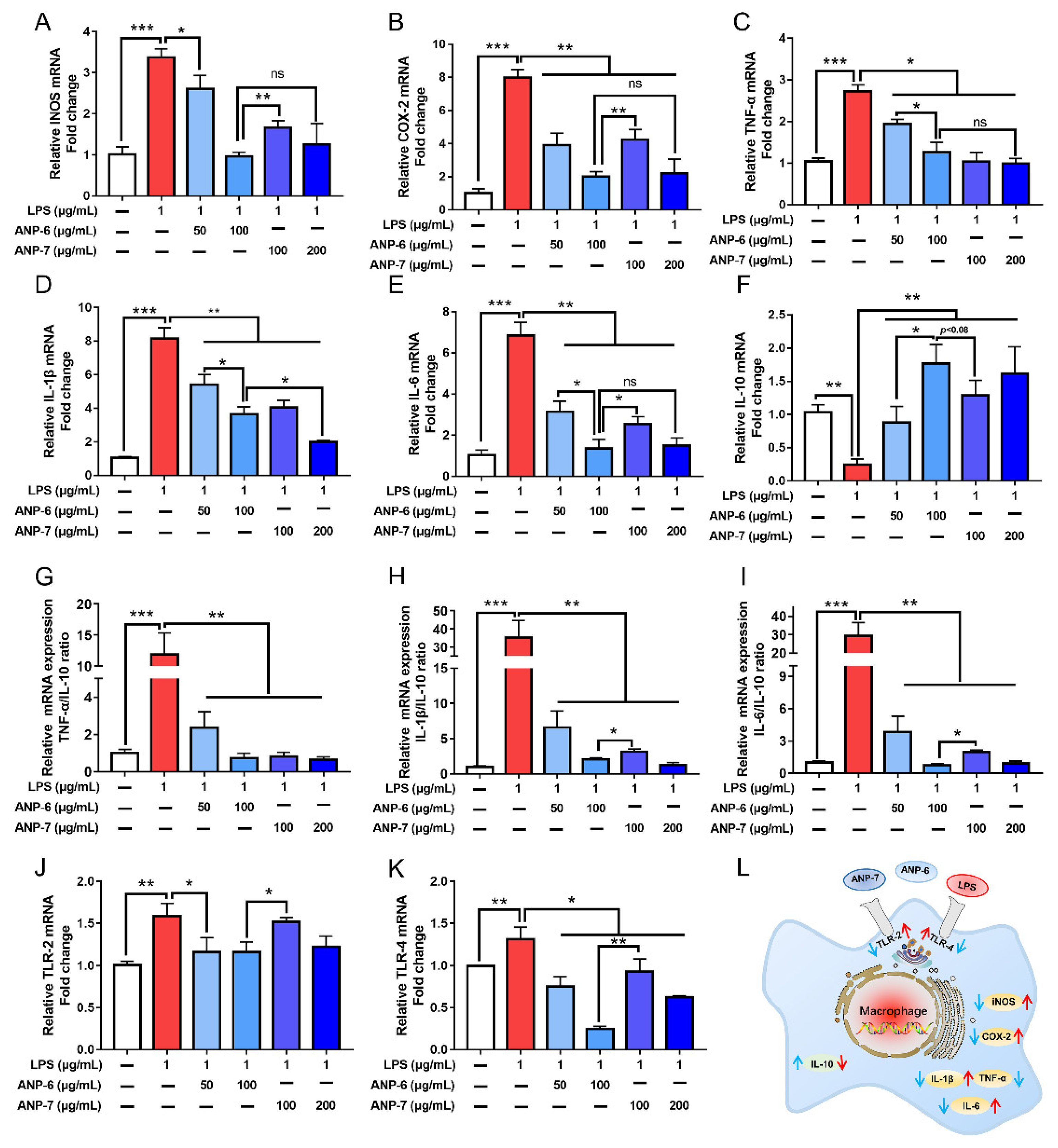
3.7. Effects of ANP-6 and ANP-7 on iNOS and COX-2 mRNA Expressions
3.8. Effects of ANP-6 and ANP-7 on the Inflammatory Cytokine mRNA Expression
3.9. Effects of ANP-6 and ANP-7 on the TLR-2 and TLR-4 mRNA Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jayawardena, T.U.; Sanjeewa, K.K.A.; Nagahawatta, D.P.; Lee, H.G.; Lu, Y.A.; Vaas, A.; Abeytunga, D.T.U.; Nanayakkara, C.M.; Lee, D.S.; Jeon, Y.J. Anti-inflammatory effects of sulfated polysaccharide from Sargassum swartzii in macrophages via blocking TLR/NF-Kappa b signal transduction. Mar. Drugs 2020, 18, 601. [Google Scholar] [CrossRef] [PubMed]
- Janssens, S.; Beyaert, R. Role of Toll-like receptors in pathogen recognition. Clin. Microbiol. Rev. 2003, 16, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, H.S.; Kim, S.Y.; Fernando, I.P.S.; Wang, L.; Abetunga, D.T.U.; Kim, W.S.; Lee, D.S.; Jeon, Y.J. Fucoidan isolated from Padina commersonii inhibit LPS-induced inflammation in macrophages blocking TLR/NF-kappa B signal pathway. Carbohydr. Polym. 2019, 224, 115195. [Google Scholar] [CrossRef] [PubMed]
- Terzi, M.; Altun, G.; Sen, S.; Kocaman, A.; Kaplan, A.A.; Yurt, K.K.; Kaplan, S. The use of non-steroidal anti-inflammatory drugs diseases. J. Chem. Neuroanat. 2018, 87, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Jia, X.; Wang, N.; Xiao, M.; Song, S.; Wu, S.; Li, Z.; Wang, S.; Cui, S.W.; Guo, Q. Insights into the structure-bioactivity relationships of marine sulfated polysaccharides: A review. Food Hydrocoll. 2022, 123, 107049. [Google Scholar] [CrossRef]
- Yuan, Y.; Macquarrie, D. Microwave assisted extraction of sulfated polysaccharides (fucoidan) from Ascophyllum nodosum and its antioxidant activity. Carbohydr. Polym. 2015, 129, 101–107. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and anti-inflammatory effects of fucoidan: A review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Ni, L.Y.; Wang, L.; Fu, X.T.; Duan, D.L.; Jeon, Y.J.; Xu, J.C.; Gao, X. In vitro and in vivo anti-inflammatory activities of a fucose-rich fucoidan isolated from Saccharina japonica. Int. J. Biol. Macromol. 2020, 156, 717–729. [Google Scholar] [CrossRef]
- Wu, N.; Li, Z.; Wang, J.; Geng, L.; Yue, Y.; Deng, Z.; Wang, Q.; Zhang, Q. Low molecular weight fucoidan attenuating pulmonary fibrosis by relieving inflammatory reaction and progression of epithelial-mesenchymal transition. Carbohydr. Polym. 2021, 273, 118567. [Google Scholar] [CrossRef]
- Wu, G.J.; Shiu, S.M.; Hsieh, M.C.; Tsai, G.J. Anti-inflammatory activity of a sulfated polysaccharide from the brown alga Sargassum cristaefolium. Food Hydrocoll. 2016, 53, 16–23. [Google Scholar] [CrossRef]
- Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-inflammatory activity of fucoidan extracts in vitro. Mar. Drugs 2021, 19, 702. [Google Scholar] [CrossRef]
- Phull, A.R.; Kim, S.J. Fucoidan as bio-functional molecule: Insights into the anti-inflammatory potential and associated molecular mechanisms. J. Funct. Food. 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on alpha-amylase, alpha-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ai, C.; Wen, C.; Qin, Y.; Liu, Z.; Wang, L.; Gong, Y.; Su, C.; Wang, Z.; Song, S. Fucoidan isolated from Ascophyllum nodosum alleviates gut microbiota dysbiosis and colonic inflammation in antibiotic-treated mice. Food Funct. 2020, 11, 5595–5606. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, S.; Ai, C.; Wen, C.; Liu, Z.; Wang, L.; Jiang, L.; Shen, P.; Zhang, G.; Song, S. Galactofucan from Laminaria japonica is not degraded by the human digestive system but inhibits pancreatic lipase and modifies the intestinal microbiota. Int. J. Biol. Macromol. 2021, 166, 611–620. [Google Scholar] [CrossRef]
- Filisetticozzi, T.; Carpita, N.C. Measurement of uronic-acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utillizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Imbs, T.I.; Skriptsova, A.V.; Zvyagintseva, T.N. Antioxidant activity of fucose-containing sulfated polysaccharides obtained from Fucus evanescens by different extraction methods. J. Appl. Phycol. 2015, 27, 545–553. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on determination of ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhao, J.; Li, D.; Wen, C.; Liu, H.; Song, S.; Zhu, B. Comparison of polysaccharides of Haliotis discus hannai and Volutharpa ampullacea perryi by PMP-HPLC-MSn analysis upon acid hydrolysis. Carbohydr. Res. 2015, 415, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Mao, G.; Feng, W.; Mao, R.; Gu, X.; Li, T.; Li, Q.; Bao, Y.; Yang, L.; Wu, X. Isolation, characterization and antioxidant activity of polysaccharide from Schisandra sphenanthera. Carbohydr. Polym. 2014, 105, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Zhang, B.; Wu, S.F.; Huang, L.; Ai, C.Q.; Pan, J.F.; Su, Y.C.; Wang, Z.F.; Wen, C.R. Structural characterization and osteogenic bioactivity of a sulfated polysaccharide from pacific abalone (Haliotis discus hannai Ino). Carbohydr. Polym. 2018, 182, 207–214. [Google Scholar] [CrossRef]
- Needs, P.W.; Selvendran, R.R. Avoiding oxidative-degradation during sodium-hydroxide methyl iodide-mediated carbohydrate methylation in dimethyl sulfoxide. Carbohydr. Res. 1993, 245, 1–10. [Google Scholar] [CrossRef]
- Yin, J.Y.; Wang, J.; Li, F.H.; Yang, Z.X.; Yang, X.Q.; Sun, W.L.; Xia, B.; Li, T.; Song, W.G.; Guo, S.D. The fucoidan from the brown seaweed Ascophyllum nodosum ameliorates atherosclerosis in apolipoprotein E-deficient mice. Food Funct. 2019, 10, 5124–5139. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of Lactobacillus and Ruminococcaceae. Food Funct. 2016, 7, 3224–3232. [Google Scholar] [CrossRef]
- Arab, K.; Ghanbarzadeh, B.; Ayaseh, A.; Jahanbin, K. Extraction, purification, physicochemical properties and antioxidant activity of a new polysaccharide from Ocimum album L. seed. Int. J. Biol. Macromol. 2021, 180, 643–653. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, W.; Gao, Y.; Teng, X.; Zhu, W.; Chen, Y.; Zhao, C.; Li, N.; Wang, C.; Yan, M.; et al. Structural elucidation of an extracellular polysaccharide produced by the marine fungus Aspergillus versicolor. Carbohydr. Polym. 2013, 93, 478–483. [Google Scholar] [CrossRef]
- Chen, G.J.; Bai, Y.X.; Zeng, Z.Q.; Peng, Y.J.; Zhou, W.T.; Shen, W.B.; Zeng, X.X.; Liu, Z.H. Structural characterization and immunostimulatory activity of heteropolysaccharides from fuzhuan brick tea. J. Agric. Food Chem. 2021, 69, 1368–1378. [Google Scholar] [CrossRef]
- Li, X.M.; Li, S.N.; Liu, J.; Lin, L.S.; Sun, H.F.; Yang, W.J.; Cai, Y.; Gao, N.; Zhou, L.T.; Qin, H.B.; et al. A regular fucan sulfate from Stichopus herrmanni and its peroxide depolymerization: Structure and anticoagulant activity. Carbohydr. Polym. 2021, 256, 117513. [Google Scholar] [CrossRef]
- Gao, J.; Lin, L.Z.; Sun, B.G.; Zhao, M.M. Comparison study on polysaccharide fractions from Laminaria japonica: Structural characterization and bile acid binding capacity. J. Agric. Food Chem. 2017, 65, 9790–9798. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.Y.; Chen, L.; Li, Q.J.; Shen, Y.Z.; Jin, L.; Zhang, X.; Chen, P.C.; Wu, M.J.; Choi, J.I.; et al. Different extraction methods bring about distinct physicochemical properties and antioxidant activities of Sargassum fusiforme fucoidans. Int. J. Biol. Macromol. 2020, 155, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Inukai, K.; Suzuki, M.; Kuga, H.; Korenaga, H. Structural studies on a sulfated polysaccharide from an Arthrobacter sp. by NMR spectroscopy and methylation analysis. Carbohydr. Res. 1997, 305, 253–260. [Google Scholar] [CrossRef]
- Cheng, H.N.; Neiss, T.G. Solution, NMR spectroscopy of food polysaccharides. Polym. Rev. 2012, 52, 81–114. [Google Scholar] [CrossRef]
- Zou, P.; Yang, X.; Yuan, Y.; Jing, C.L.; Cao, J.M.; Wang, Y.; Zhang, L.; Zhang, C.S.; Li, Y.Q. Purification and characterization of a fucoidan from the brown algae Macrocystis pyrifera and the activity of enhancing salt-stress tolerance of wheat seedlings. Int. J. Biol. Macromol. 2021, 180, 547–558. [Google Scholar] [CrossRef]
- Chevolot, L.; Foucault, A.; Chaubet, F.; Kervarec, N.; Sinquin, C.; Fisher, A.M.; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res. 1999, 319, 154–165. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, X.; Xie, H.; Li, X.; Shi, L. Structural characterization and antitumor activity of a polysaccharide from ramulus mori. Carbohydr. Polym. 2018, 190, 232–239. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, T.; Li, J.; Xin, M.; Zhao, X. Structural characterization and effect on leukopenia of fucoidan from Durvillaea antarctica. Carbohydr. Polym. 2021, 256, 117529. [Google Scholar] [CrossRef]
- Bilan, M.I.; Ustyuzhanina, N.E.; Shashkov, A.S.; Thanh, T.T.T.; Bui, M.L.; Tran, T.T.; Bui, V.; Nifantiev, N.E.; Usov, A.I. A sulfated galactofucan from the brown alga Hormophysa cuneiformis (Fucales, Sargassaceae). Carbohydr. Res. 2018, 469, 48–54. [Google Scholar] [CrossRef]
- Synytsya, A.; Kim, W.J.; Kim, S.M.; Pohl, R.; Synytsya, A.; Kvasnicka, F.; Copikova, J.; Park, Y.I. Structure and antitumour activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Rasin, A.B.; Silchenko, A.S.; Kusaykin, M.I.; Malyarenko, O.S.; Zueva, A.O.; Kalinovsky, A.I.; Jia, A.R.; Surits, V.V.; Ermakova, S.P. Enzymatic transformation and anti-tumor activity of Sargassum horneri fucoidan. Carbohydr. Polym. 2020, 246, 116635. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.D.; Yu, G.; Bao, Q.Y.; Xu, X.; Zhu, Y.B.; Ni, H.; Li, Q.B.; Oda, T. Macrophage-stimulating activities of a novel low molecular weight saccharide fragment prepared from ascophyllan with alginate lyase. J. Funct. Food. 2020, 67, 103839. [Google Scholar] [CrossRef]
- Foley, S.A.; Mulloy, B.; Tuohy, M.G. An unfractionated fucoidan from Ascophyllum nodosum: Extraction, characterization, and apoptotic effects in vitro. J. Nat. Prod. 2011, 74, 1851–1861. [Google Scholar] [CrossRef]
- Ye, J.; Chen, D.; Ye, Z.; Huang, Y.; Zhang, N.; Lui, E.M.K.; Xue, C.; Xiao, M. Fucoidan isolated from Saccharina japonica inhibits LPS-induced inflammation in macrophages via blocking NF-κB, MAPK and JAK-STAT pathways. Mar. Drugs 2020, 18, 328. [Google Scholar] [CrossRef]
- Liang, Y.; Zha, S.; Tentaku, M.; Okimura, T.; Jiang, Z.; Ueno, M.; Hirasaka, K.; Yamaguchi, K.; Oda, T. Suppressive effects of sulfated polysaccharide ascophyllan isolated from Ascophyllum nodosum on the production of NO and ROS in LPS-stimulated RAW264.7 cells. Biosci. Biotech. Bioch. 2021, 85, 882–889. [Google Scholar] [CrossRef]
- Jen, C.I.; Su, C.H.; Lai, M.N.; Ng, L.T. Comparative anti-inflammatory characterization of selected fungal and plant water soluble polysaccharides. Food Sci. Technol. Res. 2021, 27, 453–462. [Google Scholar] [CrossRef]
- Park, S.B.; Chun, K.R.; Kim, J.K.; Suk, K.; Jung, Y.M.; Lee, W.-H. The differential effect of high and low molecular weight fucoidans on the severity of collagen-induced arthritis in mice. Phytother. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Sugar Content (%) | Uronic Acid Content (%) | Sulfate Content (%) | Protein Content (%) | Total Polyphenols Content (%) |
|---|---|---|---|---|---|
| ANP-6 | 61.67 ± 1.34 | 3.39 ± 0.29 | 39.88 ± 1.61 | 0.01 ± 0.005 | 0.01 ± 0.002 |
| ANP-7 | 60.88 ± 1.36 | 2.70 ± 0.37 | 42.06 ± 1.48 | 0.01 ± 0.006 | 0.01 ± 0.003 |
| Sample | Methylated Derivative | Deduced Linkage | Characteristic Fragments (m/z) | Molar Ratio |
|---|---|---|---|---|
| dS-ANP-6 | 2,3,4-Me3-Fucp | Fucp-(1→ | 43, 57, 71, 89, 101, 117, 129, 142, 161, 189 | 1.1 |
| 2,3,4,6-Me4-Galp | Galp-(1→ | 43, 57, 71, 87, 101, 117, 129, 143, 161, 205 | 2.4 | |
| 2,4-Me2-Fucp | →3)-Fucp-(1→ | 43, 58, 85, 89, 101, 117, 131, 159, 173, 189, 201, 233 | 12.6 | |
| 3,4-Me2-Fucp | →2)-Fucp-(1→ | 43, 87, 99, 129, 143, 159, 189, 201 | 2.6 | |
| 2,4,6-Me3-Galp | →3)-Galp-(1→ | 43, 71, 87, 99, 101, 117, 129, 161, 173, 233 | 1.6 | |
| 2,3,4-Me3-Galp | →6)-Galp-(1→ | 43, 71, 87, 99, 101, 117, 129, 143, 161, 173, 189, 233 | 1.0 | |
| 2,4-Me2-Galp | →3,6)-Galp-(1→ | 43, 58, 87, 99, 101, 117, 129, 142, 161, 189, 201 | 3.0 | |
| 2,4,6-Me3-Manp | →3)-Manp-(1→ | 43, 59, 87, 101, 115, 129, 145, 173, 189, 215 | 0.2 | |
| dS-ANP-7 | 2,3,4-Me3-Fucp | Fucp-(1→ | 43, 57, 71, 89, 101, 117, 129, 142, 161, 189 | 1.2 |
| 2,3,4,6-Me4-Galp | Galp-(1→ | 43, 57, 71, 87, 101, 117, 129, 143, 161, 205 | 2.3 | |
| 2,4-Me2-Fucp | →3)-Fucp-(1→ | 43, 58, 85, 89, 101, 117, 131, 159, 173, 189, 201, 233 | 13.6 | |
| 3,4-Me2-Fucp | →2)-Fucp-(1→ | 43, 87, 99, 129, 143, 159, 189, 201 | 2.4 | |
| 2,4,6-Me3-Galp | →3)-Galp-(1→ | 43,71,87,99,101,117,129,161,173,233 | 1.6 | |
| 2,3,4-Me3-Galp | →6)-Galp-(1→ | 43, 71, 87, 99, 101, 117, 129, 143, 161, 173, 189, 233 | 1.0 | |
| 2,4-Me2-Galp | →3,6)-Galp-(1→ | 43, 58, 87, 99, 101, 117, 129, 142, 161, 189, 201 | 2.9 | |
| 2,4,6-Me3-Manp | →3)-Manp-(1→ | 43, 59, 87, 101, 115, 129, 145, 173, 189, 215 | 0.2 |
| Sample | Sugar Residue | Chemical Shift (ppm) | ||||||
|---|---|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | |||
| ANP-6 | A | →2)-α-L-Fucp4S-(1→ | 5.20/100.1 | 3.71/74.6 | 4.02/68.9 | 4.50/81.0 | 3.86/67.9 | 1.16/15.6 |
| B | →3)-α-L-Fucp2S4S-(1→ | 5.34/98.9 | 4.50/78.2 | 4.42/74.8 | 4.30/80.3 | 4.28/67.5 | 1.23/16.3 | |
| C | →3,6)-β-D-Galp4S-(1→ | 4.68/102.7 | 3.64/71.4 | 4.81/78.5 | 4.60/77.3 | 3.90/73.7 | 3.84/66.8 | |
| D | →6)-β-D-Galp-(1→ | 4.47/103.3 | 3.58/69.4 | 4.26/74.5 | 4.07/70.1 | 3.86/76.2 | 4.08/67.1 | |
| E | →3)-β-D-Galp-(1→ | 4.42/103.9 | 3.42/71.2 | 4.25/80.3 | 3.69/69.1 | 3.62/73.3 | 3.65/61.1 | |
| ANP-7 | A | →2)-α-L-Fucp4S-(1→ | 5.20/100.1 | 3.71/74.5 | 4.02/68.9 | 4.50/80.9 | 3.86/67.9 | 1.16/15.5 |
| B | →3)-α-L-Fucp2S4S-(1→ | 5.33/98.7 | 4.50/78.3 | 4.43/75.0 | 4.31/80.3 | 4.28/67.5 | 1.27/16.4 | |
| C | →3,6)-β-D-Galp4S-(1→ | 4.68/102.9 | 3.64/71.5 | 4.82/78.3 | 4.60/77.3 | 3.89/73.5 | 3.84/66.8 | |
| D | →6)-β-D-Galp-(1→ | 4.48/103.2 | 3.58/69.3 | 4.28/74.5 | 4.08/70.1 | 3.85/76.3 | 4.07/67.1 | |
| E | →3)-β-D-Galp-(1→ | 4.40/103.8 | 3.42/71.2 | 4.25/80.4 | 3.70/69.1 | 3.62/73.2 | 3.65/61.1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, L.; Yan, C.; Ai, C.; Wen, C.; Guo, X.; Song, S. Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately. Foods 2022, 11, 2381. https://doi.org/10.3390/foods11152381
Wang L, Wang L, Yan C, Ai C, Wen C, Guo X, Song S. Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately. Foods. 2022; 11(15):2381. https://doi.org/10.3390/foods11152381
Chicago/Turabian StyleWang, Lilong, Linlin Wang, Chunhong Yan, Chunqing Ai, Chengrong Wen, Xiaoming Guo, and Shuang Song. 2022. "Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately" Foods 11, no. 15: 2381. https://doi.org/10.3390/foods11152381
APA StyleWang, L., Wang, L., Yan, C., Ai, C., Wen, C., Guo, X., & Song, S. (2022). Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately. Foods, 11(15), 2381. https://doi.org/10.3390/foods11152381