
Acyclic Diterpene Phytol from Hemp Seed Oil (Cannabis sativa L.) Exerts Anti-Inflammatory Activity on Primary Human Monocytes-Macrophages

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Phytol from an Unsaponifiable Fraction of Hemp Seed Oil
2.2. Blood Collection Procedure and Human Monocytes Isolation
2.3. FACS Immunostaining of Circulating Monocytes Cells
2.4. Differentiation and Polarization of Monocyte-Derived Macrophages M1 and M2
2.5. MTT Cell Viability Assay
2.6. Isolation of RNA and qRT-PCR Protocol
2.7. Cytokine Release Measurement
2.8. Data Analysis
3. Results and Discussion
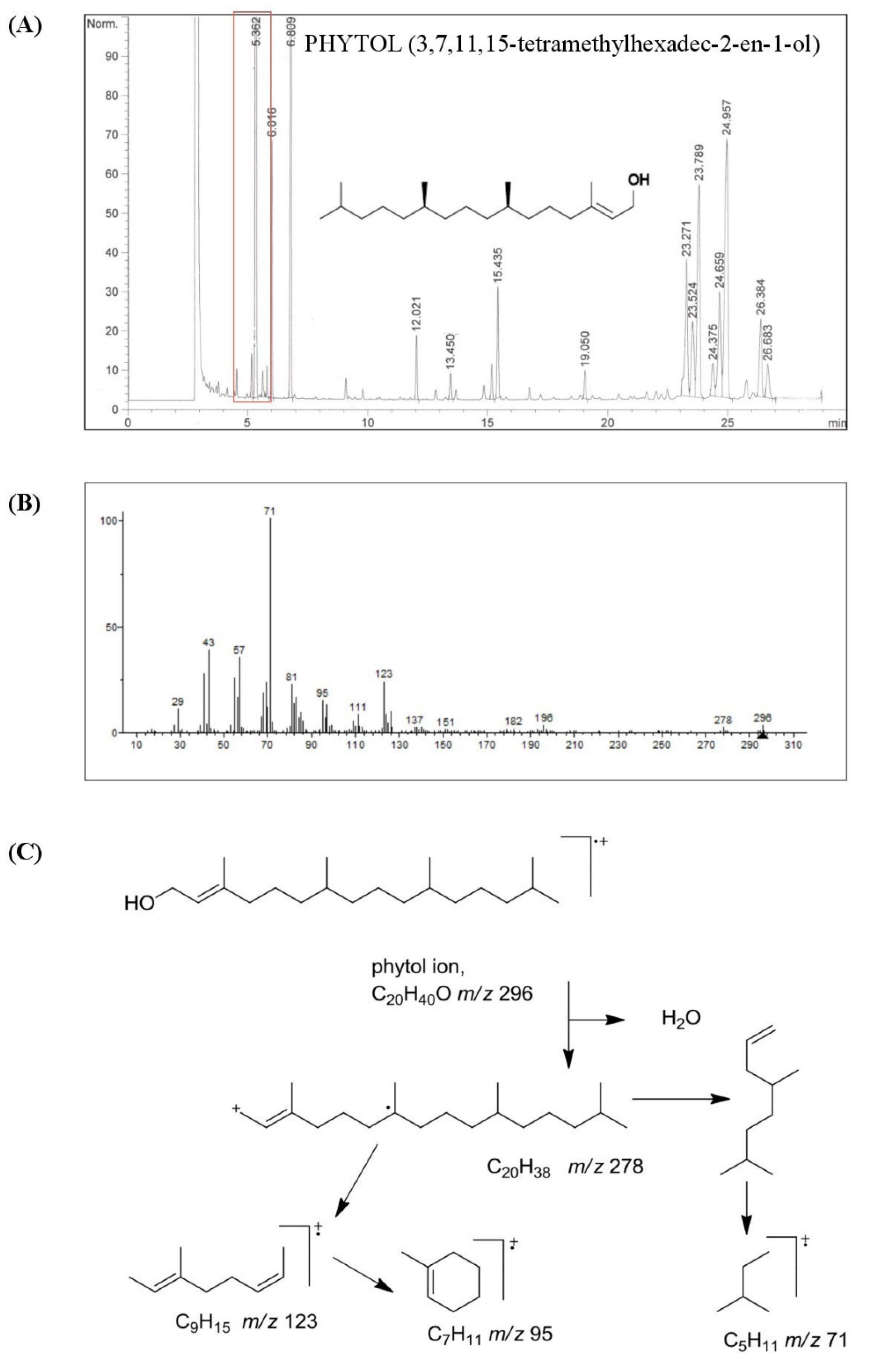
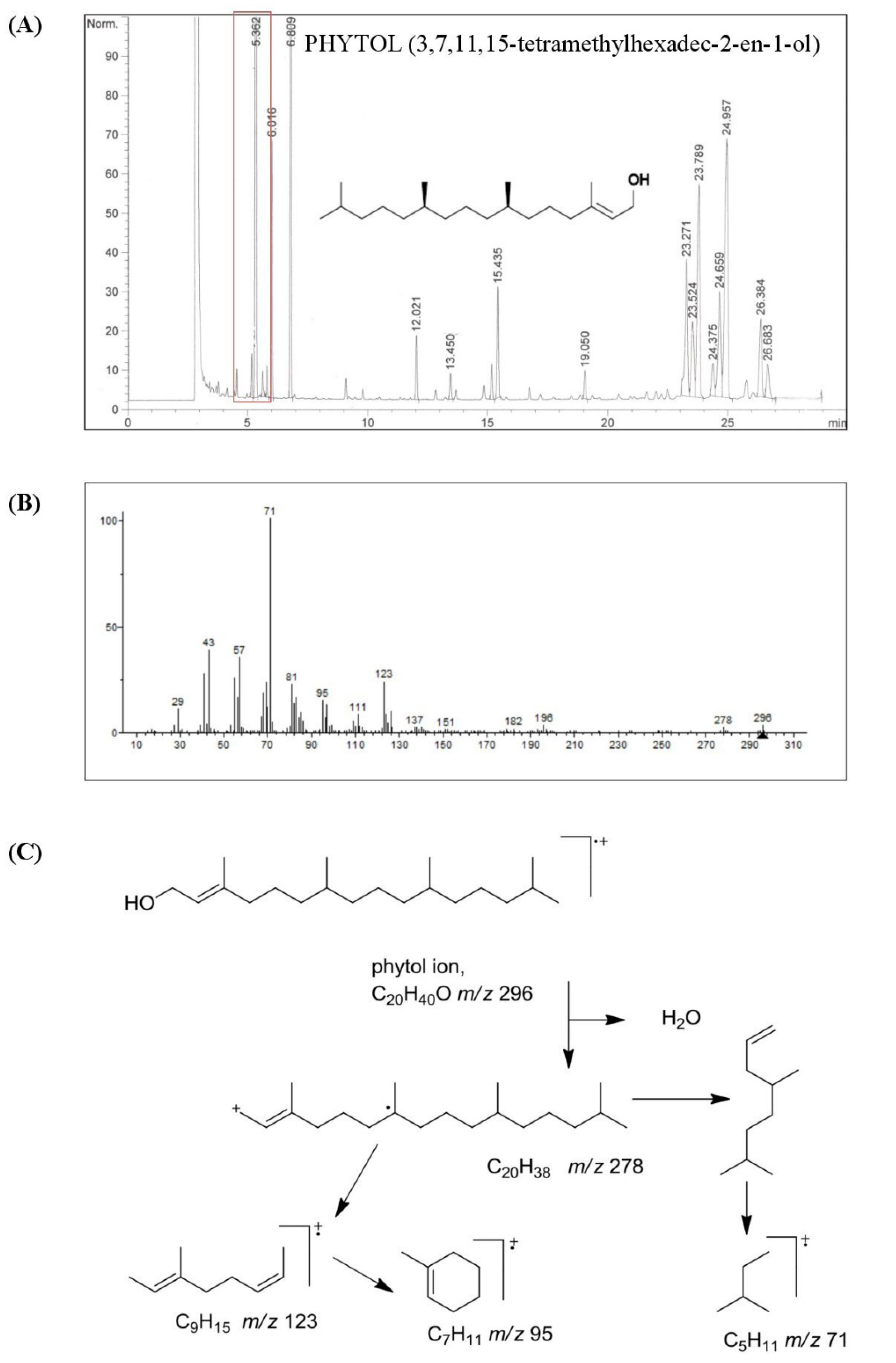
3.1. Chemical Characterization of HSO
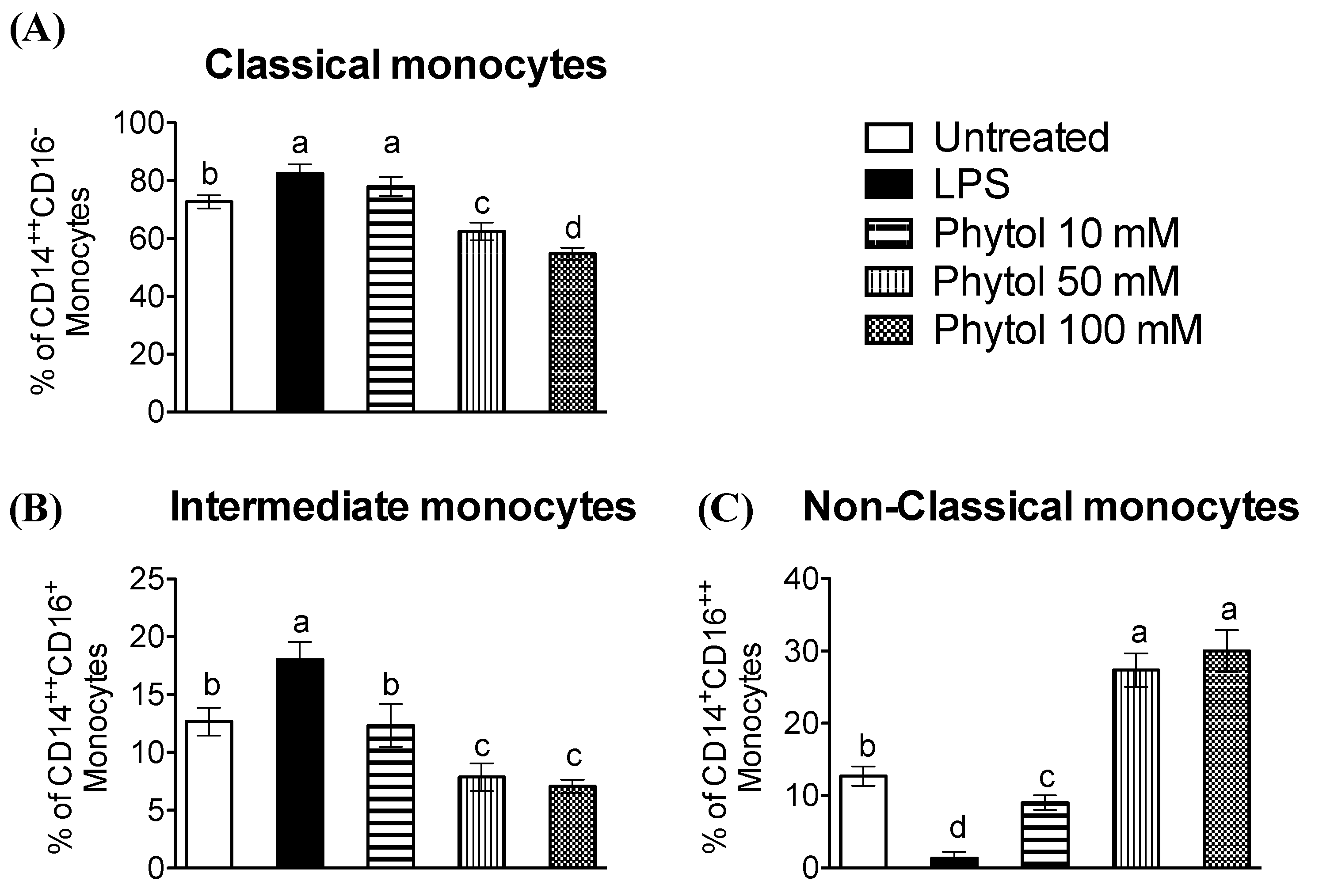
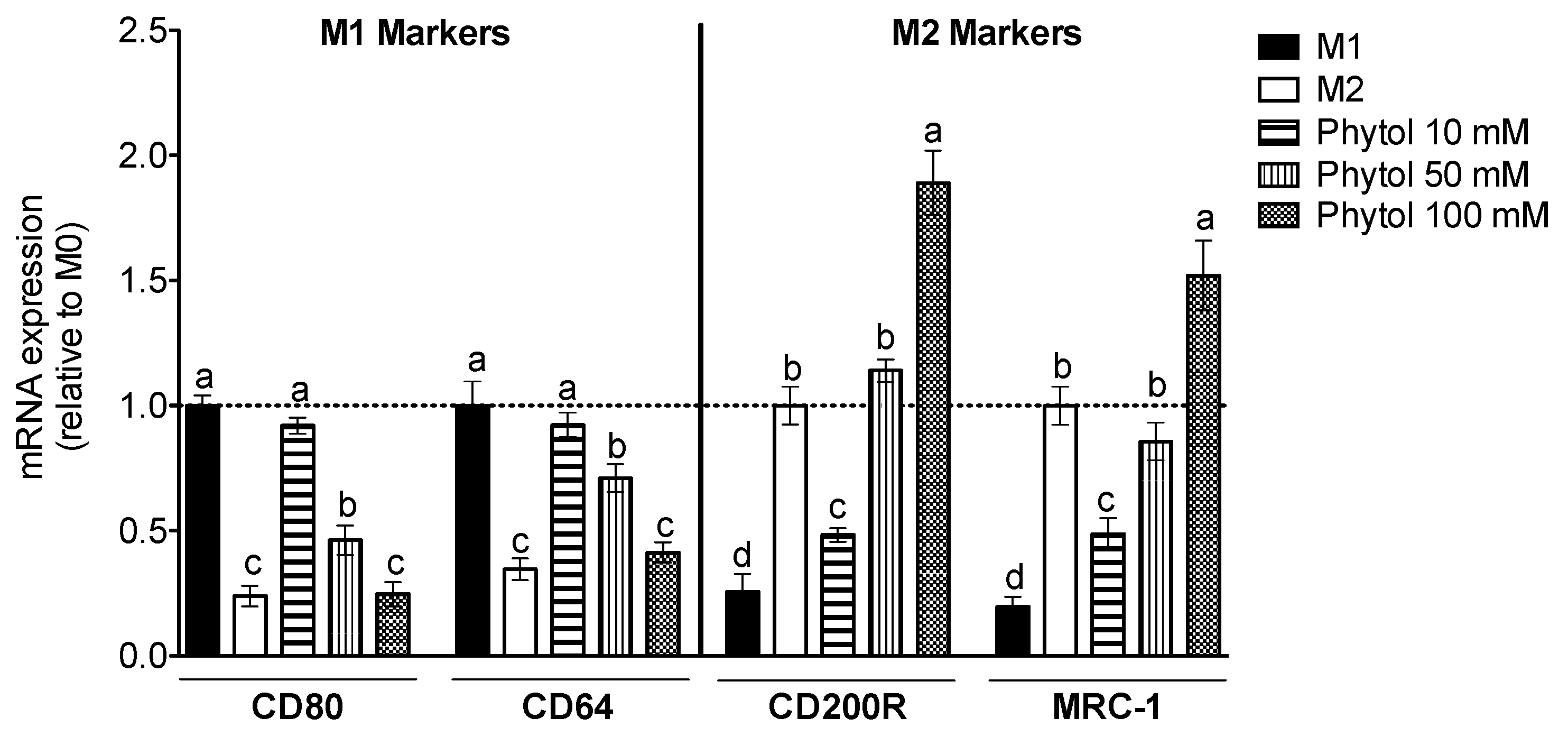
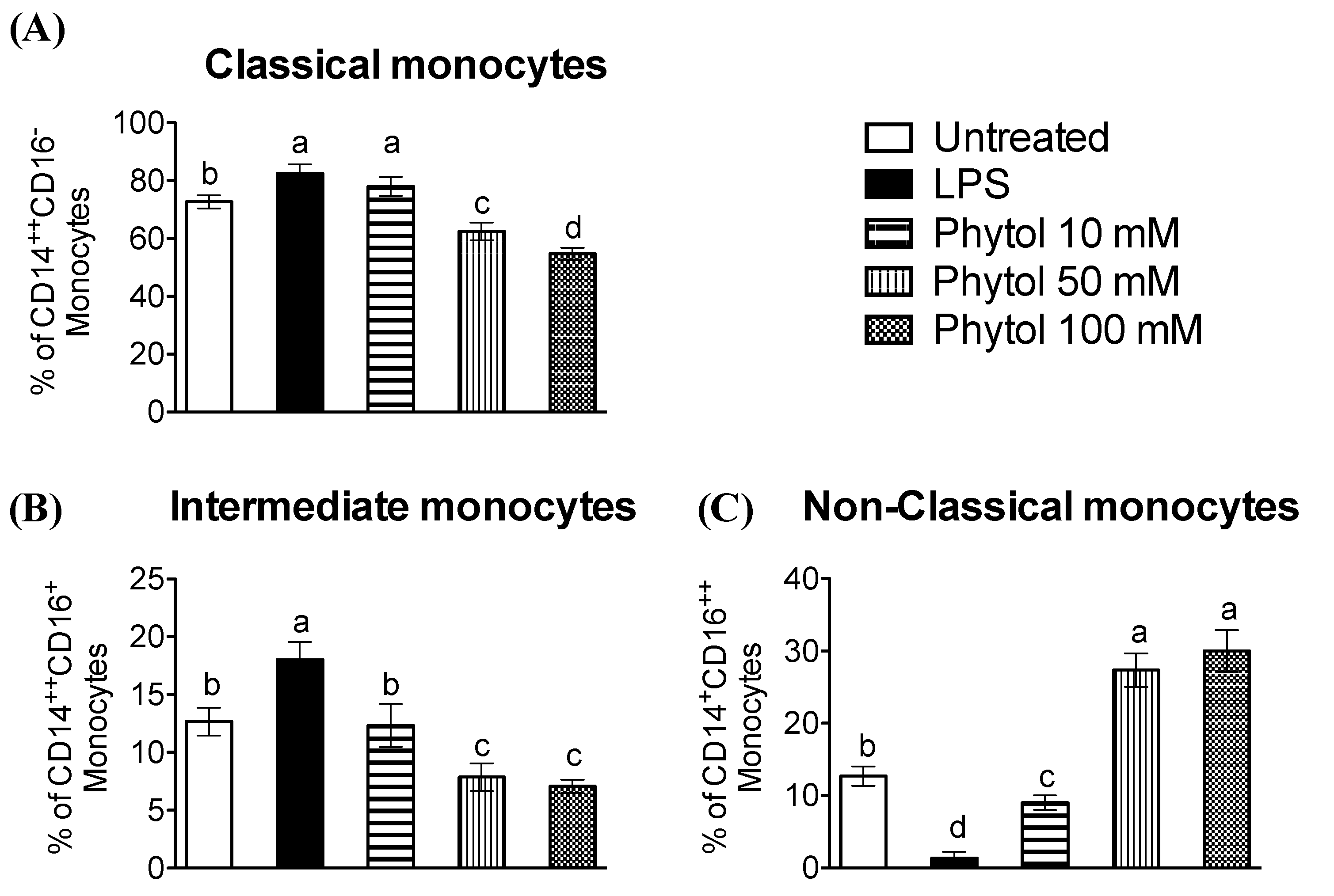
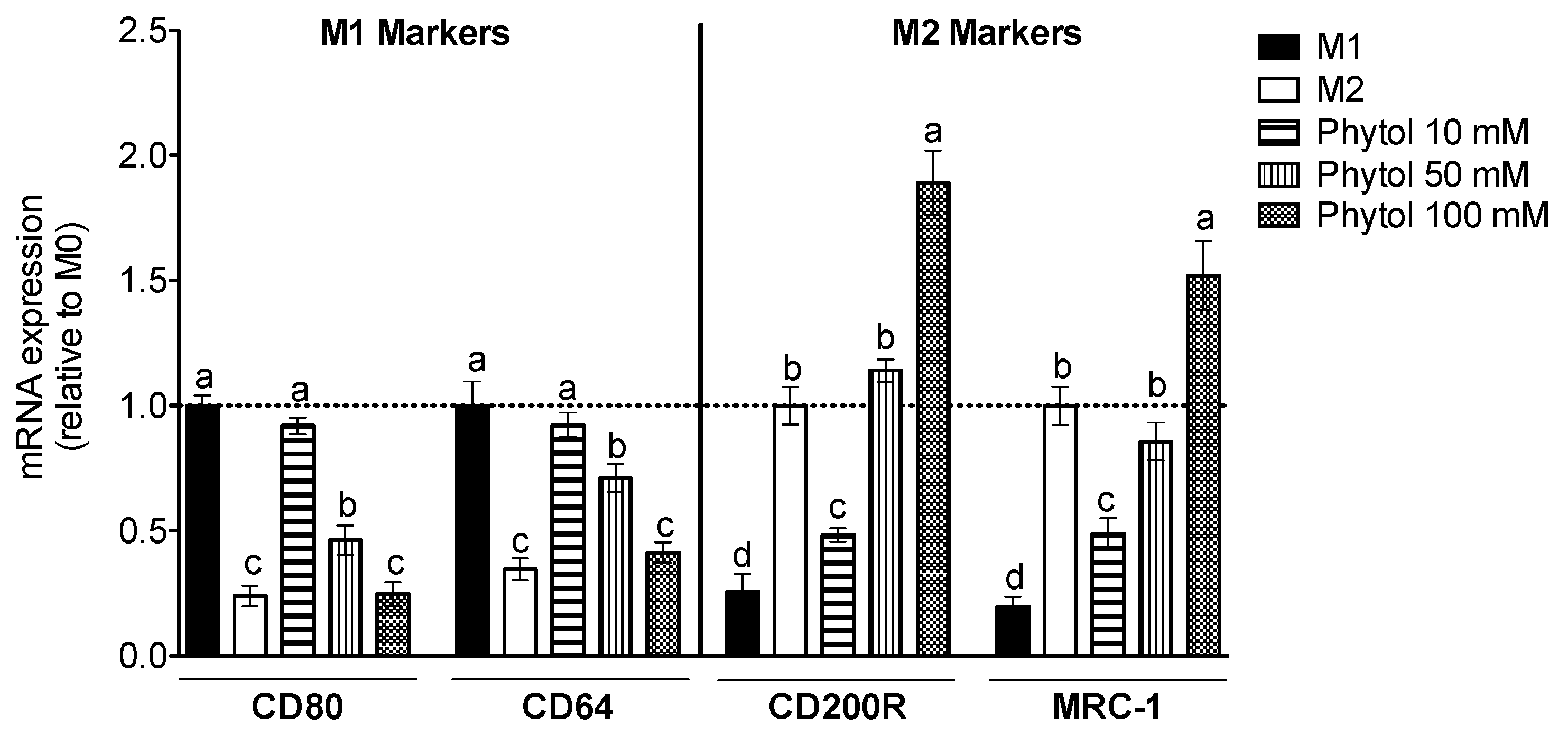
3.2. Effect of Phytol on Monocyte Differentiation and Macrophage Polarization
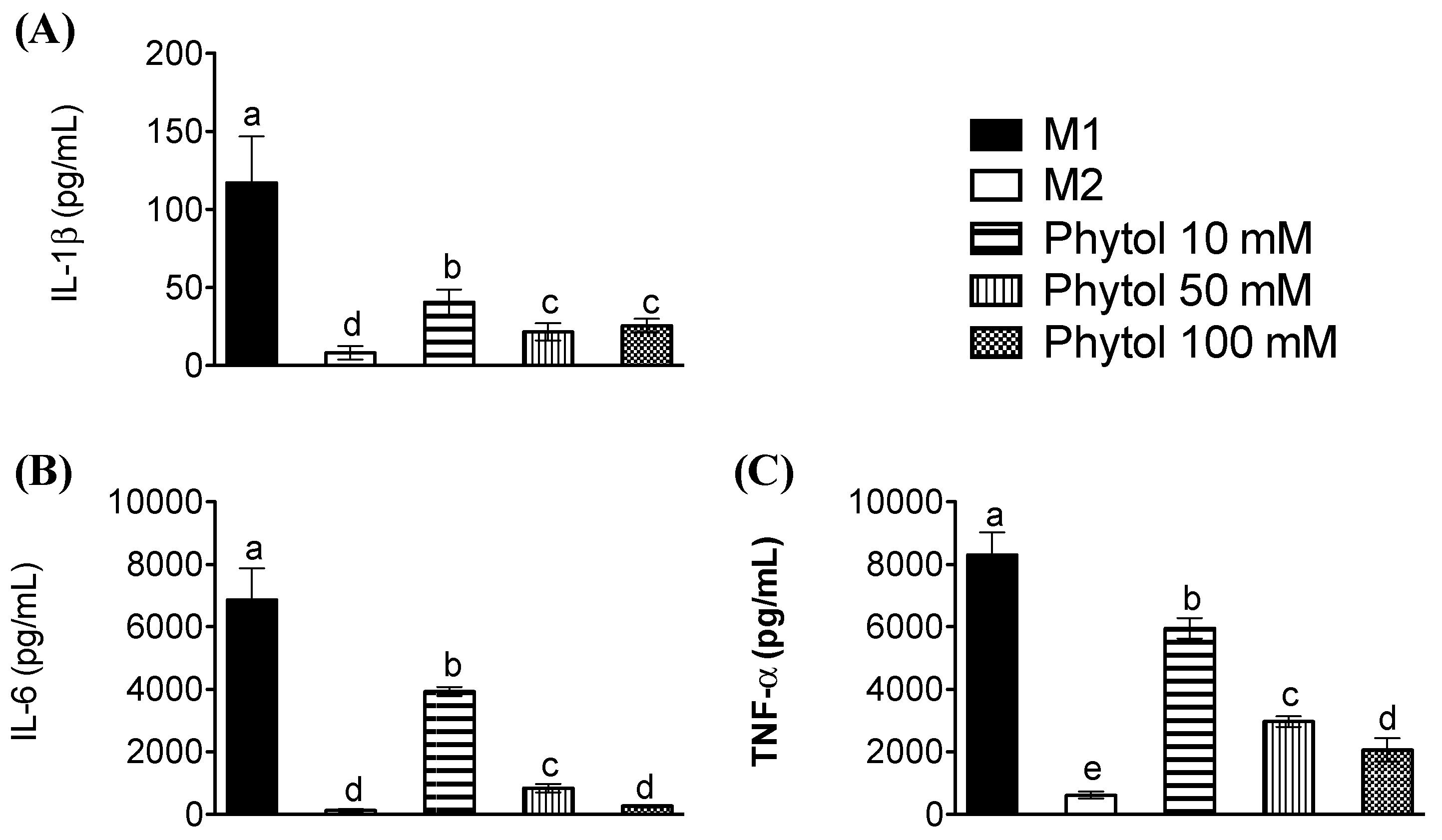
3.3. Effects of Phytol on Pro-Inflammatory Cytokine Levels
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, X.; Tang, J.; Passos, C.D.S.; Nurisso, A.; Simões-Pires, C.A.; Ji, M.; Lou, H.; Fan, P. Characterization of Lignanamides from Hemp (Cannabis sativa L.) Seed and Their Antioxidant and Acetylcholinesterase Inhibitory Activities. J. Agric. Food Chem. 2015, 63, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrition 2020, 12, 1935. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, H.P.V.; Davis, A.; Kumar, S.K.; Murray, B.; Zheljazkov, V.D. Industrial Hemp (Cannabis sativa subsp. sativa) as an Emerging Source for Value-Added Functional Food Ingredients and Nutraceuticals. Molecules 2020, 25, 4078. [Google Scholar] [CrossRef] [PubMed]
- Rea Martinez, J.; Montserrat-de la Paz, S.; De la Puerta, R.; Garcia-Gimenez, M.D.; Fernandez-Arche, M.A. Characterization of bioactive compounds in defatted hempseed (Cannabis sativa L.) by UHPLC-HRMS/MS and anti-inflammatory activity in primary human monocytes. Food Funct. 2020, 11, 4057–4066. [Google Scholar] [CrossRef]
- Rodriguez-Martin, N.M.; Montserrat-de la Paz, S.; Toscano, R.; Grao-Cruces, E.; Villanueva, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Hemp (Cannabis sativa L.) Protein Hydrolysates Promote Anti-Inflammatory Response in Primary Human Monocytes. Biomolecules 2020, 10, 803. [Google Scholar] [CrossRef]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef]
- Tapia, G.; Valenzuela, R.; Espinosa, A.; Romanque, P.; Dossi, C.; Gonzalez-Mañán, D.; Videla, L.A.; D’Espessailles, A. N-3 long-chain PUFA supplementation prevents high fat diet induced mouse liver steatosis and inflammation in relation to PPAR-α upregulation and NF-κB DNA binding abrogation. Mol. Nutr. Food Res. 2014, 58, 1333–1341. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.M.; Marín-Aguilar, F.; García-Giménez, M.D.; Fernández-Arche, M.A. Hemp (Cannabis sativa L.) Seed Oil: Analytical and Phytochemical Characterization of the Unsaponifiable Fraction. J. Agric. Food Chem. 2014, 62, 1105–1110. [Google Scholar] [CrossRef]
- Oomah, B.D.; Busson, M.; Godfrey, D.V.; Drover, J.C.G. Characteristics of hemp (Cannabis sativa L.) seed oil. Food Chem. 2002, 76, 33–43. [Google Scholar] [CrossRef]
- Rezapour-Firouzi, S.; Mohammadian, M.; Sadeghzadeh, M.; Mehranfar, S.; Mazloomi, E. The effects of evening primrose/hemp seed oil compared to rapamycin on the gene expression of immunological parameters in experimental autoimmune encephalomyelitis splenocytes. Iran. J. Allergy Asthma Immunol. 2020, 19, 183–192. [Google Scholar] [CrossRef]
- Izzo, L.; Pacifico, S.; Piccolella, S.; Castaldo, L.; Narváez, A.; Grosso, M.; Ritieni, A. Chemical Analysis of Minor Bioactive Components and Cannabidiolic Acid in Commercial Hemp Seed Oil. Molecules 2020, 25, 3710. [Google Scholar] [CrossRef] [PubMed]
- Montserrat-De La Paz, S.; García-Giménez, M.D.; Ángel-Martín, M.; Pérez-Camino, M.C.; Fernández Arche, A. Long-chain fatty alcohols from evening primrose oil inhibit the inflammatory response in murine peritoneal macrophages. J. Ethnopharmacol. 2014, 151, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, A.M.; Capocasale, M. Policosanol in Tomato (Solanum lycopersicum L.) Seed Oil: The Effect of Cultivar. J. Oleo Sci. 2015, 64, 625–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, R.O.; Sousa, F.B.M.; Damasceno, S.R.B.; Carvalho, N.S.; Silva, V.G.; Oliveira, F.R.M.A.; Sousa, D.P.; Aragão, K.S.; Barbosa, A.L.R.; Freitas, R.M.; et al. Phytol, a diterpene alcohol, inhibits the inflammatory response by reducing cytokine production and oxidative stress. Fundam. Clin. Pharmacol. 2014, 28, 455–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moraes, J.; de Oliveira, R.N.; Costa, J.P.; Junior, A.L.G.; de Sousa, D.P.; Freitas, R.M.; Allegretti, S.M.; Pinto, P.L.S. Phytol, a Diterpene Alcohol from Chlorophyll, as a Drug against Neglected Tropical Disease Schistosomiasis Mansoni. PLoS Negl. Trop. Dis. 2014, 8, e2617. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Chandra Shill, M.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef]
- Alencar, M.V.O.B.; Islam, M.T.; Ali, E.S.; Santos, J.V.O.; Paz, M.F.C.J.; Sousa, J.M.C.; Dantas, S.M.M.M.; Mishra, S.K.; Cavalcante, A.A.C.M. Association of Phytol with Toxic and Cytotoxic Activities in an Antitumoral Perspective: A Meta-Analysis and Systemic Review. Anticancer. Agents Med. Chem. 2018, 18, 1828–1837. [Google Scholar] [CrossRef]
- Al-Sayed, E.; Ke, T.Y.; Hwang, T.L.; Chen, S.R.; Korinek, M.; Chen, S.L.; Cheng, Y. Bin Cytotoxic and anti-inflammatory effects of lignans and diterpenes from Cupressus macrocarpa. Bioorganic Med. Chem. Lett. 2020, 30, 127127. [Google Scholar] [CrossRef]
- Fang, S.M.; Liu, R.; Li, L.; Yao, J.L.; Liu, E.W.; Fan, G.W.; Zhang, H.; Gao, X.M. Anti-inflammatory diterpenes from the fruits of Vitex trifolia L. var. simplicifolia Cham. J. Asian Nat. Prod. Res. 2019, 21, 985–991. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Liu, Y.P.; Wang, X.P.; Qiao, Z.H.; Yu, X.M.; Zhu, Y.Z.; Xie, L.; Qiang, L.; Fu, Y.H. Bioactive daphnane diterpenes from Wikstroemia chuii with their potential anti-inflammatory effects and anti-HIV activities. Bioorg. Chem. 2020, 105, 104388. [Google Scholar] [CrossRef]
- Campos-Xolalpa, N.; Alonso-Castro, Á.J.; Ortíz-Sanchez, E.; Zapata-Morales, J.R.; González-Chávez, M.M.; Pérez, S. Anti-inflammatory and antitumor activities of the chloroform extract and anti-inflammatory effect of the three diterpenes isolated from Salvia ballotiflora Benth. BMC Complement. Med. Ther. 2021, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Ayatollahi, S.A.; Zihad, S.M.N.K.; Sifat, N.; Khan, M.R.; Paul, A.; Salehi, B.; Islam, T.; Mubarak, M.S.; Martins, N.; et al. Phytol anti-inflammatory activity: Pre-clinical assessment and possible mechanism of action elucidation. Cell. Mol. Biol. 2020, 66, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Riboldi, E.; Ippolito, A.; Sica, A. Molecular and epigenetic basis of macrophage polarized activation. Semin. Immunol. 2015, 27, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J.; Barrett, T.J. Monocytes and macrophages in atherogenesis. Curr. Opin. Lipidol. 2019, 30, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, B.; Dahl, T.B.; Medina, I.; Groeneweg, M.; Holm, S.; Montserrat-de la Paz, S.; Rousch, M.; Otten, J.; Herias, V.; Varela, L.M.; et al. Leukocyte Overexpression of Intracellular NAMPT Attenuates Atherosclerosis by Regulating PPARγ-Dependent Monocyte Differentiation and Function. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1157–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montserrat-De La Paz, S.; De La Puerta, R.; Fernandez-Arche, A.; Quilez, A.M.; Muriana, F.J.G.; Garcia-Gimenez, M.D.; Bermudez, B. Pharmacological effects of mitraphylline from Uncaria tomentosa in primary human monocytes: Skew toward M2 macrophages. J. Ethnopharmacol. 2015, 170, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.A.; Yáñez, A.; Barman, P.K.; Goodridge, H.S. The ontogeny of monocyte subsets. Front. Immunol. 2019, 10, 1642. [Google Scholar] [CrossRef] [Green Version]
- Ożańska, A.; Szymczak, D.; Rybka, J. Pattern of human monocyte subpopulations in health and disease. Scand. J. Immunol. 2020, 92, e12883. [Google Scholar] [CrossRef]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [Green Version]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Wong, K.L.; Yeap, W.H.; Tai, J.J.Y.; Ong, S.M.; Dang, T.M.; Wong, S.C. The three human monocyte subsets: Implications for health and disease. Immunol. Res. 2012, 53, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Benoit, M.; Desnues, B.; Mege, J.-L. Macrophage Polarization in Bacterial Infections. J. Immunol. 2008, 181, 3733–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montserrat-De La Paz, S.; Fernández-Arche, M.A.; Ángel-Martín, M.; García-Giménez, M.D. Phytochemical characterization of potential nutraceutical ingredients from Evening Primrose oil (Oenothera biennis L.). Phytochem. Lett. 2014, 8, 158–162. [Google Scholar] [CrossRef]
- Martin, M.E.; Millan-Linares, M.C.; Naranjo, M.C.; Toscano, R.; Abia, R.; Muriana, F.J.G.; Bermudez, B.; Montserrat-de la Paz, S. Minor compounds from virgin olive oil attenuate LPS-induced inflammation via visfatin-related gene modulation on primary human monocytes. J. Food Biochem. 2019, 43, e12941. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Bermudez, B.; Lopez, S.; Naranjo, M.C.; Romero, Y.; Bando-Hidalgo, M.J.; Abia, R.; Muriana, F.J.G. Exogenous fatty acids and niacin on acute prostaglandin D 2 production in human myeloid cells. J. Nutr. Biochem. 2017, 39, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-De La Paz, S.; Fernández-Arche, Á.; Ángel-Martín, M.; García-Giménez, M.D. The sterols isolated from Evening Primrose oil modulate the release of proinflammatory mediators. Phytomedicine 2012, 19, 1072–1076. [Google Scholar] [CrossRef]
- Toscano, R.; Millan-Linares, M.C.; Lemus-Conejo, A.; Claro, C.; Sanchez-Margalet, V.; Montserrat-de la Paz, S. Postprandial triglyceride-rich lipoproteins promote M1/M2 microglia polarization in a fatty-acid-dependent manner. J. Nutr. Biochem. 2020, 75, 108248. [Google Scholar] [CrossRef]
- Guo, Y.L.; Xu, R.X.; Zhu, C.G.; Wu, N.Q.; Cui, Z.P.; Li, J.J. Policosanol attenuates statin-induced increases in serum proprotein convertase subtilisin/kexin type 9 when combined with atorvastatin. Evid.-Based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef]
- Fernández-Arche, A.; Marquez-Martín, A.; Vazquez, R.D.L.P.; Perona, J.S.; Terencio, C.; Perez-Camino, C.; Ruiz-Gutierrez, V. Long-chain fatty alcohols from pomace olive oil modulate the release of proinflammatory mediators. J. Nutr. Biochem. 2009, 20, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Millan-Linares, M.C.; Bermudez, B.; Martin, M.E.; Muñoz, E.; Abia, R.; Millan, F.; Muriana, F.J.G.; Montserrat-de la Paz, S. Unsaponifiable fraction isolated from grape (Vitis vinifera L.) seed oil attenuates oxidative and inflammatory responses in human primary monocytes. Food Funct. 2018, 9, 2517–2523. [Google Scholar] [CrossRef] [Green Version]
- He, X.-F.; Chen, J.-J.; Li, T.-Z.; Zhang, X.-K.; Guo, Y.-Q.; Zhang, X.-M.; Hu, J.; Geng, C.-A. Nineteen New Flavanol–Fatty Alcohol Hybrids with α-Glucosidase and PTP1B Dual Inhibition: One Unusual Type of Antidiabetic Constituent from Amomum tsao-ko. J. Agric. Food Chem. 2020, 68, 11434–11448. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.D.F.; Barreto, A.S.; Gomes Trindade, G.G.; Lima, C.M.; Araujo, A.A.S.; Menezes, I.R.A.; Candido, E.A.F.; Santana, E.T.N.; Silva-Junior, W.M.; Quintans, J.S.S.; et al. Enhancement of the functionality of women with knee osteoarthritis by a gel formulation with Caryocar coriaceum Wittm (“Pequi”) nanoencapsulated pulp fixed oil. Biomed. Pharmacother. 2022, 150, 112938. [Google Scholar] [CrossRef] [PubMed]
- Figuereido, P.R.L.; Oliveira, I.B.; Neto, J.B.S.; Oliveira, J.A.; Ribeiro, L.B.; Viana, G.S.B.; Rocha, T.M.; Leal, L.K.A.M.; Kerntopf, M.R.; Felipe, C.F.B.; et al. Caryocar coriaceum Wittm. (Pequi) fixed oil presents hypolipemic and anti-inflammatory effects in vivo and in vitro. J. Ethnopharmacol. 2016, 191, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Brito, R.G.; Dos Santos, L.P.; Quintans, J.S.S.; Lucca Junior, W.; Araujo, A.A.S.; Saravanan, S.; Menezes, I.R.A.; Countinho, H.D.M.; Quintans-Junior, L.J. Citronellol, a natural acyclic monoterpene, attenuates mechanical hyperalgesia response in mice: Evidence of the spinal cord lamina I inhibition. Chem. Biol. Interact. 2015, 239, 111–117. [Google Scholar] [CrossRef]
- Barreto, R.S.S.; Quintans, J.S.S.; Amarante, R.K.L.; Nascimento, T.S.; Amarante, R.S.; Barreto, A.S.; Pereira, E.W.M.; Duarte, M.C.; Countinho, H.D.M.; Menezes, I.R.A.; et al. Evidence for the involvement of TNF-α and IL-1β in the antinociceptive and anti-inflammatory activity of Stachys lavandulifolia Vahl. (Lamiaceae) essential oil and (-)-α-bisabolol, its main compound, in mice. J. Ethnopharmacol. 2016, 191, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Vetter, W.; Schröder, M.; Lehnert, K. Differentiation of refined and virgin edible oils by means of the trans—And cis-phytol isomer distribution. J. Agric. Food Chem. 2012, 60, 6103–6107. [Google Scholar] [CrossRef]
- Brown, P.J.; Mei, G.; Gibberd, F.B.; Burston, D.; Mayne, P.D.; McClinchy, J.E.; Sidey, M. Diet and Refsum’s disease. The determination of phytanic acid and phytol in certain foods and the application of this knowledge to the choice of suitable convenience foods for patients with Refsum’s disease. J. Hum. Nutr. Diet. 1993, 6, 295–305. [Google Scholar] [CrossRef]
- Pejin, B.; Kojic, V.; Bogdanovic, G. Natural Product Research Formerly Natural Product Letters An insight into the cytotoxic activity of phytol at in vitro conditions. Nat. Prod. Res. 2014, 28, 2053–2056. [Google Scholar] [CrossRef]
- Costa, J.; Islam, M.; Santos, P.; Ferreira, P.; Oliveira, G.; Alencar, M.; Paz, M.; Ferreira, É.; Feitosa, C.; Citó, A.; et al. Evaluation of Antioxidant Activity of Phytol Using Non- and Pre-Clinical Models. Curr. Pharm. Biotechnol. 2016, 17, 1278–1284. [Google Scholar] [CrossRef]
- Wang, J.; Hu, X.; Ai, W.; Zhang, F.; Yang, K.; Wang, L.; Zhu, X.; Gao, P.; Shu, G.; Jiang, Q.; et al. Phytol increases adipocyte number and glucose tolerance through activation of PI3K/Akt signaling pathway in mice fed high-fat and high-fructose diet. Biochem. Biophys. Res. Commun. 2017, 489, 432–438. [Google Scholar] [CrossRef]
- Bobe, G.; Zhang, Z.; Kopp, R.; Garzotto, M.; Shannon, J.; Takata, Y. Phytol and its metabolites phytanic and pristanic acids for risk of cancer: Current evidence and future directions. Eur. J. Cancer Prev. 2020, 29, 191. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Madrigal, C.; Lopez, S.; Grao-Cruces, E.; Millan-Linares, M.C.; Rodriguez-Martin, N.M.; Martin, M.E.; Alba, G.; Santa-Maria, C.; Bermudez, B.; Montserrat-de la Paz, S. Dietary Fatty Acids in Postprandial Triglyceride-Rich Lipoproteins Modulate Human Monocyte-Derived Dendritic Cell Maturation and Activation. Nutrients 2020, 12, 3139. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Rodriguez-Martin, N.M.; Villanueva, A.; Pedroche, J.; Cruz-Chamorro, I.; Millan, F.; Millan-Linares, M.C. Evaluation of Anti-Inflammatory and Atheroprotective Properties of Wheat Gluten Protein Hydrolysates in Primary Human Monocytes. Foods 2020, 9, 854. [Google Scholar] [CrossRef]
- Rosillo, M.A.; Montserrat-de la Paz, S.; Abia, R.; Castejon, M.L.; Millan-Linares, M.C.; Alarcon-de la Lastra, C.; Fernandez-Bolaños, J.G.; Muriana, F.J.G. Oleuropein and its peracetylated derivative negatively regulate osteoclastogenesis by controlling the expression of genes involved in osteoclast differentiation and function. Food Funct. 2020, 11, 4038–4048. [Google Scholar] [CrossRef] [PubMed]
- Rogacev, K.S.; Zawada, A.M.; Emrich, I.; Seiler, S.; Böhm, M.; Fliser, D.; Woollard, K.J.; Heine, G.H. Lower Apo A-I and lower HDL-C levels are associated with higher intermediate CD14++CD16+monocyte counts that predict cardiovascular events in chronic kidney disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2120–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Jiang, J.; Chen, W.; Li, W.; Chen, Z. Review Article Vascular Macrophages in Atherosclerosis. J. Immunol. Res. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Malyshev, I.; Malyshev, Y. Current concept and update of the macrophage plasticity concept: Intracellular mechanisms of reprogramming and M3 macrophage “switch” phenotype. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.M.S.; Heimfarth, L.; Pereira, E.W.M.; Oliveira, F.S.; Menezes, I.R.A.; Coutinho, H.D.M.; Picot, L.; Antoniolli, A.R.; Quintans, J.S.S.; Quintans-Júnior, L.J. Phytol, a Chlorophyll Component, Produces Antihyperalgesic, Anti-inflammatory, and Antiarthritic Effects: Possible NFκB Pathway Involvement and Reduced Levels of the Proinflammatory Cytokines TNF-α and IL-6. J. Nat. Prod. 2020, 83, 1107–1117. [Google Scholar] [CrossRef]
- Blum, L.; Tafferner, N.; Spring, I.; Kurz, J.; DeBruin, N.; Geisslinger, G.; Parnham, M.J.; Schiffmann, S. Dietary phytol reduces clinical symptoms in experimental autoimmune encephalomyelitis (EAE) at least partially by modulating NOX2 expression. J. Mol. Med. 2018, 96, 1131–1144. [Google Scholar] [CrossRef]
- Corcoran, M.P.; Meydani, M.; Lichtenstein, A.H.; Schaefer, E.J.; Dillard, A.; Lamon-Fava, S. Sex hormone modulation of proinflammatory cytokine and CRP expression in macrophages from older men and postmenopausal women. J. Endocrinol. 2010, 206, 217. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-de la Paz, S. Active Ingredients from Foods: Biochemical and Processing Aspects. Foods 2021, 10, 1240. [Google Scholar] [CrossRef] [PubMed]
- Hanna, A.; Frangogiannis, N.G. Inflammatory Cytokines and Chemokines as Therapeutic Targets in Heart Failure. Cardiovasc. Drugs Ther. 2020, 34, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, C.Y.; Deng, W.M. The role of pro-inflammatory cytokines in lipid metabolism of metabolic diseases. Int. Rev. Immunol. 2019, 38, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Li, S.; Pathak, J.L. Pro-inflammatory Cytokines and Osteocytes. Curr. Osteoporos. Rep. 2019, 17, 97–104. [Google Scholar] [CrossRef]
- Hu, F.; Lou, N.; Jiao, J.; Guo, F.; Xiang, H.; Shang, D. Macrophages in pancreatitis: Mechanisms and therapeutic potential. Biomed. Pharmacother. 2020, 131, 110693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, D.; Bai, X. Macrophages regulate the progression of osteoarthritis. Osteoarthr. Cartil. 2020, 28, 555–561. [Google Scholar] [CrossRef]
- Kishore, A.; Petrek, M. Roles of Macrophage Polarization and Macrophage-Derived miRNAs in Pulmonary Fibrosis. Front. Immunol. 2021, 12, 678457. [Google Scholar] [CrossRef]
- Ma, H.; Li, Y.N.; Song, L.; Liu, R.; Li, X.; Shang, Q.; Wang, Y.; Shao, C.; Shi, Y. Macrophages inhibit adipogenic differentiation of adipose tissue derived mesenchymal stem/stromal cells by producing pro-inflammatory cytokines. Cell Biosci. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Leuti, A.; Talamonti, E.; Gentile, A.; Tiberi, M.; Matteocci, A.; Fresegna, D.; Centonze, D.; Chiurchiù, V. Macrophage Plasticity and Polarization Are Altered in the Experimental Model of Multiple Sclerosis. Biomolecules 2021, 11, 837. [Google Scholar] [CrossRef]
- Fernandes, T.L.; Gomoll, A.H.; Lattermann, C.; Hernandez, A.J.; Bueno, D.F.; Amano, M.T. Macrophage: A Potential Target on Cartilage Regeneration. Front. Immunol. 2020, 11, 111. [Google Scholar] [CrossRef] [Green Version]
- Mesquida-Veny, F.; Del Río, J.A.; Hervera, A. Macrophagic and microglial complexity after neuronal injury. Prog. Neurobiol. 2021, 200, 101970. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.C.; Wu, H.L.; Chen, S.H.; Wang, Y.T.; Wu, C.C. Thrombomodulin facilitates peripheral nerve regeneration through regulating M1/M2 switching. J. Neuroinflamm. 2020, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | GenBank Accession Number | Direction | Sequence (5′→3′) |
|---|---|---|---|
| CD80 | NM_005191.3 | Forward Reverse | GGGAAAGTGTACGCCCTGTA GCTACTTCTGTGCCCACCAT |
| CD200R | NM_138940.2 | Forward Reverse | GTTGCCCTCCTATCGCATTA TGGAAATTCCCATCAGGTGT |
| MR | NM_002438.3 | Forward Reverse | GGCGGTGACCTCACAAGTAT ACGAAGCCATTTGGTAAACG |
| CD64 | NM_000566.3 | Forward Reverse | GTCCAAATCTCCAAGTGCGG CCCAAGTATGAGAGCAGCGT |
| GAPDH | NM_001289746 | Forward Reverse | CACATGGCCTCCAAGGAGTAAG CCAGCAGTGAGGGTCTCTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claro-Cala, C.M.; Grao-Cruces, E.; Toscano, R.; Millan-Linares, M.C.; Montserrat-de la Paz, S.; Martin, M.E. Acyclic Diterpene Phytol from Hemp Seed Oil (Cannabis sativa L.) Exerts Anti-Inflammatory Activity on Primary Human Monocytes-Macrophages. Foods 2022, 11, 2366. https://doi.org/10.3390/foods11152366
Claro-Cala CM, Grao-Cruces E, Toscano R, Millan-Linares MC, Montserrat-de la Paz S, Martin ME. Acyclic Diterpene Phytol from Hemp Seed Oil (Cannabis sativa L.) Exerts Anti-Inflammatory Activity on Primary Human Monocytes-Macrophages. Foods. 2022; 11(15):2366. https://doi.org/10.3390/foods11152366
Chicago/Turabian StyleClaro-Cala, Carmen M., Elena Grao-Cruces, Rocio Toscano, Maria C. Millan-Linares, Sergio Montserrat-de la Paz, and Maria E. Martin. 2022. "Acyclic Diterpene Phytol from Hemp Seed Oil (Cannabis sativa L.) Exerts Anti-Inflammatory Activity on Primary Human Monocytes-Macrophages" Foods 11, no. 15: 2366. https://doi.org/10.3390/foods11152366
APA StyleClaro-Cala, C. M., Grao-Cruces, E., Toscano, R., Millan-Linares, M. C., Montserrat-de la Paz, S., & Martin, M. E. (2022). Acyclic Diterpene Phytol from Hemp Seed Oil (Cannabis sativa L.) Exerts Anti-Inflammatory Activity on Primary Human Monocytes-Macrophages. Foods, 11(15), 2366. https://doi.org/10.3390/foods11152366