Transcriptome and Metabolome Profiling to Explore the Causes of Purple Leaves Formation in Non-Heading Chinese Cabbage (Brassica rapa L. ssp. chinensis Makino var. mutliceps Hort.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Total Anthocyanins Analysis
2.3. Extraction and Separation of Anthocyanins
2.4. Metabolite Identification and Quantification
2.5. RNA-Seq Analysis
2.6. qRT-PCR Analysis
2.7. Statistical Analysis
3. Results
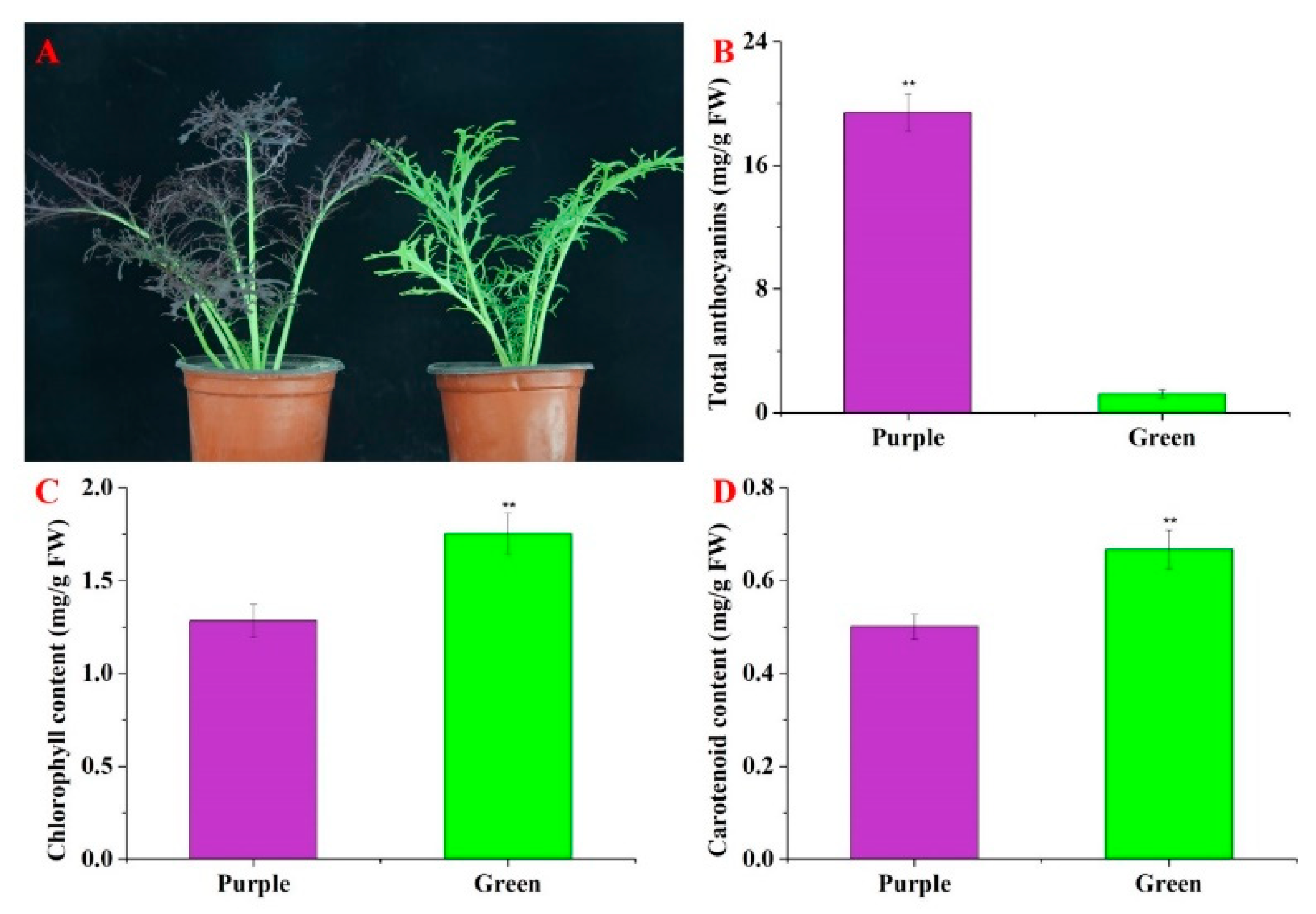
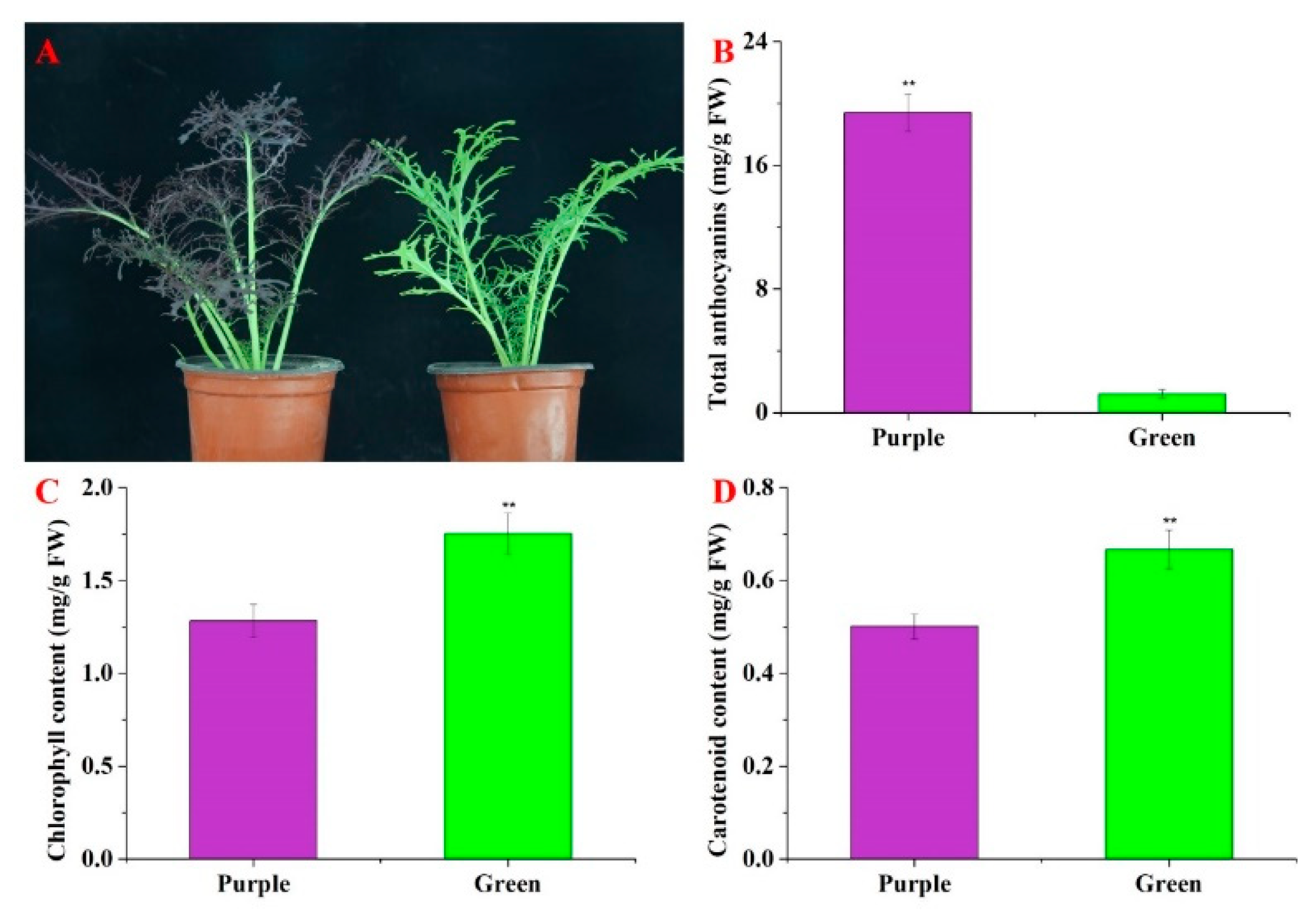
3.1. Identifications of Phenotype and Pigment Content in Purple Non-Heading Chinese Cabbage and Its Green Mutant
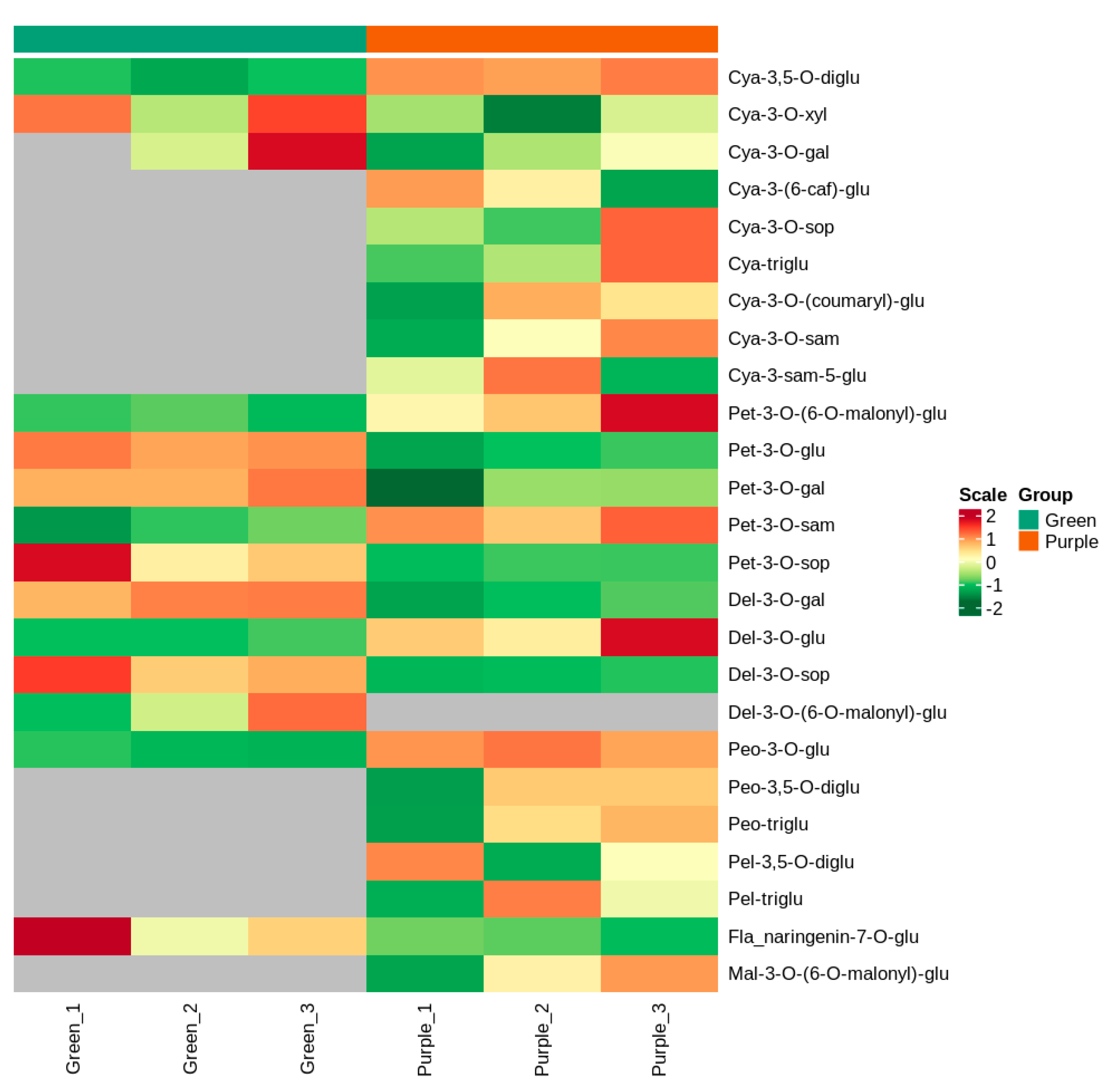
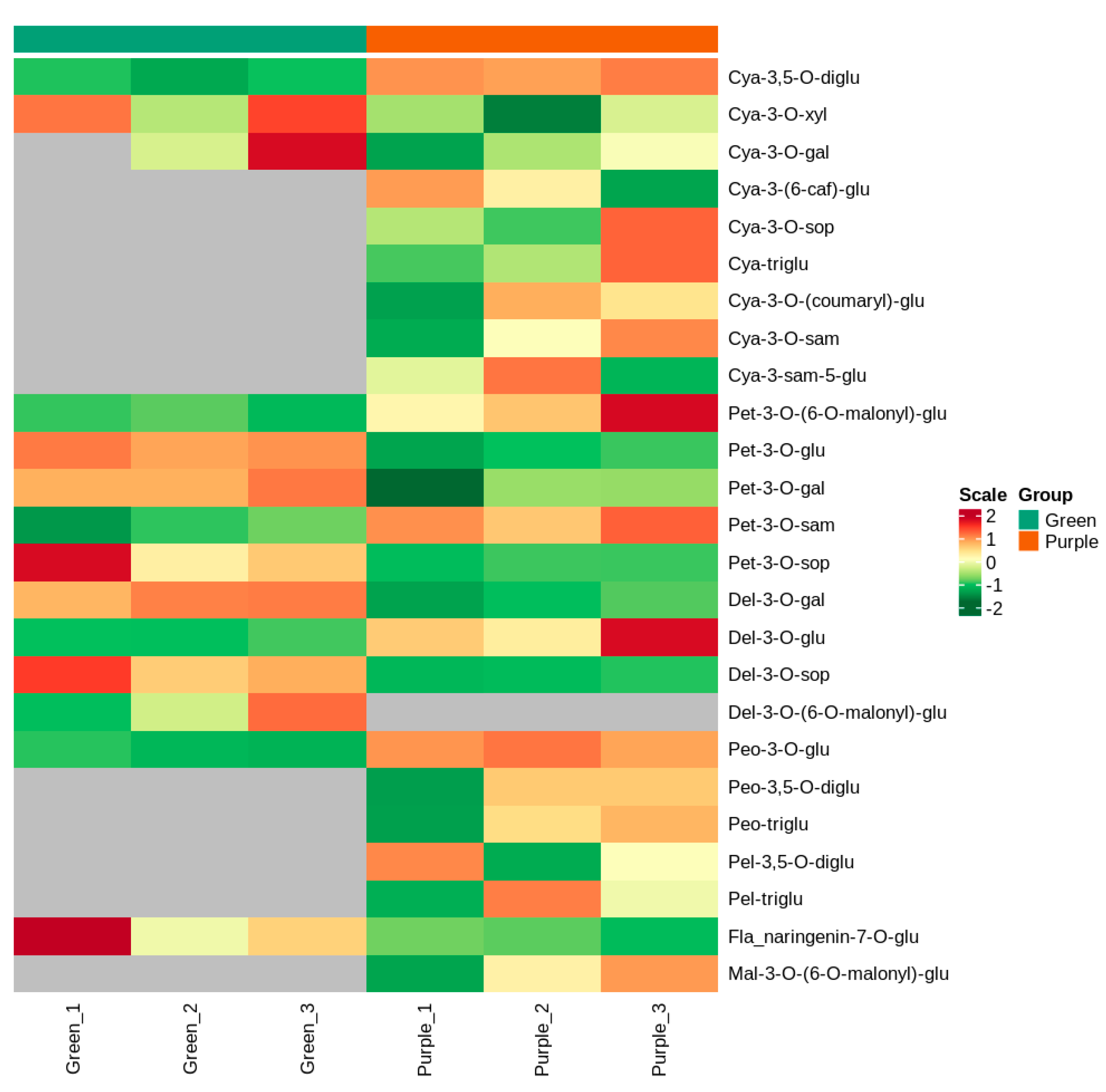
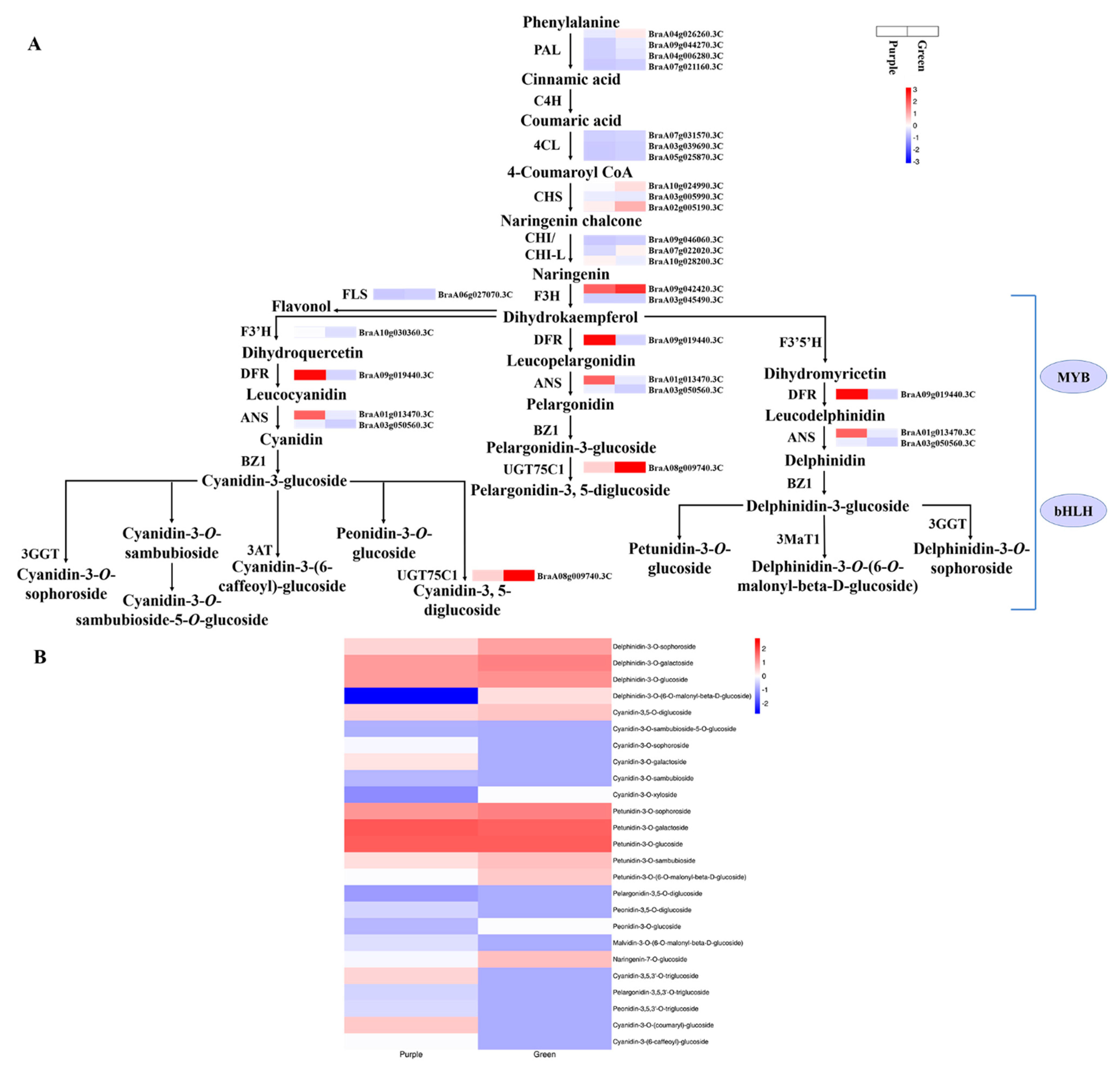
3.2. Identification of Anthocyanin Metabolites from the Leaves of Non-Heading Chinese Cabbage
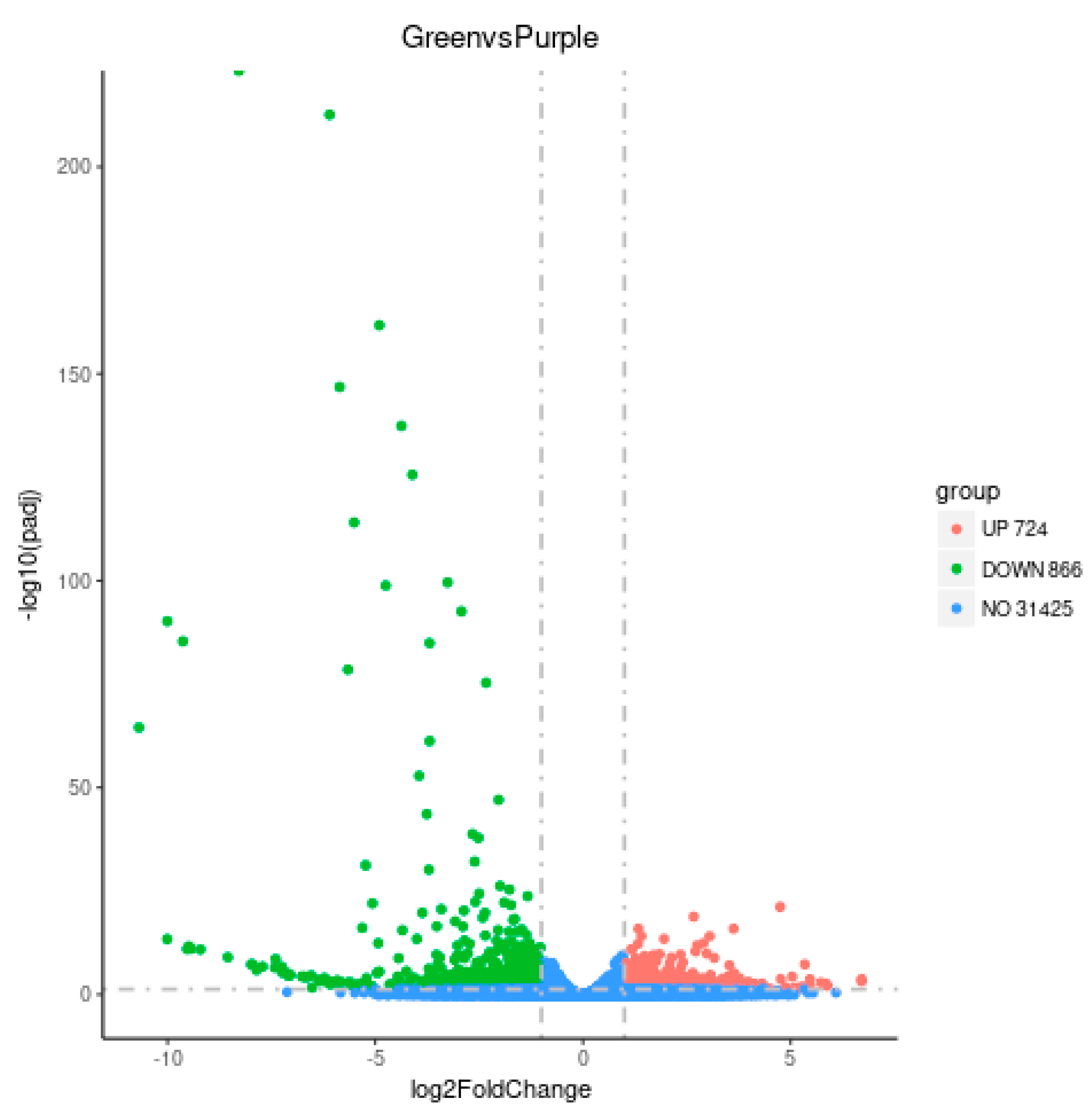
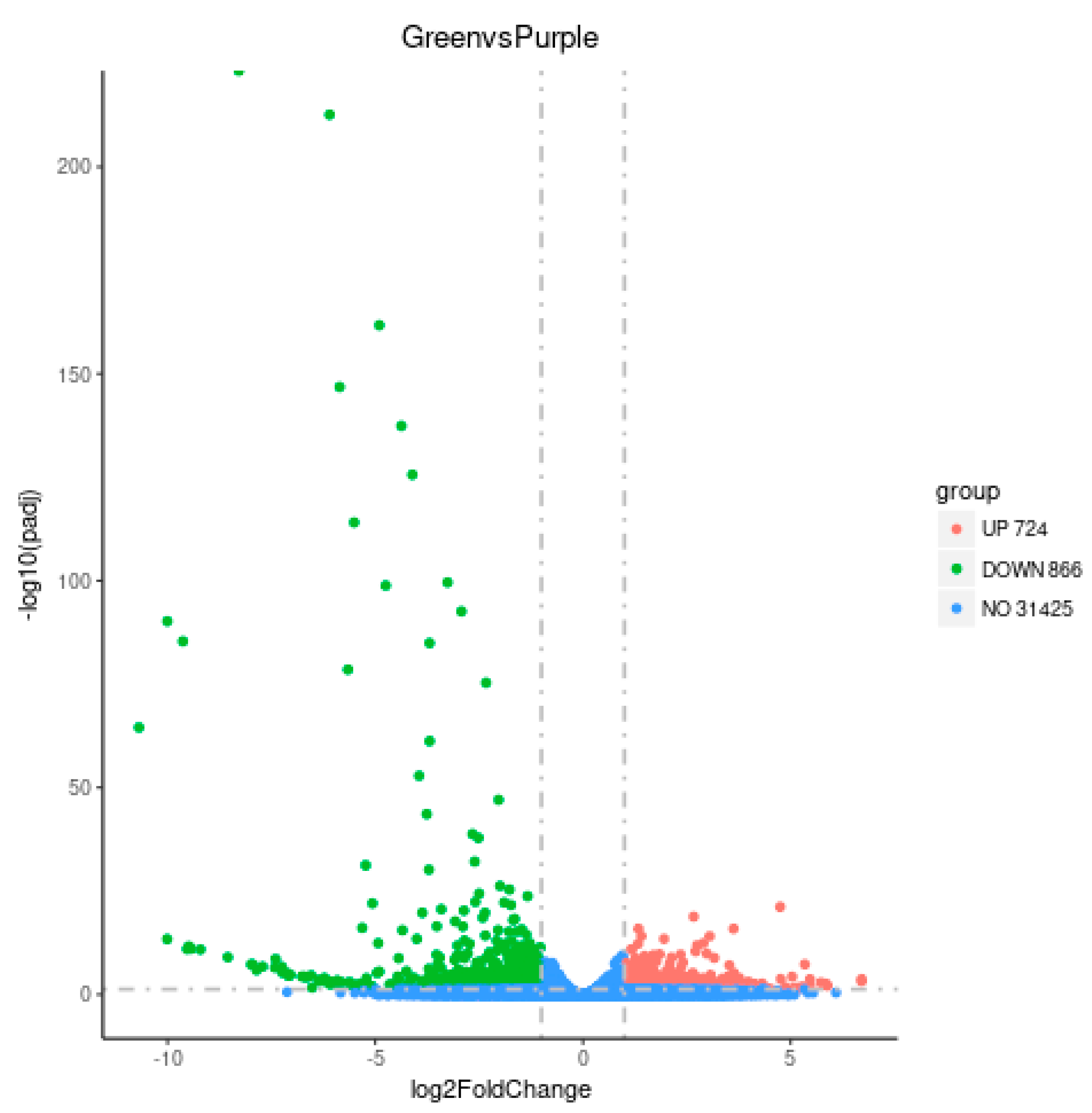
3.3. Transcriptome Analysis of Purple Non-Heading Chinese Cabbage and Its Green Mutant
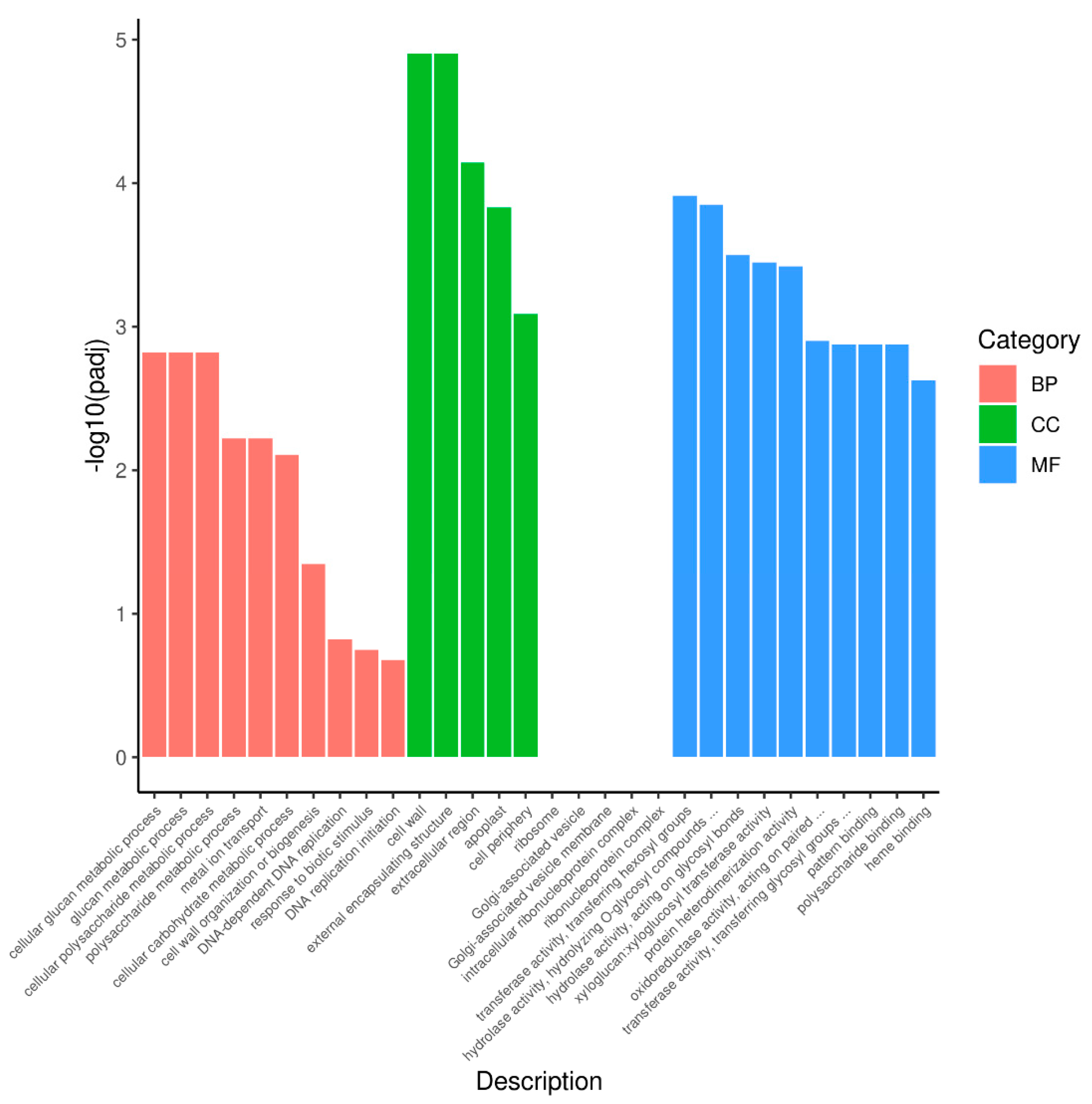
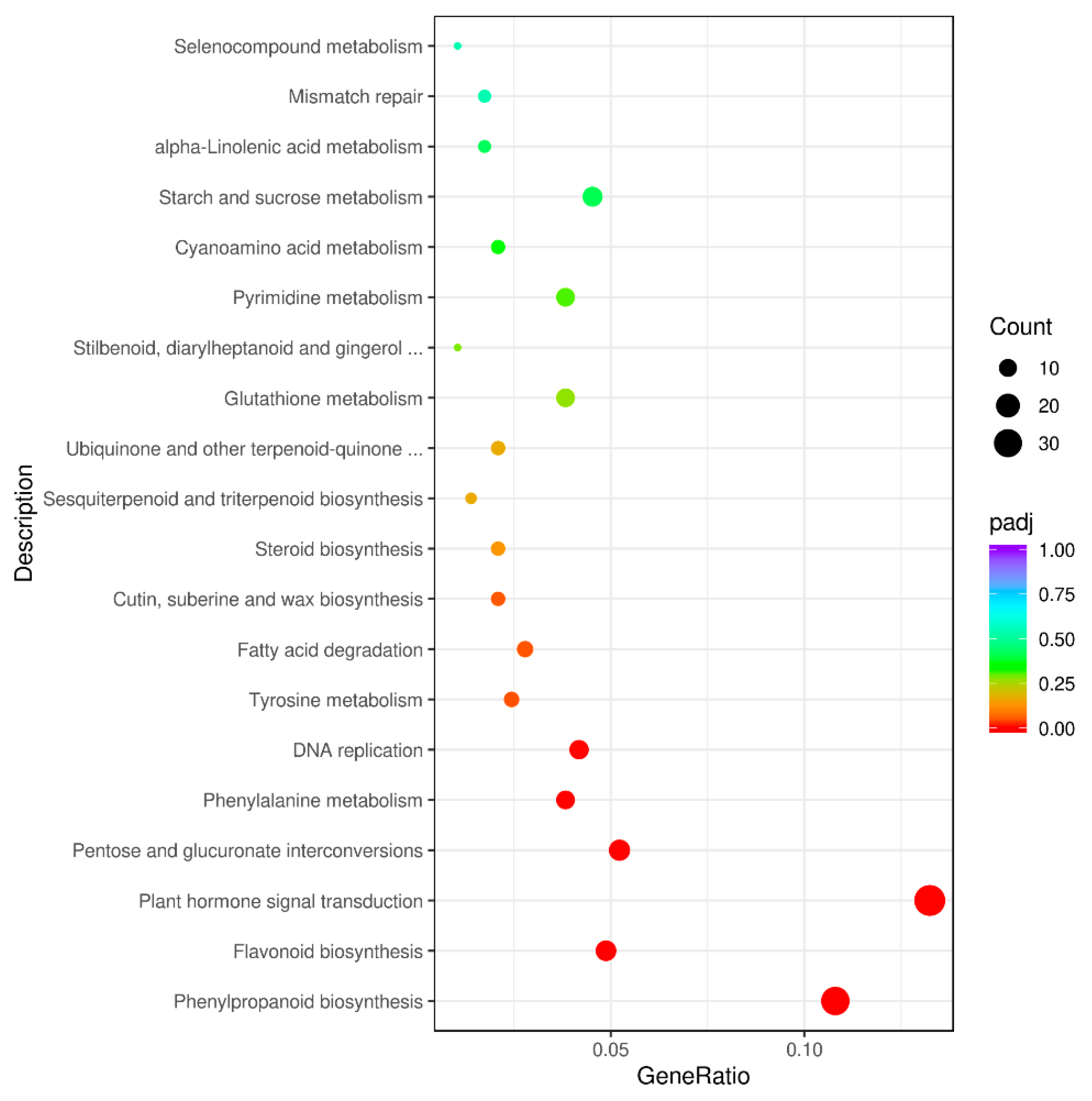
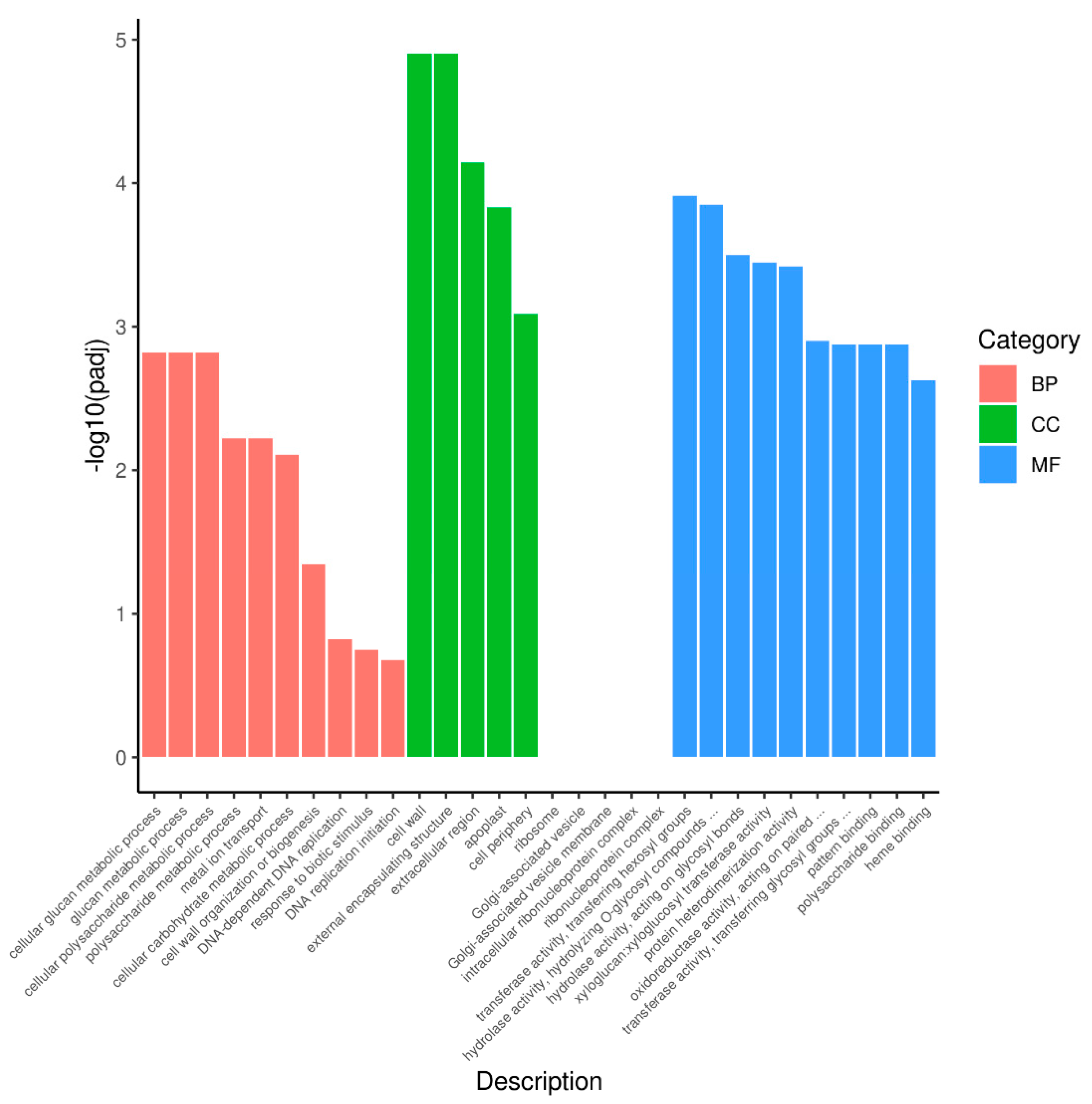
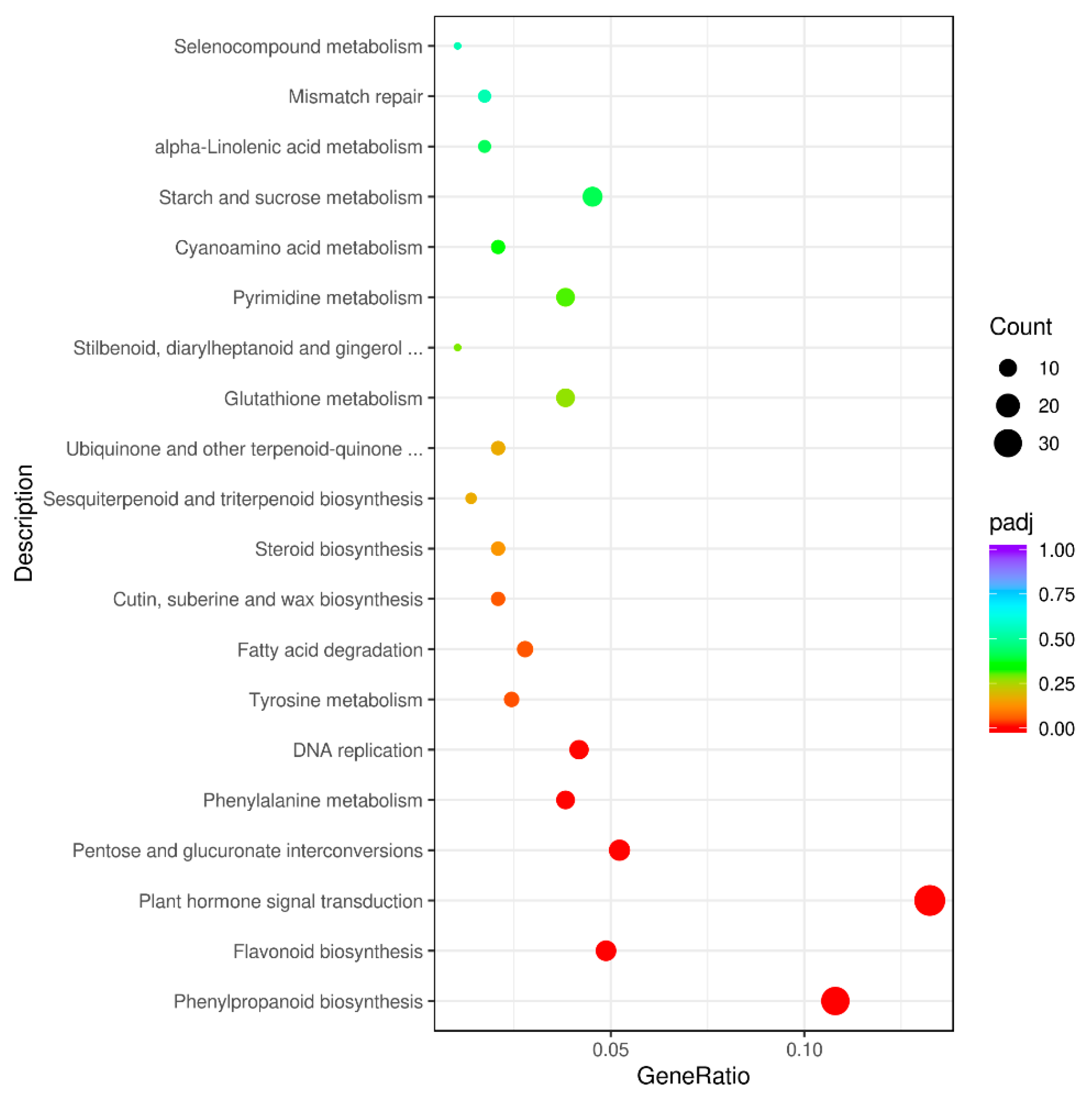
3.4. GO and KEGG Analysis of DEGs between Purple Non-Heading Chinese Cabbage and Its Green Mutant
3.5. Anthocyanin Biosynthetic DEGs in Purple Non-Heading Chinese Cabbage and Its Green Mutant
3.6. Analysis of Transcriptome Factors
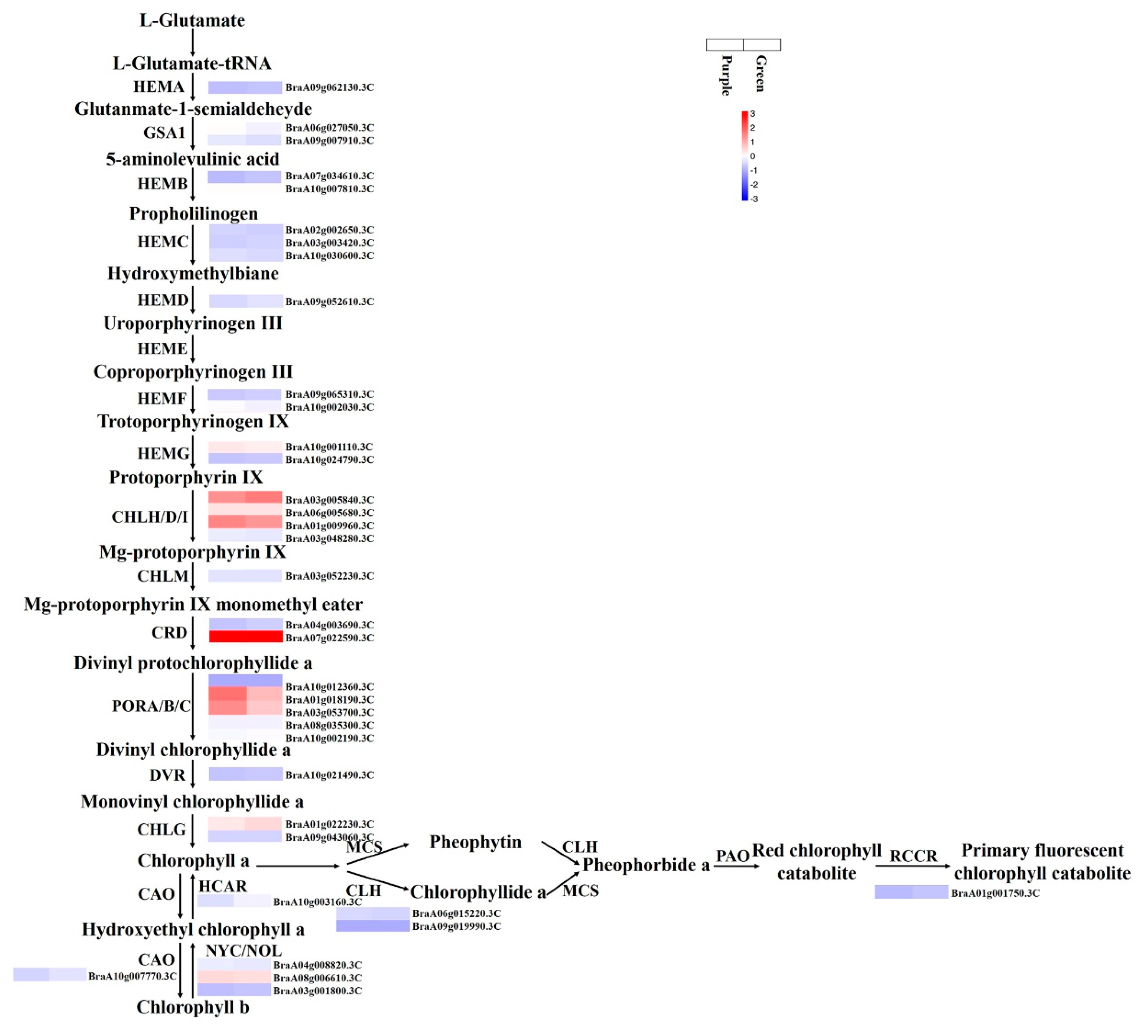
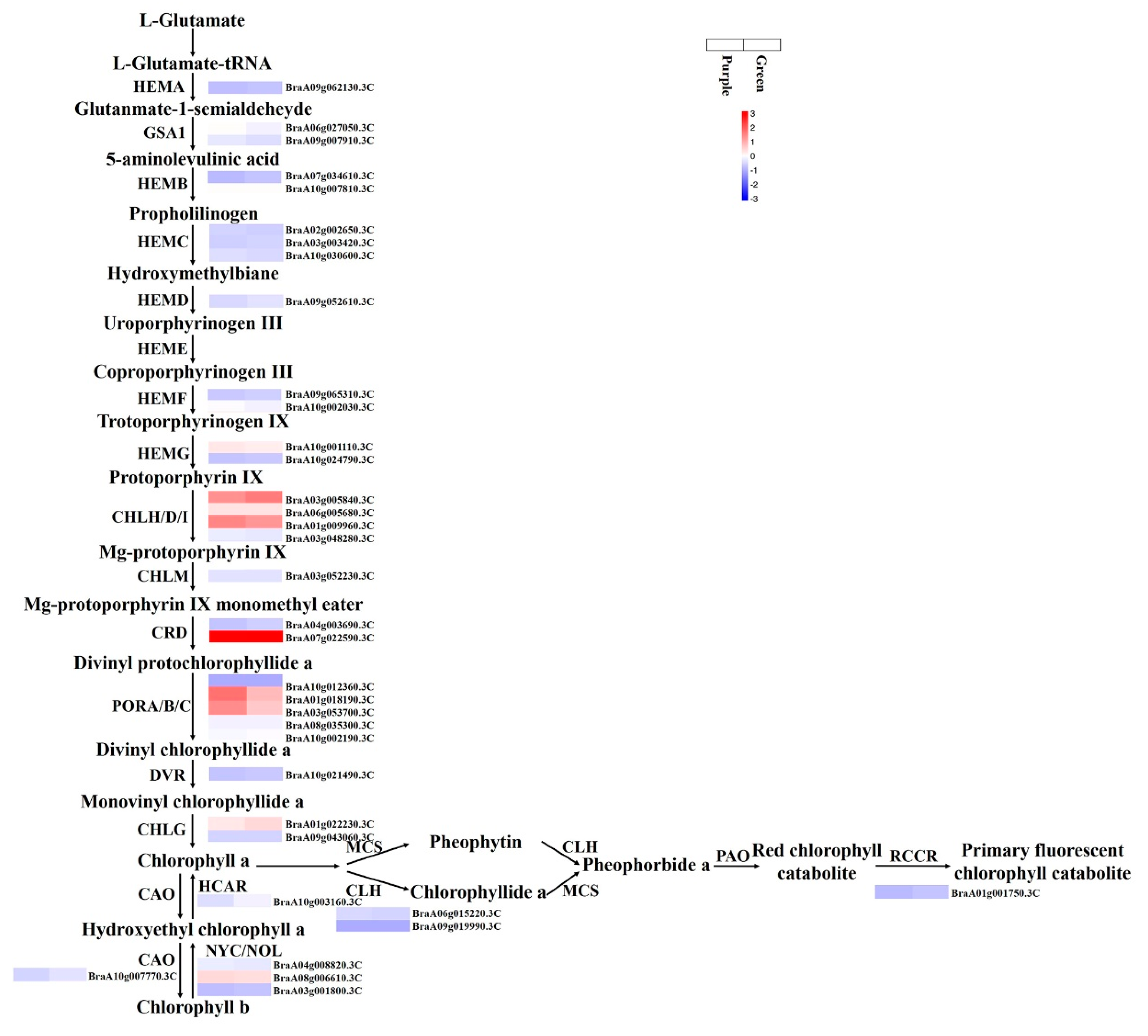
3.7. Expression Patterns of the Genes Involved in Chlorophyll Biosynthesis
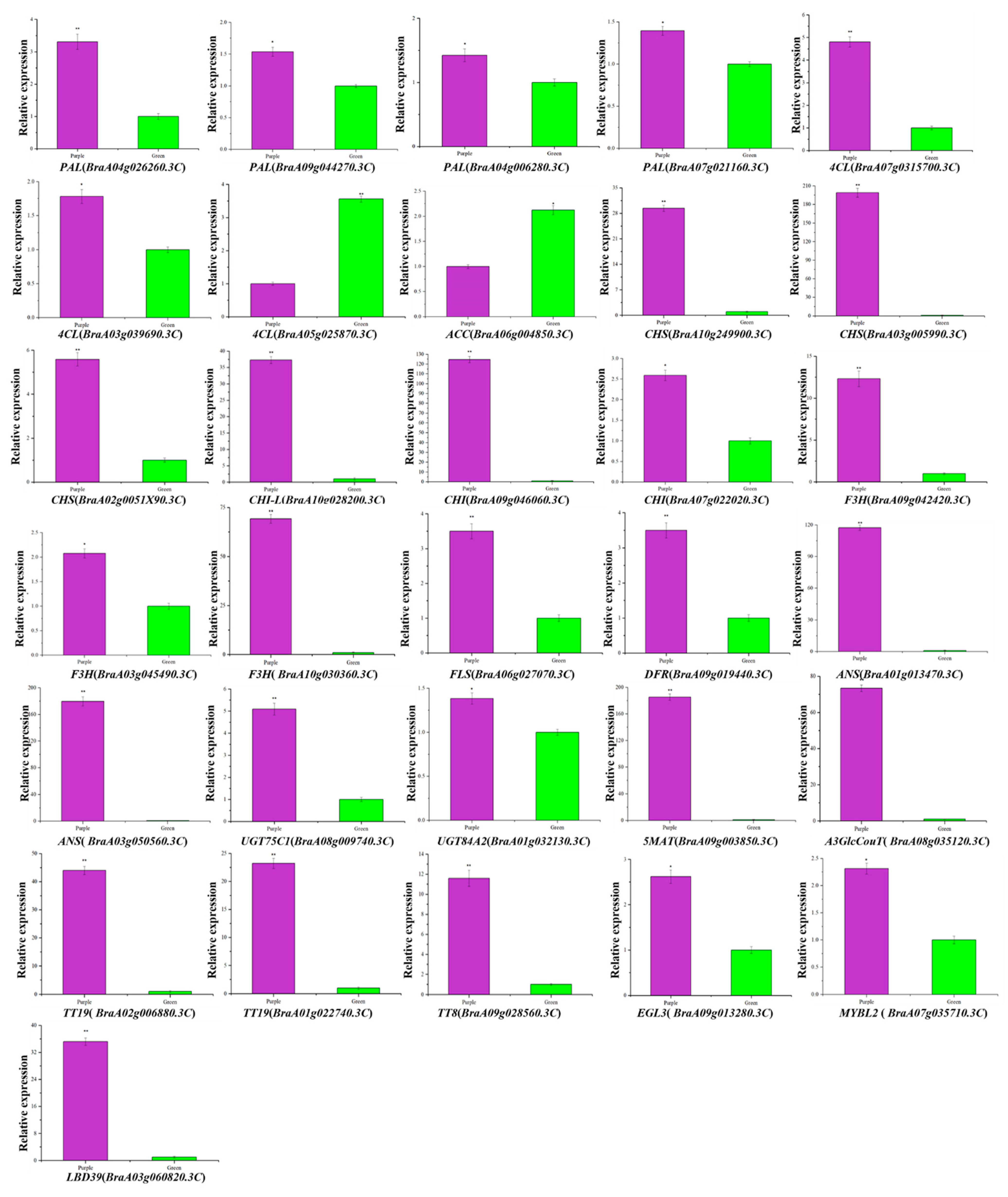
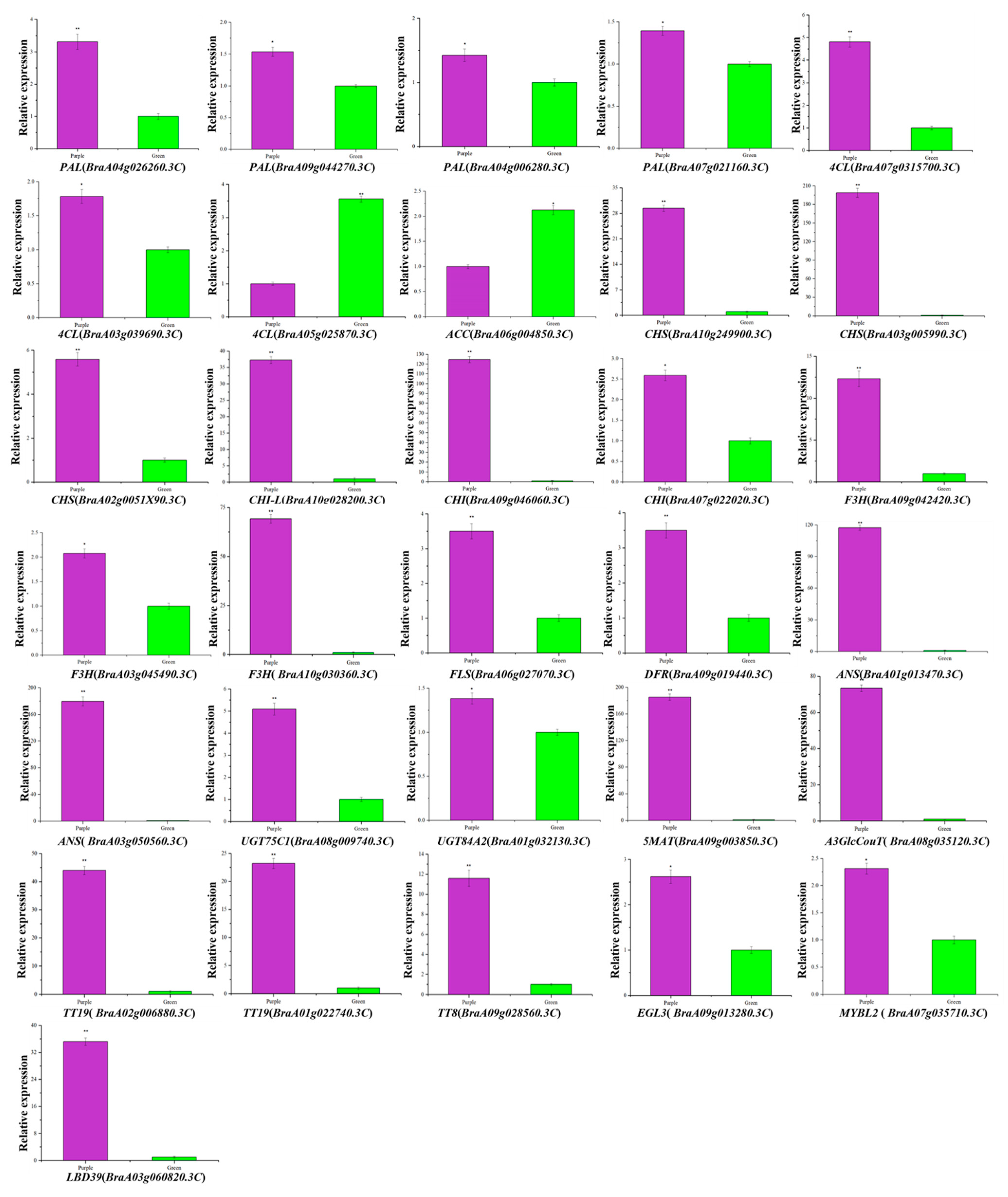
3.8. Validation of the Transcriptomic Data by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abdel-Aal, E.; Young, J.C.; Rabalski, I. Anthocyanin composition in black, blue, pink, purple, and red cereal grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef]
- Li, Y.; Shan, X.; Tong, L.; Wei, C.; Lu, K.; Li, S.; Kimani, S.; Wang, S.; Wang, L.; Gao, X. The conserved and particular roles of the R2R3-MYB regulator FhPAP1 from freesia hybrida in flower anthocyanin biosynthesis. Plant Cell Physiol. 2020, 61, 1365–1380. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Premathilake, A.T.; Gao, Y.; Yu, W.; Tao, R.; Teng, Y.; Bai, S. Ethylene-activated PpERF105 induces the expression of the repressor-type R2R3-MYB gene PpMYB140 to inhibit anthocyanin biosynthesis in red pear fruit. Plant J. 2021, 105, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Gould, K.S. Muriel wheldale onslow and the rediscovery of anthocyanin function in plants. Recent Adv. Polyphen. Res. 2010, 2, 206–225. [Google Scholar] [CrossRef]
- He, Q.; Ren, Y.; Zhao, W.; Li, R.; Zhang, L. Low temperature promotes anthocyanin biosynthesis and related gene expression in the seedlings of purple head Chinese cabbage (Brassica rapa L.). Genes 2020, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Fang, H.; Wang, J.; Yue, X.; Su, M.; Mao, Z.; Zou, Q.; Jiang, H.; Guo, Z.; Yu, L.; et al. Ultraviolet B-induced MdWRKY72 expression promotes anthocyanin synthesis in apple. Plant Sci. 2020, 292, 110377. [Google Scholar] [CrossRef]
- He, Q.; Zhang, Z.; Zhang, L. Anthocyanin accumulation, antioxidant ability and stability, and a transcriptional analysis of anthocyanin biosynthesis in purple heading Chinese cabbage (Brassica rapa L. ssp. pekinensis). J. Agric. Food Chem. 2016, 64, 132–145. [Google Scholar] [CrossRef]
- Lin, B.W.; Gong, C.C.; Song, H.F.; Cui, Y.Y. Effects of anthocyanins on the prevention and treatment of cancer. Br. J. Pharmacol. 2017, 174, 1226–1243. [Google Scholar] [CrossRef] [Green Version]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Liu, Z.; Chen, W.; Xu, H.; Feng, H. Anthocyanin degrading and chlorophyll accumulation lead to the formation of bicolor leaf in ornamental kale. Int. J. Mol. Sci. 2019, 20, 603. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hu, Z.; Zhu, M.; Zhu, Z.; Wang, Z.; Tian, S.; Chen, G. Anthocyanin accumulation and molecular analysis of correlated genes in purple kohlrabi (Brassica oleracea var. gongylodes L.). J. Agric. Food Chem. 2015, 63, 4160–4169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, L.; Liu, Z.; Zhao, Z.; Zhao, J.; Wang, Z.; Zhou, G.; Liu, P.; Liu, M. Transcriptome and metabolome profiling unveil the mechanisms of Ziziphus jujuba Mill. peel coloration. Food Chem. 2020, 312, 125903. [Google Scholar] [CrossRef]
- Hiromoto, T.; Honjo, E.; Noda, N.; Tamada, T.; Kazuma, K.; Suzuki, M.; Blaber, M.; Kuroki, R. Structural basis for acceptor-substrate recognition of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase from Clitoria ternatea. Protein Sci. 2015, 24, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, X.; Gao, X.; Ao, M.; Wang, Q.; Yang, D.; Wang, M.; Fu, Y.; Wang, L. cDNA cloning and characterization of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase in Freesia hybrida. Plant Cell Rep. 2011, 30, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gong, J.; Chen, K.; Yao, W.; Zhang, B.; Wang, J.; Tian, S.; Liu, H.; Wang, Y.; Liu, Y.; et al. A novel R3 MYB transcriptional repressor, MaMYBx, finely regulates anthocyanin biosynthesis in grape hyacinth. Plant Sci. 2020, 298, 110588. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, C.; Zhang, M.; Yin, X.; Xu, C.; Chen, K. The MrWD40-1 gene of Chinese bayberry (Myrica rubra) interacts with MYB and bHLH to enhance anthocyanin accumulation. Plant Mol. Biol. Rep. 2013, 31, 1474–1484. [Google Scholar] [CrossRef]
- Wang, L.; Lu, W.; Ran, L.; Dou, L.; Yao, S.; Hu, J.; Fan, D.; Li, C.; Luo, K. R2R3-MYB transcription factor MYB6 promotes anthocyanin and proanthocyanidin biosynthesis but inhibits secondary cell wall formation in Populus tomentosa. Plant J. 2019, 99, 733–751. [Google Scholar] [CrossRef]
- Meng, X.; Yin, B.; Feng, H.L.; Zhang, S.; Liang, X.Q. Overexpression of R2R3-MYB gene leads to accumulation of anthocyanin and enhanced resistance to chilling and oxidative stress. Biol. Plant. 2014, 58, 121–130. [Google Scholar] [CrossRef]
- Dong, T.; Han, R.; Yu, J.; Zhu, M.; Zhang, Y.; Gong, Y.; Li, Z. Anthocyanins accumulation and molecular analysis of correlated genes by metabolome and transcriptome in green and purple asparaguses (Asparagus officinalis, L.). Food Chem. 2019, 271, 18–28. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 2008, 55, 940–953. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.W.; Rahim, M.A.; Jung, H.J.; Afrin, K.S.; Kim, H.T.; Park, J.I.; Kang, J.G.; Nou, I.S. Abscisic acid and ethylene biosynthesis-related genes are associated with anthocyanin accumulation in purple ornamental cabbage (Brassica oleracea var. acephala). Genome 2019, 62, 513–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhao, J.; Qin, M.; Ren, Y.; Zhang, H.; Dai, Z.; Hao, L.; Zhang, L. Genetic analysis and mapping of the purple gene in purple heading Chinese cabbage. Hortic. Plant J. 2016, 2, 351–356. [Google Scholar] [CrossRef]
- Hao, Y.; Wang, J.; Hu, C.; Zhou, Q.; Mubeen, H.M.; Hou, X. Regulation of BcMYB44 on anthocyanin synthesis and drought tolerance in non-heading Chinese cabbage (Brassica campestris ssp. chinensis Makino). Horticulturae 2022, 8, 351. [Google Scholar] [CrossRef]
- Kim, J.; Kim, D.H.; Lee, J.Y.; Lim, S.H. The R3-Type MYB transcription factor BrMYBL2.1 negatively regulates anthocyanin biosynthesis in Chinese cabbage (Brassica rapa L.) by repressing MYB-bHLH-WD40 complex activity. Int. J. Mol. Sci. 2022, 23, 3382. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Zhao, M.; Hu, Y.; Meng, F.; Song, X.; Tigabu, M.; Chiang, V.L.; Sederoff, R.; Ma, W.; et al. Molecular and metabolic insights into anthocyanin biosynthesis for leaf color change in chokecherry (Padus virginiana). Int. J. Mol. Sci. 2021, 22, 10697. [Google Scholar] [CrossRef]
- He, Q.; Lu, Q.; He, Y.; Wang, Y.; Zhang, N.; Zhao, W.; Zhang, L. Dynamic changes of the anthocyanin biosynthesis mechanism during the development of heading Chinese Cabbage (Brassica rapa L.) and Arabidopsis under the control of BrMYB2. Front. Plant Sci. 2020, 11, 593766. [Google Scholar] [CrossRef]
- Shen, J.; Zou, Z.; Zhang, X.; Zhou, L.; Wang, Y.; Fang, W.; Zhu, X. Metabolic analyses reveal different mechanisms of leaf color change in two purple-leaf tea plant (Camellia sinensis L.) cultivars. Hortic. Res. 2018, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.T.; Yu, Z.C.; Tang, J.W.; Cai, M.L.; Chen, Y.L.; Yang, C.W.; Chow, W.S.; Peng, C.L. The major photoprotective role of anthocyanins in leaves of Arabidopsis thaliana under long-term high light treatment: Antioxidant or light attenuator? Photosynth. Res. 2021, 149, 25–40. [Google Scholar] [CrossRef]
- Park, C.H.; Bong, S.J.; Lim, C.J.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of green and red mizuna (Brassica rapa L. var. japonica). Foods 2020, 9, 1079. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.W.; Zhou, X.; Burke, S.; Wu, X.; Prior, R.L.; Li, L. The purple cauliflower arises from activation of a MYB transcription factor. Plant Physiol. 2010, 154, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charron, C.S.; Clevidence, B.A.; Britz, S.J.; Novotny, J.A. Effect of dose size on bioavailability of acylated and nonacylated anthocyanins from red cabbage (Brassica oleracea L. Var. capitata). J. Agric. Food Chem. 2007, 55, 5354–5362. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Xu, H.; Chen, L.Z.; Fan, X.X.; Jing, Z.; Chen, S.; Xu, Z.G. Study of the relationship between leaf color formation and anthocyanin metabolism among different purple Pakchoi lines. Molecules 2020, 25, 4809. [Google Scholar] [CrossRef]
- Liu, C.; Yao, X.; Li, G.; Huang, L.; Xie, Z. Transcriptomic profiling of purple broccoli reveals light-induced anthocyanin biosynthetic signaling and structural genes. PeerJ 2020, 8, e8870. [Google Scholar] [CrossRef]
- Yuan, Y.; Chiu, L.W.; Li, L. Transcriptional regulation of anthocyanin biosynthesis in red cabbage. Planta 2009, 230, 1141–1153. [Google Scholar] [CrossRef]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in Arabidopsis: Structural and genetic diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.C.; Hu, G.B.; Hu, F.C.; Wang, H.C.; Yang, Z.Y.; Lai, B. The UDP glucose: Flavonoid-3-O-glucosyltransferase (UFGT) gene regulates anthocyanin biosynthesis in litchi (Litchi chinesis Sonn.) during fruit coloration. Mol. Biol. Rep. 2012, 39, 6409–6415. [Google Scholar] [CrossRef]
- Offen, W.; Martinez-Fleites, C.; Yang, M.; Kiat-Lim, E.; Davis, B.G.; Tarling, C.A.; Ford, C.M.; Bowles, D.J.; Davies, G.J. Structure of a flavonoid glucosyltransferase reveals the basis for plant natural product modification. EMBO J. 2006, 25, 1396–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitayama, M.; et al. Functional genomics by integrated analysis of metabolome and transcriptome of Arabidopsis plants over-expressing an MYB transcription factor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wu, J.; Zheng, S.; Cheng, F.; Liu, B.; Liang, J.; Cui, Y.; Wang, X. Anthocyanin profile characterization and quantitative trait locus mapping in zicaitai (Brassica rapa L. ssp. chinensis var. purpurea). Mol. Breed. 2015, 35, 113. [Google Scholar] [CrossRef]
- Li, G.H.; Chen, H.C.; Liu, J.L.; Luo, W.L.; Xie, D.S.; Luo, S.B.; Wu, T.Q.; Akram, W.; Zhong, Y.J. A high-density genetic map developed by specific-locus amplified fragment (SLAF) sequencing and identification of a locus controlling anthocyanin pigmentation in stalk of Zicaitai (Brassica rapa L. ssp. chinensis var. purpurea). BMC Genom. 2019, 20, 343. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, G.; Dong, T.; Pan, Y.; Zhao, Z.; Tian, S.; Hu, Z. Anthocyanin accumulation and transcriptional regulation of anthocyanin biosynthesis in purple Bok Choy (Brassica rapa var. chinensis). J. Agric. Food Chem. 2014, 62, 12366–12376. [Google Scholar] [CrossRef]
- Song, H.; Yi, H.; Lee, M.; Han, C.T.; Lee, J.; Kim, H.; Park, J.I.; Nou, I.S.; Kim, S.J.; Hur, Y. Purple Brassica oleracea var. capitata F. rubra is due to the loss of BoMYBL2-1 expression. BMC Plant Biol. 2018, 18, 82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Content in Purple (µg/g) | Content in Green (µg/g) | Fold Change | ||
|---|---|---|---|---|---|
| Mean | SE | Mean | SE | ||
| Delphinidin-3-O-sophoroside | 3.3800 | 0.0617 | 6.4200 | 0.3990 | 1.9000 |
| Delphinidin-3-O-galactoside | 26.4000 | 1.3500 | 48.6000 | 1.1900 | 1.8400 |
| Delphinidin-3-O-glucoside | 27.8000 | 2.5900 | 17.2000 | 0.1860 | 0.6190 |
| Delphinidin-3-O-(6-O-malonyl-beta-D-glucoside) | N/A | N/A | 0.129 | 0.0044 | N/A |
| Cyanidin-3,5-O-diglucoside | 3.3300 | 0.0814 | 0.5810 | 0.1200 | 0.1750 |
| Cyanidin-3-O-sambubioside-5-O-glucoside | 0.0316 | 0.0002 | N/A | N/A | N/A |
| Cyanidin-3-O-sophoroside | 0.4870 | 0.0155 | N/A | N/A | N/A |
| Cyanidin-3-O-galactoside | 1.9100 | 0.1540 | 2.4600 | 0.4050 | 1.2800 |
| Cyanidin-3-O-sambubioside | 0.0385 | 0.0022 | N/A | N/A | N/A |
| Cyanidin-3-O-xyloside | 0.0068 | 0.0006 | 0.0091 | 0.0009 | 1.3500 |
| Cyanidin-3-O-(coumaryl)-glucoside | 4.8400 | 0.3150 | N/A | N/A | N/A |
| Cyanidin-3-(6-caffeoyl)-glucoside | 0.5440 | 0.0175 | N/A | N/A | N/A |
| Cyanidin-3,5,3’-O-triglucoside | 3.0200 | 0.0764 | N/A | N/A | N/A |
| Petunidin-3-O-sophoroside | 31.9000 | 0.6510 | 59.70000 | 6.9300 | 1.8700 |
| Petunidin-3-O-galactoside | 377.0000 | 15.8000 | 454.0000 | 4.3300 | 1.2100 |
| Petunidin-3-O-glucoside | 284.0000 | 15.9000 | 577.0000 | 10.1000 | 2.0300 |
| Petunidin-3-O-sambubioside | 2.0100 | 0.1160 | 0.7390 | 0.1190 | 0.3670 |
| Petunidin-3-O-(6-O-malonyl-beta-D-glucoside) | 0.6030 | 0.0523 | 0.4020 | 0.0076 | 0.6670 |
| Peonidin-3,5,3’-O-triglucoside | 0.1320 | 0.0044 | N/A | N/A | N/A |
| Peonidin-3,5-O-diglucoside | 0.1170 | 0.0030 | N/A | N/A | N/A |
| Peonidin-3-O-glucoside | 0.0350 | 0.0009 | 0.0109 | 0.0007 | 0.3110 |
| Malvidin-3-O-(6-O-malonyl-beta-D-glucoside) | 0.1670 | 0.0056 | N/A | N/A | N/A |
| Naringenin-7-O-glucoside | 0.4640 | 0.0214 | 0.8780 | 0.1420 | 1.8900 |
| Pelargonidin-3,5-O-diglucoside | 0.0128 | 0.0006 | N/A | N/A | N/A |
| Pelargonidin-3,5,3’-O-triglucoside | 0.1110 | 0.0006 | N/A | N/A | N/A |
| Gene ID | Gene Name | Average FPKM of Purple | Average FPKM of Green | Log2 Fold Change | p Value | Padj |
|---|---|---|---|---|---|---|
| BraA04g026260.3C | PAL | 123.39 | 33.22 | −1.89 | 6.59 × 10−26 | 6.07 × 10−23 |
| BraA09g044270.3C | PAL | 31.13 | 11.23 | −1.47 | 3.30 × 10−19 | 1.92 × 10−16 |
| BraA04g006280.3C | PAL | 25.71 | 9.26 | −1.48 | 4.66 × 10−15 | 1.80 × 10−12 |
| BraA07g021160.3C | PAL | 1.42 | 0.58 | −1.28 | 1.19 × 10−3 | 1.44 × 10−2 |
| BraA07g031570.3C | 4CL | 18.39 | 3.47 | −2.41 | 3.67 × 10−22 | 2.62 × 10−19 |
| BraA03g039690.3C | 4CL | 4.28 | 1.50 | −1.52 | 1.66 × 10−4 | 3.14 × 10−3 |
| BraA05g025870.3C | 4CL | 0.01 | 0.25 | 4.41 | 5.38 × 10−3 | 4.43 × 10−2 |
| BraA06g004850.3C | ACC | 0.02 | 0.21 | 3.13 | 1.33 × 10−5 | 4.40 × 10−4 |
| BraA10g024990.3C | CHS | 208.44 | 37.86 | −2.46 | 3.81 × 10−7 | 2.53 × 10−5 |
| BraA03g005990.3C | CHS | 154.68 | 11.44 | −3.76 | 1.35 × 10−5 | 4.44 × 10−4 |
| BraA02g005190.3C | CHS | 297.88 | 58.44 | −2.35 | 4.82 × 10−8 | 4.65 × 10−6 |
| BraA09g046060.3C | CHI | 3.56 | 0 | −7.94 | 4.01 × 10−10 | 6.53 × 10−8 |
| BraA07g022020.3C | CHI | 59.44 | 28.25 | −1.08 | 2.50 × 10−4 | 4.28 × 10−3 |
| BraA10g028200.3C | CHI-L | 281.65 | 13.87 | −4.35 | 5.12 × 10−19 | 2.86 × 10−16 |
| BraA09g042420.3C | 854.91 | 112.69 | −2.92 | 9.14 × 10−97 | 2.61 × 10−93 | |
| BraA03g045490.3C | F3H | 21.68 | 1.43 | −3.94 | 8.93 × 10−57 | 1.41 × 10−53 |
| BraA10g030360.3C | F3’H | 215.68 | 7.08 | −4.93 | 9.41 × 10−16 | 4.04 × 10−13 |
| BraA06g027070.3C | FLS | 1.55 | 0.22 | −2.80 | 1.57 × 10−4 | 3.01 × 10−3 |
| BraA09g019440.3C | DFR | 1194.67 | 3.87 | −8.28 | 0 | 0 |
| BraA01g013470.3C | ANS | 869.00 | 12.68 | −6.10 | 1.75 × 10−217 | 2.49 × 10−213 |
| BraA03g050560.3C | ANS | 147.04 | 0.19 | −9.62 | 1.90 × 10−89 | 4.53 × 10−86 |
| BraA08g009740.3C | UGT75C1 | 428.73 | 137.21 | −1.64 | 7.75 × 10−22 | 5.40 × 10−19 |
| BraA01g032130.3C | UGT84A2 | 21.56 | 6.75 | −1.68 | 1.56 × 10−21 | 1.06 × 10−18 |
| BraA09g003850.3C | 5MAT | 145.89 | 0.09 | −10.69 | 1.66 × 10−68 | 2.95 × 10−65 |
| BraA08g035120.3C | A3GlcCouT | 323.27 | 5.60 | −5.86 | 2.19 × 10−151 | 1.56 × 10−147 |
| BraA02g006880.3C | TT19 | 567.41 | 99.37 | −2.51 | 1.09 × 10−41 | 1.42 × 10−38 |
| BraA10g022740.3C | TT19 | 457.80 | 22.22 | −4.37 | 6.96 × 10−142 | 3.97 × 10−138 |
| BraA09g028560.3C | TT8 | 56.12 | 1.88 | −4.90 | 1.98 × 10−166 | 1.88 × 10−162 |
| BraA09g013280.3C | EGL3 | 1.33 | 0.42 | −1.67 | 1.01 × 10−3 | 1.28 × 10−2 |
| BraA07g035710.3C | MYBL2 | 128.88 | 30.63 | −2.08 | 2.04 × 10−16 | 9.38 × 10−14 |
| BraA03g060820.3C | LBD39 | 4.69 | 0.66 | −2.83 | 2.51 × 10−6 | 1.17 × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Qi, X.; Liu, Z.; Zheng, W.; Guan, J.; Liu, Z.; Ren, J.; Feng, H.; Zhang, Y. Transcriptome and Metabolome Profiling to Explore the Causes of Purple Leaves Formation in Non-Heading Chinese Cabbage (Brassica rapa L. ssp. chinensis Makino var. mutliceps Hort.). Foods 2022, 11, 1787. https://doi.org/10.3390/foods11121787
Zhao Y, Qi X, Liu Z, Zheng W, Guan J, Liu Z, Ren J, Feng H, Zhang Y. Transcriptome and Metabolome Profiling to Explore the Causes of Purple Leaves Formation in Non-Heading Chinese Cabbage (Brassica rapa L. ssp. chinensis Makino var. mutliceps Hort.). Foods. 2022; 11(12):1787. https://doi.org/10.3390/foods11121787
Chicago/Turabian StyleZhao, Ying, Xinghua Qi, Zeji Liu, Wenfeng Zheng, Jian Guan, Zhiyong Liu, Jie Ren, Hui Feng, and Yun Zhang. 2022. "Transcriptome and Metabolome Profiling to Explore the Causes of Purple Leaves Formation in Non-Heading Chinese Cabbage (Brassica rapa L. ssp. chinensis Makino var. mutliceps Hort.)" Foods 11, no. 12: 1787. https://doi.org/10.3390/foods11121787
APA StyleZhao, Y., Qi, X., Liu, Z., Zheng, W., Guan, J., Liu, Z., Ren, J., Feng, H., & Zhang, Y. (2022). Transcriptome and Metabolome Profiling to Explore the Causes of Purple Leaves Formation in Non-Heading Chinese Cabbage (Brassica rapa L. ssp. chinensis Makino var. mutliceps Hort.). Foods, 11(12), 1787. https://doi.org/10.3390/foods11121787