
Anticholinesterase Activities of Different Solvent Extracts of Brewer’s Spent Grain



Abstract

1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
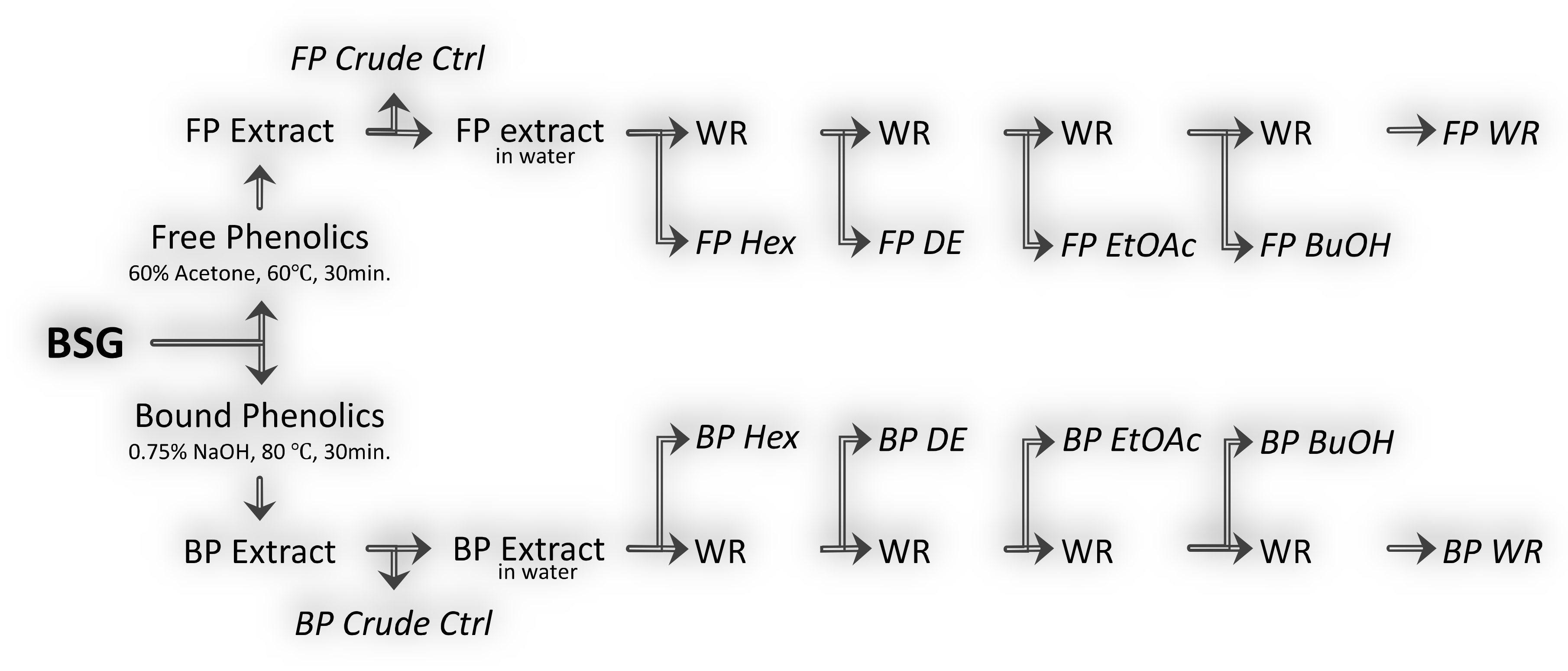
2.2. Solid–Liquid Extraction of Free and Bound Phenolics
2.3. Liquid–Liquid Partitioning of Free and Bound Phenolic Extracts
2.4. Determination of Polyphenolic Content
2.4.1. Total Phenolic Content (TPC) by Folin–Ciocalteu (FC)
2.4.2. Individual Polyphenol Quantification by UPLC-MS/MS
2.5. Preparation of Polyphenol Blends
2.6. Anti-Cholinesterase Assays
2.7. Statistical Analysis
3. Results and Discussion
3.1. Extraction Yield
3.2. Total Polyphenol Content
3.3. UPLC-MS/MS Quantification of BSG Free and Bound Polyphenols
3.4. Anti-AChE and -BChE Activities
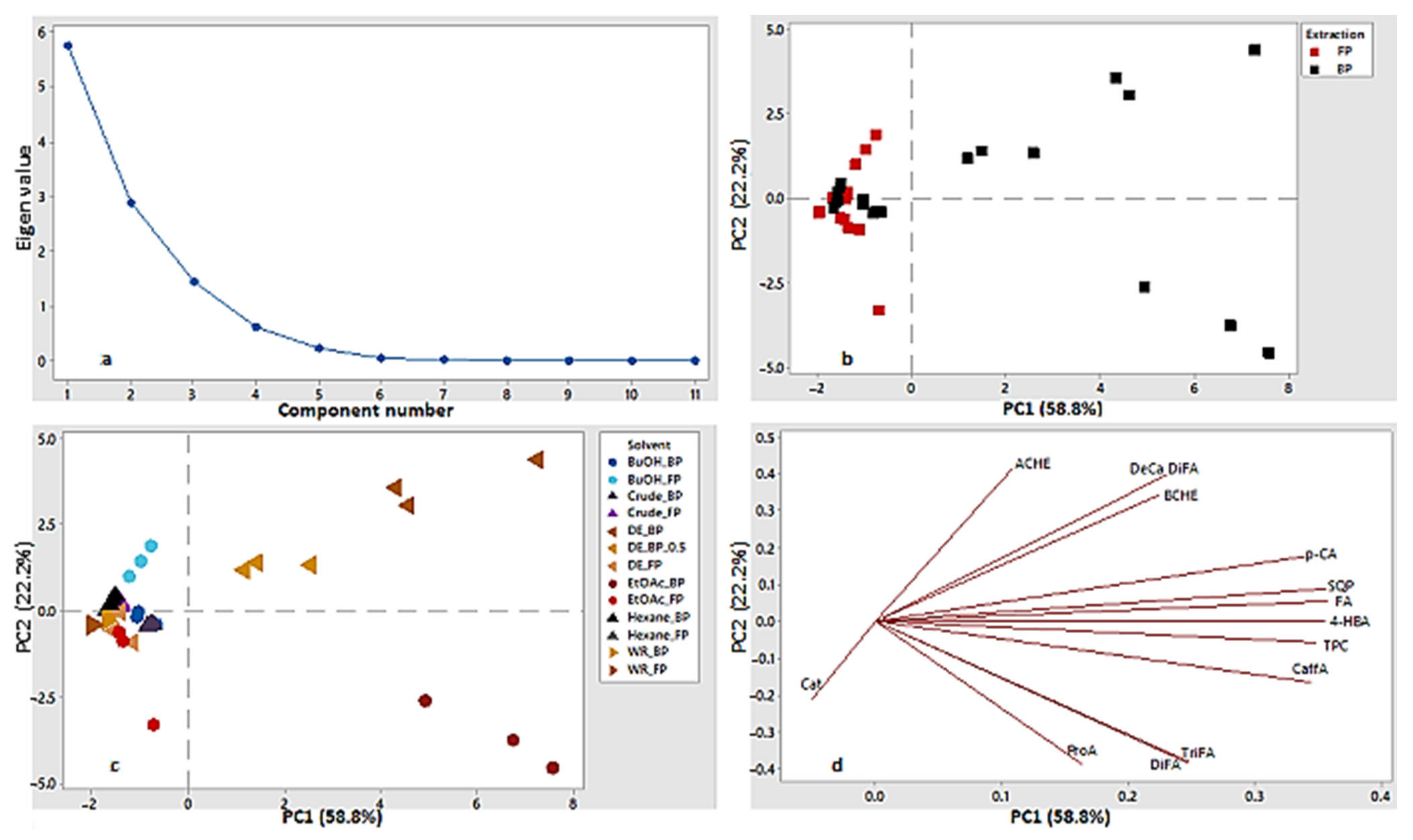
3.5. Pearson Correlation, Multiple Regression Model of Variables and PCA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Koch, W. Dietary Polyphenols—Important Non-Nutrients in the Prevention of Chronic Noncommunicable Diseases. A Systematic Review. Nutrients 2019, 11, 1039. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.; Gaspar, A.; Garrido, E.M.; Garrido, J.; Borges, F. Hydroxycinnamic acid antioxidants: An electrochemical overview. BioMed Res. Int. 2013, 2013, 251754. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.U.; Choudhary, M.I. (Eds.) Drug Design and Discovery in Alzheimer’s Disease; Elsevier: Amsterdam, The Netherlands, 2015; p. 784. [Google Scholar]
- Cavdar, H.; Senturk, M.; Guney, M.; Durdagi, S.; Kayik, G.; Supuran, C.T.; Ekinci, D. Inhibition of acetylcholinesterase and butyrylcholinesterase with uracil derivatives: Kinetic and computational studies. J. Enzym. Inhib. Med. Chem. 2019, 34, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. CHOLINE|Properties and Determination. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 1251–1254. [Google Scholar]
- Westfall, T.C. Cholinergic Neurotransmission in the Autonomic and Somatic Motor Nervous System; Elsevier: Amsterdam, The Netherlands, 2009; pp. 827–834. [Google Scholar]
- Geula, C.; Darvesh, S. Butyrylcholinesterase, cholinergic neurotransmission and the pathology of Alzheimer’s disease. Drugs Today 2004, 40, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Idehen, E.; Tang, Y.; Sang, S. Bioactive phytochemicals in barley. J. Food Drug Anal. 2017, 25, 148–161. [Google Scholar] [CrossRef]
- Kim, D.; Nguyen, M.D.; Dobbin, M.M.; Fischer, A.; Sananbenesi, F.; Rodgers, J.T.; Delalle, I.; Baur, J.A.; Sui, G.; Armour, S.M. SIRT1 deacetylase protects against neurodegeneration in models for Alzheimer’s disease and amyotrophic lateral sclerosis. EMBO J. 2007, 26, 3169–3179. [Google Scholar] [CrossRef]
- Hartman, R.E.; Shah, A.; Fagan, A.M.; Schwetye, K.E.; Parsadanian, M.; Schulman, R.N.; Finn, M.B.; Holtzman, D.M. Pomegranate juice decreases amyloid load and improves behavior in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2006, 24, 506–515. [Google Scholar] [CrossRef]
- Hu, N.; Yu, J.-T.; Tan, L.; Wang, Y.-L.; Sun, L.; Tan, L. Nutrition and the risk of Alzheimer’s disease. BioMed Res. Int. 2013, 2013, 524820. [Google Scholar] [CrossRef]
- Bouzaiene, N.N.; Jaziri, S.K.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The effects of caffeic, coumaric and ferulic acids on proliferation, superoxide production, adhesion and migration of human tumor cells in vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef]
- Kumar, N.; Pruthi, V. Potential applications of ferulic acid from natural sources. Biotechnol. Rep. 2014, 4, 86–93. [Google Scholar] [CrossRef]
- Yabe, T.; Hirahara, H.; Harada, N.; Ito, N.; Nagai, T.; Sanagi, T.; Yamada, H. Ferulic acid induces neural progenitor cell proliferation in vitro and in vivo. Neuroscience 2010, 165, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Winter, A.N.; Brenner, M.C.; Punessen, N.; Snodgrass, M.; Byars, C.; Arora, Y.; Linseman, D.A. Comparison of the neuroprotective and anti-inflammatory effects of the anthocyanin metabolites, protocatechuic acid and 4-hydroxybenzoic acid. Oxidative Med. Cell. Longev. 2017, 2017, 6297080. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A.; Boirie, Y. Associations between food and beverage groups and major diet-related chronic diseases: An exhaustive review of pooled/meta-analyses and systematic reviews. Nutr. Rev. 2014, 72, 741–762. [Google Scholar] [CrossRef]
- Faulds, C.B.; Sancho, A.I.; Bartolomé, B. Mono- and dimeric ferulic acid release from brewer’s spent grain by fungal feruloyl esterases. Appl. Microbiol. Biotechnol. 2002, 60, 489–493. [Google Scholar] [PubMed]
- Bartolomé, B.; Gómez-Cordovés, C.; Sancho, A.I.; Díez, N.; Ferreira, P.; Soliveri, J.; Copa-Patiño, J.L. Growth and release of hydroxycinnamic acids from brewer’s spent grain by Streptomyces avermitilis CECT 3339. Enzym. Microb. Technol. 2003, 32, 140–144. [Google Scholar] [CrossRef]
- Birsan, R.I.; Wilde, P.; Waldron, K.W.; Rai, D.K. Recovery of Polyphenols from Brewer’s Spent Grains. Antioxidants 2019, 8, 380. [Google Scholar] [CrossRef]
- Guido, L.F.; Moreira, M.M. Techniques for Extraction of Brewer’s Spent Grain Polyphenols: A Review. Food Bioprocess Technol. 2017, 10, 1192–1209. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Phenolics in cereals, fruits and vegetables: Occurrence, extraction and analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Cirillo, E.; Natella, F.; Mencarelli, D.; Comisso, A.; Scaccini, C. Detection of bound phenolic acids: Prevention by ascorbic acid and ethylenediaminetetraacetic acid of degradation of phenolic acids during alkaline hydrolysis. Food Chem. 2002, 79, 119–124. [Google Scholar] [CrossRef]
- Jay, A.J.; Parker, M.L.; Faulks, R.; Husband, F.; Wilde, P.; Smith, A.C.; Faulds, C.B.; Waldron, K.W. A systematic micro-dissection of brewers’ spent grain. J. Cereal Sci. 2008, 47, 357–364. [Google Scholar] [CrossRef]
- Soccol, C.R.; Faraco, V.; Karp, S.G.; Vandenberghe, L.P.S.; Thomaz-Soccol, V.; Woiciechowski, A.L.; Pandey, A. Lignocellulosic bioethanol: Current status and future perspectives. In Biofuels: Alternative Feedstocks and Conversion Processes for the Production of Liquid and Gaseous Biofuels; Elsevier: Amsterdam, The Netherlands, 2019; pp. 331–354. [Google Scholar]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef] [PubMed]
- Zuorro, A.; Iannone, A.; Lavecchia, R. Water–organic solvent extraction of phenolic antioxidants from brewers’ spent grain. Processes 2019, 7, 126. [Google Scholar] [CrossRef]
- Socaci, S.A.; Fărcaş, A.C.; Diaconeasa, Z.M.; Vodnar, D.C.; Rusu, B.; Tofană, M. Influence of the extraction solvent on phenolic content, antioxidant, antimicrobial and antimutagenic activities of brewers’ spent grain. J. Cereal Sci. 2018, 80, 180–187. [Google Scholar] [CrossRef]
- Andres, A.I.; Petron, M.J.; Lopez, A.M.; Timon, M.L. Optimization of Extraction Conditions to Improve Phenolic Content and In Vitro Antioxidant Activity in Craft Brewers’ Spent Grain Using Response Surface Methodology (RSM). Foods 2020, 9, 1398. [Google Scholar] [CrossRef] [PubMed]
- Bonifácio-Lopes, T.; Boas, A.A.V.; Coscueta, E.R.; Costa, E.M.; Silva, S.; Campos, D.; Teixeira, J.A.; Pintado, M. Bioactive extracts from brewer’s spent grain. Food Funct. 2020, 11, 8963–8977. [Google Scholar] [CrossRef]
- Meneses, N.G.T.; Martins, S.; Teixeira, J.A.; Mussatto, S.I. Influence of extraction solvents on the recovery of antioxidant phenolic compounds from brewer’s spent grains. Sep. Purif. Technol. 2013, 108, 152–158. [Google Scholar] [CrossRef]
- Moreira, M.M.; Morais, S.; Barros, A.A.; Delerue-Matos, C.; Guido, L.F. A novel application of microwave-assisted extraction of polyphenols from brewer’s spent grain with HPLC-DAD-MS analysis. Anal. Bioanal. Chem. 2012, 403, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Chen, J.; Zhu, S.; Zhang, C.; Chen, H.; Liu, Y. Inhibition of wheat bran and it’s active compoments on alpha-glucosidase in vitro. Pharm. Mag. 2013, 9, 309–314. [Google Scholar] [CrossRef]
- Reichardt, C.; Welton, T. Empirical Parameters of Solvent Polarity. In Solvents and Solvent Effects in Organic Chemistry; John Wiley & Sons: Hoboken, NJ, USA, 2002; pp. 389–469. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Faraone, I.; Rai, D.K.; Russo, D.; Chiummiento, L.; Fernandez, E.; Choudhary, A.; Milella, L. Antioxidant, Antidiabetic, and Anticholinesterase Activities and Phytochemical Profile of Azorella glabra Wedd. Plants 2019, 8, 265. [Google Scholar] [CrossRef]
- Granato, D.; de Araújo Calado, V.M.; Jarvis, B. Observations on the use of statistical methods in food science and technology. Food Res. Int. 2014, 55, 137–149. [Google Scholar] [CrossRef]
- Watson, D.W. Tips for Troubleshooting Liquid-Liquid Extractions. Available online: https://www.chromatographyonline.com/view/tips-troubleshooting-liquid-liquid-extractions (accessed on 23 August 2020).
- Khoddami, A.; Wilkes, M.A.; Roberts, T.H. Techniques for analysis of plant phenolic compounds. Molecules 2013, 18, 2328–2375. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Dragone, G.; Roberto, I.C. Ferulic and p-coumaric acids extraction by alkaline hydrolysis of brewer’s spent grain. Ind. Crop. Prod. 2007, 25, 231–237. [Google Scholar] [CrossRef]
- Modenbach, A.A.; Nokes, S.E. Effects of sodium hydroxide pretreatment on structural components of biomass. Trans. ASABE 2014, 57, 1187–1198. [Google Scholar]
- Connolly, A.; Piggott, C.O.; FitzGerald, R.J. Characterisation of protein-rich isolates and antioxidative phenolic extracts from pale and black brewers’ spent grain. Int. J. Food Sci. Technol. 2013, 48, 1670–1681. [Google Scholar] [CrossRef]
- Macheiner, D.; Adamitsch, B.F.; Karner, F.; Hampel, W.A. Pretreatment and hydrolysis of brewer’s spent grains. Eng. Life Sci. 2003, 3, 401–405. [Google Scholar] [CrossRef]
- Forssell, P.; Kontkanen, H.; Schols, H.A.; Hinz, S.; Eijsink, V.G.H.; Treimo, J.; Robertson, J.A.; Waldron, K.W.; Faulds, C.B.; Buchert, J. Hydrolysis of brewers’ spent grain by carbohydrate degrading enzymes. J. Inst. Brew. 2008, 114, 306–314. [Google Scholar] [CrossRef]
- Xiros, C.; Topakas, E.; Katapodis, P.; Christakopoulos, P. Evaluation of Fusarium oxysporum as an enzyme factory for the hydrolysis of brewer’s spent grain with improved biodegradability for ethanol production. Ind. Crop. Prod. 2008, 28, 213–224. [Google Scholar] [CrossRef]
- Dehnavi, G.Z.; Laucerica, J.L.; Rodriguez, D.; Beaton, M.; Taherzadeh, M.J.; Martin, C. Fractionation of the main components of barley spent grains from a microbrewery. Cellul. Chem. Technol. 2011, 45, 339. [Google Scholar]
- Wahlström, R.; Rommi, K.; Willberg-Keyriläinen, P.; Ercili-Cura, D.; Holopainen-Mantila, U.; Hiltunen, J.; Mäkinen, O.; Nygren, H.; Mikkelson, A.; Kuutti, L. High yield protein extraction from Brewer’s spent grain with novel carboxylate salt-urea aqueous deep eutectic solvents. ChemistrySelect 2017, 2, 9355–9363. [Google Scholar] [CrossRef]
- Stefanello, F.S.; Dos Santos, C.O.; Bochi, V.C.; Fruet, A.P.B.; Soquetta, M.B.; Dorr, A.C.; Nornberg, J.L. Analysis of polyphenols in brewer’s spent grain and its comparison with corn silage and cereal brans commonly used for animal nutrition. Food Chem. 2018, 239, 385–401. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- de Simón, B.F.; Pérez-Ilzarbe, J.; Hernández, T.; Gómez-Cordovés, C.; Estrella, I. HPLC study of the efficiency of extraction of phenolic compounds. Chromatographia 1990, 30, 35–37. [Google Scholar] [CrossRef]
- Krygier, K.; Sosulski, F.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids. 1. Extraction and purification procedure. J. Agric. Food Chem. 1982, 30, 330–334. [Google Scholar] [CrossRef]
- Liu, J.; Wang, C.; Wang, Z.; Zhang, C.; Lu, S.; Liu, J. The antioxidant and free-radical scavenging activities of extract and fractions from corn silk (Zea mays L.) and related flavone glycosides. Food Chem. 2011, 126, 261–269. [Google Scholar] [CrossRef]
- McCarthy, A.L.; O’Callaghan, Y.C.; Neugart, S.; Piggott, C.O.; Connolly, A.; Jansen, M.A.K.; Krumbein, A.; Schreiner, M.; FitzGerald, R.J.; O’Brien, N.M. The hydroxycinnamic acid content of barley and brewers’ spent grain (BSG) and the potential to incorporate phenolic extracts of BSG as antioxidants into fruit beverages. Food Chem. 2013, 141, 2567–2574. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Fernandes, M.; Dragone, G.; Mancilha, I.M.; Roberto, I.C. Brewer’s spent grain as raw material for lactic acid production by Lactobacillus delbrueckii. Biotechnol. Lett. 2007, 29, 1973–1976. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.M.; Morais, S.; Carvalho, D.O.; Barros, A.A.; Delerue-Matos, C.; Guido, L.F. Brewer’s spent grain from different types of malt: Evaluation of the antioxidant activity and identification of the major phenolic compounds. Food Res. Int. 2013, 54, 382–388. [Google Scholar] [CrossRef]
- Sancho, A.I.; Bartolomé, B.; Gómez-Cordovés, C.; Williamson, G.; Faulds, C.B. Release of ferulic acid from cereal residues by barley enzymatic extracts. J. Cereal Sci. 2001, 34, 173–179. [Google Scholar] [CrossRef]
- Bartolomé, B.; Gómez-Cordovés, C. Barley spent grain: Release of hydroxycinnamic acids (ferulic and p-coumaric acids) by commercial enzyme preparations. J. Sci. Food Agric. 1999, 79, 435–439. [Google Scholar] [CrossRef]
- McCarthy, A.; O’Callaghan, Y.; Piggott, C.; FitzGerald, R.; O’Brien, N. Brewers’ spent grain; bioactivity of phenolic component, its role in animal nutrition and potential for incorporation in functional foods: A review. Proc. Nutr. Soc. 2013, 72, 117–125. [Google Scholar] [PubMed]
- Bartolomé, B.; Faulds, C.B.; Williamson, G. Enzymic release of ferulic acid from barley spent grain. J. Cereal Sci. 1997, 25, 285–288. [Google Scholar] [CrossRef]
- Martín-García, B.; Pasini, F.; Verardo, V.; Díaz-de-Cerio, E.; Tylewicz, U.; Gómez-Caravaca, A.M.; Caboni, M.F. Optimization of sonotrode ultrasonic-assisted extraction of proanthocyanidins from brewers’ spent grains. Antioxidants 2019, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Ikram, S.; Huang, L.; Zhang, H.; Wang, J.; Yin, M. Composition and nutrient value proposition of brewers spent grain. J. Food Sci. 2017, 82, 2232–2242. [Google Scholar] [CrossRef]
- Maillard, M.-N.; Berset, C. Evolution of antioxidant activity during kilning: Role of insoluble bound phenolic acids of barley and malt. J. Agric. Food Chem. 1995, 43, 1789–1793. [Google Scholar] [CrossRef]
- Adelakun, O.E.; Kudanga, T.; Parker, A.; Green, I.R.; le Roes-Hill, M.; Burton, S.G. Laccase-catalyzed dimerization of ferulic acid amplifies antioxidant activity. J. Mol. Catal. B Enzym. 2012, 74, 29–35. [Google Scholar] [CrossRef]
- Garcia-Conesa, M.T.; Plumb, G.W.; Kroon, P.A.; Wallace, G.; Williamson, G. Antioxidant properties of ferulic acid dimers. Redox Rep. 1997, 3, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Jabir, N.R.; Khan, F.R.; Tabrez, S. Cholinesterase targeting by polyphenols: A therapeutic approach for the treatment of Alzheimer’s disease. CNS Neurosci. Ther. 2018, 24, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.B.; Bunzel, M.; Schaäfer, J.; Knudsen, K.E.B.; Sørensen, J.F.; Yu, S.; Lærke, H.N. Ferulic acid dehydrodimer and dehydrotrimer profiles of distiller’s dried grains with solubles from different cereal species. J. Agric. Food Chem. 2015, 63, 2006–2012. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; He, Y.; Lu, F. The structure-antioxidant activity relationship of dehydrodiferulates. Food Chem. 2018, 269, 480–485. [Google Scholar] [CrossRef]
- Nenadis, N.; Zhang, H.-Y.; Tsimidou, M.Z. Structure—Antioxidant activity relationship of ferulic acid derivatives: Effect of carbon side chain characteristic groups. J. Agric. Food Chem. 2003, 51, 1874–1879. [Google Scholar] [CrossRef] [PubMed]
- Terpinc, P.; Polak, T.; Šegatin, N.; Hanzlowsky, A.; Ulrih, N.P.; Abramovič, H. Antioxidant properties of 4-vinyl derivatives of hydroxycinnamic acids. Food Chem. 2011, 128, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Ouattara, N.; Meda, R.N.-T.; Hilou, A.; Guenné, S.; Konaté, K.; Coulibaly, A.Y.; Kiendrébeogo, M.; Millogo, J.F.; Nacoulma, O.G. Anti-acetylcholinesterase and antioxidant activities and HPLC-MS analysis of polyphenol from extracts of Nelsonia canescens (Lam.) Spreng. Asian Pac. J. Trop. Dis. 2013, 3, 382–388. [Google Scholar] [CrossRef]
- Gonçalves, S.; Moreira, E.; Grosso, C.; Andrade, P.B.; Valentão, P.; Romano, A. Phenolic profile, antioxidant activity and enzyme inhibitory activities of extracts from aromatic plants used in Mediterranean diet. J. Food Sci. Technol. 2017, 54, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Vladimir-Knežević, S.; Blažeković, B.; Kindl, M.; Vladić, J.; Lower-Nedza, A.D.; Brantner, A.H. Acetylcholinesterase inhibitory, antioxidant and phytochemical properties of selected medicinal plants of the Lamiaceae family. Molecules 2014, 19, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D.; Borowiec, K. Phenolic acids from malt are efficient acetylcholinesterase and butyrylcholinesterase inhibitors. J. Inst. Brew. 2012, 118, 40–48. [Google Scholar] [CrossRef]
- Szwajgier, D. Anticholinesterase activity of phenolic acids and their derivatives. Z Nat. C 2013, 68, 125–132. [Google Scholar] [CrossRef]
- Shahwar, D.; Rehman, S.U.; Raza, M.A. Acetyl cholinesterase inhibition potential and antioxidant activities of ferulic acid isolated from Impatiens bicolor Linn. J. Med. Plant Res. 2010, 4, 260–266. [Google Scholar]
- Orhan, I.; Kartal, M.; Tosun, F.; Şener, B. Screening of various phenolic acids and flavonoid derivatives for their anticholinesterase potential. Z Nat. C 2007, 62, 829–832. [Google Scholar] [CrossRef]
- Kikuzaki, H.; Hisamoto, M.; Hirose, K.; Akiyama, K.; Taniguchi, H. Antioxidant properties of ferulic acid and its related compounds. J. Agric. Food Chem. 2002, 50, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Salazar-López, N.J.; González-Aguilar, G.A.; Loarca-Piña, G.; Cinco-Moroyoqui, F.J.; Rouzaud-Sández, O.; Domínguez-Avila, J.A.; Robles-Sánchez, M. Contribution and interactions of hydroxycinnamic acids found in bran and wholegrain sorghum (Sorghum bicolor L. Moench): Effects on the antioxidant capacity and inhibition of human erythrocyte hemolysis. Oxidative Med. Cell. Longev. 2017, 2017, 8219023. [Google Scholar] [CrossRef] [PubMed]
- Lilienfeld, S. Galantamine—a novel cholinergic drug with a unique dual mode of action for the treatment of patients with Alzheimer’s disease. CNS Drug Rev. 2002, 8, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.M.; Ferreres, F.; Oliveira, J.M.A.; Gaspar, L.; Faria, J.; Valentão, P.; Sottomayor, M.; Andrade, P.B. Pharmacological effects of Catharanthus roseus root alkaloids in acetylcholinesterase inhibition and cholinergic neurotransmission. Phytomedicine 2010, 17, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.-H.; Yang, Z.-D.; Xue, W.-W.; Sheng, J.; Shi, Y.; Yao, X.-J. Isolation, characterization and acetylcholinesterase inhibitory activity of alkaloids from roots of Stemona sessilifolia. Fitoterapia 2013, 89, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-d.; Zhang, X.; Du, J.; Ma, Z.-j.; Guo, F.; Li, S.; Yao, X.-j. An aporphine alkaloid from Nelumbo nucifera as an acetylcholinesterase inhibitor and the primary investigation for structure–activity correlations. Nat. Prod. Res. 2012, 26, 387–392. [Google Scholar] [CrossRef]
- Santos, T.C.d.; Gomes, T.M.; Pinto, B.A.S.; Camara, A.L.; Paes, A.M.d.A. Naturally occurring acetylcholinesterase inhibitors and their potential use for Alzheimer’s disease therapy. Front. Pharmacol. 2018, 9, 1192. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D. Anticholinesterase activities of selected polyphenols–a short report. Pol. J. Food Nutr. Sci. 2014, 64, 59–64. [Google Scholar] [CrossRef]
- Stanley, T.H.; Van Buiten, C.B.; Baker, S.A.; Elias, R.J.; Anantheswaran, R.C.; Lambert, J.D. Impact of roasting on the flavan-3-ol composition, sensory-related chemistry, and in vitro pancreatic lipase inhibitory activity of cocoa beans. Food Chem. 2018, 255, 414–420. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, R.D. Protection of cholinesterase by ethanol against inhibition by organophosphates in vitro. J. Biol. Chem. 1956, 219, 927–931. [Google Scholar] [CrossRef]
- Pohanka, M.; Fusek, J.; Adam, V.; Kizek, R. Carbofuran assay using gelatin based biosensor with acetylcholinesterase as a recognition element. Int. J. Electrochem. Sci. 2013, 8, 71–79. [Google Scholar]
- Musial, A.; Bajda, M.; Malawska, B. Recent developments in cholinesterases inhibitors for Alzheimer’s disease treatment. Curr. Med. Chem. 2007, 14, 2654–2679. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Samples | Extraction Yield (mg/g BSG) | TPC (mg GAE/g BSG) | SQP (mg/g BSG) | |||
|---|---|---|---|---|---|---|
| FP | BP | FP | BP | FP | BP | |
| Hex F. | 19.4 ± 16.8 a | 12.2 ± 7.1 b | 0.09 ± 0.09 a | 0.02 ± 0.0 b | n.d. b | <0.01 c |
| DE F. | 25.8 ± 14.2 a | 8.3 ± 1.2 b | 0.23 ± 0.09 a | 0.67 ± 0.02 b | <0.01 a | 0.8 ± 0.0 a b |
| EtOAc F. | 6.6 ± 4.0 a | 34.6 ± 6.9 a | 0.12 ± 0.07 a | 3.5 ± 0.5 a | <0.01 a | 2.9 ± 0.3 a |
| BuOH F. | 6.2 ± 4.4 a | 23.8 ± 8.3 a b | 0.09 ± 0.004 a | 1.0 ± 0.4 a b | <0.01 a b | 0.07 ± 0.03 b c |
| WR F. | 22.2 ± 7.8 a | 345 ± 162.5 a | 0.11 ± 0.02 a | 6.2 ± 2.8 a | n.d. b | 0.03 ± 0.04 c |
| Total | 80.2 ± 3.4 | 424.2 ± 179.9 | 0.64 ± 0.07 | 11.3 ± 3.6 | 0.013 ± 0.02 | 3.80 ± 0.2 |
| Crude (Control) | 94.9 ± 9.2 | n.t. | 1.7 ± 0.2 | n.t. | <0.02 | n.t. |
| Samples (µg/mg) | FA | p-CA | Cat | CafA | 4-HBA | ProA | DeCa-DiFA | DiFA | TriFA | Total | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FP | BP | FP | BP | FP | BP | FP | BP | FP | BP | FP | BP | FP | BP | FP | BP | FP | BP | FP | BP | |
| Hex F. | n.d. | 0.04 ± 0.07 b | n.d. | 0.05 ± 0.06 c | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.08 ± 0.08 b | n.d. | n.d. | n.d. | n.d. | n.d. | 0.15 ± 0.2 c |
| DE F. | 0.053 ± 0.03 a | 41.5 ± 15.3 a | 0.03 ± 0.02 a | 25.9 ± 6.3 a | 0.33 ± 0.10 a | n.d. | 0.02 ± 0.01 a | 0.50 ± 0.2 a b | 0.05 ± 0.05 a | 0.30 ± 0.1 a b | 0.06 ± 0.03 ab | 0.04 ± 0.00 a b | n.d. | 30.62 ± 0.8 a | n.d. | 0.04±0.01 b | n.d. | n.d. | 0.5 ± 0.3 a | 99.0 ± 21.2 a |
| EtOAc F. | 0.054 ± 0.03 a | 40.6 ± 7.7 a | n.d. | 16.1 ± 1.9 a b | 0.88 ± 0.67 a | n.d. | 0.03 ± 0.02 a | 1.04 ± 0.3 a | 0.04 ± 0.03 a | 0.33 ± 0.05 a | 0.21 ± 0.04 a | 0.24 ± 0.06 a | n.d. | 0.95 ± 0.6 a b | n.d. | 12.91±2.4 a | n.d. | 12.76 ± 2.3 a | 1.0 ± 0.9 a | 84.9 ± 14.6 a b |
| BuOH F. | n.d. | 1.2 ± 1.0 a b | n.d. | 0.44 ± 0.4 b | 0.06 ± 0.06 a | n.d. | n.d. | 0.03 ± 0.02 b c | n.d. | 0.02 ± 0.01 b | 0.03 ± 0.03 a b | 0.03 ± 0.01 b | n.d. | 0.08 ± 0.04 b | n.d. | 0.92±0.6 a b | n.d. | 0.70 ± 0.7 a b | 0.07 ± 0.1 a | 3.3 ± 2.8 b c |
| WR F. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.09 * b | n.d. | 0.11 * b | n.d. | 0.1 ± 0.1 c |
| Crude Ctrl | n.d. | 5.33 * | n.d. | 2.28 * | 0.15 * | n.d. | n.d. | 0.08 * | 0.01 * | 0.03 * | 0.02* | 0.02* | n.d. | 0.58 * | - | 1.09 * | n.d. | 0.78 * | 0.19 * | 10.2 * |
| Samples 1 mg/mL | TPC μgGAE/mg Extract | SQP μg/mg Extract | AChE %Inhibition | BChE %Inhibition | |
|---|---|---|---|---|---|
| FP | Hex F. | 4.1 ± 0.6 e | n.d. | 11.7 ± 1.3 b | 17.5 ± 1.8 c d |
| DE F. | 9.8 ± 1.9 e | 0.5 ± 0.3 b | 10.7 ± 3.6 b | 16.4 ± 3.1 c d | |
| EtOAc F. | 20.3 ± 3.4 d e | 1.0 ± 1 b | 8.7 ± 0.6 b | 15.7 ± 2.9 c d | |
| BuOH F. | 11.4 ± 2.5 e | 0.07 ± 0.1 b | 34.9 ± 6.4 a | 40.5 ± 11.2b | |
| WR F. | 5.1 ± 1.1 e | - | 12.8 ± 0.7 b | - | |
| Crude Ctrl | 17.3 ± 0.7 d e | 0.19 * b | 20.8 ± 2.2 b | 17.2 ± 1.2 c d | |
| BP | Hex F. | 1.8 ± 0.4 e | 0.15 ± 0.2 b | 13.8 ± 3.5 b | 25.1 ± 1.5 c |
| DE F. | 82.9 ± 13.2 b | 99.0 ± 21.2 a | 37.9 ± 10.4 a | 53.6 ± 7.7 a | |
| EtOAc F. | 102.3 ± 14.1 a | 84.9 ± 14.6 a | 10.3 ± 2.9 b | 25.3 ± 3.3 c | |
| BuOH F. | 40.7 ± 1.6 c | 3.3 ± 2.8 b | 14.3 ± 2.9 b | 16.9 ± 3.1 c d | |
| WR F. | 18.0 ± 0.9 d e | 0.1 ± 0.1 b | 11.6 ± 1.3 b | 9.4 ± 3.8 d e | |
| Crude Ctrl | 31.7 ± 0.8 c d | 10.2 * b | 10.2 ± 1.4 b | 11.4 ± 0.4 d e | |
| Standards | AChE % Inhibition | BChE % Inhibition | ||
|---|---|---|---|---|
| 0.1 mg/mL | 1 mg/mL | 0.1 mg/mL | 1 mg/mL | |
| Ferulic A. | 1.0 ± 0.9 b | 15.4 ± 0.1 a b | 14.6 ± 1 a b | 27.2 ± 0.9 a b |
| p-Coumaric A. | 5.2 ± 0.4 a | 14.4 ± 0.5 b c | 6.4 ± 0.6 b | 22.1 ± 1.3 b c |
| Catechin | 3.8 ± 1.1 ab | 14.9 ± 0.2 a b | 12.2 ± 0.6 a b | 31.6 ± 0.4 a b |
| 4-Hydroxybenzoic A. | 1.0 ± 0.2 b | 5.2 ± 0.9 c | n.d. | 11.9 ± 0.6 b c |
| Caffeic A. | 3.3 ± 0.4 a b | 25.5 ± 0.2 a | 15.4 ± 1.3a | 52.3 ± 0.8 a |
| Protocatechuic A. | n.d. | 13.8 ± 0.7 b c | n.d. | 7.6 ± 2.4 c |
| Blends | TPC μgGAE/mg | SQP μg/mg | AChE %Inhibition | BChE %Inhibition |
| FP EtOAc1 | 260.6 ± 11.9 a b | 1000 | n.d. | 16.7 ± 1.5 a |
| BP DE1 | 243.8 ± 1.4 b | 1000 | 11.1 ± 0.6 a | 9.9 ± 0.2 b |
| BP EtOAc3 | 267.4 ± 8.4 a | 1000 | 8.3 ± 0.1 a | 11.2 ± 1.1 a b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birsan, R.I.; Wilde, P.; Waldron, K.W.; Rai, D.K. Anticholinesterase Activities of Different Solvent Extracts of Brewer’s Spent Grain. Foods 2021, 10, 930. https://doi.org/10.3390/foods10050930
Birsan RI, Wilde P, Waldron KW, Rai DK. Anticholinesterase Activities of Different Solvent Extracts of Brewer’s Spent Grain. Foods. 2021; 10(5):930. https://doi.org/10.3390/foods10050930
Chicago/Turabian StyleBirsan, Rares I., Peter Wilde, Keith W. Waldron, and Dilip K. Rai. 2021. "Anticholinesterase Activities of Different Solvent Extracts of Brewer’s Spent Grain" Foods 10, no. 5: 930. https://doi.org/10.3390/foods10050930
APA StyleBirsan, R. I., Wilde, P., Waldron, K. W., & Rai, D. K. (2021). Anticholinesterase Activities of Different Solvent Extracts of Brewer’s Spent Grain. Foods, 10(5), 930. https://doi.org/10.3390/foods10050930