
Molecular Identification and Selection of Probiotic Strains Able to Reduce the Serum TMAO Level in Mice Challenged with Choline

 , , ,
, , ,
Abstract
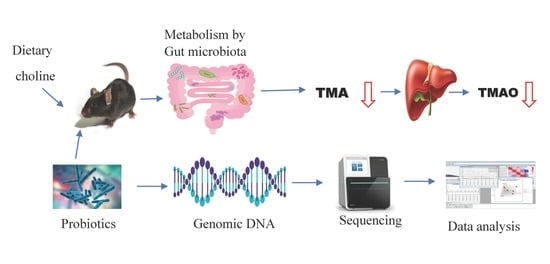
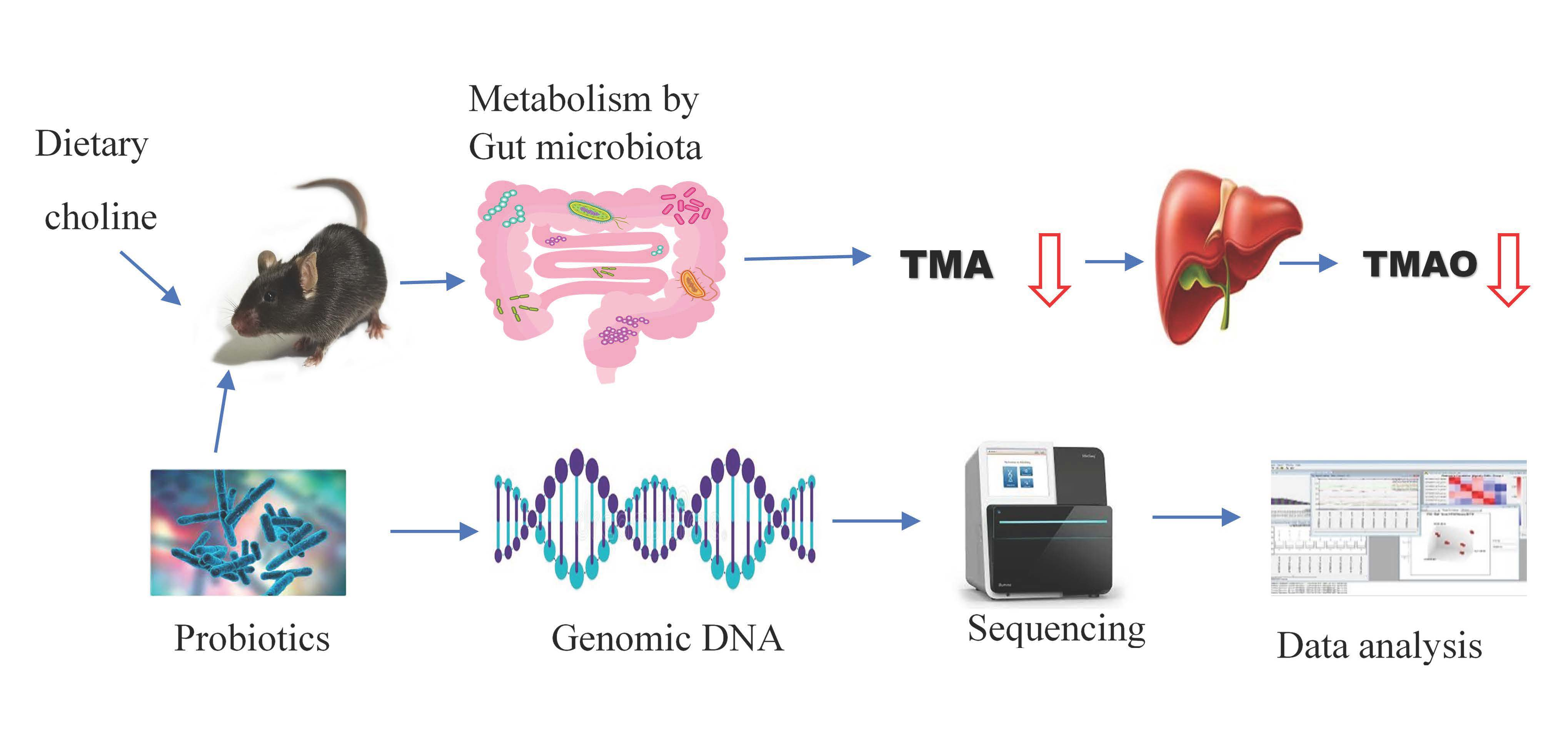
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Screening of Lactic Acid Bacteria (LAB) Strains and Their Antagonistic Effect against TMA-Producing Bacteria
2.2.1. Resistance to Artificial Gastric and Intestinal Fluids
2.2.2. Antimicrobial Activity of LAB Strains
2.2.3. Adhesion to Caco-2 Cells for LAB Strains and TMA-Producing Strains
2.3. Mouse Models
2.4. Probiotics Treatment
2.5. UPLC-MS/MS Analysis
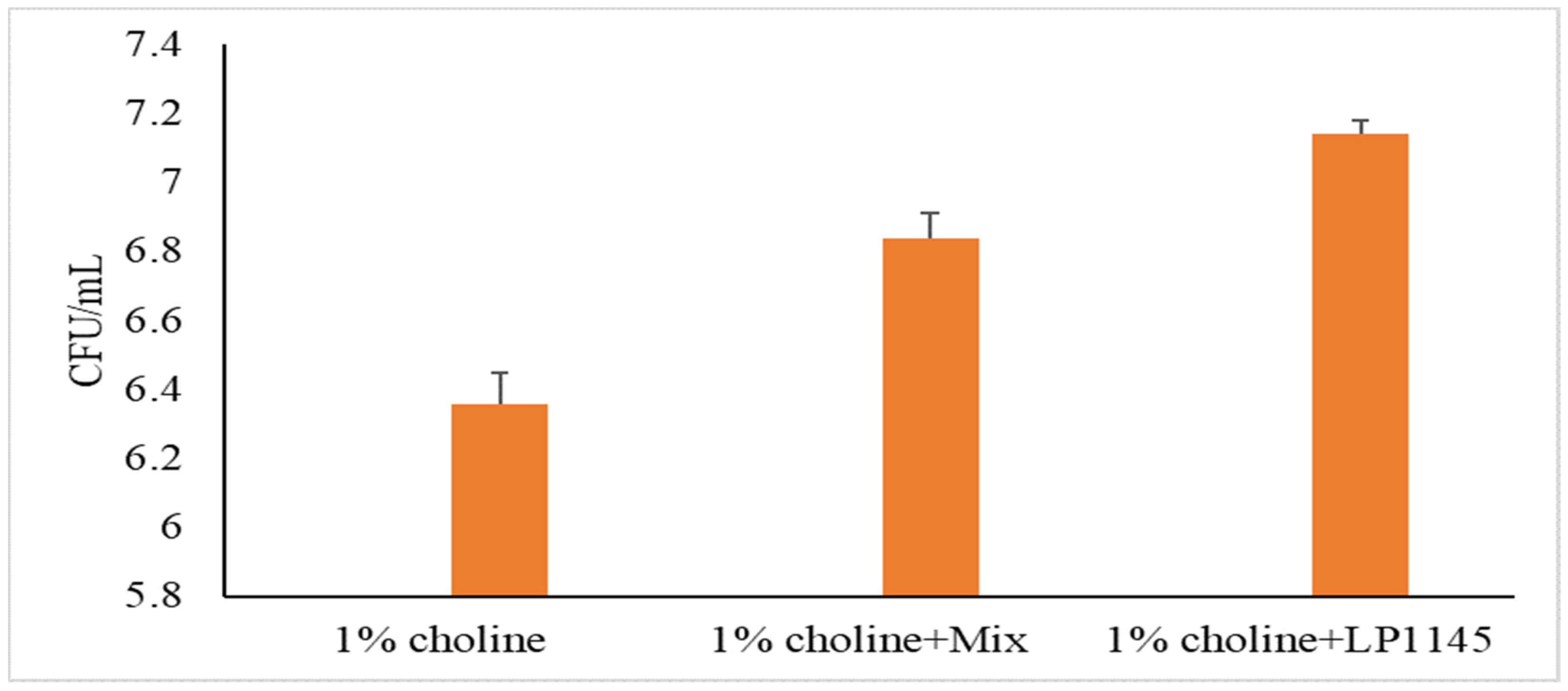
2.6. Counting Lactobacilli Population in Cecum Digesta of Mice
2.7. Quantification of Lactobacilli Populations by qPCR
2.8. Identification of Lactobacillus Amylovorus Strain LAM 1345
2.8.1. Genotypic Characterization Method
DNA Extraction
2.8.2. Leader Peptidase A (lep A) Gene Analysis
2.8.3. Phylogenetic Analysis
2.9. Lpb. plantarum LP1145 Identification
2.9.1. Genomic DNA Extraction, Sequencing, and Annotation
2.9.2. Identification of Genes Related to Antimicrobial Resistance (AMR) and Virulence Factors, Biogenic Amine Producing Genes
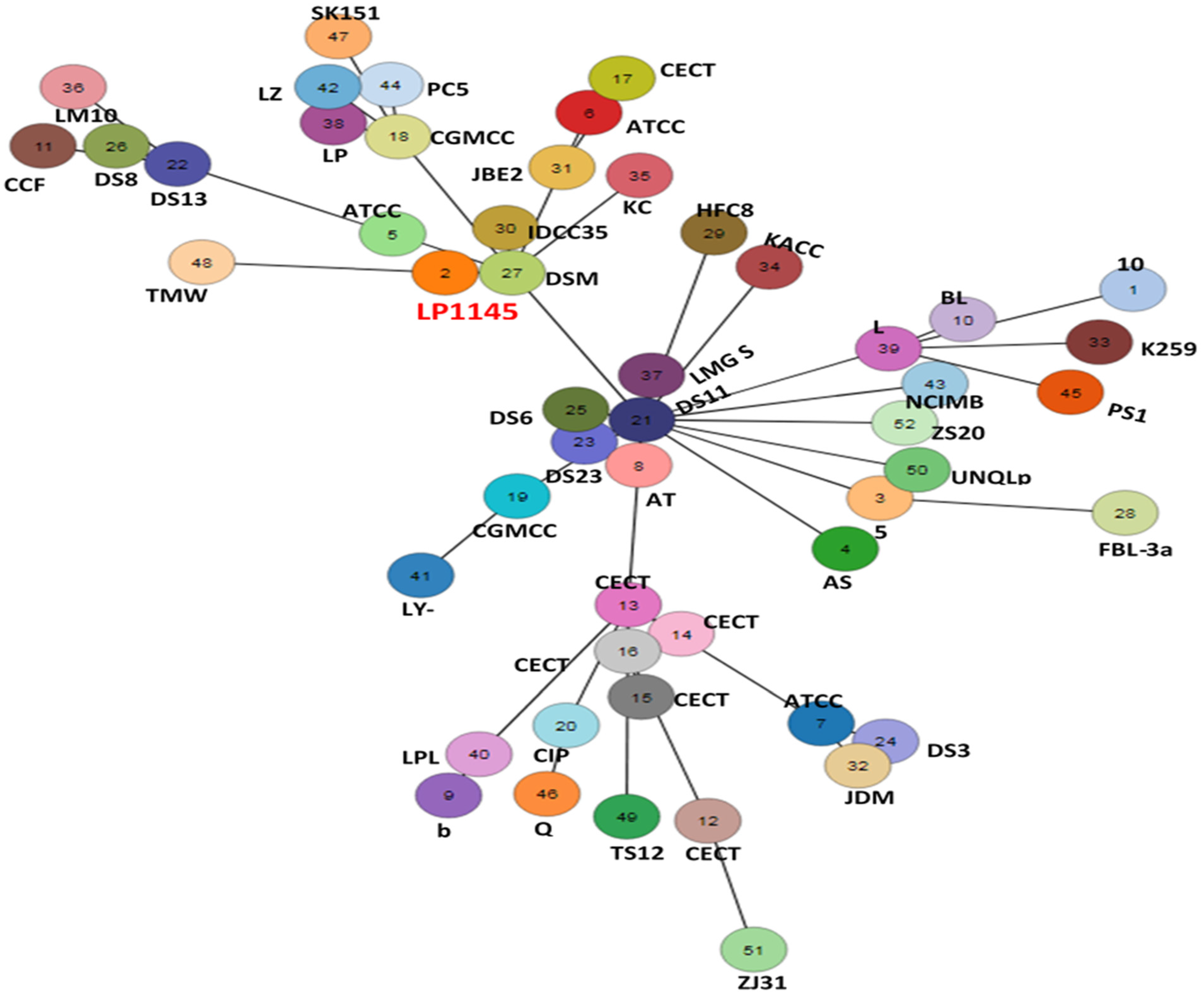
2.9.3. Whole-Genome Multilocus Sequence Typing (WgMLST)
2.10. Lim. fermentum LF1143 Identification
2.11. Statistical Analysis
3. Results
3.1. Acid and Bile Tolerance among Lactobacilli
3.2. Growth Inhibition of TMA Bacteria by LAB Supernatants
3.3. Adherence of TMA Bacteria and Lactobacilli to Caco-2 Cells
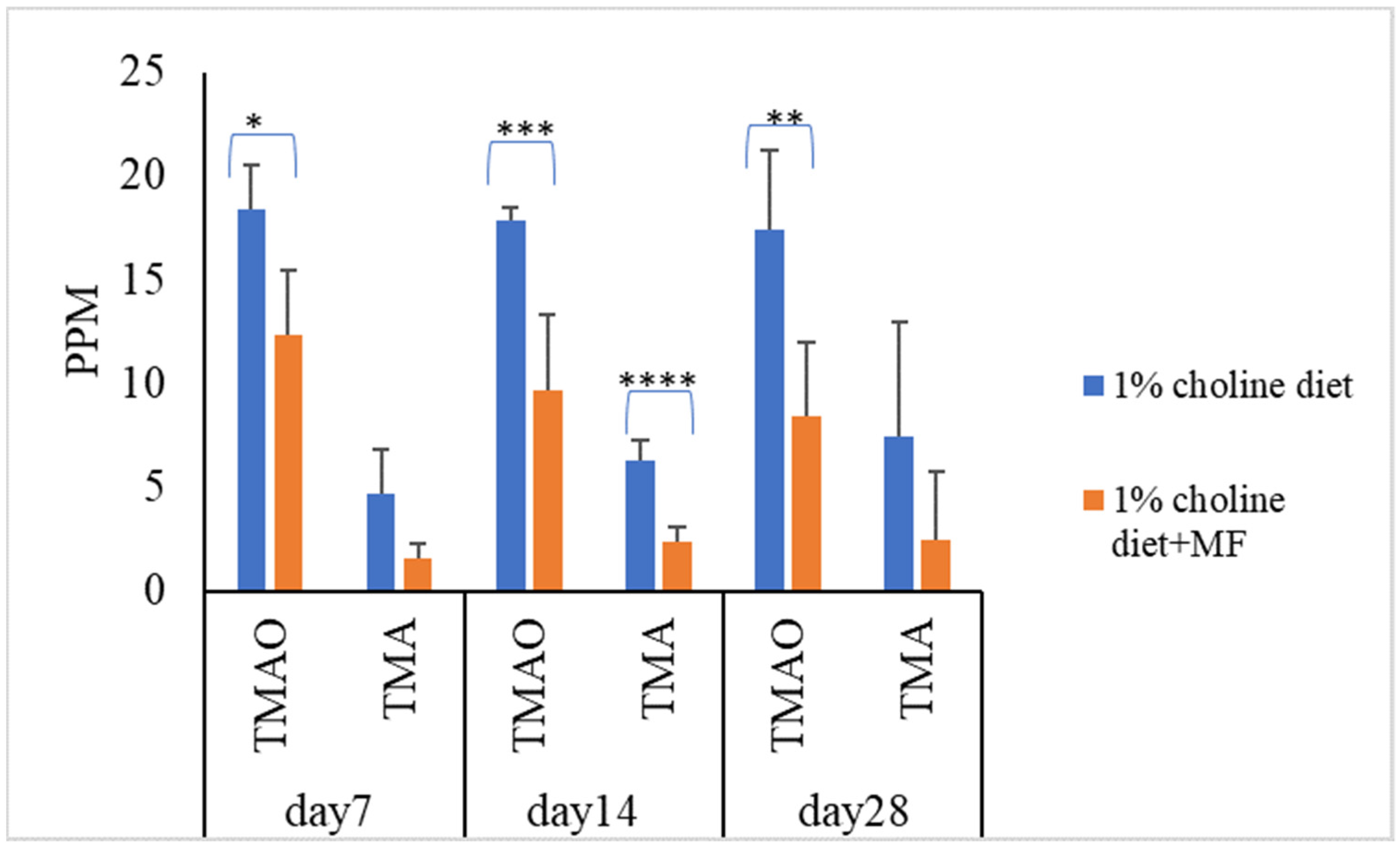
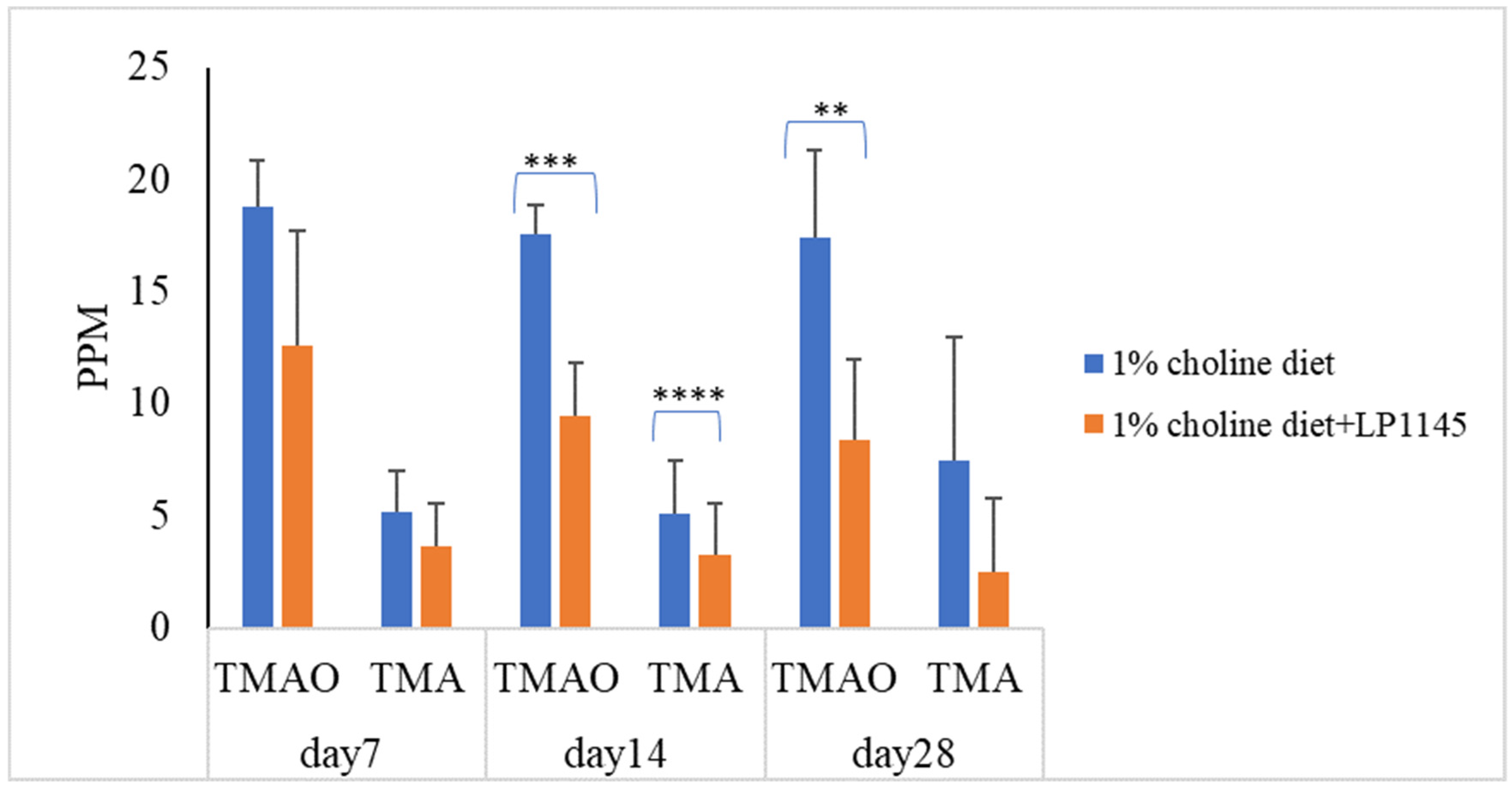
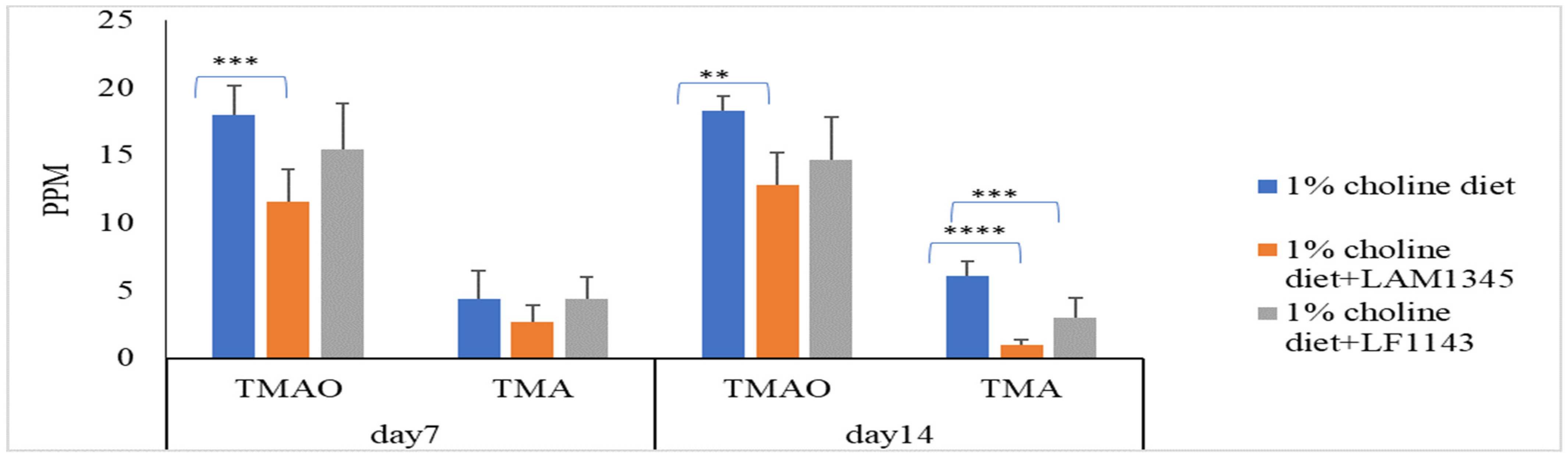
3.4. Probiotic Strains Reducing Serum TMAO Levels in Mice Challenged with Choline
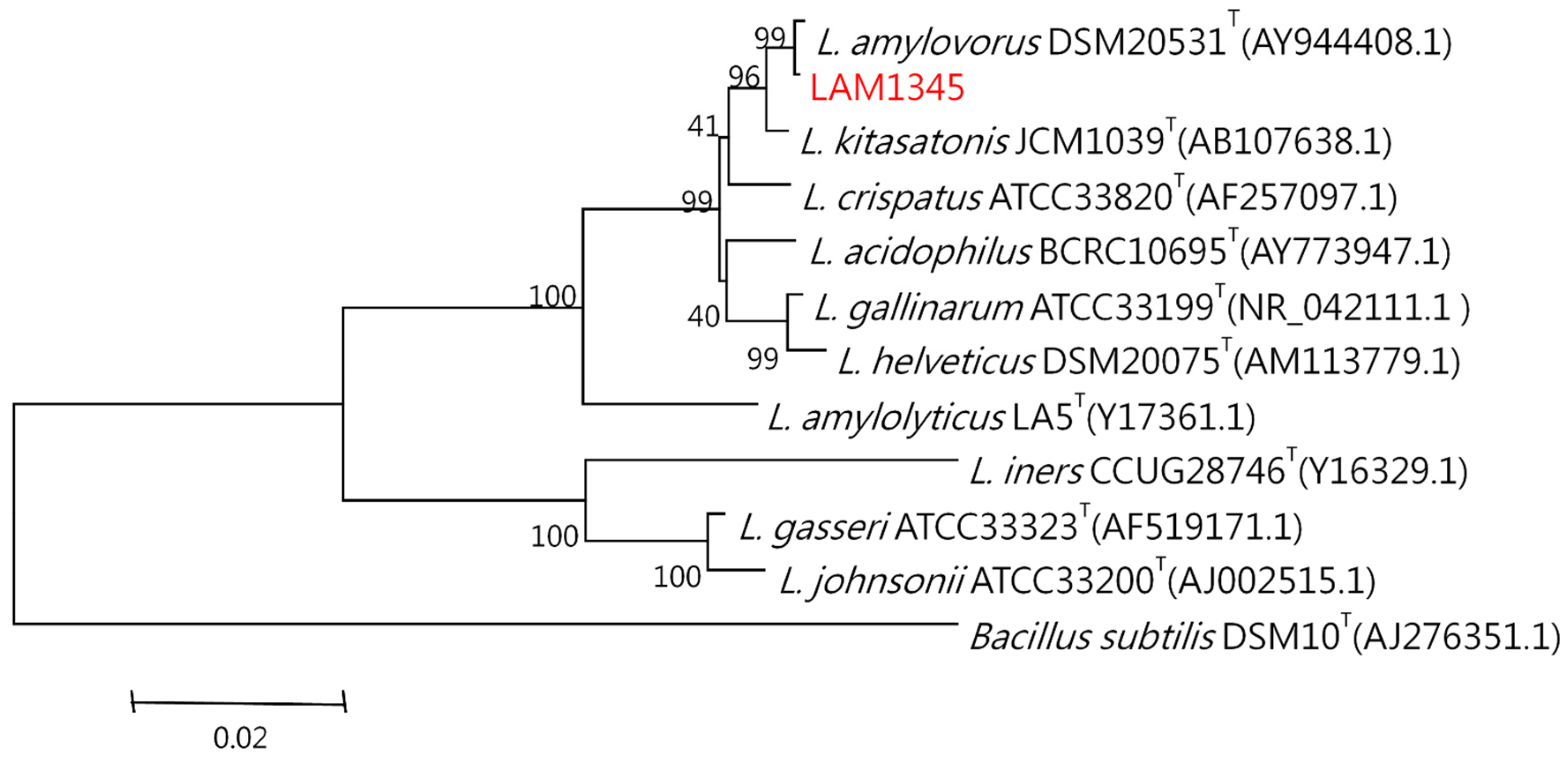
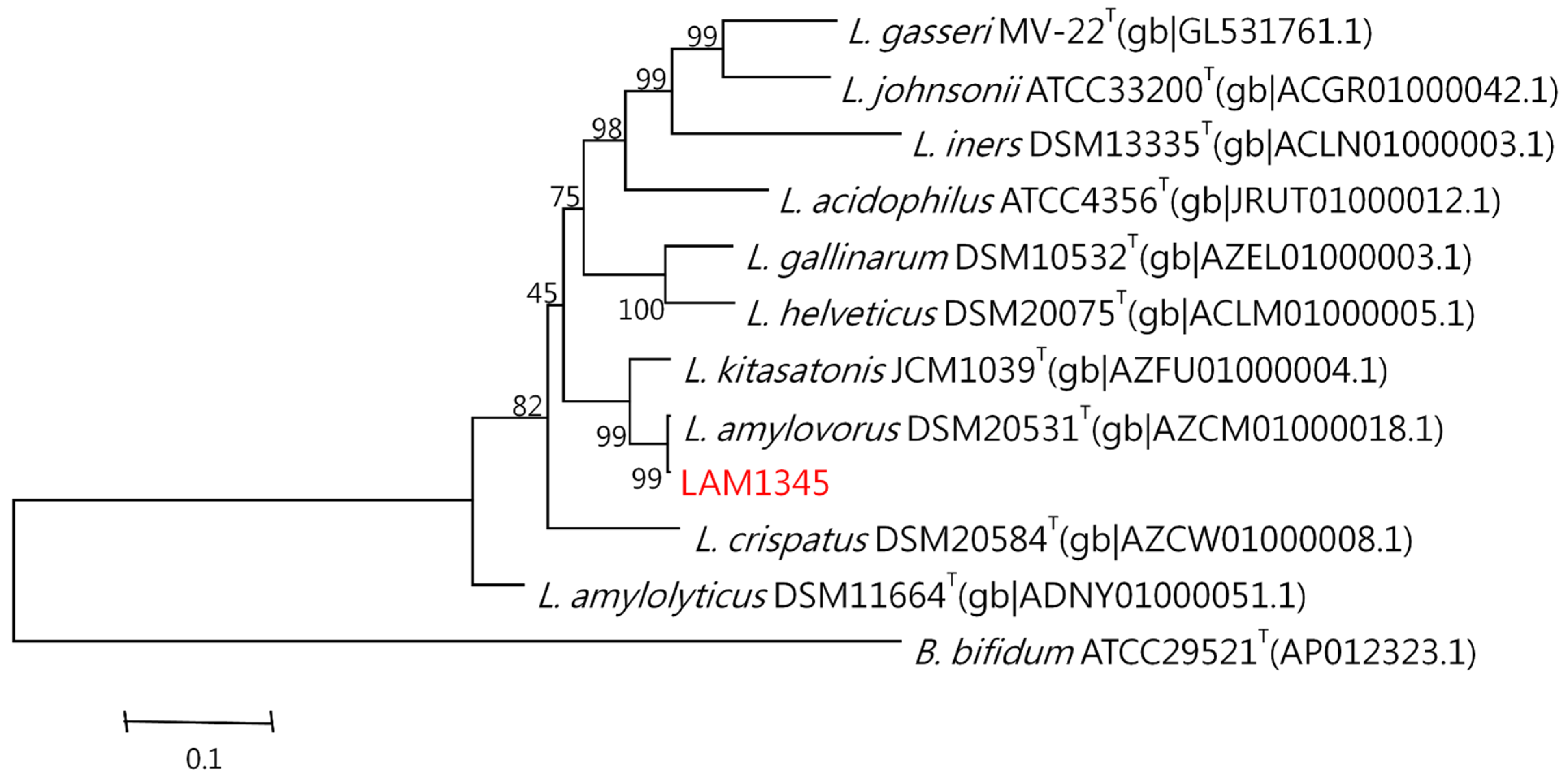
3.5. Lactobacillus Amylovorus Identification
3.5.1. Identification of Bacteria through Bacteriological and Biochemical Tests
3.5.2. Sequence Analysis and Phylogenetic Tree
3.6. Lpb. plantarum LP1145 Identification
3.7. Lim. fermentum Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dahlof, B. Cardiovascular disease risk factors: Epidemiology and risk assessment. Am. J. Cardiol. 2010, 105, 3A–9A. [Google Scholar] [CrossRef]
- Lloyd-Jones, D.M. Cardiovascular risk prediction: Basic concepts, current status, and future directions. Circulation 2010, 121, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, K.; Wang, X.; Pang, Y.; Jiang, C. The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell 2021, 12, 360–373. [Google Scholar] [CrossRef]
- Bennett, B.J.; de Aguiar Vallim, T.Q.; Wang, Z.; Shih, D.M.; Meng, Y.; Gregory, J.; Allayee, H.; Lee, R.; Graham, M.; Crooke, R.; et al. Trimethylamine-N-oxide, a metabolite associated with atherosclerosis, exhibits complex genetic and dietary regulation. Cell Metab. 2013, 17, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Wolino, K.S.; de F Cardozo, L.F.M.; Cardozo, L.; de Oliveira Leal, V.; Mafra, D.; Stockler-Pinto, M.B. Can diet modulate trimethylamine N-oxide (TMAO) production? What do we know so far? Eur. J. Nutr. 2021, 60, 3567–3584. [Google Scholar] [CrossRef]
- Iglesias-Carres, L.; Hughes, M.D.; Steele, C.N.; Ponder, M.A.; Davy, K.P.; Neilson, A.P. Use of dietary phytochemicals for inhibition of trimethylamine N-oxide formation. J. Nutr. Biochem. 2021, 91, 108600. [Google Scholar] [CrossRef]
- Tripolt, N.J.; Leber, B.; Triebl, A.; Kofeler, H.; Stadlbauer, V.; Sourij, H. Effect of Lactobacillus casei Shirota supplementation on trimethylamine-N-oxide levels in patients with metabolic syndrome: An open-label, randomized study. Atherosclerosis 2015, 242, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Boutagy, N.E.; Neilson, A.P.; Osterberg, K.L.; Smithson, A.T.; Englund, T.R.; Davy, B.M.; Hulver, M.W.; Davy, K.P. Probiotic supplementation and trimethylamine-N-oxide production following a high-fat diet. Obesity 2015, 23, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, Z.; Lv, Y.; Tong, L.; Liu, T.; Yi, H.; Zhou, X.; Yu, Z.; Tian, X.; Cui, Q.; et al. Reduction of intestinal trimethylamine by probiotics ameliorated lipid metabolic disorders associated with atherosclerosis. Nutrition 2020, 79–80, 110941. [Google Scholar] [CrossRef]
- Chen, S.; Jiang, P.P.; Yu, D.; Liao, G.C.; Wu, S.L.; Fang, A.P.; Chen, P.Y.; Wang, X.Y.; Luo, Y.; Long, J.A.; et al. Effects of probiotic supplementation on serum trimethylamine-N-oxide level and gut microbiota composition in young males: A double-blinded randomized controlled trial. Eur. J. Nutr. 2021, 60, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Zarate, G.; Chaia, A.P.; Gonzalez, S.; Oliver, G. Viability and beta-galactosidase activity of dairy propionibacteria subjected to digestion by artificial gastric and intestinal fluids. J. Food Prot. 2000, 63, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Rammelsberg, M.; Radler, F. Antibacterial polypeptides of Lactobacillus species. J. Appl. Bacteriol. 1990, 69, 177–184. [Google Scholar] [CrossRef]
- Gopal, P.K.; Prasad, J.; Smart, J.; Gill, H.S. In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli. Int. J. Food Microbiol. 2001, 67, 207–216. [Google Scholar] [CrossRef]
- Tsai, C.C.; Huang, L.F.; Lin, C.C.; Tsen, H.Y. Antagonistic activity against Helicobacter pylori infection in vitro by a strain of Enterococcus faecium TM39. Int. J. Food Microbiol. 2004, 96, 1–12. [Google Scholar] [CrossRef]
- Johnson, D.W. A flow injection electrospray ionization tandem mass spectrometric method for the simultaneous measurement of trimethylamine and trimethylamine N-oxide in urine. J. Mass Spectrom. 2008, 43, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Ramireddy, L.; Tsen, H.-Y.; Chiang, Y.-C.; Hung, C.Y.; Chen, F.-C.; Yen, H.-T. The gene expression and bioinformatic analysis of choline trimethylamine-lyase (CutC) and its activating enzyme (CutD) for gut microbes and comparison with their TMA production levels. Curr. Res. Microb. Sci. 2021, 2, 100043. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Park, Y.K.; Kim, J.F. PAIDB v2.0: Exploration and analysis of pathogenicity and resistance islands. Nucleic Acids Res. 2015, 43, D624–D630. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Lin, J.W.; Chen, C.C. cano-wgMLST_BacCompare: A Bacterial Genome Analysis Platform for Epidemiological Investigation and Comparative Genomic Analysis. Front. Microbiol. 2019, 10, 1687. [Google Scholar] [CrossRef]
- Huang, C.H.; Chen, C.C.; Chiu, S.H.; Liou, J.S.; Lin, Y.C.; Lin, J.S.; Huang, L.; Watanabe, K. Development of a high-resolution single-nucleotide polymorphism strain-typing assay using whole genome–based analyses for the Lactobacillus acidophilus probiotic strain. Microorganisms 2020, 8, 1445. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Sheu, S.J. Use of Tuf Gene Sequences for the Quantititative and Qualitative Assay of Lactic Acid Bacteria and Comparison of Tuf with 16SrRNA Gene for the Phylogenetic Analysis of Bifidobacterium. Ph.D. Thesis, National Chung Hsing University, Taichung, Taiwan, 2009. [Google Scholar]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Dickson, E.M.; Riggio, M.P.; Macpherson, L. A novel species-specific PCR assay for identifying Lactobacillus fermentum. J. Med. Microbiol. 2005, 54, 299–303. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Song, Y.; Kato, N.; Liu, C.; Matsumiya, Y.; Kato, H.; Watanabe, K. Rapid identification of 11 human intestinal Lactobacillus species by multiplex PCR assays using group- and species-specific primers derived from the 16S-23S rRNA intergenic spacer region and its flanking 23S rRNA. FEMS Microbiol. Lett. 2000, 187, 167–173. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for Evaluation of Probiotics in Food; Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food London, ON, Canada, 30 April and 1 May 2002; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Hsu, T.C.; Yi, P.J.; Lee, T.Y.; Liu, J.R. Probiotic characteristics and zearalenone-removal ability of a Bacillus licheniformis strain. PLoS ONE 2018, 13, e0194866. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, M.; Zhao, J.; Xia, Y.; Lai PF, H.; Ai, L. A surface protein from Lactobacillus plantarum increases the adhesion of Lactobacillus strains to human epithelial cells. Front. Microbiol. 2018, 9, 2858. [Google Scholar] [CrossRef] [PubMed]
- Hynonen, U.; Kant, R.; Lahteinen, T.; Pietila, T.E.; Beganovic, J.; Smidt, H.; Uroic, K.; Avall-Jaaskelainen, S.; Palva, A. Functional characterization of probiotic surface layer protein-carrying Lactobacillus amylovorus strains. BMC Microbiol. 2014, 14, 199. [Google Scholar] [CrossRef] [PubMed]
- Hynonen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Warrier, M. Trimethylamine N-Oxide, the Microbiome, and Heart and Kidney Disease. Annu. Rev. Nutr. 2017, 37, 157–181. [Google Scholar] [CrossRef]
- Qiu, L.; Tao, X.; Xiong, H.; Yu, J.; Wei, H. Lactobacillus plantarum ZDY04 exhibits a strain-specific property of lowering TMAO via the modulation of gut microbiota in mice. Food Funct. 2018, 9, 4299–4309. [Google Scholar] [CrossRef]
- Qiu, L.; Yang, D.; Tao, X.; Yu, J.; Xiong, H.; Wei, H. Enterobacter aerogenes ZDY01 Attenuates Choline-Induced Trimethylamine N-Oxide Levels by Remodeling Gut Microbiota in Mice. J. Microbiol. Biotechnol. 2017, 27, 1491–1499. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Chan, J.Y.H.; Lee, C.T.; Tain, Y.L. Hypertension Programmed by Perinatal High-Fat Diet: Effect of Maternal Gut Microbiota-Targeted Therapy. Nutrients 2019, 11, 2908. [Google Scholar] [CrossRef]
- Malik, M.; Suboc, T.M.; Tyagi, S.; Salzman, N.; Wang, J.; Ying, R.; Tanner, M.J.; Kakarla, M.; Baker, J.E.; Widlansky, M.E. Lactobacillus plantarum 299v Supplementation Improves Vascular Endothelial Function and Reduces Inflammatory Biomarkers in Men with Stable Coronary Artery Disease. Circ. Res. 2018, 123, 1091–1102. [Google Scholar] [CrossRef]
- Montrucchio, C.; De Nicolo, A.; D’Ettorre, G.; D’Ascenzo, F.; Lazzaro, A.; Tettoni, M.; D’Avolio, A.; Bonora, S.; Celani, L.; Di Perri, G.; et al. Serum Trimethylamine-N-oxide Concentrations in People Living with HIV and the Effect of Probiotic Supplementation. Int. J. Antimicrob Agents 2020, 55, 105908. [Google Scholar] [CrossRef]
- Grill, J.P.; Cayuela, C.; Antoine, J.M.; Schneider, F. Isolation and characterization of a Lactobacillus amylovorus mutant depleted in conjugated bile salt hydrolase activity: Relation between activity and bile salt resistance. J. Appl. Microbiol. 2000, 89, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Pellicciari, R.; Pruzanski, M.; Auwerx, J.; Schoonjans, K. Targeting bile-acid signalling for metabolic diseases. Nat. Rev. Drug Discov. 2008, 7, 678–693. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Polacek, N.; Vesper, O.; Staub, E.; Einfeldt, E.; Wilson, D.N.; Nierhaus, K.H. The highly conserved LepA is a ribosomal elongation factor that back-translocates the ribosome. Cell 2006, 127, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Arasu, M.V.; Al-Dhabi, N.A.; Ilavenil, S.; Choi, K.C.; Srigopalram, S. In vitro importance of probiotic Lactobacillus plantarum related to medical field. Saudi J. Biol. Sci. 2016, 23, S6–S10. [Google Scholar] [CrossRef] [PubMed]
- Omar, J.M.; Chan, Y.-M.; Jones, M.L.; Prakash, S.; Jones, P.J.H. Lactobacillus fermentum and Lactobacillus amylovorus as probiotics alter body adiposity and gut microflora in healthy persons. J. Funct. Foods 2013, 5, 116–123. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Koning, C.J.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics- a acomparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Strain No. a | Medium b | Source | Category |
|---|---|---|---|---|
| Lactiplantibacillus plantarum | LPL07 | MRS | Lab isolate | Anaerobic |
| Streptococcus thermophilus | BCRC 13869; | TSB | Pasteurized milk | Aerobic |
| Lactobacillus acidophilus | BCRC 10695; | MRS | Human | Anaerobic |
| Lactobacillusgasseri | BCRC 14619; | MRS | Human mouth, gut, and feces | Anaerobic |
| Ligilactobacillus salivarius | BCRC 14759; | MRS | Saliva | Anaerobic |
| Lactobacillus amylovorus | LAM1345 | MRS | Lab isolate | Anaerobic |
| Limosilactobacillus fermentum | LF1143 | MRS | Lab isolate | Anaerobic |
| Lactiplantibacillus plantarum | LP1145 | MRS | Lab isolate | Anaerobic |
| Lactobacillus gasseri | BCRC 17614; | Human | Anaerobic | |
| Lactobacillus gasseri | BCRC 17615; | MRS | Human feces | Anaerobic |
| Lactobacillus gasseri | BCRC 17616; | Vaginal track | Anaerobic | |
| Ligilactobacillus salivarius | BCRC 12574; | MRS | Saliva | Anaerobic |
| Ligilactobacillus salivarius | LSA1105 | MRS | Lab isolate | Anaerobic |
| Proteus mirabilis | BCRC10725 | NB | Urine of patient with kidney stones | Aerobic |
| Klebsiella pneumoniae subsp. pneumoniae | BCRC 10694 | NB | Human urinary tract | Aerobic |
| Escherichia fergusonii | BCRC 15582 | NB | Feces of 1-year-old boy | Aerobic |
| Escherichia coli | BCRC 15374 | TSB | Human feces | Aerobic |
| Clostridium sporogenes | BCRC 10943 | TSB | - | Anaerobic |
| Clostridium tetani | BCRC 80185 | TSB | - | Anaerobic |
| Providencia alcalifaciens | BCRC 13995 | NB | Human feces | Aerobic |
| Providencia rettgeri | BCRC 12624 | NB | - | Aerobic |
| Providencia rustigianii | BCRC 13997 | NB | Human feces | Aerobic |
| Anaerococcus hydrogenalis | BCRC 80846 | TSB | Human feces | Anaerobic |
| Anaerococcus vaginalis | BCRC 80848 | TSB | Ovarian abscess | Anaerobic |
| Anaerococcus tetradius | BCRC 80847 | TSB | Human vagina | Anaerobic |
| Hungatella hathewayi | BCRC 80852 | TSB | Human feces | Anaerobic |
| Yokenella regensburgi | BCRC 80857 | NB | Human wrist wound | Aerobic |
| Vibrio furnissii | BCRC 80856 | NB | Human feces | Aerobic |
| Olsenella uli | BCRC 80854 | TSB | Human gingival crevice | Anaerobic |
| Klebsiella pneumoniae subsp. rhinoscleromatis | BCRC 17593 | NB | Nose of a patient, Sumatra | Aerobic |
| Klebsiella variicola | BCRC 80853 | NB | - | Aerobic |
| Klebsiella oxytoca | BCRC 17136 | NB | - | Aerobic |
| Proteus penneri | BCRC 80855 | NB | Urine | Aerobic |
| g/kg | |
|---|---|
| Casein, “vitamin-free” test | 193.0 |
| DL-methionine | 3.0 |
| Sucrose | 508.4543 |
| Corn starch | 151.2497 |
| Corn oil | 50.0 |
| Cellulose | 50.0 |
| Mineral mix, AIN-76(170915) | 35.0 |
| Calcium carbonate | 4.0 |
| Vitamin E, DL-alpha tocopheryl acetate (500 IU/g) | 0.242 |
| Vitamin A palmitate (500,000 IU/g) | 0.0396 |
| Vitamin D3, cholecalciferol (500,000 IU/g) | 0.0044 |
| Vitamin mix, w/o choline, A, D, E (83171) | 5.0 |
| Ethoxyquin, antioxidant | 0.01 |
| Primer Sequences | No. of PCR Cycles and PCR Conditions | Size (bp) | Ref | |
|---|---|---|---|---|
| Lactobacillus | F: 5′-GACTTGGTTGAAATGGAAGT-3′ R: 5′-TCAGTGGTGTGGAAGTAGAA-3′ | 40 (35 s at 95 °C, 35 s at 54 °C, 35 s at 72 °C) 40 s at 72 °C | 572 | [31] |
| 16 s | F:5′AGAGTTTGATCCTGGCTCAG-3′ R: 5′-GGTTACCTTGTTACGACT-3′ | 30 (60 s at 94 °C, 60 s at 50 °C, 60 s at 72 °C) 7 min at 72 °C | 1500 | [32] |
| lepA | F: 5′-GGDCACGTRGAYTTYTCWTAYGA-3′ R:5′-GCATAVCCYTTNGTDRAWGACTT-3′ | 35 (60 s at 94 °C, 75 s at 50 °C, 60 s at 72 °C) 7 min at 72 °C | 1163 | In this study |
| Lim. fermentum | F: 5′-AATACCGCATTA CAACTTTG -3′ R: 5′-GGTTAAATACCCTCAACGTA -3′ | 35 (60 s at 94 °C, 60 s at 50 °C, 60 s at 72 °C) 7 min at 72 °C | 337 | [33] |
| L. reuteri | F: 5′-CAGACAATCTTTGATTGTTTAG-3′ R: 5′-GCTTGTTGGTTTGGGCTCTTC-3′ | 40 (30 s at 95 °C, 60 s at 60 °C, 120 s at 2 °C) 2 min at 72 °C | 300 | [34] |
| Query Strain | Subject Strain | dDDH (%) | ANI (%) |
|---|---|---|---|
| LP1145 | Lactiplantibacillus plantarum ATCC 14917 | 100 | 99.99 |
| LP1145 | Lactiplantibacillus plantarum DSM 20174 | 100 | 99.97 |
| LP1145 | Lactobacillus arizonensis DSM 13273 | 94.1 | 99.23 |
| LP1145 | Lactiplantibacillus argentoratensis DSM 16365 | 62.9 | 95.64 |
| LP1145 | Lactiplantibacillus paraplantarum DSM 10667 | 31.1 | 86.22 |
| Lpb. plantarum LP1145 | Streptococcus thermophilus BCRC 13869 | L. amylovorus LAM1345 | L. gasseri BCRC14619 | L. salivarius BCRC14759 | L. acidophilus BCRC 10695 | Lim. fermentum LF1143 | Lpb. plantarum LPL07 | L. gasseri BCRC17614 | L. gasseri BCRC 17615 | L. gasseri BCRC 17616 | L. salivarius BCRC 12574 | L. salivarius LSA1105 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Providencia alcalifaciens | 21 [++] | 21 [++] | 24 [+++] | 22 [++] | 27 [+++] | 20 [++] | 25.5 [+++] | 22 [++] | 19.5 [++] | 20.5 [++] | 20.5 [++] | 25 [+++] | 27.5 [+++] |
| Escherichia fergusonii | 23.5 [+++] | 18.5 [++] | 19.5 [++] | 20.5 [++] | 17.5 [++] | 22.5 [+++] | 21 [++] | 23 [+++] | 17 [++] | 19 [++] | 17 [++] | 17.5 [++] | 18 [++] |
| Proteus mirabilis | 24 [+++] | 20.5 [++] | 21.5 [++] | 21 [++] | 19 [++] | 24 [+++] | 20.5 [++] | 22 [++] | 17 [++] | 18.5 [++] | 18 [++] | 19 [++] | 19 [++] |
| Klebsiella Pneumoniae | 21.5 [++] | 18 [++] | 19.5 [++] | 19 [++] | 17.5 [++] | 21 [++] | 19 [++] | 20.5 [++] | 17 [++] | 17.5 [++] | 17.5 [++] | 17.5 [++] | 18 [++] |
| Providencia rustigianii | 27 [+++] | 26 [+++] | 21.5 [++] | 22 [++] | 23 [+++] | 26.5 [+++] | 26 [+++] | 24.5 [+++] | 22 [++] | 22.5 [+++] | 23.5 [+++] | 24 [+++] | 23.5 [+++] |
| Clostridium sporogenes | 11 [−] | 13 [+] | 9 [−] | 12.5 [+] | 13.5 [+] | 9 [−] | 9 [−] | 11 [−] | 12 [+] | 22 [++] | 13.5 [+] | 12.5 [+] | 13 [+] |
| Escherichia coli | 12 [+] | 11.5 [+] | 12 [+] | 12 [+] | 12 [+] | 10 [−] | 11 [−] | 11 [−] | 12.5 [+] | 11 [−] | 10 [−] | 11 [−] | 11.5 [+] |
| Clostridium tetani | 9 [−] | 9 [−] | 10 [−] | 11 [−] | 13 [+] | 9 [−] | 9 [−] | 9 [−] | 11 [−] | 11.5 [+] | 12 [+] | 14 [+] | 13 [+] |
| Providencia rettgeri | 14 [+] | 11 [−] | 11.5 [+] | 12 [+] | 11.5 [+] | 12 [+] | 12 [+] | 12 [+] | 12 [+] | 11 [−] | 11 [−] | 11 [−] | 11.5 [+] |
| Strains | TMA and LAB Bacteria per Caco2 Cells (mean ± SD) |
|---|---|
| Negative control | 0 ±0 |
| Escherichia coli | 19.2 ± 4.8 |
| Providencia rettgeri | 3.4 ± 1.3 |
| Providencia alcalifaciens | 4.6 ± 3.2 |
| Providencia rustigianii | 5.5 ± 2.1 |
| Clostridium sporogenes | 5.8 ± 2.0 |
| Clostridium terani | 9.9 ± 4.1 |
| Escherichia fergusoni | 11.2 ± 2.8 |
| Proteus mirabilis | 13.5 ± 6.5 |
| Klebsiella pneumonia | 12.0 ± 4.7 |
| Klebsiella sub rhinoscleromatis | 6.7 ± 2.9 |
| Klebsiella varicola | 14.6 ± 5.3 |
| Klebsiella oxytoca | 13.1 ± 2.1 |
| Anaerococcus hydrogenalis | 5.5 ± 2.0 |
| Anaerococcus vaginalis | 7.2 ± 2.2 |
| Anaerococcus tetradius | 6.0 ± 1.8 |
| Hungatella hathewayi | 7.4 ± 3.3 |
| Yokenella regensburgi | 10.6 ± 2.5 |
| Proteus penneri | 17.8 ± 3.9 |
| Vibrio furnisii | 12.7 ± 3.2 |
| Olsenella uli | 15.1 ± 5.3 |
| Lactobacillusgasseri (14619) | 23.5 ± 7.8 |
| Ligilactobacillus salivarius (14789) | 25.1 ± 3.5 |
| Lactiplantibacillus plantarum LPL07 | 18.7 ± 3.2 |
| Lactobacillus amylovorus LAM1345 | 273 ± 1.32 |
| Limosilactobacillus fermentum LF1143 | 218 ± 2.36 |
| Lactiplantibacillus plantarum LP1145 | 203 ± 2.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramireddy, L.; Tsen, H.-Y.; Chiang, Y.-C.; Hung, C.-Y.; Wu, S.-R.; Young, S.-L.; Lin, J.-S.; Huang, C.-H.; Chiu, S.-H.; Chen, C.-C.; et al. Molecular Identification and Selection of Probiotic Strains Able to Reduce the Serum TMAO Level in Mice Challenged with Choline. Foods 2021, 10, 2931. https://doi.org/10.3390/foods10122931
Ramireddy L, Tsen H-Y, Chiang Y-C, Hung C-Y, Wu S-R, Young S-L, Lin J-S, Huang C-H, Chiu S-H, Chen C-C, et al. Molecular Identification and Selection of Probiotic Strains Able to Reduce the Serum TMAO Level in Mice Challenged with Choline. Foods. 2021; 10(12):2931. https://doi.org/10.3390/foods10122931
Chicago/Turabian StyleRamireddy, Latha, Hau-Yang Tsen, Yu-Chen Chiang, Chen-Ying Hung, Shih-Rong Wu, San-Land Young, Jin-Seng Lin, Chien-Hsun Huang, Shih-Hau Chiu, Chien-Chi Chen, and et al. 2021. "Molecular Identification and Selection of Probiotic Strains Able to Reduce the Serum TMAO Level in Mice Challenged with Choline" Foods 10, no. 12: 2931. https://doi.org/10.3390/foods10122931
APA StyleRamireddy, L., Tsen, H.-Y., Chiang, Y.-C., Hung, C.-Y., Wu, S.-R., Young, S.-L., Lin, J.-S., Huang, C.-H., Chiu, S.-H., Chen, C.-C., & Chen, C.-C. (2021). Molecular Identification and Selection of Probiotic Strains Able to Reduce the Serum TMAO Level in Mice Challenged with Choline. Foods, 10(12), 2931. https://doi.org/10.3390/foods10122931