
Linseed Silesia, Diverse Crops for Diverse Diets. New Solutions to Increase Dietary Lipids in Crop Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants Cultivation and Yield Determination
2.2. Basic Nutrient Analysis
2.3. Determination of Main Polymers Content in the Straw and Fiber
2.4. Preparation of Oil from Seeds
2.5. Determination of Fatty Acid Composition in Oil
2.6. Extraction of Hydrophilic and Lipid-Soluble Components from Oil
2.7. UPLC-PDA-MS Analysis of Bioactive Components in Oil
2.8. Oil Stability Analysis
2.9. Statistical Analysis
3. Results
3.1. Crop Growth and Yield
3.2. Fatty Acid Composition and Sensitivity to Oxidation
3.2.1. Fatty Acid Composition in Oil Pressed from Mature Seeds
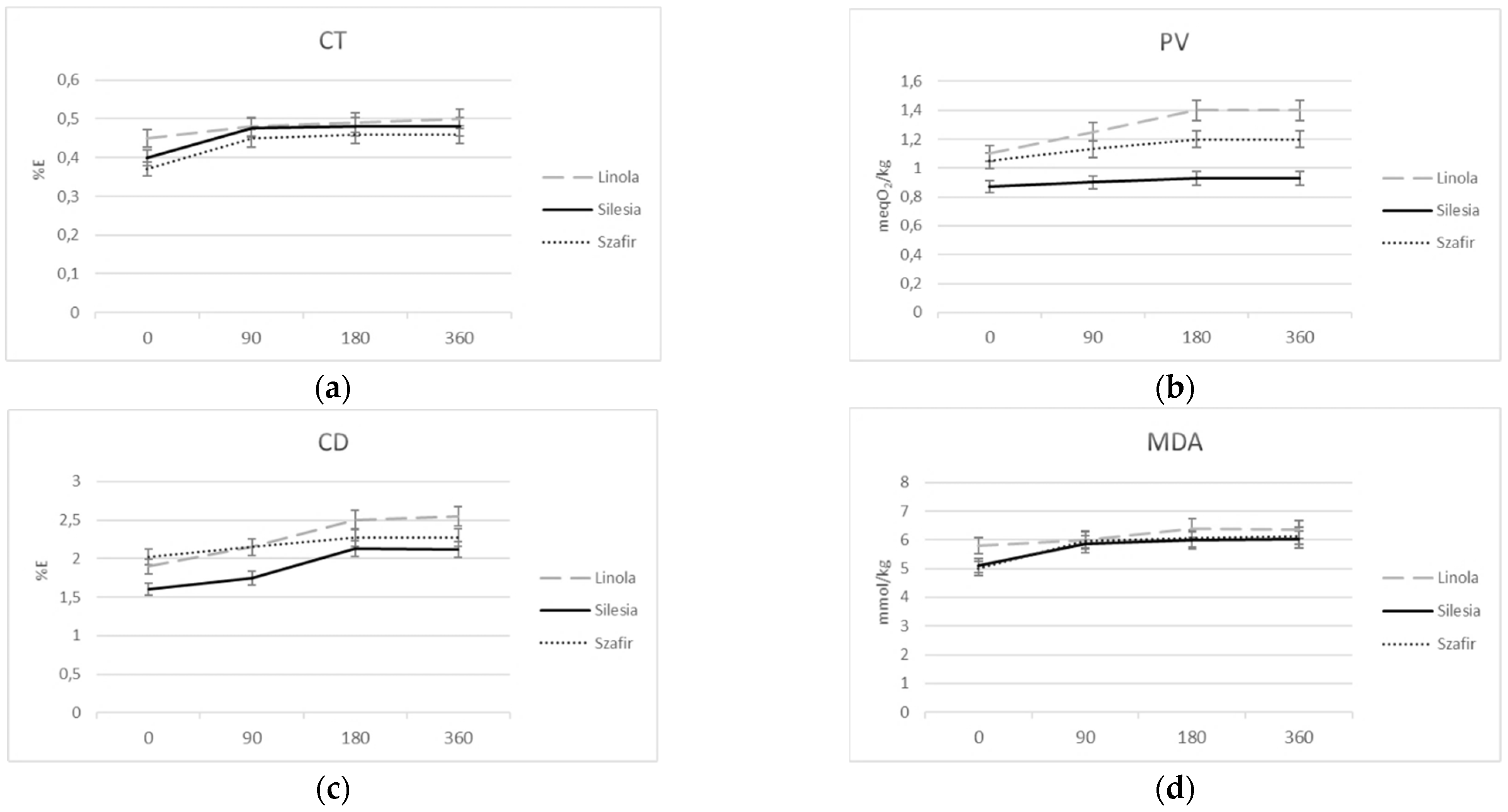
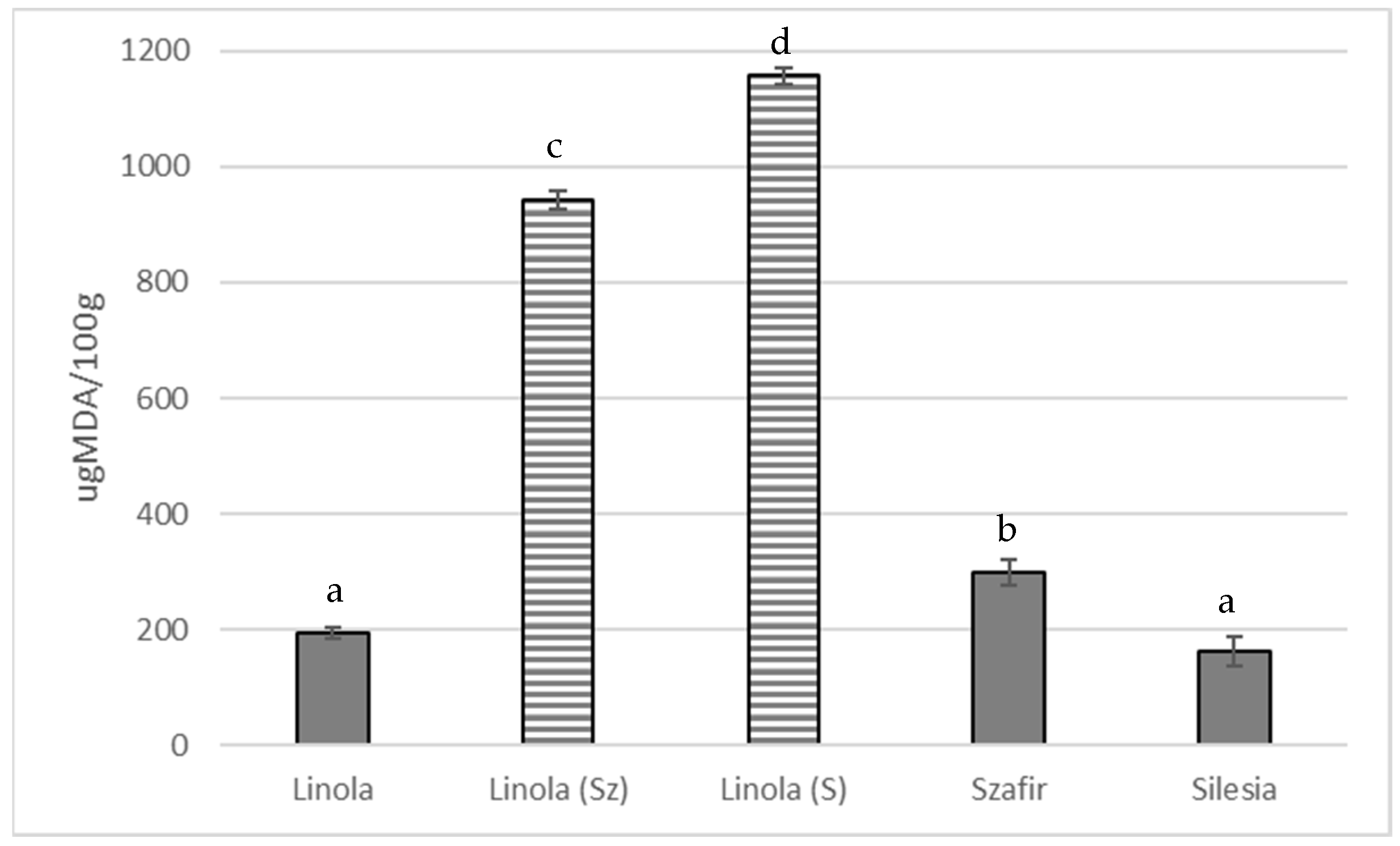
3.2.2. Flax Oil Sensitivity to Oxidation
3.3. Antioxidant Content in Oil
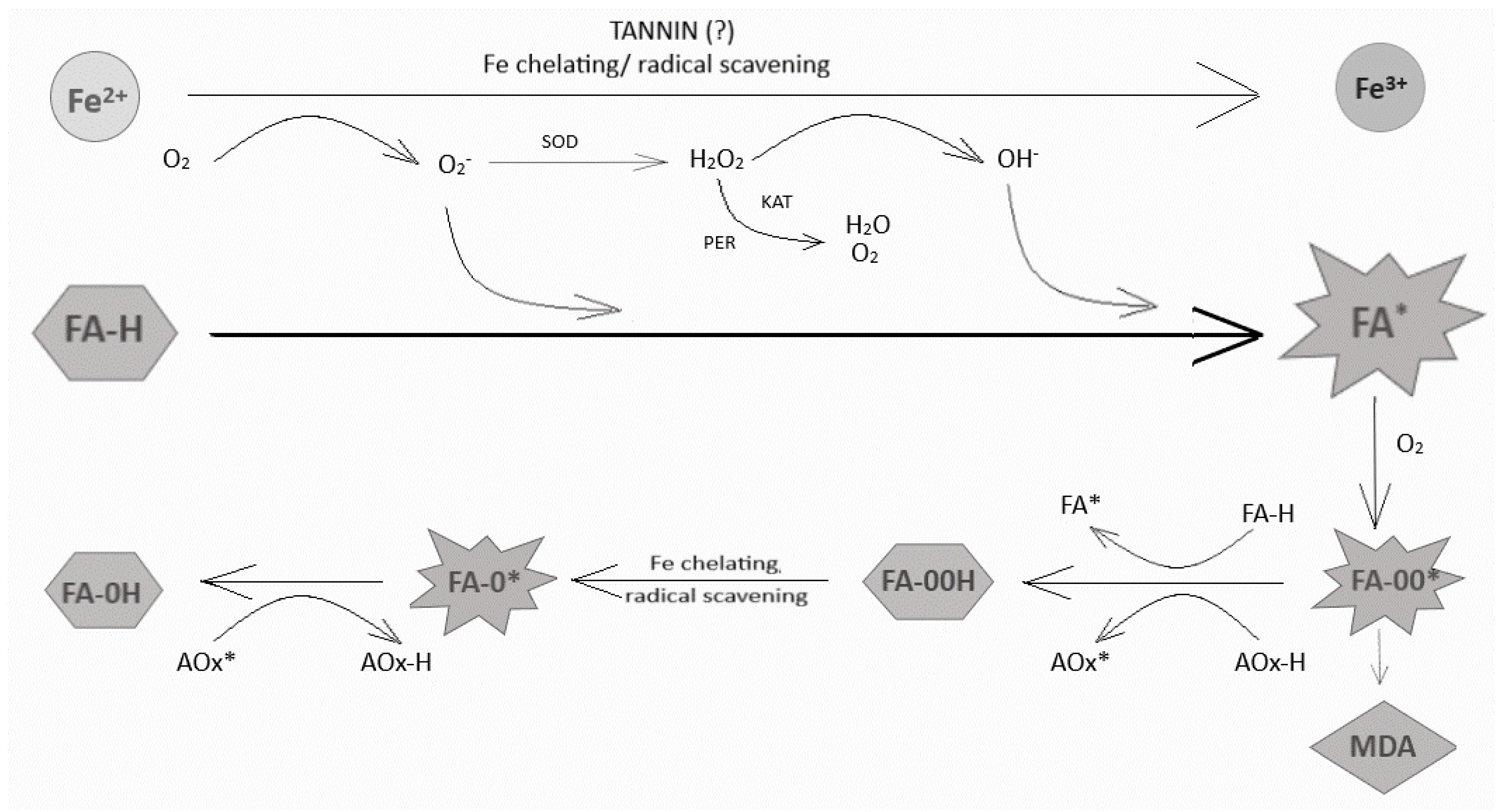
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Goyal, A.; Sharma, V.; Upadhyay, N.; Gill, S.; Sihag, M. Flax and flaxseed oil: An ancient medicine & modern functional food. J. Food Sci. Technol. 2014, 51, 1633–1653. [Google Scholar] [CrossRef] [Green Version]
- Tavarini, S.; Castagna, A.; Conte, G.; Foschi, L.; Sanmartin, C.; Incrocci, L.; Ranieri, A.; Serra, A.; Angelini, L.G. Evaluation of Chemical Composition of Two Linseed Varieties as Sources of Health-Beneficial Substances. Molecules 2019, 24, 3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, M.; Richter, D.; Matuła, J.; Szopa, J. Linseed, the multipurpose plant. Ind. Crops Prod. 2015, 75, 165–177. [Google Scholar] [CrossRef]
- Wang, D.D.; Hu, F.B. Dietary Fat and Risk of Cardiovascular Disease: Recent Controversies and Advances. Annu. Rev. Nutr. 2017, 37, 423–446. [Google Scholar] [CrossRef]
- Serhan, C.N. Novel lipid mediators and resolution mechanisms in acute inflammation: To resolve or not? Am. J. Pathol. 2010, 177, 1576–1591. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.B.; Kanungo, N.; Nayak, V.; Chainy, G.B.N.; Dandapat, J. Catechin and curcumin interact with S protein of SARS-CoV2 and ACE2 of human cell membrane: Insights from computational studies. Sci. Rep. 2021, 11, 2043. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Evaluation of green tea polyphenols as novel corona virus (SARS CoV-2) main protease (Mpro) inhibitors—An in silico docking and molecular dynamics simulation study. J. Biomol. Struct. Dyn. 2021, 39, 4362–4374. [Google Scholar] [CrossRef]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Fatimawali; Kepel, B.J.; Idroes, R.; Effendi, Y.; Sakib, S.A.; Emran, T.B. Potential of Plant Bioactive Compounds as SARS-CoV-2 Main Protease (Mpro) and Spike (S) Glycoprotein Inhibitors: A Molecular Docking Study. Scientifica 2020, 2020, 6307457. [Google Scholar] [CrossRef] [PubMed]
- El-Hadary, A.E.; Taha, M. Pomegranate peel methanolic-extract improves the shelf-life of edible-oils under accelerated oxidation conditions. Food Sci. Nutr. 2020, 8, 1798–1811. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemist (AOAC). Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; The Association: Arlington, VA, USA, 1990. [Google Scholar]
- Ververis, C.; Georghiou, K.; Christodoulakis, N.; Santas, P.; Santas, R. Fiber Dimensions, Lignin and Cellulose Content of Various Plant Materials and Their Suitability for Paper Production. Ind. Crops Prod. 2004, 19, 245–254. [Google Scholar] [CrossRef]
- Zuk, M.; Działo, M.; Richter, D.; Dymińska, L.; Matuła, J.; Kotecki, A.; Hanuza, J.; Szopa, J. Chalcone Synthase (CHS) Gene Suppression in Flax Leads to Changes in Wall Synthesis and Sensing Genes, Cell Wall Chemistry and Stem Morphology Parameters. Front. Plant Sci. 2016, 7, 894. [Google Scholar] [CrossRef] [Green Version]
- Iiyama, K.; Wallis, A.F.A. Determination of lignin in herbaceous plants by an improved acetyl bromide procedure. J. Sci. Food Agric. 1990, 51, 145–161. [Google Scholar] [CrossRef]
- Hasiewicz-Derkacz, K.; Kulma, A.; Czuj, T.; Prescha, A.; Żuk, M.; Grajzer, M.; Łukaszewicz, M.; Szopa, J. Natural phenolics greatly increase flax (Linum usitatissimum) oil stability. BMC Biotechnol. 2015, 15, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, M.; Szperlik, J.; Hnitecka, A.; Szopa, J. Temporal biosynthesis of flavone constituents in flax growth stages. Plant Physiol. Biochem. 2019, 142, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Recknagel, R.O.; Glende, E.A., Jr. Spectrophotometric detection of lipid conjugated dienes. Methods Enzym. 1984, 105, 331–337. [Google Scholar] [CrossRef]
- Agregán, R.; Franco, D.; Carballo, J.; Tomasevic, I.; Barba, F.J.; Gómez, B.; Muchenje, V.; Lorenzo, J.M. Shelf life study of healthy pork liver pâté with added seaweed extracts from Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Food Res. Int. 2018, 112, 400–411. [Google Scholar] [CrossRef]
- Das, A.K.; Nanda, P.K.; Chowdhury, N.R.; Dandapat, P.; Gagaoua, M.; Chauhan, P.; Pateiro, M.; Lorenzo, J.M. Application of Pomegranate by-Products in Muscle Foods: Oxidative Indices, Colour Stability, Shelf Life and Health Benefits. Molecules 2021, 26, 467. [Google Scholar] [CrossRef]
- Tanaka, T.; Hosokawa, M.; Yasui, Y.; Ishigamori, R.; Miyashita, K. Cancer Chemopreventive Ability of Conjugated Linolenic Acids. Int. J. Mol. Sci. 2011, 12, 7495–7509. [Google Scholar] [CrossRef] [PubMed]
- Tsuzuki, T.; Igarashi, M.; Miyazawa, T. Conjugated eicosapentaenoic acid (EPA) inhibits transplanted tumor growth via membrane lipid peroxidation in nude mice. J. Nutr. 2004, 134, 1162–1166. [Google Scholar] [CrossRef] [Green Version]
- Zuk, M.; Prescha, A.; Stryczewska, M.; Szopa, J. Engineering Flax Plants To Increase Their Antioxidant Capacity and Improve Oil Composition and Stability. J. Agric. Food Chem. 2012, 60, 5003–5012. [Google Scholar] [CrossRef]
- Said, F.; Hamed, S.; Ghada, A.; Abo-Elwafa, G. Enhancement of oxidation stability of flax seed oil by blending with stable vegetable oils. J. Appl. Sci. Res. 2012, 8, 5039–5048. [Google Scholar]
- Kazuko, O.; Tanaka, A.; Tsuji, H. Formation of the Photosynthetic Electron Transport System during the Early Phase of Greening in Barley Leaves. Plant Physiol. 1989, 91, 409–414. [Google Scholar]
- Baud, S.; Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 2010, 49, 235–249. [Google Scholar] [CrossRef]
- Vigeolas, H.; van Dongen, J.T.; Waldeck, P.; Huhn, D.; Geigenberger, P. Lipid Storage Metabolism Is Limited by the Prevailing Low Oxygen Concentrations within Developing Seeds of Oilseed Rape. Plant Physiol. 2003, 133, 2048–2060. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, S.; Ito, H.; Tanaka, R.; Tanaka, A. Chlorophyll b Reductase Plays an Essential Role in Maturation and Storability of Arabidopsis Seeds. Plant Physiol. 2012, 160, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.; Borisjuk, L.; Flügge, U.I. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Zuk, M.; Pelc, K.; Szperlik, J.; Sawula, A.; Szopa, J. Metabolism of the Cyanogenic Glucosides in Developing Flax: Metabolic Analysis, and Expression Pattern of Genes. Metabolites 2020, 10, 288. [Google Scholar] [CrossRef]
- Padiya, R.; Chowdhury, D.; Borkar, R.; Srinivas, R.; Pal Bhadra, M.; Banerjee, S.K. Garlic attenuates cardiac oxidative stress via activation of PI3K/AKT/Nrf2-Keap1 pathway in fructose-fed diabetic rat. PLoS ONE 2014, 9, e94228. [Google Scholar] [CrossRef] [PubMed]
- Devasagayam, T.; Boloor, K.; Ramasarma, T. Methods for estimating lipid peroxidation: An analysis of merits and demerits. IJBB 2003, 40, 300–308. [Google Scholar]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Zidorn, C. Plant chemophenetics—A new term for plant chemosystematics/plant chemotaxonomy in the macro-molecular era. Phytochemistry 2019, 163, 147–148. [Google Scholar] [CrossRef]
- Velasco, L.; Goffman, F.D. Tocopherol, plastochromanol and fatty acid patterns in the genus Linum. Plant Syst. Evol. 2000, 221, 77–88. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Cantile, T.; Roberto, L.; Ingenito, A.; Catania, M.R.; Roscetto, E.; Palumbo, G.; Zarrelli, A.; Pollio, A. Determination of the In Vitro and In Vivo Antimicrobial Activity on Salivary Streptococci and Lactobacilli and Chemical Characterisation of the Phenolic Content of a Plantago lanceolata Infusion. BioMed Res. Int. 2015, 2015, 286817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollio, A.; Zarrelli, A.; Romanucci, V.; Di Mauro, A.; Barra, F.; Pinto, G.; Crescenzi, E.; Roscetto, E.; Palumbo, G. Polyphenolic Profile and Targeted Bioactivity of Methanolic Extracts from Mediterranean Ethnomedicinal Plants on Human Cancer Cell Lines. Molecules 2016, 21, 395. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. Evolutionary aspects of the dietary omega-6:omega-3 fatty acid ratio: Medical implications. World Rev. Nutr. Diet. 2009, 100, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Li, F.; Li, L.; Fan, J.; Sun, X.; Yin, Y. n-6:n-3 PUFA ratio is involved in regulating lipid metabolism and inflammation in pigs. Br. J. Nutr. 2014, 111, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharm. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| 2018 | 2019 | 2020 | Average | |
|---|---|---|---|---|
| Straw crop [t/ha] | ||||
| Linola | 4.09 ± 0.10 a | 3.06 ± 0.13 a | 3.54 ± 0.12 a | 3.57 ± 0.13 a |
| Szafir | 4.33 ± 0.09 a | 3.29 ± 0.09 a | 3.84 ± 0.08 a | 3.82 ± 0.09 a |
| Silesia | 4.75 ± 0.09 b | 3.96 ± 0.10 b | 4.36 ± 0.09 a | 4.36 ± 0.09 b |
| Seed yield [t/ha] | ||||
| Linola | 2.17 ± 0.03 a | 1.89 ± 0.03 a | 1.83 ± 0.03 b | 1,96 ± 0.03 a |
| Szafir | 2.25 ± 0.05 a | 1.92 ± 0.02 a | 2.08 ± 0.04 a | 2.08 ± 0.04 a |
| Silesia | 2.23 ± 0.03 a | 2.04 ± 0.04 b | 2.13 ± 0.03 a | 2.13 ± 0.03 a |
| 1000 SW [g] | ||||
| Linola | 5.6 ± 0.53 a | 5.7 ± 0.25 a | 5.8 ± 0.43 a | 5.7 ± 0.45 a |
| Szafir | 7.2 ± 0.63 b | 7.3 ± 0.33 b | 7.5 ± 0.52 b | 7.3 ± 0.63 b |
| Silesia | 5.8 ± 0.33 a | 5.9 ± 0.63 a | 6.0 ± 0.33 a | 5.9 ± 0.53 a |
| Variety | 30 DAS | 60 DAS | 80 DAS |
|---|---|---|---|
| Linola | 26.71 ± 1.25 a | 13.62 ± 0.55 a | 8.31 ± 0.33 a |
| Szafir | 21.82 ± 0.72 b | 12.59 ± 0.45 a | 11.64 ± 0.39 b |
| Silesia | 29.53 ± 1.06 c | 19.63 ± 0.63 b | 11.74 ± 0.42 b |
| Cellulose | Hemicellulose | Pectin | Lignin | |
|---|---|---|---|---|
| The content of the main biopolymers (mg/gDW) in mature straw | ||||
| Linola | 447 ± 19.2 a | 83.5 ± 10.67 a | 182.19 ± 13.98 a | 202.6 ± 53.7 a |
| Szafir | 507 ± 18.6 b | 96.9 ± 19.2 a | 179.65 ± 14.6 a | 194.6 ± 39.6 a |
| Silesia | 543 ± 12.5 c | 138.6 ± 12.81 b | 169.58 ± 17.73 a | 188.8 ± 24.5 a |
| The content of the main biopolymers (mg/gDW) in fiber extracted from straw | ||||
| Linola | 656 ± 19.5 a | 13.69 ± 3.99 a | 16.66 ± 2.70 a | 141.19 ± 16.13 a |
| Szafir | 664 ± 28.6 a | 12.89 ± 4.36 a | 16.44 ± 3.99 a | 138.26 ± 12.99 a |
| Silesia | 692 ± 35 a | 14.54 ± 3.43 a | 16.28 ± 2.01 a | 98.77 ± 7.04 a |
| Variety | Fats | Crude Protein | Dietary Fiber |
|---|---|---|---|
| Linola | 41.3 ± 0.8 a | 20.78 ± 0.09 a | 4.23 ± 0.06 a |
| Szafir | 45.4 ± 0.5 b | 24.29 ± 0.12 b | 3.98 ± 0.06 a |
| Silesia | 44.1 ± 0.7 c | 24.20 ± 0.11 b | 4.08 ± 0.05 a |
| Variety | Linola | Szafir | Silesia |
|---|---|---|---|
| Percentage of total fatty acids | |||
| C16:0 | 6.78 ± 0.10 a | 6.01 ± 0.10 b | 4.92 ± 0.18 c |
| C16:1 | 0.08 ± 0.02 a | 0.09 ± 0.01 a | 0.08 ± 0.02 a |
| C16:2 | 0.06 ± 0.01 a | 0.06 ± 0.01 a | 0.04 ± 0.00 b |
| C18:0 | 4.12 ± 0.14 a | 5.60 ± 0.06 b | 5.67 ± 0.11 c |
| C18:1 | 19.54 ± 0.21 a | 22.81 ± 0.10 b | 20.74 ± 0.19 c |
| C18:2 | 67.38 ± 0.44 a | 12.70 ± 0.07 b | 38.75 ± 0.32 c |
| C18:3 | 2.40 ± 0.04 a | 51.88 ± 0.26 b | 29.91 ± 0.24 c |
| C20:0 | 0.13 ± 0.02 a | 0.13 ± 0.03 a | 0.11 ± 0.01 a |
| C20:1 | 0.09 ± 0.01 a | 0.12 ± 0.02 a | 0.09 ± 0.01 a |
| C22:0 | 0.06 ± 0.01 a | 0.08 ± 0.01 b | 0.07 ± 0.00 c |
| C22:1 | 0.05 ± 0.00 a | 0.05 ± 0.00 a | 0.04 ± 0.01 a |
| Sum of fatty acids mg/gFW | |||
| ∑ total | 214.38 ± 5.84 a | 258.46 ± 5.66 b | 311.18 ± 4.95 c |
| PUFA | 149.72 ± 3.48 a | 167.07 ± 3.34 b | 213.78 ± 3.44 c |
| MUFA | 42.36 ± 0.55 a | 59.63 ± 0.36 b | 65.19 ± 0.52 c |
| SFA | 23.77 ± 0.26 a | 30.55 ± 0.30 b | 33.51 ± 0.39 c |
| Compounds (µg/100g FW) | Linola | Szafir | Silesia |
|---|---|---|---|
| Ferulic acid | 0.79 ± 0.01 a | 1.39 ± 0.24 b | 2.15 ± 0.03 c |
| Coumaric acid | 0. 51 ± 0.01 a | 0.68 ± 0.03 a | 1.02 ± 0.01 b |
| Caffeic acid | 0.15 ± 0.05 a | 0.26 ± 0.02 b | 0.35 ± 0.006 c |
| Chlorogenic acid | 0.01 ± 0.001 a | 0.11 ± 0.01 a | 0.77 ± 0.018 b |
| Vanillin | 2.27 ± 0.01 a | 3.43 ± 0.72 b | 6.37 ± 0.001 c |
| Syringic aldehyde | 0.37 ± 0.01 a | 0.35 ± 0.03 a | 0. 53 ± 0.001 b |
| Coniferyl aldehyde | 0.47 ± 0.01 a | 1.58 ± 0.05 b | 2.74 ± 0.008 c |
| Proanthocyanidin | 0.15 ± 0.02 a | 0.26 ± 0.02 b | 0.27 ± 0.003 b |
| Hydrolysable tannins | 0.077 ± 0.05 a | 0.127 ± 0.05 b | 0.289 ± 0.011 c |
| γ-Tocopherol | 3579 ± 18.05 a | 3670 ± 15.85 b | 3628 ± 14.95 c |
| Plastochromanol-8 | 45.92 ± 3.05 a | 48.80 ± 5.02 b | 52.84 ± 4.35 c |
| Lutein | 1.84 ± 0.15 a | 1.84 ± 0.14 a | 1.56 ± 0.14 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuk, M.; Szperlik, J.; Szopa, J. Linseed Silesia, Diverse Crops for Diverse Diets. New Solutions to Increase Dietary Lipids in Crop Species. Foods 2021, 10, 2675. https://doi.org/10.3390/foods10112675
Zuk M, Szperlik J, Szopa J. Linseed Silesia, Diverse Crops for Diverse Diets. New Solutions to Increase Dietary Lipids in Crop Species. Foods. 2021; 10(11):2675. https://doi.org/10.3390/foods10112675
Chicago/Turabian StyleZuk, Magdalena, Jakub Szperlik, and Jan Szopa. 2021. "Linseed Silesia, Diverse Crops for Diverse Diets. New Solutions to Increase Dietary Lipids in Crop Species" Foods 10, no. 11: 2675. https://doi.org/10.3390/foods10112675
APA StyleZuk, M., Szperlik, J., & Szopa, J. (2021). Linseed Silesia, Diverse Crops for Diverse Diets. New Solutions to Increase Dietary Lipids in Crop Species. Foods, 10(11), 2675. https://doi.org/10.3390/foods10112675