
On the Role of Dietary Nitrate in the Maintenance of Systemic and Oral Health


{kind=link}
{kind=link}
Abstract
:1. Introduction
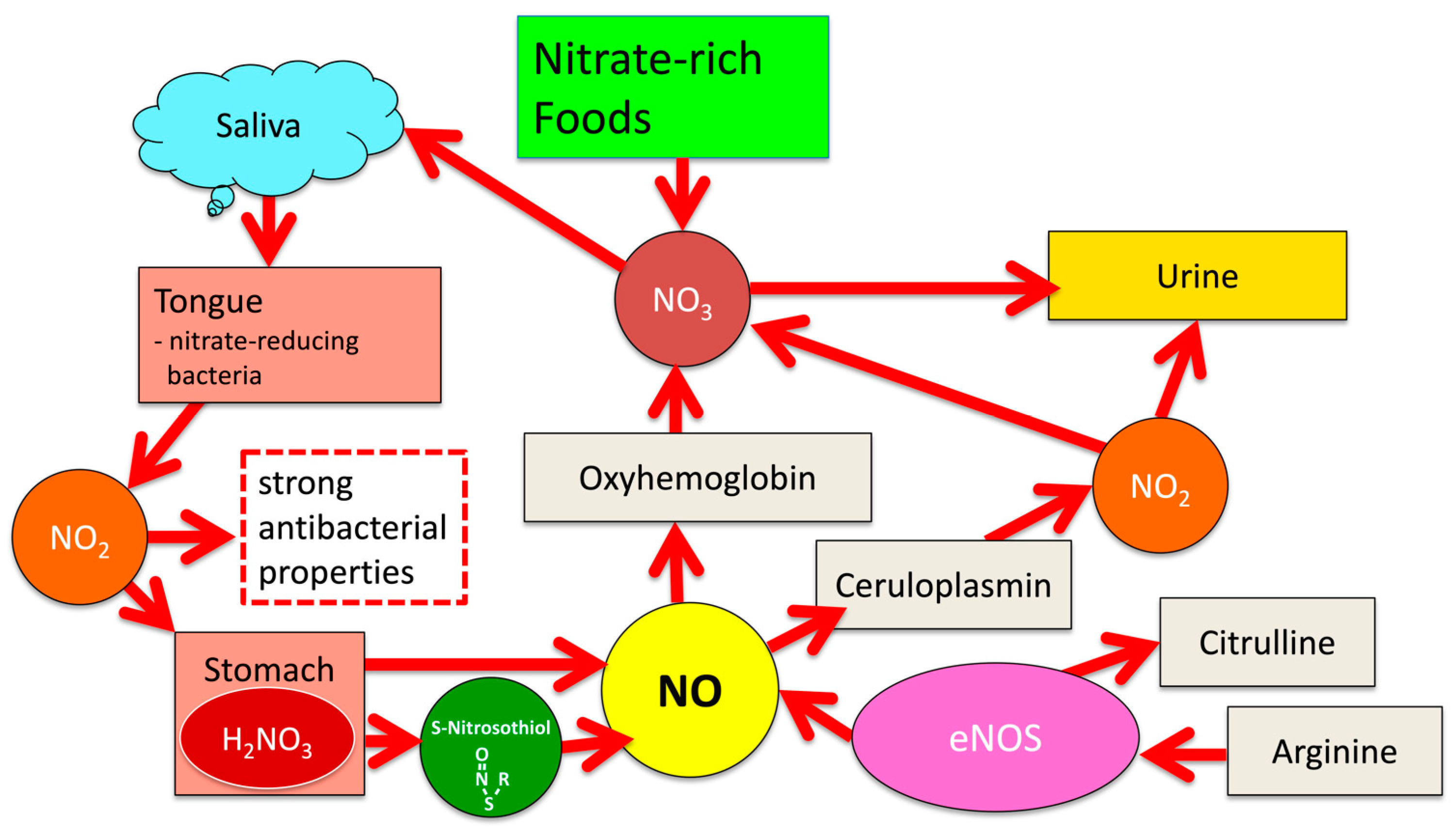
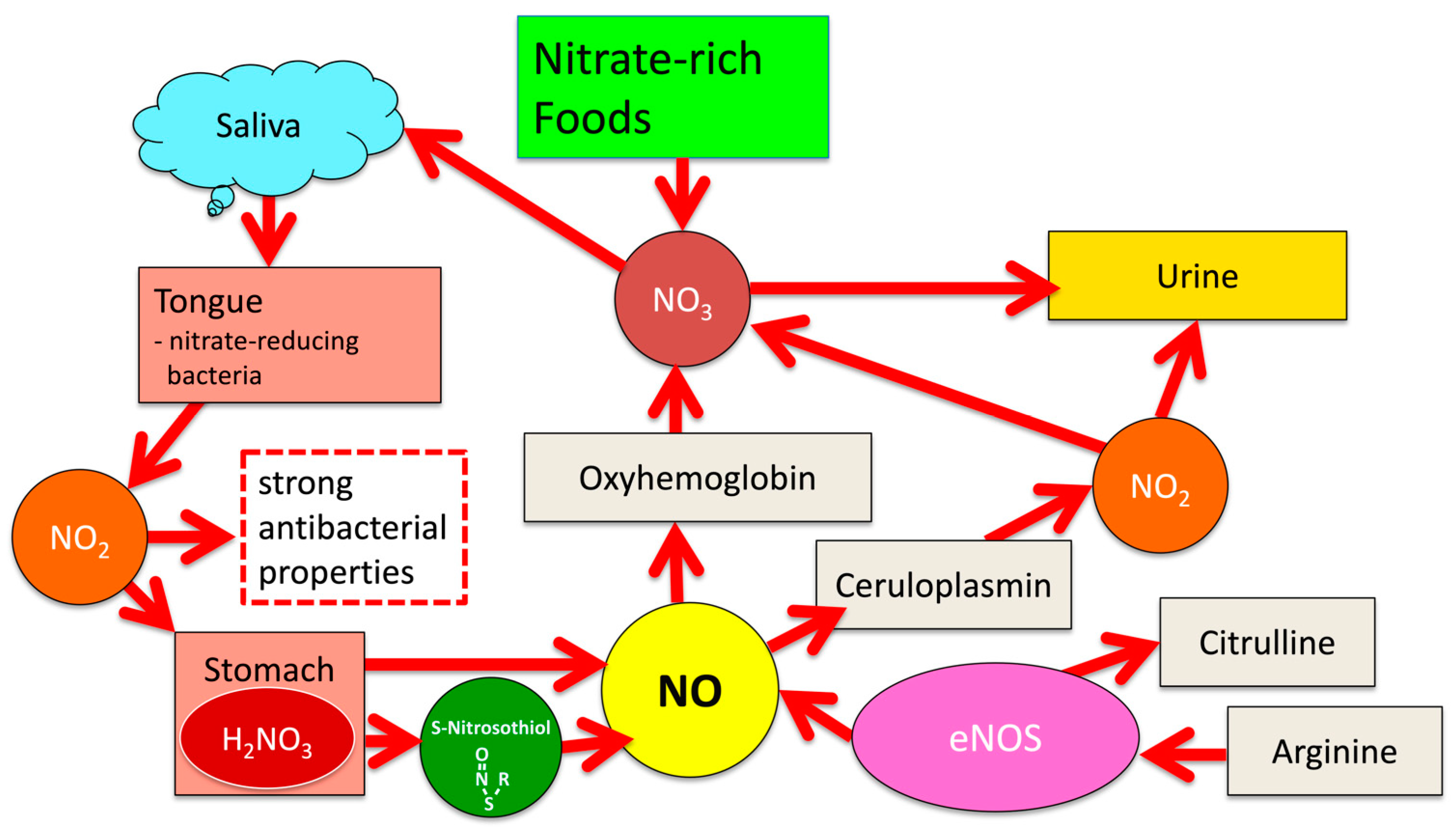
1.1. Significance of the NO-NO2-NO3 Cycle for Human Physiology
- Endothelial Nitric Oxide Synthase (eNOS)
- 2.
- Neuronal Nitric Oxide Synthase (nNOS)
- 3.
- Inducible Nitric Oxide Synthase (iNOS)
1.2. Nitric Oxide (NO)
2. NOS-Independent NO Generation from Dietary Nitrate
3. Dietary Nitrate and the Maintenance of Oral Health
3.1. Impact of Dietary Nitrate on Caries Development
3.2. Impact of Dietary Nitrate on the Development of Gingivitis and Periodontitis
3.3. Impact of Oral Antiseptics on Salivary NO2 Levels
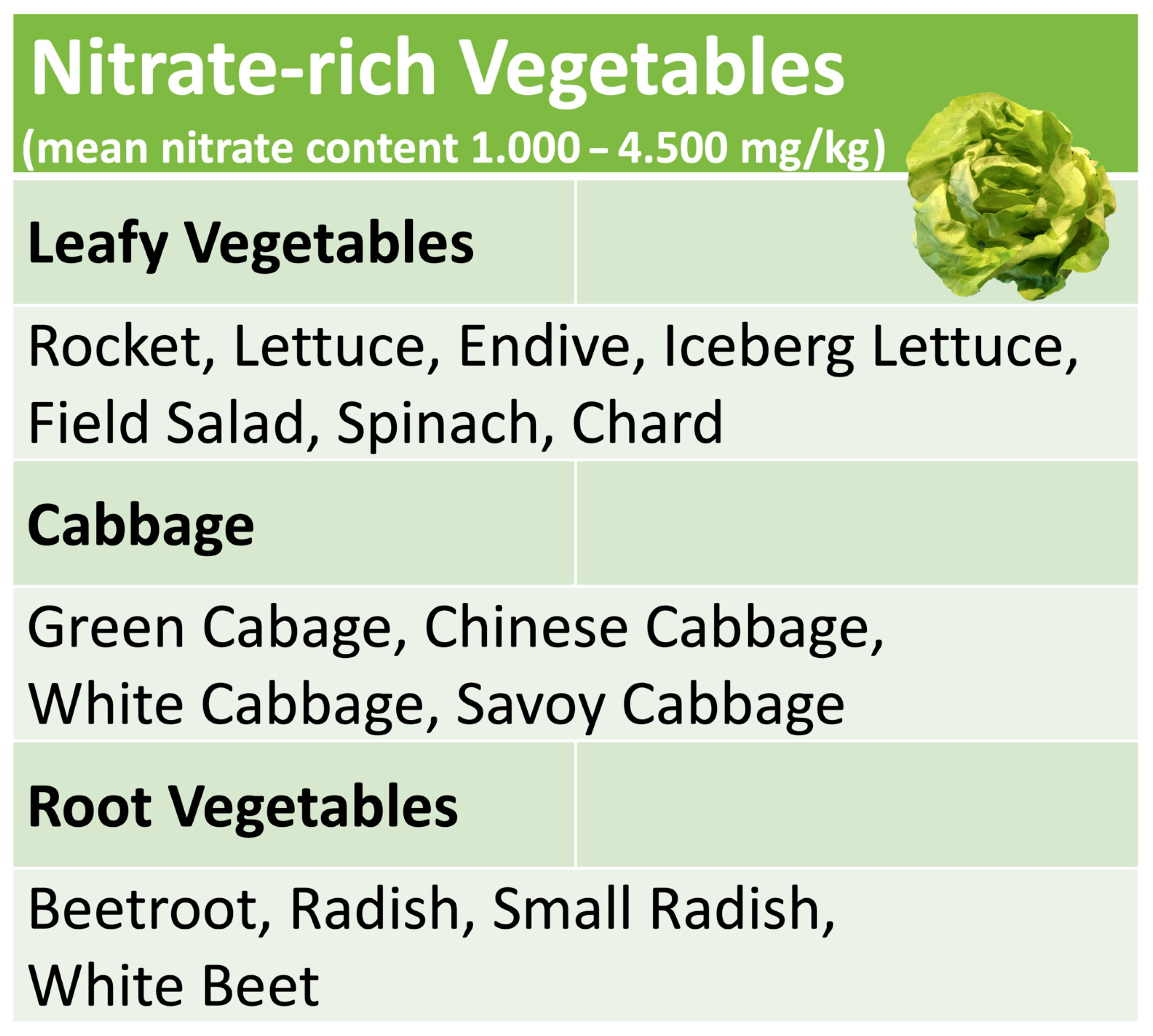
4. Nitrate-Rich Vegetables
5. Diet and Oral Health—Historical Aspects
6. Controversies about the Impact of Dietary Nitrate on General Health
7. Dietary Nitrates—Implications for Clinical Practice
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, J.D.; Liu, C.W.; Chen, Y.; Miller, A.J. Nitrogen sensing in legumes. J. Exp. Bot. 2017, 68, 1919–1926. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Furchgott, R.F.; Zawadzki, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Zeiher, A.M.; Fisslthaler, B.; Schray-Utz, B.; Busse, R. Nitric oxide modulates the expression of monocyte chemoattractant protein 1 in cultured human endothelial cells. Circ. Res. 1995, 76, 980–986. [Google Scholar] [CrossRef]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.M.; Huang, A.; Kaley, G.; Sun, D. eNOS uncoupling and endothelial dysfunction in aged vessels. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1829–H1836. [Google Scholar] [CrossRef] [Green Version]
- Karbach, S.; Wenzel, P.; Waisman, A.; Munzel, T.; Daiber, A. eNOS uncoupling in cardiovascular diseases—The role of oxidative stress and inflammation. Curr. Pharm. Des. 2014, 20, 3579–3594. [Google Scholar] [CrossRef]
- Gambardella, J.; Khondkar, W.; Morelli, M.B.; Wang, X.; Santulli, G.; Trimarco, V. Arginine and Endothelial Function. Biomedicines 2020, 8, 277. [Google Scholar] [CrossRef]
- Toda, N.; Ayajiki, K.; Okamura, T. Control of systemic and pulmonary blood pressure by nitric oxide formed through neuronal nitric oxide synthase. J. Hypertens. 2009, 27, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, X.; He, F. Mechanism and role of nitric oxide signaling in periodontitis. Exp. Ther. Med. 2019, 18, 3929–3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Torres, I.; Manzano-Pech, L.; Rubio-Ruiz, M.E.; Soto, M.E.; Guarner-Lans, V. Nitrosative Stress and Its Association with Cardiometabolic Disorders. Molecules 2020, 25, 2555. [Google Scholar] [CrossRef] [PubMed]
- Kirkeboen, K.A.; Strand, O.A. The role of nitric oxide in sepsis--an overview. Acta Anaesthesiol. Scand. 1999, 43, 275–288. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E. Biology of nitrogen oxides in the gastrointestinal tract. Gut 2013, 62, 616–629. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Khatri, J.; Mills, C.E.; Maskell, P.; Odongerel, C.; Webb, A.J. It is rocket science—Why dietary nitrate is hard to ‘beet’! Part I: Twists and turns in the realization of the nitrate-nitrite-NO pathway. Br. J. Clin. Pharmacol. 2017, 83, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, N.; O’Driscoll, F.; Dougall, H.; Duncan, C.; Smith, L.; Golden, M.; McKenzie, H. Stomach NO synthesis. Nature 1994, 368, 502. [Google Scholar] [CrossRef]
- Dykhuizen, R.S.; Frazer, R.; Duncan, C.; Smith, C.C.; Golden, M.; Benjamin, N.; Leifert, C. Antimicrobial effect of acidified nitrite on gut pathogens: Importance of dietary nitrate in host defense. Antimicrob. Agents Chemother. 1996, 40, 1422–1425. [Google Scholar] [CrossRef] [Green Version]
- Dykhuizen, R.S.; Fraser, A.; McKenzie, H.; Golden, M.; Leifert, C.; Benjamin, N. Helicobacter pylori is killed by nitrite under acidic conditions. Gut 1998, 42, 334–337. [Google Scholar] [CrossRef] [Green Version]
- Bjorne, H.; Govoni, M.; Tornberg, D.C.; Lundberg, J.O.; Weitzberg, E. Intragastric nitric oxide is abolished in intubated patients and restored by nitrite. Crit. Care Med. 2005, 33, 1722–1727. [Google Scholar] [CrossRef] [PubMed]
- Abu-Alghayth, M.; Vanhatalo, A.; Wylie, L.J.; McDonagh, S.T.; Thompson, C.; Kadach, S.; Kerr, P.; Smallwood, M.J.; Jones, A.M.; Winyard, P.G. S-nitrosothiols, and other products of nitrate metabolism, are increased in multiple human blood compartments following ingestion of beetroot juice. Redox Biol. 2021, 43, 101974. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Paula, G.H.; Tanus-Santos, J.E. Nitrite-stimulated Gastric Formation of S-nitrosothiols As An Antihypertensive Therapeutic Strategy. Curr. Drug Targets 2019, 20, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Diesen, D.L.; Hess, D.T.; Stamler, J.S. Hypoxic vasodilation by red blood cells: Evidence for an s-nitrosothiol-based signal. Circ. Res. 2008, 103, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.; Bond, R.; McLean, P.; Uppal, R.; Benjamin, N.; Ahluwalia, A. Reduction of nitrite to nitric oxide during ischemia protects against myocardial ischemia-reperfusion damage. Proc. Natl. Acad. Sci. USA 2004, 101, 13683–13688. [Google Scholar] [CrossRef] [Green Version]
- Cialoni, D.; Brizzolari, A.; Samaja, M.; Bosco, G.; Paganini, M.; Pieri, M.; Lancellotti, V.; Marroni, A. Nitric Oxide and Oxidative Stress Changes at Depth in Breath-Hold Diving. Front. Physiol. 2020, 11, 609642. [Google Scholar] [CrossRef]
- Scoffield, J.; Michalek, S.; Harber, G.; Eipers, P.; Morrow, C.; Wu, H. Dietary Nitrite Drives Disease Outcomes in Oral Polymicrobial Infections. J. Dent. Res. 2019, 98, 1020–1026. [Google Scholar] [CrossRef]
- Hohensinn, B.; Haselgrübler, R.; Müller, U.; Stadlbauer, V.; Lanzerstorfer, P.; Lirk, G.; Höglinger, O.; Weghuber, J. Sustaining elevated levels of nitrite in the oral cavity through consumption of nitrate-rich beetroot juice in young healthy adults reduces salivary pH. Nitric. Oxide. 2016, 60, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Burleigh, M.C.; Sculthorpe, N.; Henriquez, F.L.; Easton, C. Nitrate-rich beetroot juice offsets salivary acidity following carbohydrate ingestion before and after endurance exercise in healthy male runners. PLoS ONE 2020, 15, e0243755. [Google Scholar] [CrossRef]
- Huffines, J.T.; Stoner, S.N.; Baty, J.J.; Scoffield, J.A. Nitrite Triggers Reprogramming of the Oral Polymicrobial Metabolome by a Commensal Streptococcus. Front. Cell. Infect. Microbiol. 2022, 12, 833339. [Google Scholar] [CrossRef]
- Parvu, A.E.; Alb, S.F.; Craciun, A.; Taulescu, M.A. Efficacy of subantimicrobial-dose doxycycline against nitrosative stress in chronic periodontitis. Acta Pharmacol. Sin. 2013, 34, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendall, H.K.; Marshall, R.I.; Bartold, P.M. Nitric oxide and tissue destruction. Oral. Dis. 2001, 7, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Topcu Ali, O.; Akalin, F.A.; Sahbazoglu, K.B.; Yamalik, N.; Kilinc, K.; Karabulut, E.; Tozum, T.F. Nitrite and nitrate levels of gingival crevicular fluid and saliva in subjects with gingivitis and chronic periodontitis. J. Oral Maxillofac. Res. 2014, 5, e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, Q.A.; McKay, I.J.; Gonzales-Marin, C.; Allaker, R.P. Regulation of adrenomedullin and nitric oxide production by periodontal bacteria. J. Periodontal. Res. 2015, 50, 650–657. [Google Scholar] [CrossRef]
- Hussain, Q.A.; McKay, I.J.; Gonzales-Marin, C.; Allaker, R.P. Detection of adrenomedullin and nitric oxide in different forms of periodontal disease. J. Periodontal. Res. 2016, 51, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Han, D.H.; Kim, M.S.; Shin, H.S.; Park, K.P.; Kim, H.D. Association between periodontitis and salivary nitric oxide metabolites among community elderly Koreans. J. Periodontol. 2013, 84, 776–784. [Google Scholar] [CrossRef]
- Aurer, A.; Aleksić, J.; Ivić-Kardum, M.; Aurer, J.; Culo, F. Nitric oxide synthesis is decreased in periodontitis. J. Clin. Periodontol. 2001, 28, 565–568. [Google Scholar] [CrossRef]
- Jockel-Schneider, Y.; Gossner, S.K.; Petersen, N.; Stolzel, P.; Hagele, F.; Schweiggert, R.M.; Haubitz, I.; Eigenthaler, M.; Carle, R.; Schlagenhauf, U. Stimulation of the nitrate-nitrite-NO-metabolism by repeated lettuce juice consumption decreases gingival inflammation in periodontal recall patients: A randomized, double-blinded, placebo-controlled clinical trial. J. Clin. Periodontol. 2016, 43, 603–608. [Google Scholar] [CrossRef]
- Jockel-Schneider, Y.; Schlagenhauf, U.; Stolzel, P.; Gossner, S.; Carle, R.; Ehmke, B.; Prior, K.; Hagenfeld, D. Nitrate-rich diet alters the composition of the oral microbiota in periodontal recall patients. J. Periodontol. 2021, 92, 1536–1545. [Google Scholar] [CrossRef]
- Woelber, J.P.; Gartner, M.; Breuninger, L.; Anderson, A.; Konig, D.; Hellwig, E.; Al-Ahmad, A.; Vach, K.; Dotsch, A.; Ratka-Kruger, P.; et al. The influence of an anti-inflammatory diet on gingivitis. A randomized controlled trial. J. Clin. Periodontol. 2019, 46, 481–490. [Google Scholar] [CrossRef]
- Kapil, V.; Haydar, S.M.; Pearl, V.; Lundberg, J.O.; Weitzberg, E.; Ahluwalia, A. Physiological role for nitrate-reducing oral bacteria in blood pressure control. Free Radic. Biol. Med. 2013, 55, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondonno, C.P.; Liu, A.H.; Croft, K.D.; Considine, M.J.; Puddey, I.B.; Woodman, R.J.; Hodgson, J.M. Antibacterial mouthwash blunts oral nitrate reduction and increases blood pressure in treated hypertensive men and women. Am. J. Hypertens. 2015, 28, 572–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshipura, K.; Munoz-Torres, F.; Fernandez-Santiago, J.; Patel, R.P.; Lopez-Candales, A. Over-the-counter mouthwash use, nitric oxide and hypertension risk. Blood Press. 2020, 29, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Joshipura, K.J.; Munoz-Torres, F.J.; Morou-Bermudez, E.; Patel, R.P. Over-the-counter mouthwash use and risk of pre-diabetes/diabetes. Nitric Oxide 2017, 1, 14–20. [Google Scholar] [CrossRef]
- Beals, J.W.; Binns, S.E.; Davis, J.L.; Giordano, G.R.; Klochak, A.L.; Paris, H.L.; Schweder, M.M.; Peltonen, G.L.; Scalzo, R.L.; Bell, C. Concurrent Beet Juice and Carbohydrate Ingestion: Influence on Glucose Tolerance in Obese and Nonobese Adults. J. Nutr. Metab. 2017, 2017, 6436783. [Google Scholar] [CrossRef] [Green Version]
- Hughan, K.S.; Levine, A.; Helbling, N.; Anthony, S.; DeLany, J.P.; Stefanovic-Racic, M.; Goodpaster, B.H.; Gladwin, M.T. Effects of Oral Sodium Nitrite on Blood Pressure, Insulin Sensitivity, and Intima-Media Arterial Thickening in Adults With Hypertension and Metabolic Syndrome. Hypertension 2020, 76, 866–874. [Google Scholar] [CrossRef]
- Paris, S.; Banerjee, A.; Bottenberg, P.; Breschi, L.; Campus, G.; Domejean, S.; Ekstrand, K.; Giacaman, R.A.; Haak, R.; Hannig, M.; et al. How to Intervene in the Caries Process in Older Adults: A Joint ORCA and EFCD Expert Delphi Consensus Statement. Caries Res. 2020, 54, 459–465. [Google Scholar] [CrossRef]
- Sanz, M.; Herrera, D.; Kebschull, M.; Chapple, I.; Jepsen, S.; Beglundh, T.; Sculean, A.; Tonetti, M.S.; Participants, E.F.P.W.; Methodological, C. Treatment of stage I-III periodontitis-The EFP S3 level clinical practice guideline. J. Clin. Periodontol. 2020, 47 (Suppl. 22), 4–60. [Google Scholar] [CrossRef]
- Chain, E.; PoCit, F.; Alexander, J.; Bedford, D.; Cockburn, A.; Cravedi, J.P.; Dogliotti, E.; Di Domenico, A.; Fernandez-Cruz, L.; Fink-Gremmels, J.; et al. Nitrate in vegetables—Scientific opinion of the panel of contaminants in the food chain. EFSA J. 2008, 6, 698. [Google Scholar]
- Sherameti, I.; Sopory, S.K.; Trebicka, A.; Pfannschmidt, T.; Oelmuller, R. Photosynthetic electron transport determines nitrate reductase gene expression and activity in higher plants. J. Biol. Chem. 2002, 277, 46594–46600. [Google Scholar] [CrossRef]
- Salehzadeh, H.; Maleki, A.; Rezaee, R.; Shahmoradi, B.; Ponnet, K. The nitrate content of fresh and cooked vegetables and their health-related risks. PLoS ONE 2020, 15, e0227551. [Google Scholar] [CrossRef]
- San Juan, A.F.; Dominguez, R.; Lago-Rodriguez, A.; Montoya, J.J.; Tan, R.; Bailey, S.J. Effects of Dietary Nitrate Supplementation on Weightlifting Exercise Performance in Healthy Adults: A Systematic Review. Nutrients 2020, 12, 2227. [Google Scholar] [CrossRef]
- Nyakayiru, J.; Jonvik, K.L.; Trommelen, J.; Pinckaers, P.J.; Senden, J.M.; van Loon, L.J.; Verdijk, L.B. Beetroot Juice Supplementation Improves High-Intensity Intermittent Type Exercise Performance in Trained Soccer Players. Nutrients 2017, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Adler, C.J.; Dobney, K.; Weyrich, L.S.; Kaidonis, J.; Walker, A.W.; Haak, W.; Bradshaw, C.J.; Townsend, G.; Soltysiak, A.; Alt, K.W.; et al. Sequencing ancient calcified dental plaque shows changes in oral microbiota with dietary shifts of the Neolithic and Industrial revolutions. Nat. Genet. 2013, 45, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Herzog, P.; Feig, S.A. Methaemoglobinaemia in the newborn infant. Clin Haematol. 1978, 7, 75–83. [Google Scholar] [CrossRef]
- Sanchez-Echaniz, J.; Benito-Fernandez, J.; Mintegui-Raso, S. Methemoglobinemia and consumption of vegetables in infants. Pediatrics 2001, 107, 1024–1028. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, F.; Majdi, M.; Naghshi, S.; Sheikhhossein, F.; Djafarian, K.; Shab-Bidar, S. Nitrate-nitrite exposure through drinking water and diet and risk of colorectal cancer: A systematic review and meta-analysis of observational studies. Clin. Nutr. 2021, 40, 3073–3081. [Google Scholar] [CrossRef]
- Humans IWGotEoCRt. IARC monographs on the evaluation of carcinogenic risks to humans. Ingested nitrate and nitrite, and cyanobacterial peptide toxins. IARC Monogr. Eval. Carcinog. Risks Hum. 2010, 94, 1–412. [Google Scholar]
- Zhao, Z.; Yin, Z.; Zhao, Q. Red and processed meat consumption and gastric cancer risk: A systematic review and meta-analysis. Oncotarget 2017, 8, 30563–30575. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.K.; Patterson, A.J.; MacDonald-Wicks, L.K.; Bondonno, C.P.; Blekkenhorst, L.C.; Ward, N.C.; Hodgson, J.M.; Byles, J.E.; McEvoy, M.A. Dietary Nitrate and Diet Quality: An Examination of Changing Dietary Intakes within a Representative Sample of Australian Women. Nutrients 2018, 10, 1005. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S.C.; Bergkvist, L.; Wolk, A. Fruit and vegetable consumption and incidence of gastric cancer: A prospective study. Cancer Epidemiol. Biomarkers Prev. 2006, 15, 1998–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palli, D.; Russo, A.; Decarli, A. Dietary patterns, nutrient intake and gastric cancer in a high-risk area of Italy. Cancer Causes Control. 2001, 12, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Donos, N.; Calciolari, E.; Brusselaers, N.; Goldoni, M.; Bostanci, N.; Belibasakis, G.N. The adjunctive use of host modulators in non-surgical periodontal therapy. A systematic review of randomized, placebo-controlled clinical studies. J. Clin. Periodontol. 2020, 47 (Suppl. 22), 199–238. [Google Scholar] [CrossRef] [Green Version]
- Kwan, M.W.; Wong, M.C.; Wang, H.H.; Liu, K.Q.; Lee, C.L.; Yan, B.P.; Yu, C.M.; Griffiths, S.M. Compliance with the Dietary Approaches to Stop Hypertension (DASH) diet: A systematic review. PLoS ONE 2013, 8, e78412. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlagenhauf, U. On the Role of Dietary Nitrate in the Maintenance of Systemic and Oral Health. Dent. J. 2022, 10, 84. https://doi.org/10.3390/dj10050084
Schlagenhauf U. On the Role of Dietary Nitrate in the Maintenance of Systemic and Oral Health. Dentistry Journal. 2022; 10(5):84. https://doi.org/10.3390/dj10050084
Chicago/Turabian StyleSchlagenhauf, Ulrich. 2022. "On the Role of Dietary Nitrate in the Maintenance of Systemic and Oral Health" Dentistry Journal 10, no. 5: 84. https://doi.org/10.3390/dj10050084
APA StyleSchlagenhauf, U. (2022). On the Role of Dietary Nitrate in the Maintenance of Systemic and Oral Health. Dentistry Journal, 10(5), 84. https://doi.org/10.3390/dj10050084