Abstract
Metal-substituted hemoproteins have been examined by biochemists for decades, but their potential for diverse functionalities has not been thoroughly investigated. By replacing hemoproteins with non-native metals, heme-containing proteins could be capable of performing a range of chemistries not allowed for in the native protein. The metal within the heme of the oxygen-carrying hemoprotein, myoglobin, can readily be replaced with other first row transition metals such as cobalt, chromium and manganese. Upon oxidation with two-electron oxidants (ex. meta-chloroperbenzoic acid), an oxidizing intermediate is produced in manganese-substituted myoglobin. Electron paramagnetic resonance analyses confirm the oxidation of Mn(III) to Mn(IV). With the addition of weak C-H bonds of 1,4 cyclohexadiene, hydrogen atom abstraction is exhibited by the oxidizing intermediate that displays a second-order rate constant of 2.79 +/− 0.22 M−1 s−1 by the metal-oxo species. The replacement of the iron ion with a manganese ion at the active site of myoglobin displays oxidative capabilities that are not shown in native myoglobin.
1. Introduction
A major research interest of metallo-enzymologists is to improve functionalities of natural proteins for industrial and biomedical applications [1,2,3,4,5,6,7,8,9]. One way of directing reactivity of enzymes is by site-directed mutagenesis of amino acids around the active site or essential amino acids that are involved in substrate binding [2,5]. The purpose of this study is to use protein scaffolds of robust hemoproteins and to introduce alternative metals in order to promote reactivity. The focus of this study is on the globular hemoprotein, myoglobin. Myoglobin is a dioxygen storage protein predominantly found in muscle tissue. Its primary function is to store dioxygen at the Fe(II) site without getting oxidized where its ability to store dioxygen would be compromised [10,11]. The active site of myoglobin contains a heme moiety and an axial histidine ligand to the iron of the heme and a distal histidine residue that functions to anchor and stabilize dioxygen to prevent autooxidation of the iron. Myoglobin has a robust protein superstructure where the heme can readily be removed and replaced with nonnative porphyrins [3,4,6]. The metal within the heme of myoglobin has been the target of this study because the iron ion at the center of the heme cofactor can readily be replaced with other transition metals such as cobalt, chromium, ruthenium, manganese, and copper [12,13,14,15,16,17,18,19,20,21,22,23,24,25].
Manganese-substituted myoglobin is the focus of this research study because manganese does not bind dioxygen in air. The ability to replace the iron ion in myoglobin with another metal has been reported for decades, but investigations into the reactivity of transition metal-substituted myoglobin have received little attention. Transition metal complexes display varying reactivities with different transition metals even when the same types of ligands ligate them [26,27]. Therefore, it is reasonable to expect that myoglobin would show different reactivity when the iron ion is replaced with another metal. Native myoglobin can be oxidized by two-electron oxidants to form an Fe(IV)-oxo species (compound II), but this moiety does not participate in reactivity [2] that is typically associated with metal-oxo species such as oxygen transfer and cleavage of C-H or N-H bonds [28,29,30,31,32,33,34,35,36] The iron ion in myoglobin is particularly suited to perform its function of dioxygen storage and does not promote oxidative chemistry. Since the iron was replaced with manganese, which does not bind dioxygen, new types of reactivity was explored that is not displayed in native myoglobin. Reported herein are the results of a manganese-substituted myoglobin and the characterization of an oxidizing intermediate that is reactive towards a weak C-H bond.
2. Results and Discussion
Replacement of the heme in myoglobin was accomplished using the Teale-butanone extraction to remove the iron heme and manganese protoporphyrin IX chloride was added to the apomyoglobin for reconstitution [37]. Figure 1 highlights the structural similarities between metmyoglobin and manganese-substituted myoglobin [23,38]. This comparison suggests that any difference in reactivity observed in the new myoglobin protein is due to the replaced metal and not due to any structural perturbations upon replacement of the iron ion. Mn-myoglobin was first quantitated by the bichinchoninic acid and copper assay [39]. The UV/vis spectrum of Mn-myoglobin was compared to literature reports to conclude that reconstitution was successful. Purity of the manganese-substituted myoglobin was confirmed from the ratio of the 375 nm and 280 nm peaks.
The reactivity of the Mn-myoglobin was tested by the addition of two-electron oxidants. Mn-myoglobin was reactive towards most two-electron oxidants but the best yield was found by the reaction of Mn-myoglobin with m-chloroperbenzoic acid (mCPBA) and, therefore, this report focuses on the results of an intermediate in Mn-myoglobin generated using mCPBA as the oxidant. One species is observed with the addition of one equivalent of mCPBA that returns to Mn(III)-myoglobin. The generated species that forms decays over time to the resting state Mn-myoglobin suggesting that the species is an intermediate with a limited lifetime and not a new compound (Figure S1).
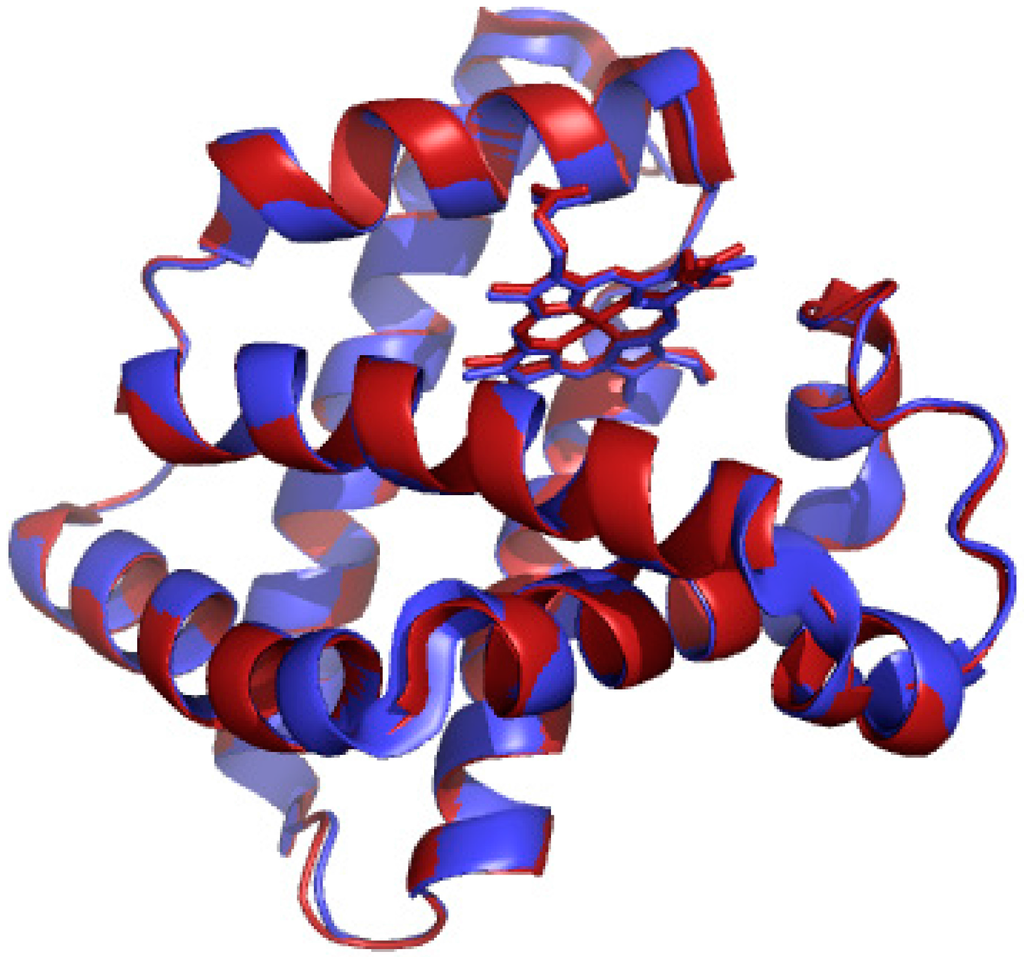
Figure 1.
X-ray crystal structures of metmyoglobin (red, pdb accession number: 1YMB) [38] and manganese-substituted myoglobin (blue, pdb accession number: 2O58) [23].
Figure 1.
X-ray crystal structures of metmyoglobin (red, pdb accession number: 1YMB) [38] and manganese-substituted myoglobin (blue, pdb accession number: 2O58) [23].
2.1. UV/vis Spectroscopy of Manganese-Substituted Myoglobin
The reaction of manganese-substituted myoglobin with two-electron oxidants yielded a distinct species as observed by UV/vis spectroscopy having an absorption maximum at 410 nm as highlighted in Figure 2. Over time (10 min at 20 °C), the starting material was regenerated indicating that the generated species is an intermediate that was somewhat unstable. The highest yield of intermediate observed was the reaction of Mn-myoglobin with meta-chloroperbenzoic acid (mCPBA). The UV/vis spectrum of the intermediate exhibits similar features to the spectrum reported by Groves and coworkers for the reaction of manganese-substituted horseradish peroxidase with mCPBA [24]. This result is not unexpected because the first coordination sphere of the active site of horseradish peroxidase is similar to active site of myoglobin and therefore should have a similar electronic structure [40].
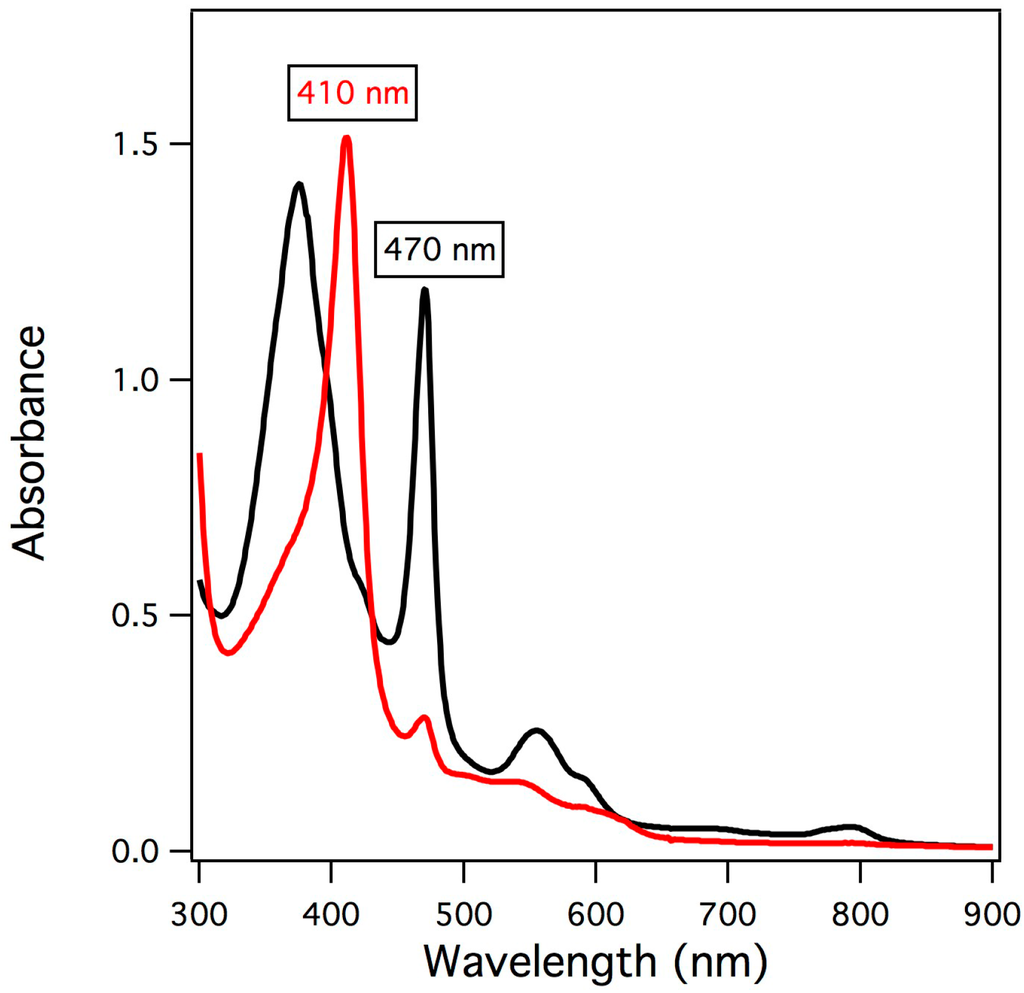
Figure 2.
UV/vis absorption spectra of Mn(III)-myoglobin (black) and upon addition of mCPBA (red).
Figure 2.
UV/vis absorption spectra of Mn(III)-myoglobin (black) and upon addition of mCPBA (red).
2.2. Electron Paramagnetic Resonance of the Mn-Myoglobin Intermediate
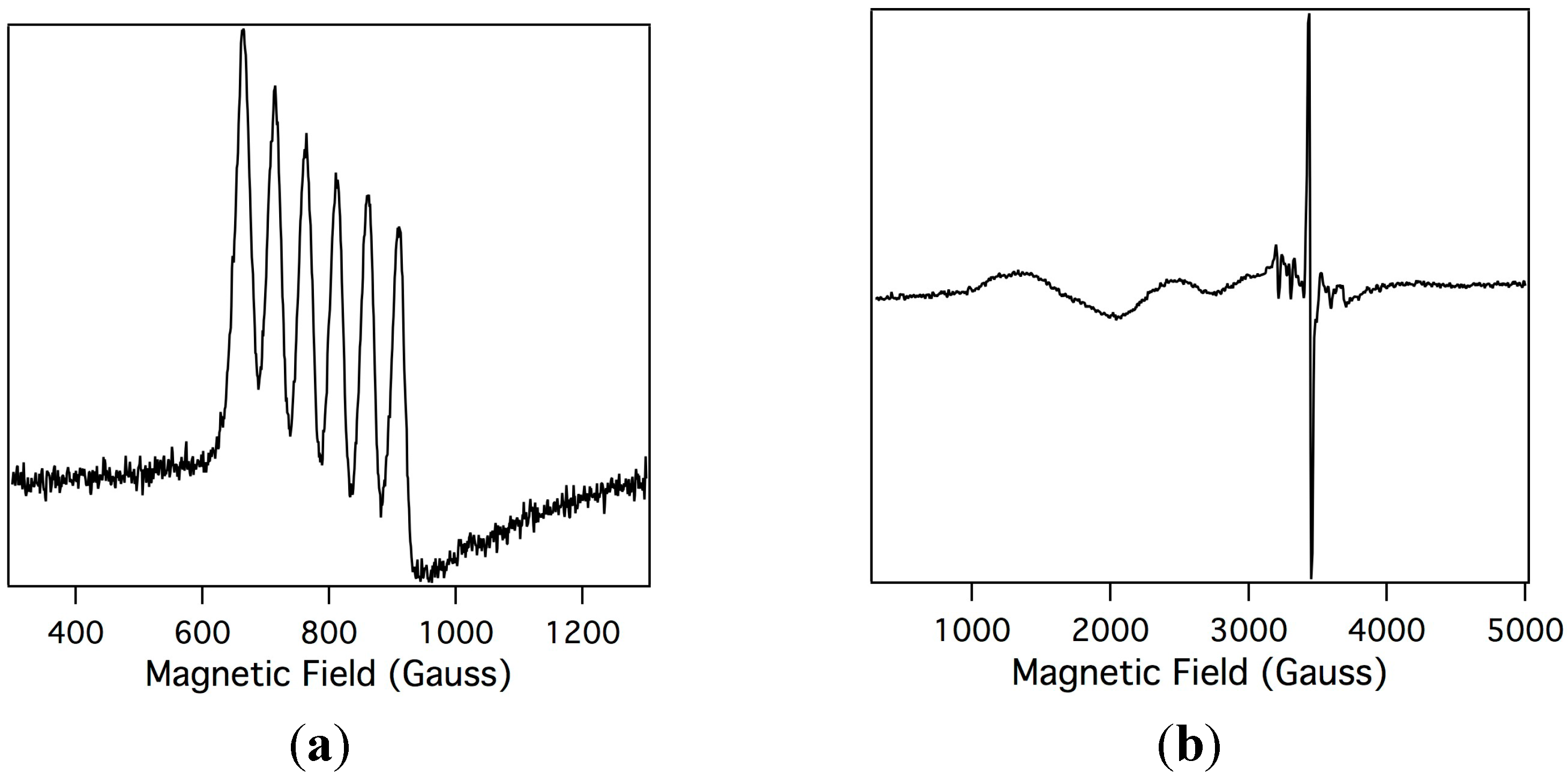
In order to understand the electronic changes upon oxidation of Mn-myoglobin, electron paramagnetic resonance spectroscopy (EPR) was performed on Mn-myoglobin and the species that is formed upon oxidation with mCPBA. In standard EPR spectroscopy the sample is aligned perpendicular to the incident radiation and this allows probing of non-integer spin systems, but when the sample is aligned parallel to the magnetic field this allows for detection of integer spins [41]. The possibilities for the spin states for a two-electron oxidized species are Mn(IV), which would show signals in perpendicular field due to three unpaired electrons, and Mn(V), which would be EPR silent due to two paired electrons. Shown in Figure 3 is the orbital diagrams of Mn(III), Mn(IV), and Mn(V) highlighting the information that can be obtained by performing EPR spectroscopy and how manganese oxidation states can be determined comparatively. Both parallel and perpendicular modes EPR were analyzed so that determination of the oxidation state of manganese ions is straightforward. Shown in Figure 4a, the parallel mode EPR spectrum of the Mn-myoglobin starting material confirms that it is a Mn(III), S = 2, displaying its characteristic six-line (I = 5/2) hyperfine splitting of a = 48 G. The hyperfine coupling constant is in agreement with other Mn(III) compounds [42,43]. Importantly, there is no signal in perpendicular mode. Upon reaction with mCPBA, the Mn(III) signal disappears in the parallel mode and a new, broad absorption band at geff ≃ 5.1 in perpendicular mode appears, shown in Figure 4b, which is in accord to other high-spin d3 (S = 3/2) manganese(IV) compounds [24,30,44]. An organic radical at g ≃ 2 also appears in the spectrum. The presence of a Mn(IV) organic radical is attributable to a compound I species of the manganese-substituted myoglobin. A small amount of residual Mn(II) is also observed at g ≃ 2. The Mn(IV) signal is comparable to other protein and synthetic reports of oxidized manganese systems. The perpendicular-mode EPR spectrum of the oxidized Mn-myoglobin indicates a higher oxidation state than the starting Mn(III)-myoglobin, presumably Mn(IV)-oxo.
A report showing the reaction between manganese-substituted horseradish peroxidase and mCPBA by Groves and coworkers shows an EPR spectrum containing a signal arising from an oxidized intermediate that was assigned as a Mn(IV)=O protein free radical. Further analysis of the manganese-substituted horseradish peroxidase species generated by reaction with mCPBA shows an 18O isotope sensitive resonance Raman band confirming the presence of a Mn-O species [24]. Myoglobin and horseradish peroxidase have a similar active site environment and it is assumed that the intermediate produced in manganese-substituted myoglobin is similar in electronic structure to the manganese-substituted horseradish peroxidase.
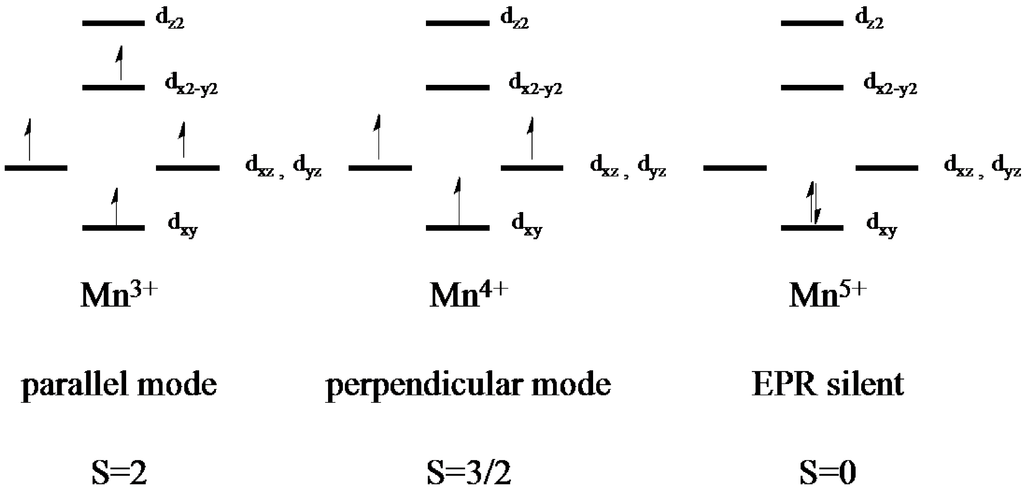
Figure 3.
d-orbital representation of spin states for Mn ions. Parallel mode EPR reveals integer spins systems while perpendicular mode EPR shows half-integer spin systems.
Figure 3.
d-orbital representation of spin states for Mn ions. Parallel mode EPR reveals integer spins systems while perpendicular mode EPR shows half-integer spin systems.
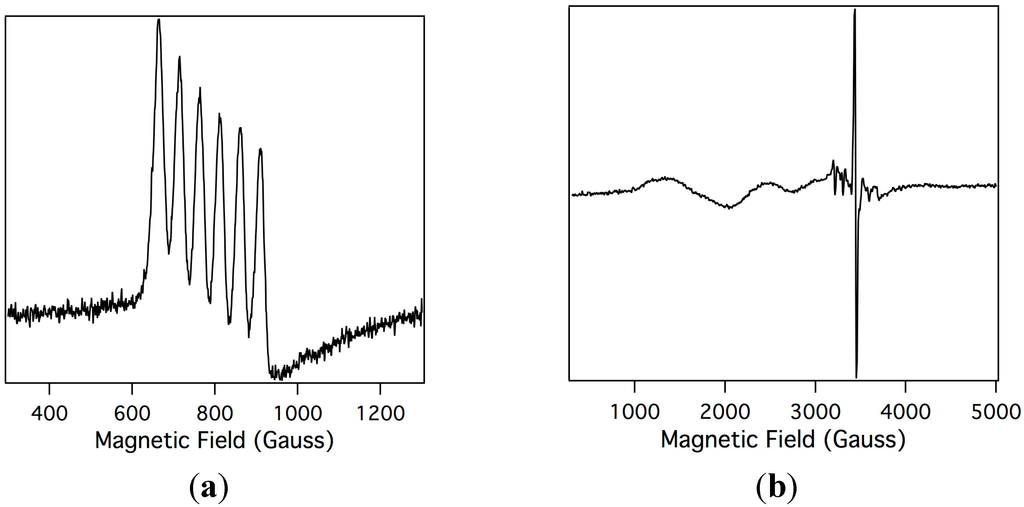
Figure 4.
X-band electron paramagnetic resonance spectroscopy (EPR) spectrum of Mn(III)-myoglobin (1 mM in 10 mM phosphate buffer pH = 7) in parallel mode (a); and the EPR spectrum of the intermediate after Mn(III)-myoglobin is reacted with mCPBA in perpendicular mode (b). Experimental conditions: microwave frequency, 9.47 GHz; microwave power, 20.26 mW; modulation amplitude, 10.02 G; temperature, 10K.
Figure 4.
X-band electron paramagnetic resonance spectroscopy (EPR) spectrum of Mn(III)-myoglobin (1 mM in 10 mM phosphate buffer pH = 7) in parallel mode (a); and the EPR spectrum of the intermediate after Mn(III)-myoglobin is reacted with mCPBA in perpendicular mode (b). Experimental conditions: microwave frequency, 9.47 GHz; microwave power, 20.26 mW; modulation amplitude, 10.02 G; temperature, 10K.
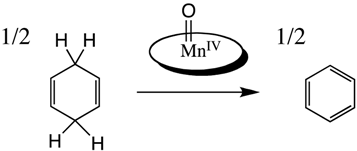
2.3. Reaction of the Mn-Myoglobin Intermediate with 1,4 Cyclohexadiene
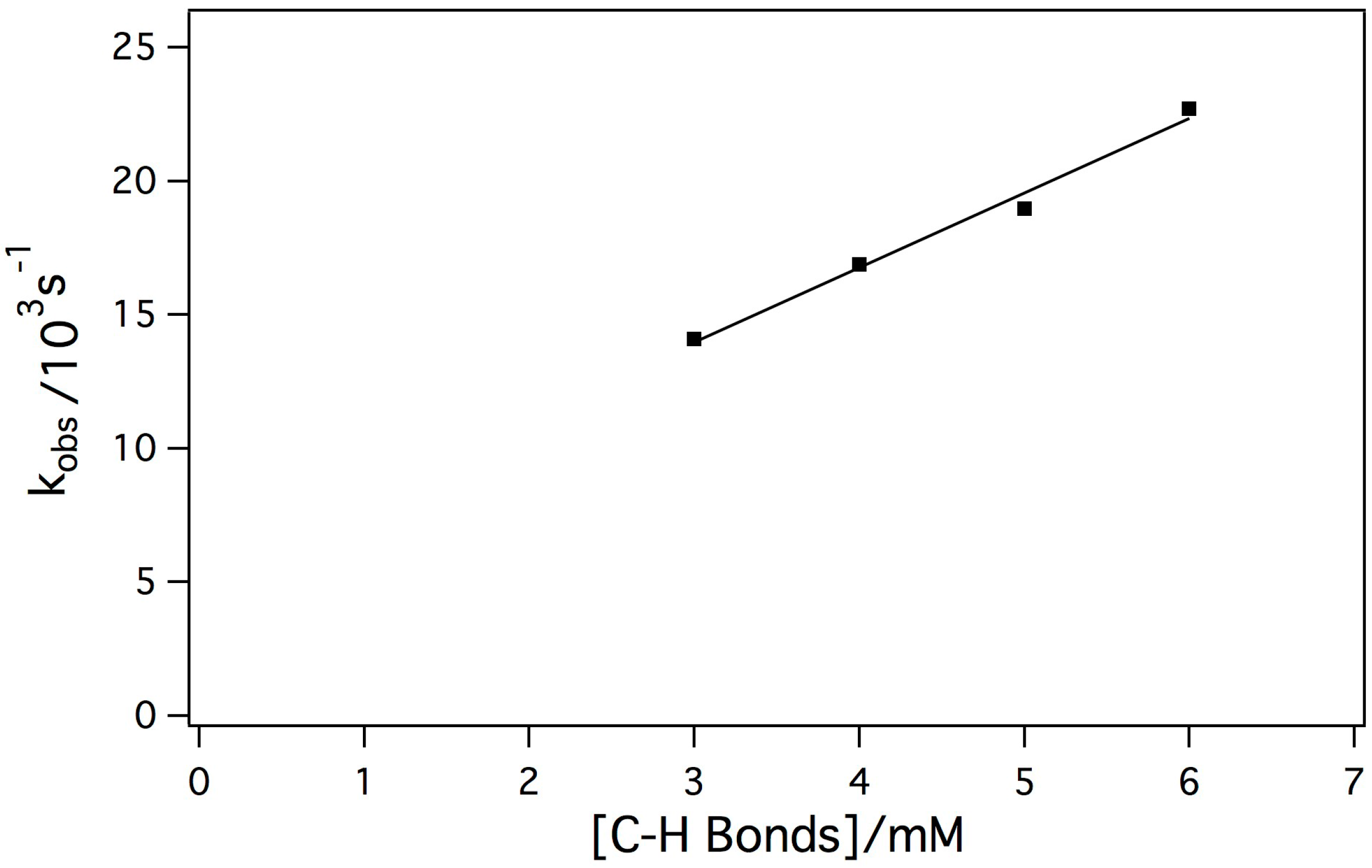
The reaction between Mn-myoglobin and mCPBA presumably forms a high-valent metal-oxo species. Taking this into account, the intermediate was reacted with a weak C-H bond to determine its ability to cleave C-H bonds since native myoglobin does not display this type of reactivity. 1,4 cyclohexadiene has a C-H bond strength of 77 kcal/mol, which is one of the weakest C-H bonds. [26,45,46] In these kinetic experiments, the concentration of the intermediate was held constant while the substrate concentration changed. Initially, it was observed that decay of the intermediate was much faster when the substrate was added indicating that the intermediate was reactive towards the substrate. The dependence of the observed rate constant (kobs) on the concentration of substrate showed a linear dependence. Shown in Figure 5 is the dependence of kobs with substrate concentration showing a linear relationship indicating a second-order reaction mechanism, which is typical for hydrogen atom abstraction reactivity. The second-order rate constant was determined to be k = 2.79 +/− 0.22 M−1 s−1. The observed second-order rate constant is comparable to other Mn(IV)-oxo species upon reaction with cyclohexadiene [44].
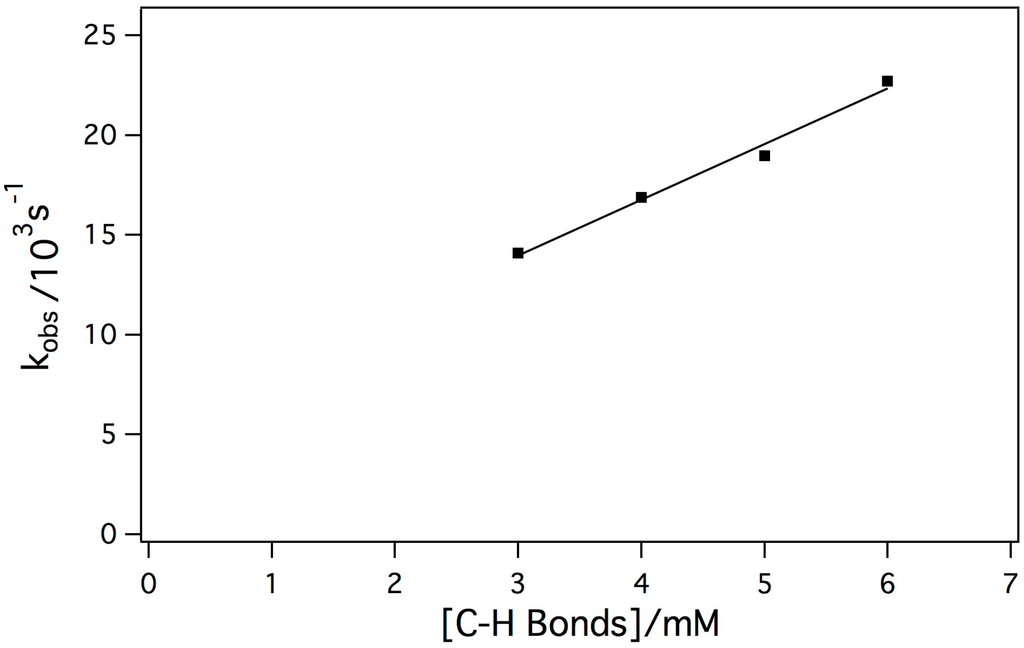
Figure 5.
Plot of kobs against substrate concentration in order to determine the second-order rate constant of the reaction between the Mn-myoglobin intermediate with 1,4 cyclohexadiene (k = 2.79 +/− 0.22 M−1s−1).
Figure 5.
Plot of kobs against substrate concentration in order to determine the second-order rate constant of the reaction between the Mn-myoglobin intermediate with 1,4 cyclohexadiene (k = 2.79 +/− 0.22 M−1s−1).
3. Experimental Section
3.1. General Methods
Horse heart myoglobin was obtained from Sigma Aldrich, the BCA assay was obtained from Pierce Protein Biology Products, Amicon® and Centricon® filters were obtained from EMD Millipore and Mn(III) Protoporphyrin IX chloride was obtained from Frontier Scientific. All other chemicals were obtained from Sigma Aldrich and unless otherwise indicated used as received. mCPBA was recrystallized by a 1:3 petroleum ether and ether mixture.
3.2. Instrumentation
UV/vis spectra were recorded on a Hewlett Packard 8453 diode array spectrophotometer equipped with a UNISOKU Scientific Instruments cryostat for temperature-dependent kinetic analyses. X-band EPR spectra were collected using a Bruker EMX spectrometer equipped with an ER041XG microwave bridge. Spectra for EPR samples were collected using the following spectrometer settings: attenuation = 10 dB, microwave power = 20.26 mW, frequency = 9.47 GHz, modulation amplitude = 10.02 G, gain = 1.00 × 103, and resolution = 1024 points. Low-temperature spectra (10K) were obtained using an Oxford Instrument liquid He quartz cryostat.
3.3. Preparation of Mn-Myoglobin
Mn-myoglobin was prepared according to previously reported procedures by extraction of the heme and reconstitution with Mn(III) protoporphyrin IX chloride [37]. After reconstitution, the red protein solution was run through a CM-52 (Whatman) packed column equilibrated with 10 mM phosphate buffer (pH 7) and eluted with 0.5 M phosphate buffer. Purity of the protein was checked by UV/vis spectroscopy by measuring the 375 nm/280 nm ratio (Rz value ~ 1). The concentration of the protein is determined by the 375 nm peak (ε= 70,000 M−1cm−1), which was quantitated using the BCA assay [39].
3.4. Preparation of EPR Samples
Mn-myoglobin solution is concentrated in an Amicon® and Centricon® filters to a concentration of 1 mM in 10 mM phosphate buffer, pH = 7. A five-fold excess of mCPBA was added to the solution and mixed. An aliquot of the protein solution was taken for UV/vis analysis to confirm formation of the intermediate. The protein solution was added to an EPR tube and slowly frozen in liquid N2. The samples were stored in liquid N2 to prevent decay of the intermediate.
3.5. Kinetic analyses
All reactions were followed by UV/vis spectrophotometry at 20 °C. Control experiments were performed in the absence of substrate. All representative substrate concentration reactions were run in triplicate and averages of the runs are reported. 20 μM Mn-myoglobin in 10 mM phosphate buffer, pH = 7, was used for all reactions and at least a five-fold excess was added to reaction mixture to generate the intermediate. Upon generation of the intermediate, the appropriate amount of substrate was added to the mixture with constant stirring. A spectrum was collected every 4 s and the resulting curve was fit to a first-order model to find kobs. kobs was plotted vs. concentration of C-H bonds to find the concentration dependence of the rate on substrate concentration. The second-order rate constant was determined from the slope of the line.
4. Conclusions
The replacement of the iron ion with a manganese ion within the heme of the oxygen storage protein, myoglobin, has resulted in a protein that has enhanced reactivity as demonstrated by the resulting C-H bond cleavage ability. The oxidation of Mn-myoglobin by mCPBA showed an intermediate that was reactive toward 1,4 cyclohexadiene. The intermediate was characterized by UV/vis spectroscopy and EPR spectroscopy and was tentatively assigned as an Mn(IV)-oxo species in accordance with the EPR spectrum and in comparison with the fully characterized manganese-substituted horseradish peroxidase by Groves and coworkers [24]. These findings show that the Mn(IV)-oxo species in Mn- myoglobin is an effective oxidant towards weak C-H bonds.
Acknowledgments
The authors would like to thank Research Corporation for Science Advancement (20046) for financial support and to A. S. Borovik and Sarah Cook at the University of California-Irvine for electron paramagnetic resonance measurements.
Author Contributions
K. L. S. conceived and designed the experiments. J. H. performed the preparation and purification of manganese-substituted myoglobin. J. H. prepared the samples for EPR. H. C. performed the kinetic analyses. K. L. S. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomas, C.M.; Ward, T.R. Design of artificial metalloenzymes. Appl. Organometallic Chem. 2005, 19, 35–39. [Google Scholar] [CrossRef]
- Ozaki, S.-I.; Matsui, T.; Watanabe, Y. Conversion of Myoglobin into a Highly Stereo-specific Peroxygenase by the L29H/H64L Mutation. J. Am. Chem. Soc. 1996, 118, 9784–9785. [Google Scholar] [CrossRef]
- Ueno, T.; Ohashi, M.; Kono, M.; Kondo, K.; Suzuki, A.; Yamane, T.; Watanabe, Y. Crystal Structures of Artificial Metalloproteins: Tight Binding of FeIII(Schiff-Base) by Mutation of Ala71 to Gly in Apo-Myoglobin. Inorg. Chem. 2004, 43, 2852–2858. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, N.; Ueno, T.; Unno, M.; Matsui, T.; Ikeda-Saito, M.; Watanabe, Y. Ligand design for the improvement of stability of metal complex: Protein hybrids. Chem. Comm. 2008, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Huang, X.; Sun, S.; Wang, C.; Lebioda, L.; Dawson, J.H. Amphitrite ornata Dehaloperoxidase (DHP): Investigations of Structural Factors that Influence the Mechanism of Halophenol Dehalogenation Using “Peroxidase-like” Myoglobin Mutants and “Myoglobin-like” DHP Mutants. Biochemistry 2011, 50, 8172–7180. [Google Scholar] [CrossRef] [PubMed]
- Garner, D.K.; Liang, L.; Barrios, D.A.; Zhang, J.-L.; Lu, Y. The Important Role of Covalent Anchor Positions in Tuning Catalytic Properties of a Rationally Designed MnSalen-Containing Metalloenzyme. ACS Catal. 2011, 1, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-W.; Yeung, N.; Gao, Y.-G.; Miner, K.D.; Lei, L.; Robinson, H.; Lu, Y. Introducing a 2-His-1-Glu Non-heme Iron Center into Myoglobin Confers Nitric Oxide Reductase Activity. J. Am. Chem. Soc. 2010, 132, 9970–9972. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yeung, N.; Sieracki, N.; Marshall, N.M. Design of Functional Metalloproteins. Nature 2009, 460, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yeung, N.; Russell, B.S.; Garner, D.K.; Lu, Y. Catalytic Reduction of NO to N2O by a Designed Heme-Copper Center in Myoglobin: Implications for the Role of Metal Ions. J. Am. Chem. Soc. 2006, 128, 6766–6767. [Google Scholar] [CrossRef] [PubMed]
- Springer, B.A.; Sligar, S.G.; Olson, J.S.; Phillips, G.M., Jr. Mechansims of Ligand Recognition in Myoglobin. Chem. Rev. 1994, 94, 699–714. [Google Scholar] [CrossRef]
- Møller, J.K.S.; Skibsted, L. Nitric Oxide and Myoglobins. Chem. Rev. 2002, 102, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Dejima, H.; Matsuo, T.; Sato, H.; Murata, D.; Hisaeda, Y. Blue Myoglobin Reconstituted with an Iron Porphycene Shows Extremely High Oxygen Affinity. J. Am. Chem. Soc. 2002, 124, 11226–11227. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Hayashi, A.; Abe, M.; Matsuda, T.; Hisaeda, Y.; Hayashi, T. Meso-Unsubstituted Iron Corrole in Hemoproteins: Remarkable Differences in Effects on Peroxidase Activities between Myoglobin and Horseradish Peroxidase. J. Am. Chem. Soc. 2009, 131, 15124–15125. [Google Scholar] [CrossRef] [PubMed]
- Oohora, K.; Kihira, Y.; Mizohata, E.; Inoue, T.; Hayashi, T. C(sp3)-H hydroxylation catalyzed by Myoglobin reconstituted with manganese porphycene. J. Am. Chem. Soc. 2013, 135, 17282–17285. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Ikeda-Saito, M.; Yonetani, T. Electron paramagnetic resonance and spectrophotometric studies of the peroxide compounds of manganese-substituted horseradish peroxidase, cytochrome-c peroxidase, and manganese-porphyrin model complexes. Biochim. Biophys. Acta 1987, 912, 74–81. [Google Scholar] [CrossRef]
- Waterman, M.R.; Yonetani, T. Studies on Modified Hemoglobins I. Properties of Hybrid Hemoglobins Containing Manganese Protoporphyrin IX. J. Biol. Chem. 1970, 245, 5847–5852. [Google Scholar] [PubMed]
- Yonetani, T.; Asakura, T. Studies on Cytochrome c Peroxidase: IV. Comparison of Manganese Porphyrin-Containing Cytochrome c Peroxidase, Horseradish Peroxidase, and Myoglobin. J. Biol. Chem. 1969, 244, 4580–4588. [Google Scholar] [PubMed]
- Dickinson, L.C. Metal Replaced Hemoproteins. J. Chem. Educ. 1976, 53, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Z.; Taniguchi, I.; Mulchandani, A. Redox properties of engineered ruthenium myoglobin. Bioelectrochemistry 2009, 75, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Mondal, M.S.; Mazumdar, S.; Mitra, S. Binding of Cyanide and Thiocyanate to Manganese Reconstituted Myoglobin and Formation of Peroxide Compound: Optical Spectral, Multinuclear NMR, and Kinetic Studies. Inorg. Chem. 1993, 32, 5362–5367. [Google Scholar] [CrossRef]
- Mondal, M.S.; Mitra, S. Kinetic studies of the two-step reactions of H2O2 with manganese-reconstituted myoglobin. Biochim. Biophys. Acta 1996, 1296, 174–180. [Google Scholar] [CrossRef]
- Paulson, D.R.; Addison, A.W.; Dolphin, D.; James, B.R. Preparation of Ruthenium(II) and Ruthenum(III) Myoglobin and the Reaction of Dioxygen, and Carbon Monoxide, with Ruthenium (II) Myoglobin. J. Biol. Chem. 1979, 254, 7002–7006. [Google Scholar] [PubMed]
- Zahran, Z.N.; Chooback, L.; Copeland, D.M.; West, A.H.; Richter-Addo, G.B. Crystal structures of manganese- and cobalt-substituted myoglobin in complex with NO and nitrite reveal unusal ligand conformations. J. Inorg. Biochem. 2008, 102, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Nick, R.J.; Ray, G.B.; Fish, K.M.; Spiro, T.G.; Groves, J.T. Evidence for a Weak Mn=O Bond and a Non-Porphyrin Radical in Manganese-Substituted Horseradish Peroxidase Compound I. J. Am. Chem. Soc. 1991, 113, 1838–1840. [Google Scholar] [CrossRef]
- Heinecke, J.L.; Yi, J.; Pereira, J.C.M.; Richter-Addo, G.B.; Ford, P.C. Nitrite reduction by CoII and MnII substituted myoglobins. Towards understanding necessary components of Mb nitrite reductase activity. J. Inorg. Biochem. 2012, 107, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Borovik, A.S. Monomeric Mn and Fe Complexes with Terminal Hydroxo and Oxo Ligands: Probing Reactivity via O-H Bond Dissociation Energies. J. Am. Chem. Soc. 2003, 125, 13234–13242. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.P. Corrolazines: New Frontiers in High-Valent Metalloporphyrinoid Stability and Reactivity. Acc. Chem. Res. 2007, 40, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Zdilla, M.J.; Abu-Omar, M.M. Manganese(III) Corrole-Oxidant Adduct as the Active Intermediate in Catalytic Hydrogen Atom Transfer. Inorg. Chem. 2007, 47, 10718–10722. [Google Scholar] [CrossRef] [PubMed]
- Groves, J.T.; Gross, Z.; Stern, M.T. Preparation and Reactivity of Oxoiron(IV) Porphyrins. Inorg. Chem. 1994, 33, 5065–5072. [Google Scholar] [CrossRef]
- Parsell, T.H.; Behan, R.K.; Green, M.T.; Hendrich, M.P.; Borovik, A.S. Preparation and Properties of a Monomeric MnIV-Oxo complex. J. Am. Chem. Soc. 2006, 128, 8728–8729. [Google Scholar] [CrossRef] [PubMed]
- Parsell, T.H.; Yang, M.-Y.; Borovik, A.S. C-H Bond Cleavage with Reductants: Re-Investigating the Reactivity of Monomeric MnIII/IV-Oxo Complexes and the Role of Oxo Ligand Basicity. J. Am. Chem. Soc. 2009, 131, 2762–2763. [Google Scholar] [CrossRef] [PubMed]
- Prokop, K.A.; Neu, H.M.; de Visser, S.P.; Goldberg, D.P. A Manganese(V)-Oxo pi-Cation Radical Complex: Influence of One-Electron Oxidation on Oxygen-Atom Transfer. J. Am. Chem. Soc. 2011, 133, 15874–15877. [Google Scholar] [CrossRef] [PubMed]
- McGown, A.J.; Kerber, W.D.; Fujii, H.; Goldberg, D.P. Catalytic Reactivity of a Mesa-N-Substituted Corrole and Evidence for a High-Valent Iron-Oxo Species. J. Am. Chem. Soc. 2009, 131, 8040–8048. [Google Scholar] [CrossRef] [PubMed]
- Leeladee, P.; Goldberg, D.P. Epoxidation Catalyzed by Manganese(V) Oxo and Imido Complexes: Role of the Oxidant-Mn-Oxo (Imido) Intermediates. Inorg. Chem. 2010, 49, 3083–3085. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Leeladee, P.; McGown, A.J.; DeBeer, S.; Goldberg, D.P. A high-valent Iron-oxo Corrolazine Activates C-H Bonds via Hydrogen-atom Transfer. J. Am. Chem. Soc. 2012, 134, 7392–7399. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Goldberg, I.; Botoshansky, M.; Buchman, Y.; Gross, Z. Oxygen Atom Transfer Reactions from Isolated (Oxo)manganese(V) Corroles to Sulfides. J. Am. Chem. Soc. 2010, 132, 15233–15245. [Google Scholar] [CrossRef] [PubMed]
- Teale, F.W. Cleavage of the haem-protein link by acid methylethylketone. Biochim. Biophys. Acta 1959, 35, 543. [Google Scholar] [CrossRef]
- Evans, S.V.; Brayer, G.D. High-resolution study of the three-dimensional structure of horse heart metmyoglobin. J. Mol. Biol. 1990, 213, 885–897. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Dunford, H.B. Heme Peroxidases; John Wiley & Sons, Inc.: New York, NY, USA, 1999. [Google Scholar]
- Drago, R.S. Physical Methods for Chemists, 2nd ed.; Saunders College Publishing: Philadelphia, PA, USA, 1992. [Google Scholar]
- Campbell, K.A.; Lashley, M.R.; Wyatt, J.K.; Nantz, M.H.; Britt, R.D. Dual-Mode EPR Study of Mn(III) Salen and the Mn(III) Salen-Catalyzed Epoxidation of cis-β-Methylstyrene. J. Am. Chem. Soc. 2001, 123, 5710–5719. [Google Scholar] [CrossRef] [PubMed]
- Shook, R.L.; Gunderson, W.A.; Greaves, J.; Ziller, J.W.; Hendrich, M.P.; Borovik, A.S. A Monomeric MnIII-Peroxo Complex Derived Directly from Dioxygen. J. Am. Chem. Soc. 2008, 130, 8888–8889. [Google Scholar] [CrossRef] [PubMed]
- Sawant, S.C.; Wu, X.; Cho, J.; Cho, K.-B.; Kim, S.H.; Seo, M.S.; Lee, Y.-M.; Kubo, M.; Ogura, T.; Shaik, S.; Nam, W. Water as an Oxygen Source: Synthesis, Characterization, and Reactivity Studies of a Mononuclear Nonheme Manganese(IV) Oxo Complex. Angew. Chem. Int. Ed. 2010, 49, 8190–9194. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.M. Hydrogen Atom Abstraction by Metal-Oxo Complexes: Understanding the Analogy with Organic Radical Reactions. Acc. Chem. Res. 1998, 31, 441–450. [Google Scholar] [CrossRef]
- Bordwell, F.G.; Cheng, J.P.; Harrelson, J.A. Homolytic bond dissociation energies in solution from equilibrium acidity and electrochemical data. J. Am. Chem. Soc. 1988, 110, 1229–1231. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).