
Lanthanide Exposure In Vitro Differentially Diminishes MTT Cell Viability in Axenic Neuronal or Glial Cell Model Systems

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Saline/Sodium Nitrate/Calcium Nitrate
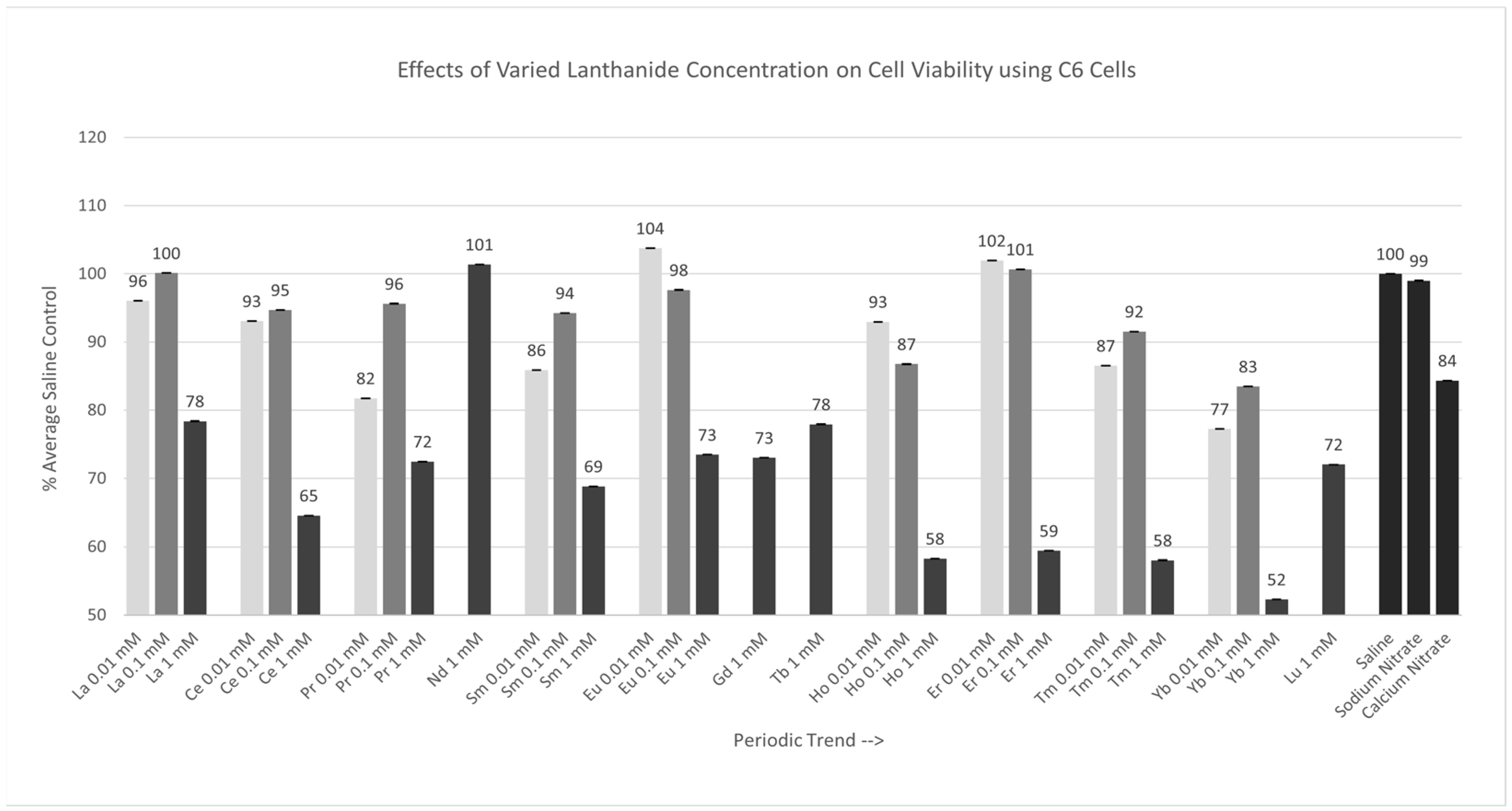
2.2. Concentration Variation Experiments
2.2.1. C6 Cells [0.01 mM, 0.1 mM, 1 mM]
2.2.2. PC12 Cells [0.01 mM, 0.1 mM, 1 mM]
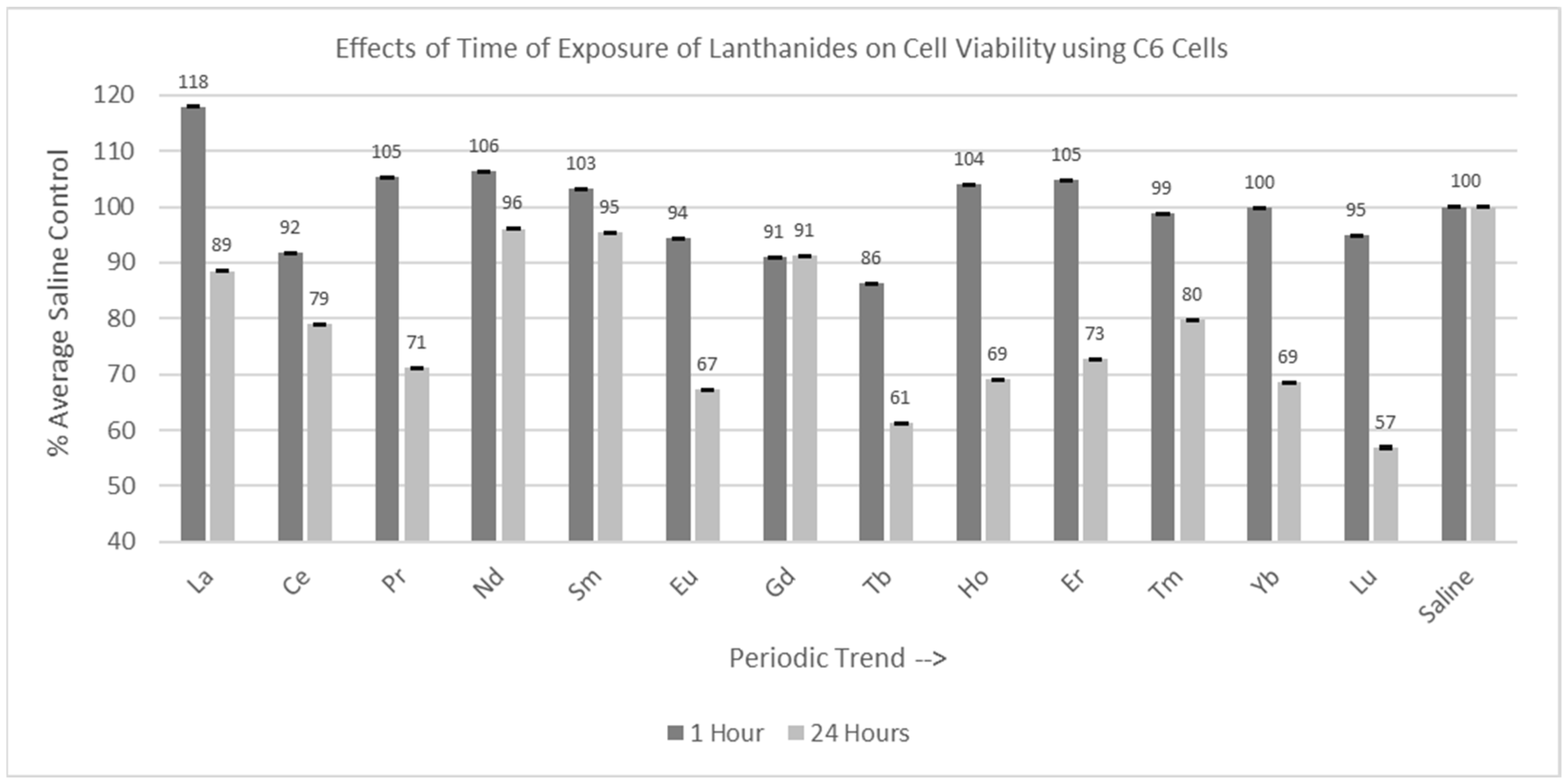
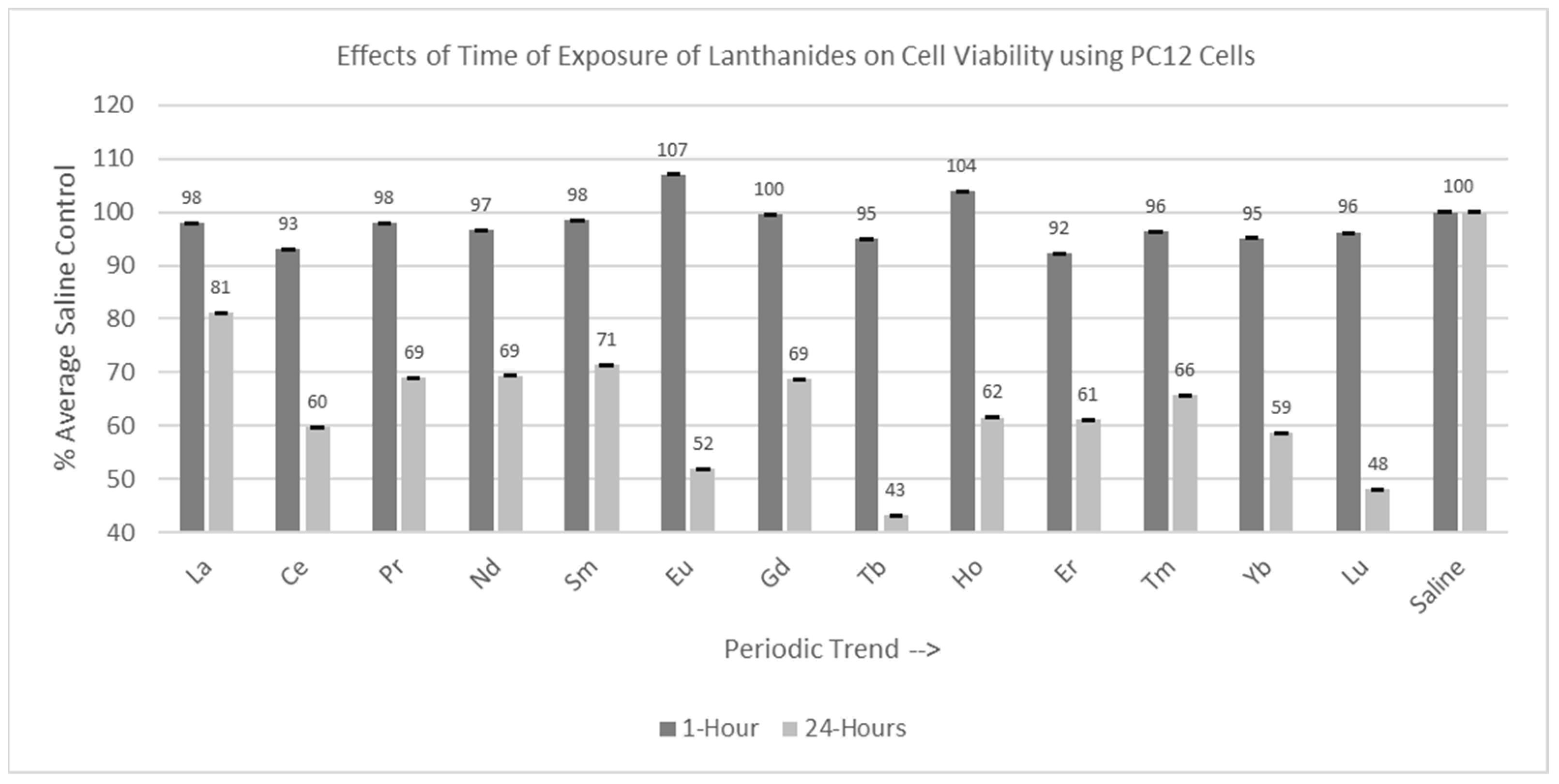
2.3. Incubation Time Experiments
2.3.1. C6 Cells (1 hr and 24 hr Incubation)
2.3.2. PC12 Cells (1 hr and 24 hr Incubation)
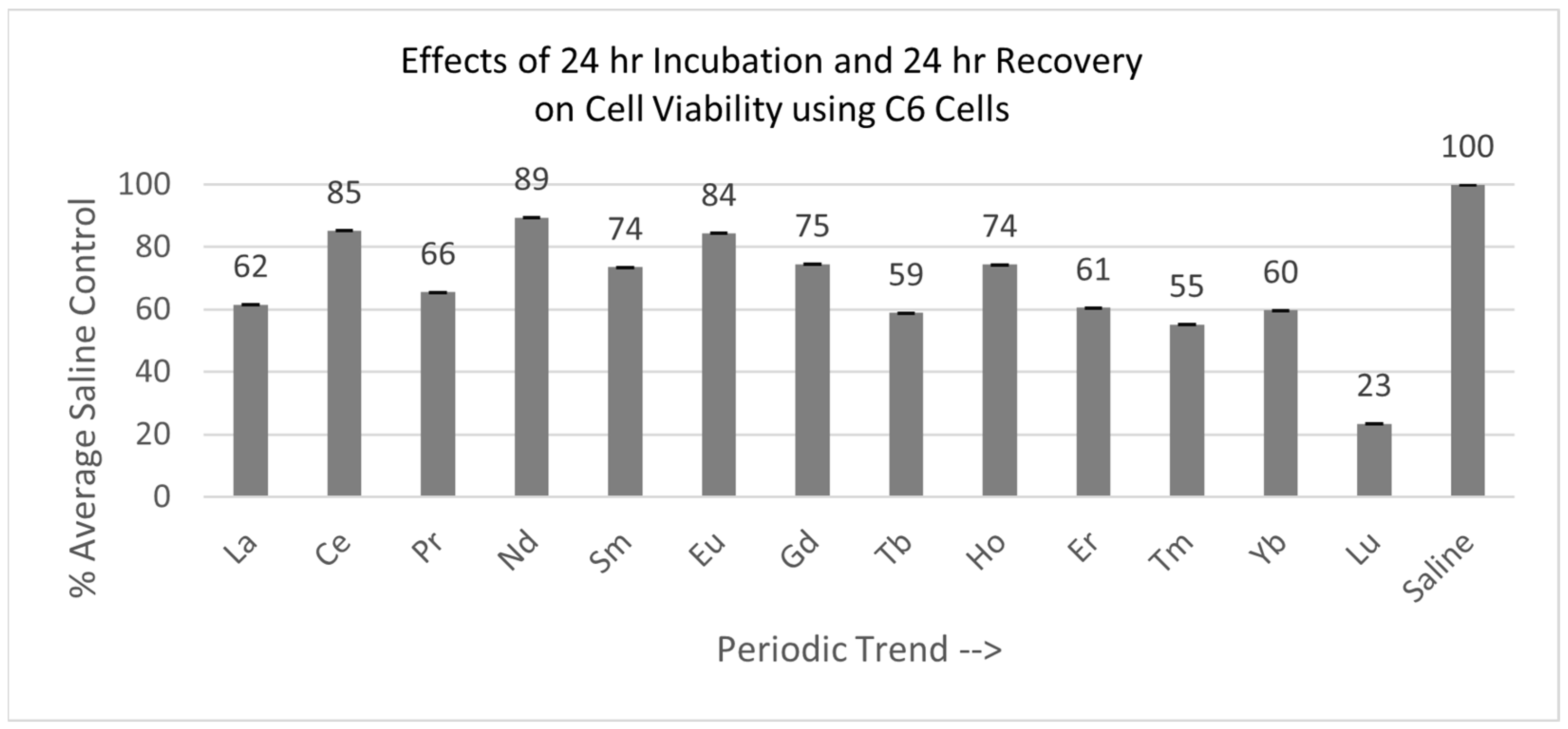
2.4. Recovery After Lanthanide Removal Experiments
2.4.1. C6 Cells (24 hr Incubation with 24 hr Recovery)
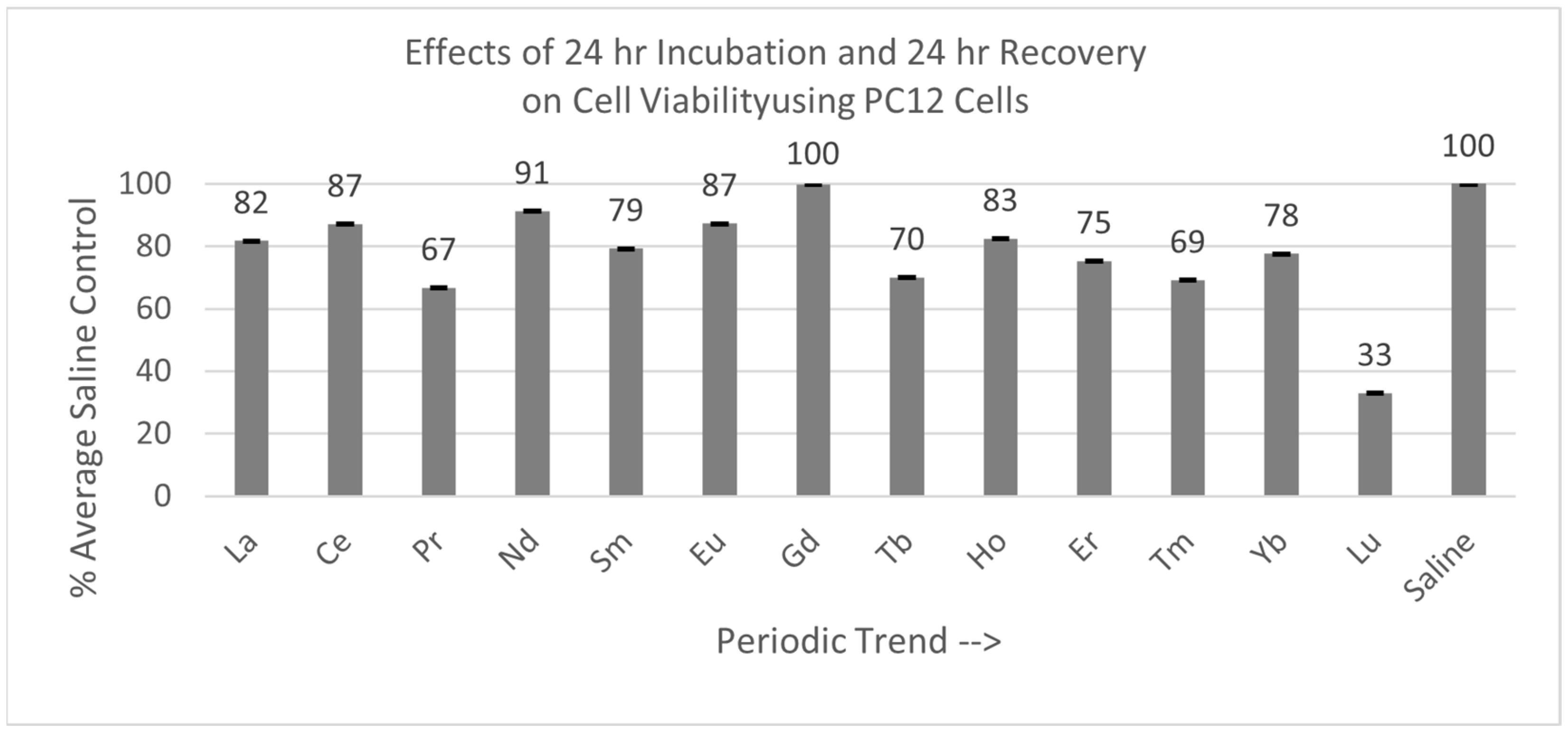
2.4.2. PC12 Cells (24 hr Incubation with 24 hr Recovery)
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Viability Assays
4.3. Stock Lanthanide Preparation
5. Conclusions
Limitations of the Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGill, I. Rare Earth Elements. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH: Weinheim, Germany, 2005. [Google Scholar] [CrossRef]
- Aspinall, H.C. Chemistry of the f-Block Elements; CRC Press: Boca Raton, FL, USA, 2001; p. 8. ISBN 978-90-5699-333-7. [Google Scholar]
- Haxel, G.; Hedrick, J.; Orris, J. Rare Earth Elements Critical Resources for High Technology (PDF); United States Geological Survey: Reston, VA, USA, 2006; USGS Fact Sheet: 087-02. Archived (PDF) from the original on 14 December 2010. Retrieved 19 April 2008. [Google Scholar]
- Palasz, A.; Czeka, P. Toxicological and cytophysiological aspects of lanthanides action. Acta Biochim Pol. 2000, 47, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.; Zhao, Q.; Su, D.; Li, P.; Stagnitti, F. Biological toxicity of lanthanide elements on algae. Chemosphere 2010, 80, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, V.; Vignati, D.A.; Leyval, C.; Giamberini, L. Environmental fate and ecotoxicity of lanthanides: Are they a uniform group beyond chemistry? Environ. Int. 2014, 71, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Pol, A.; Barends, T.R.M.; Dietl, A.; Khadem, A.F.; Eygensteyn, J.; Jetten, M.S.M.; Op den Camp, H.J.M. Rare earth metals are essential for methanotrophic life in volcanic mudpots. Environ. Microbiol. 2014, 16, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Good, N.M.; Lee, H.D.; Hawker, E.R.; Su, M.Z.; Gilad, A.A.; Martinez-Gomez, N.C. Hyperaccumulation of gadolinium by Methylorubrum extorquens AM1 reveals impacts of lanthanides on cellular processes beyond methylotrophy. Front. Microbiol. 2022, 13, 820327. [Google Scholar] [CrossRef] [PubMed]
- Peplow, M. Unlocking the lanthanome. Chemical and Engineering News, 7 February 2022. [Google Scholar]
- Cotruvo, J.A. The chemistry of lanthanides in biology: recent discoveries, emerging principles, and technological applications. ACS Cent. Sci. 2019, 5, 1496–1506. [Google Scholar] [CrossRef] [PubMed]
- Featherson, E.R.; Cortruvo, J.A., Jr. The biochemistry of lanthanide acquisition, trafficking, and utilization. BBA-Mol. Cell Res. 2021, 1868, 118864. [Google Scholar]
- Ramirez-Olvera, S.; Trejo-Tellez, L.I.; Garcia-Morales, S.; Perez-Sate, J.A.; Gomez-Merino, F.C. Cerium enhances germination and shoot growth and alters mineral nutrient concentration in rice. PLoS ONE 2018, 13, e0194691. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Gao, N.; Zhang, S.; Xing, J.; Hou, J. Investigating the toxically homogenous effects of three lanthanides on zebrafish. Comp. Biochem. Physiol. Part C 2022, 253, 109251. [Google Scholar] [CrossRef] [PubMed]
- Edington, S.C.; Gonzalez, A.; Middendort, T.R.; Halling, D.B.; Aldrich, R.W.; Baiz, C.R. Coordination of lanthanide ions distorts binding site conformation in calmodulin. Proc. Natl. Acad. Sci. USA 2018, 115, E3126–E3134. [Google Scholar] [CrossRef] [PubMed]
- Vikolova, V.; Kircheva, N.; Dobrev, S.; Angelova, S.; Dudev, T. Lanthanides as calcium mimetic species in calcium-signaling/buffering proteins: The effect of lanthanide type on the Ca2+/Ln3+ competition. Int. J. Mol. Sci. 2023, 24, 6297. [Google Scholar] [CrossRef] [PubMed]
- Pallares, R.M.; Faulkner, D.; An, D.D.; Hebert, S.; Loguinov, A.; Proctor, M.; Villalobos, J.A.; Bjornstad, K.A.; Rosen, C.J.; Vulpe, C.; et al. Genome-wide toxicologenomic study of the lanthanides sheds light on the selective toxicity mechanisms associated with critical materials. Proc. Natl. Acad. Sci. USA 2021, 118, e2025952118. [Google Scholar] [CrossRef] [PubMed]
- Pallares, R.M.; An, D.D.; Hebert, S.; Loguinov, A.; Proctor, M.; Villalobos, J.A.; Bjornstad, K.A.; Rosen, C.J.; Vulpe, C.; Abergel, R.J. Identifying toxicity mechanisms associated with early lanthanide exposure through multidimensional genome-wide screening. ACS Omega 2022, 7, 34412–34419. [Google Scholar] [CrossRef] [PubMed]
- Pallares, R.M.; Li, Y.; Abergel, R.J. Understanding the biological behavior of lanthanides and actinides through omics approaches. Trends Anal. Chem. 2023, 67, 117251. [Google Scholar] [CrossRef]
- Pallares, R.M.; An, D.D.; Hebert, S.; Loguinov, A.; Proctor, M.; Villalobos, J.A.; Bjornstad, K.A.; Rosen, C.J.; Vulpe, C.; Abergel, R.J. Screening the complex biological behavior of late lanthanides through genome-wide interactions. Metallomics 2023, 15, mfad039. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, K.J.; Han, X.; Müller, U.F. A ribozyme that uses lanthanides as cofactor. Nucleic Acids Res. 2023, 51, 7163–7173. [Google Scholar] [CrossRef] [PubMed]
- Patasz, A.; Segovia, Y.; Skowronek, R.; Worthington, J.J. Molecular neurochemistry of the lanthanides. Synapse 2019. [Google Scholar] [CrossRef]
- Platt, D.C.; Apuzzo, C.F.; Jones, M.A.; Cedeño, D.L.; Vallejo, R. Development of a C6 glioma cell model system to assess effects of cathodic passively balanced electrical stimulation on responses to neurotransmitters: Implications for modulation of intracellular nitric oxide, chloride, and calcium ions. Brain Sci. 2022, 12, 1504. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Platt, D.C.; Ferrence, L.M.; Breausche, F.; Terry, K.; Ferrence, G.M.; Jones, M.A. Lanthanide Exposure In Vitro Differentially Diminishes MTT Cell Viability in Axenic Neuronal or Glial Cell Model Systems. Inorganics 2025, 13, 127. https://doi.org/10.3390/inorganics13040127
Platt DC, Ferrence LM, Breausche F, Terry K, Ferrence GM, Jones MA. Lanthanide Exposure In Vitro Differentially Diminishes MTT Cell Viability in Axenic Neuronal or Glial Cell Model Systems. Inorganics. 2025; 13(4):127. https://doi.org/10.3390/inorganics13040127
Chicago/Turabian StylePlatt, David C., Linda M. Ferrence, Faith Breausche, Katelyn Terry, Gregory M. Ferrence, and Marjorie A. Jones. 2025. "Lanthanide Exposure In Vitro Differentially Diminishes MTT Cell Viability in Axenic Neuronal or Glial Cell Model Systems" Inorganics 13, no. 4: 127. https://doi.org/10.3390/inorganics13040127
APA StylePlatt, D. C., Ferrence, L. M., Breausche, F., Terry, K., Ferrence, G. M., & Jones, M. A. (2025). Lanthanide Exposure In Vitro Differentially Diminishes MTT Cell Viability in Axenic Neuronal or Glial Cell Model Systems. Inorganics, 13(4), 127. https://doi.org/10.3390/inorganics13040127