
Network Pharmacology Study to Interpret Signaling Pathways of Ilex cornuta Leaves against Obesity




Abstract
:1. Introduction
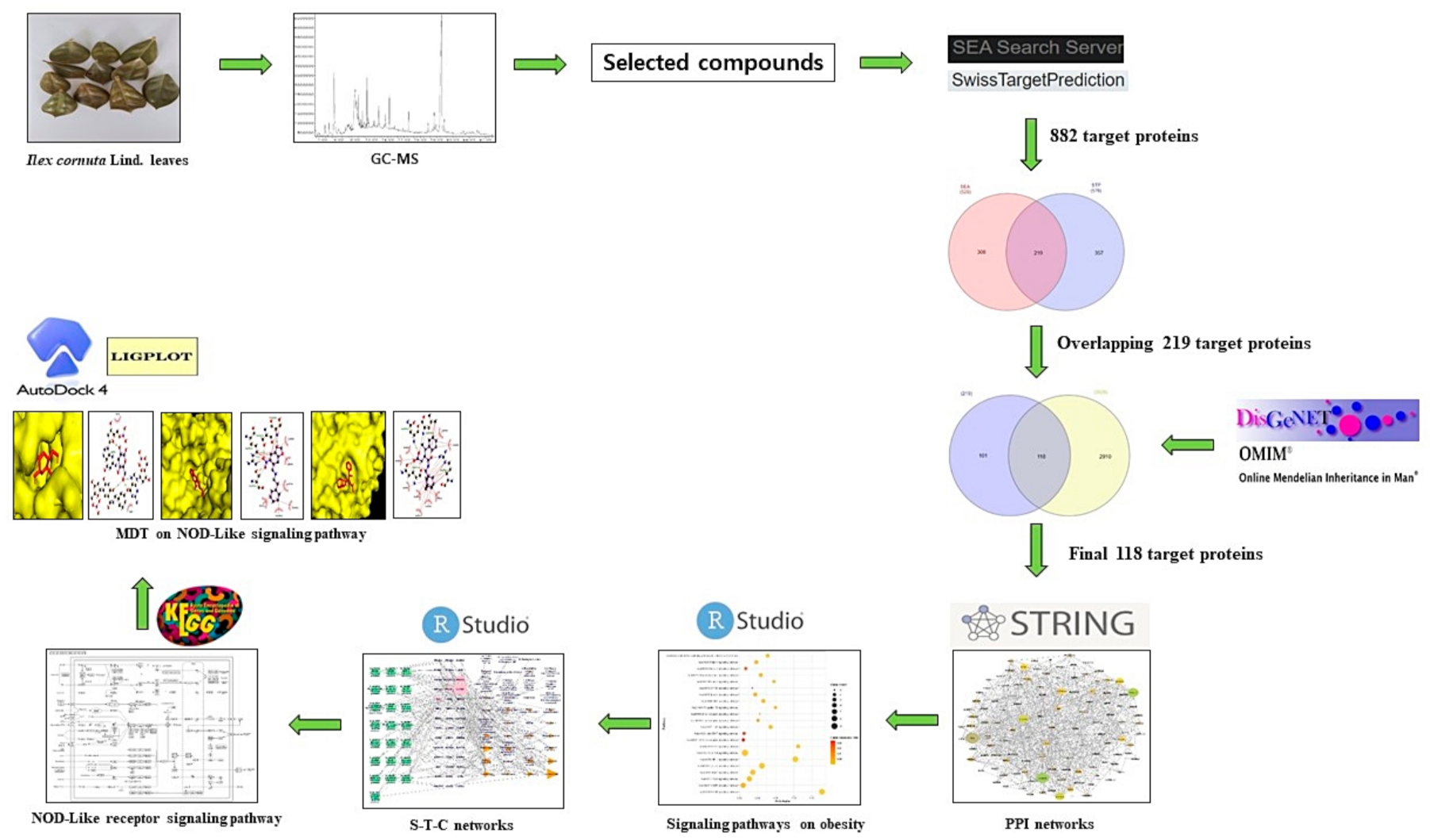
2. Results
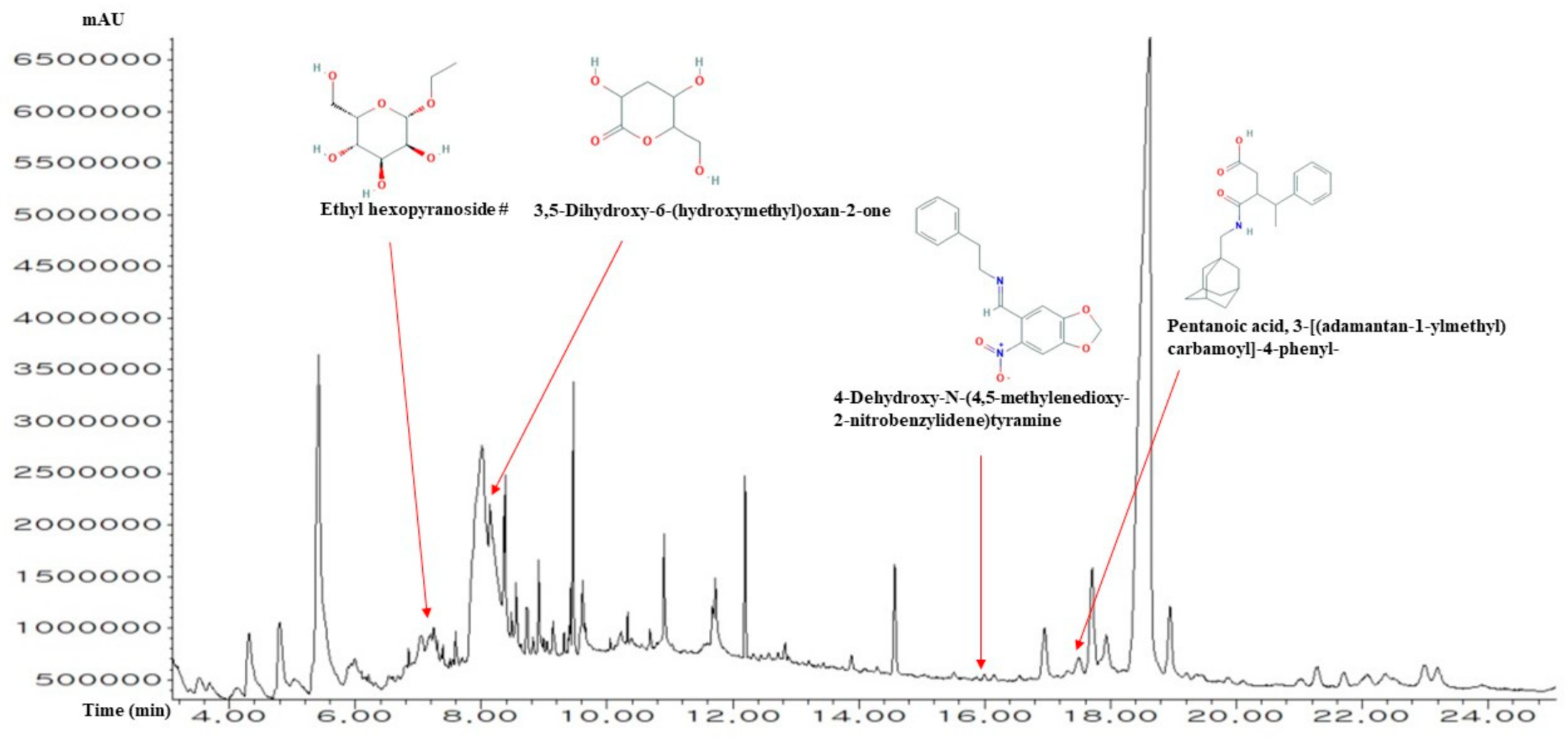
2.1. Chemical Compounds from ICLs
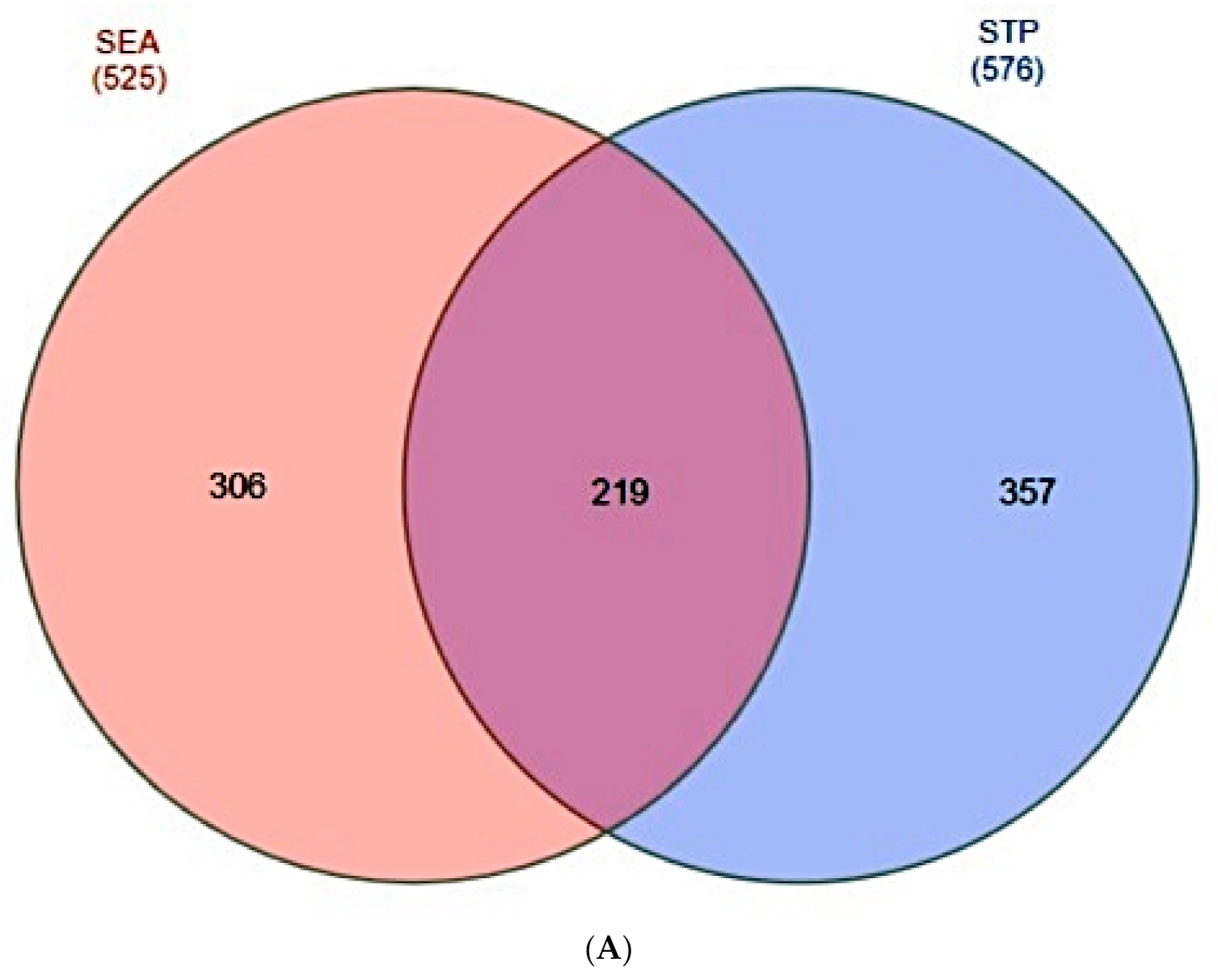
2.2. Overlapping Target Proteins between SEA and STP
2.3. Overlapping Target Proteins between Obesity-Related Target Proteins and 219 Target Proteins
2.4. PPI Networks
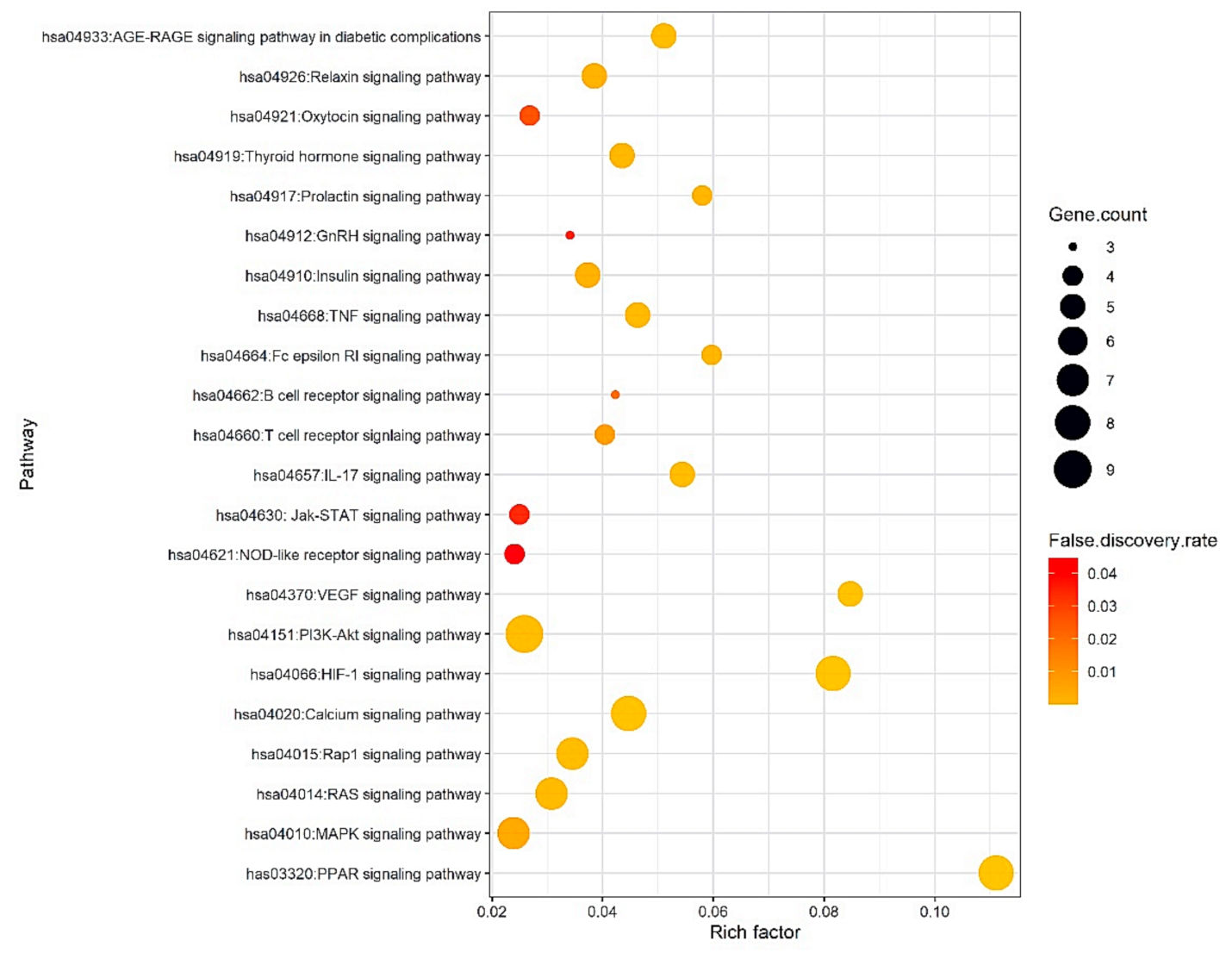
2.5. Analysis of Signaling Pathways against Obesity
2.6. STC Networks Analysis of ICLs against Obesity
2.7. KEGG Pathway Analysis of NLR Signaling Pathway
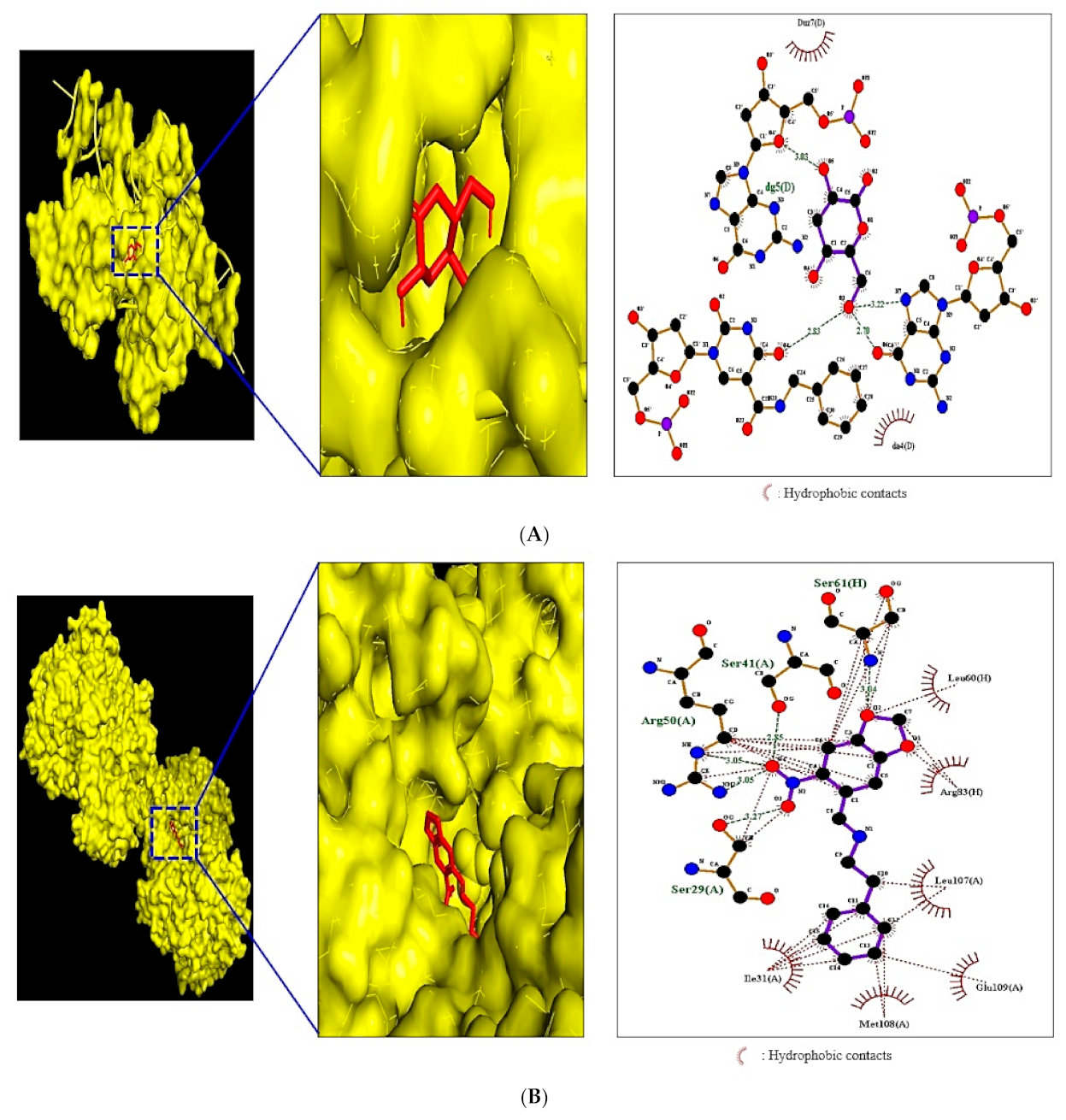
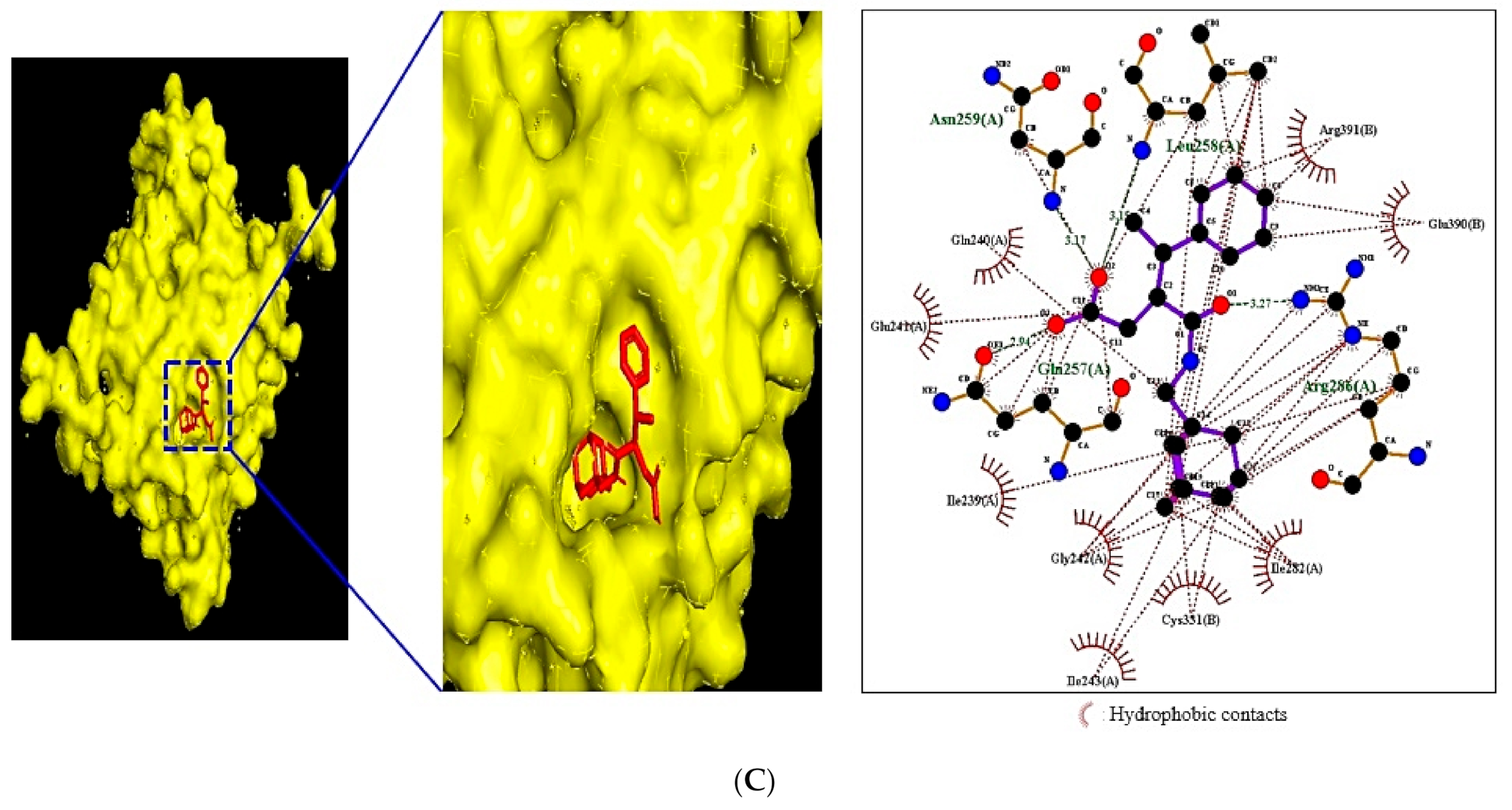
2.8. MDT of Four Target Proteins and 11 Compounds Connected to NLR Signaling Pathway
2.9. Toxicological Properties of Four Key Compounds
3. Discussion
4. Materials and Methods
4.1. Plant Material Collection and Classification
4.2. Plant Preparation and Extraction
4.3. GC-MS Analysis Condition
4.4. Chemical Compounds Database Construction and Drug-Likeness Identification
4.5. Acquisition of Target Proteins Related to Selected Chemical Compounds or Obesity
4.6. PPI Networks and Signaling Pathways on a Bubble Chart
4.7. A Signaling Pathways-Target Proteins-Chemical Compounds Network
4.8. Preparation of Target Proteins for MDT
4.9. Preparation of Ligands for MDT
4.10. Preparation of Positive Standard Ligands on the NLR Signaling Pathway for MDT
4.11. Preparation of Ligand Molecules for MDT
4.12. Ligand-Protein Docking
4.13. Toxicological Properties Prediction by admetSAR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| GnRH | Gonadotropin- Releasing Hormone |
| ICLs | Ilex cornuta leaves |
| IL17 | Interleukin 17 |
| Janus Kinase | Signal Transducer and Activator of Transcription proteins: JAK-STAT |
| MAPK1 | Mitogen-activated protein kinase 1 |
| MDT | Molecular Docking Test |
| NLR | NOD-like receptor |
| OTPs | Overlapping Target Proteins |
| ORTPs | Obesity-Related Target Proteins |
| PhosphoInositide | 3-Kinase—Protein kinase B: PI3K-AKT |
| PPI | Protein–Protein Interaction |
| RAP1 | Ras-proximate-1 |
| SEA | Similarity Ensemble Approach |
| SMILES | Simplified Molecular Input Line Entry System |
| STC | Signaling pathways -Target proteins-Compounds |
| STP | SwissTargetPrediction |
| TNF | Tumor Necrosis Factor |
References
- Hurt, R.T.; Kulisek, C.; Buchanan, L.A.; McClave, S.A. The obesity epidemic: Challenges, health initiatives, and implications for gastroenterologists. Gastroenterol. Hepatol. 2010, 6, 780–792. [Google Scholar]
- Hryhorczuk, C.; Sharma, S.; Fulton, S.E. Metabolic disturbances connecting obesity and depression. Front. Neurosci. 2013, 7, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, K.K.; Adnan, M.; Ju, I.; Cho, D.H. A network pharmacology study on main chemical compounds fromHibiscus cannabinusL. leaves. RSC Adv. 2021, 11, 11062–11082. [Google Scholar] [CrossRef]
- Artham, S.M.; Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and hypertension, heart failure, and coronary heart disease—Risk factor, paradox, and recommendations for weight loss. Ochsner J. 2009, 9, 124–132. [Google Scholar]
- Shi, J.; Fan, J.; Su, Q.; Yang, Z. Cytokines and Abnormal Glucose and Lipid Metabolism. Front. Endocrinol. 2019, 10, 703. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Tschöp, M.H.; Wilding, J.P.H. Anti-obesity drugs: Past, present and future. DMM Dis. Models Mech. 2012, 5, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.G.; Park, C.Y. Anti-obesity drugs: A review about their effects and safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Chy, M.N.U.; Adnan, M.; Chowdhury, M.R.; Pagano, E.; Kamal, A.T.M.M.; Oh, K.K.; Cho, D.H.; Capasso, R. Central and peripheral pain intervention by Ophiorrhiza rugosa leaves: Potential underlying mechanisms and insight into the role of pain modulators. J. Ethnopharmacol. 2021, 276, 114182. [Google Scholar]
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef]
- KR101876617B1—Composition Comprising Extract of Ilex Cornuta for Promoting the Differentiation to Adipocytic Cells—Google Patents. Available online: https://patents.google.com/patent/KR101876617B1/en (accessed on 26 April 2021).
- Kim, J.; Kang, W.; Min, H. In Vitro Anti-Inflammatory Activity of Ilex cornuta Extract Mediated by Inhibition of Extracellular Signal-Regulated Kinase Phosphorylation. J. Med. Food 2017, 20, 981–988. [Google Scholar] [CrossRef]
- Ahn, E.M.; Asamenew, G.; Kim, H.W.; Lee, S.H.; Yoo, S.M.; Cho, S.M.; Cha, Y.S.; Kang, M.S. Anti-obesity effects of petasites japonicus (Meowi) ethanol extract on raw 264.7 macrophages and 3t3-l1 adipocytes and its characterization of polyphenolic compounds. Nutrients 2020, 12, 1261. [Google Scholar] [CrossRef]
- Hao, D.; Gu, X.; Xiao, P.; Liang, Z.; Xu, L.; Peng, Y. Research progress in the phytochemistry and biology of Ilex pharmaceutical resources. Acta Pharm. Sin. B 2013, 3, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.K.; Adnan, M.; Cho, D.H. Network pharmacology approach to decipher signaling pathways associated with target proteins of NSAIDs against COVID-19. Sci. Rep. 2021, 11, 9606. [Google Scholar] [CrossRef]
- Chandran, U.; Mehendale, N.; Patil, S.; Chaguturu, R.; Patwardhan, B. Network Pharmacology. In Innovative Approaches in Drug Discovery: Ethnopharmacology, Systems Biology and Holistic Targeting; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 127–164. ISBN 9780128018224. [Google Scholar]
- Oh, K.K.; Adnan, M.; Cho, D.H. Network pharmacology of bioactives from Sorghum bicolor with targets related to diabetes mellitus. PLoS ONE 2020, 15, e0240873. [Google Scholar] [CrossRef]
- Oh, K.K.; Adnan, M.; Cho, D.H. Active ingredients and mechanisms of Phellinus linteus (grown on Rosa multiflora) for alleviation of Type 2 diabetes mellitus through network pharmacology. Gene 2020, 768, 145320. [Google Scholar] [CrossRef]
- Shen, L.; Chen, W.; Zhang, B.; Liu, L.; Cao, Y. Integrating network pharmacology and bioinformatics analysis to explore the mechanism of Yupingfengsan in treating lung adenocarcinoma. Eur. J. Integr. Med. 2019, 31, 100967. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, L.; Yin, Z.; Lin, J. Improving chemical similarity ensemble approach in target prediction. J. Cheminformatics 2016, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W3664. [Google Scholar] [CrossRef] [Green Version]
- Piñero, J.; Queralt-Rosinach, N.; Bravo, À.; Deu-Pons, J.; Bauer-Mehren, A.; Baron, M.; Sanz, F.; Furlong, L.I. DisGeNET: A discovery platform for the dynamical exploration of human diseases and their genes. Database 2015, 2015, 1–17. [Google Scholar] [CrossRef]
- Hamosh, A.; Scott, A.F.; Amberger, J.S.; Bocchini, C.A.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM), a knowledgebase of human genes and genetic disorders. Nucleic Acids Res. 2005, 33, D514. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, P.; He, W.; Qin, C.; Chen, S.; Tao, L.; Wang, Y.; Tan, Y.; Gao, D.; Wang, B.; et al. NPASS: Natural product activity and species source database for natural product research, discovery and tool development. Nucleic Acids Res. 2018, 46, D1217–D1222. [Google Scholar] [CrossRef] [Green Version]
- Hilgren, J.D.; Salverda, J.A. Antimicrobial Efficacy of a Peroxyacetic/Octanoic Acid Mixture in Fresh-Cut-Vegetable Process Waters. J. Food Sci. 2000, 65, 1376–1379. [Google Scholar] [CrossRef]
- Saleem, M. Lupeol, a novel anti-inflammatory and anti-cancer dietary triterpene. Cancer Lett. 2009, 285, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Sun, H.; Liu, Y.; Shiroshita, T.; Kawano, S.; Takeshi, S.; Ma, J.; Zhang, Z. Anti-cancer activity comparisons of aqueous extracts from Inonotus obliquus, Cordyceps militaris and Uncaria tomentosa in vitro and in vivo. J. Pharmacogn. Phytochem. 2014, 2, 19–25. [Google Scholar]
- Wang, Q.B.; Sun, L.Y.; Du, Y. Veratric Acid Inhibits LPS-Induced IL-6 and IL-8 Production in Human Gingival Fibroblasts. Inflammation 2016, 39, 237–242. [Google Scholar] [CrossRef]
- Tamai, R.; Suzuki, K.; Mashima, I.; Kiyoura, Y. MPMBP down-regulates Toll-like receptor (TLR) 2 ligand-induced proinflammatory cytokine production by inhibiting NF-κB but not AP-1 activation. Int. Immunopharmacol. 2020, 79. [Google Scholar] [CrossRef]
- Shityakov, S.; Förster, C. In silico predictive model to determine vector-mediated transport properties for the blood-brain barrier choline transporter. Adv. Appl. Bioinform. Chem. 2014. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Yang, Q.; He, Y.; Zou, X.; Cao, Z. BmK NT1-induced neurotoxicity is mediated by PKC/CaMKⅡ-dependent ERK1/2 and p38 activation in primary cultured cerebellar granule cells. Toxicology 2019, 421, 22–29. [Google Scholar] [CrossRef]
- Xu, S.; Li, X.; Liu, Y.; Xia, Y.; Chang, R.; Zhang, C. Inflammasome inhibitors: Promising therapeutic approaches against cancer. J. Hematol. Oncol. 2019, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.-C.; Ho, H.-H.; Lin, M.-C.; Huang, C.-N.; Huang, H.-P.; Wang, C.-J. Impact of polyphenolic components from mulberry on apoptosis of vascular smooth muscle cells. J. Sci. Food Agric. 2016, 96, 381–391. [Google Scholar] [CrossRef]
- Renolleau, S.; Fau, S.; Goyenvalle, C.; Joly, L.M.; Chauvier, D.; Jacotot, E.; Mariani, J.; Charriaut-Marlangue, C. Specific caspase inhibitor Q-VD-OPh prevents neonatal stroke in P7 rat: A role for gender. J. Neurochem. 2007, 100, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Aldeghi, M.; Malhotra, S.; Selwood, D.L.; Chan, A.W.E. Two- and three-dimensional rings in drugs. Chem. Biol. Drug Des. 2014, 83, 450–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, H.; Tian, L.; Li, Q.; Luo, J.; Zhang, Y. Analysis of the Physicochemical Properties of Acaricides Based on Lipinski’s Rule of Five. J. Comput. Biol. 2020, 27, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Shrivastava, S.; Hassanali, M.; Stothard, P.; Chang, Z.; Woolsey, J. DrugBank: A comprehensive resource for in silico drug discovery and exploration. Nucleic Acids Res. 2006, 34. [Google Scholar] [CrossRef]
- Blaschke, F.; Takata, Y.; Caglayan, E.; Law, R.E.; Hsueh, W.A. Obesity, peroxisome proliferator-activated receptor, and atherosclerosis in type 2 diabetes. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Elias, I.; Franckhauser, S.; Bosch, F. New insights into adipose tissue VEGF-A actions in the control of obesity and insulin resistance. Adipocyte 2013, 2, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Qu, A.; Matsubara, T.; Chanturiya, T.; Jou, W.; Gavrilova, O.; Shah, Y.M.; Gonzalez, F.J. Disruption of hypoxia-inducible factor 1 in adipocytes improves insulin sensitivity and decreases adiposity in high-fat diet-fed mice. Diabetes 2011, 60, 2484–2495. [Google Scholar] [CrossRef] [Green Version]
- Nadler, S.T.; Stoehr, J.P.; Schueler, K.L.; Tanimoto, G.; Yandell, B.S.; Attie, A.D. The expression of adipogenic genes is decreased in obesity and diabetes mellitus. Proc. Natl. Acad. Sci. USA 2000, 97, 11371–11376. [Google Scholar] [CrossRef] [Green Version]
- Kok, P.; Roelfsema, F.; Frölich, M.; Meinders, A.E.; Pijl, H. Prolactin Release Is Enhanced in Proportion to Excess Visceral Fat in Obese Women. J. Clin. Endocrinol. Metab. 2004, 89, 4445–4449. [Google Scholar] [CrossRef]
- Ahmed, M.; Gaffen, S.L. IL-17 in obesity and adipogenesis. Cytokine Growth Factor Rev. 2010, 21, 449–453. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga, L.A.; Shen, W.-J.; Joyce-Shaikh, B.; Pyatnova, E.A.; Richards, A.G.; Thom, C.; Andrade, S.M.; Cua, D.J.; Kraemer, F.B.; Butcher, E.C. IL-17 Regulates Adipogenesis, Glucose Homeostasis, and Obesity. J. Immunol. 2010, 185, 6947–6959. [Google Scholar] [CrossRef] [Green Version]
- Tomino, Y.; Hagiwara, S.; Gohda, T. AGE-RAGE interaction and oxidative stress in obesity-related renal dysfunction. Kidney International 2011, 80, 133–135. [Google Scholar] [CrossRef] [Green Version]
- Leuner, B.; Max, M.; Thamm, K.; Kausler, C.; Yakobus, Y.; Bierhaus, A.; Sel, S.; Hofmann, B.; Silber, R.E.; Simm, A.; et al. RAGE beeinflusst adipositas bei mäusen. einfluss des rezeptors für AGEs (RAGE) auf gewichtszunahme, insulin-und AGE-akkumulation in mäusen unter hochfettdiät. Zeitschrift für Gerontologie und Geriatrie 2012, 45, 102–108. [Google Scholar] [CrossRef]
- Kern, P.A.; Ranganathan, S.; Li, C.; Wood, L.; Ranganathan, G. Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2001, 280, 745–751. [Google Scholar] [CrossRef]
- Song, Z.; Wang, Y.; Zhang, F.; Yao, F.; Sun, C. Calcium signaling pathways: Key pathways in the regulation of obesity. Int. J. Mol. Sci. 2019, 20, 2768. [Google Scholar] [CrossRef] [Green Version]
- Song, R.H.; Wang, B.; Yao, Q.M.; Li, Q.; Jia, X.; Zhang, J.A. The Impact of Obesity on Thyroid Autoimmunity and Dysfunction: A Systematic Review and Meta-Analysis. Front. Immunol. 2019, 10, 2349. [Google Scholar] [CrossRef]
- Liu, R.; Nikolajczyk, B.S. Tissue immune cells fuel obesity-associated inflammation in adipose tissue and beyond. Front. Immunol. 2019, 10, 1587. [Google Scholar] [CrossRef] [Green Version]
- Shirakawa, K.; Yan, X.; Shinmura, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Yamamoto, T.; Anzai, A.; Isobe, S.; Yoshida, N.; et al. Obesity accelerates T cell senescence in murine visceral adipose tissue. J. Clin. Investig. 2016, 126, 4626–4639. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, E.G.; Murphy, W.J. Obesity induced T cell dysfunction and implications for cancer immunotherapy. Curr. Opin. Immunol. 2018, 51, 181–186. [Google Scholar] [CrossRef]
- Lee, K.C.; Hsieh, Y.C.; Chan, C.C.; Sun, H.J.; Huang, Y.H.; Hou, M.C.; Lin, H.C. Human relaxin-2 attenuates hepatic steatosis and fibrosis in mice with non-alcoholic fatty liver disease. Lab. Investig. 2019, 99, 1203–1216. [Google Scholar] [CrossRef]
- Smith, C.M.; Walker, A.W.; Hosken, I.T.; Chua, B.E.; Zhang, C.; Haidar, M.; Gundlach, A.L. Relaxin-3/RXFP3 networks: An emerging target for the treatment of depression and other neuropsychiatric diseases? Front. Pharmacol. 2014, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef]
- Martínez, P.; Gómez-López, G.; García, F.; Mercken, E.; Mitchell, S.; Flores, J.M.; deCabo, R.; Blasco, M.A. RAP1 Protects from Obesity through Its Extratelomeric Role Regulating Gene Expression. Cell Rep. 2013, 3, 2059–2074. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H.; Douchi, T.; Yamamoto, S.; Toshimichi, O.; Kuwahata, R.; Nagata, Y. Body fat distribution and body composition during GnRH agonist therapy. Obstet. Gynecol. 2001, 97, 338–342. [Google Scholar] [CrossRef]
- Goossens, G.H. The Renin-Angiotensin System in the Pathophysiology of Type 2 Diabetes. Obes. Facts 2012, 5, 611–624. [Google Scholar] [CrossRef]
- Ding, C.; Leow, M.K.S.; Magkos, F. Oxytocin in metabolic homeostasis: Implications for obesity and diabetes management. Obes. Rev. 2019, 20, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Liu, G.; Guo, J.; Su, Z.Q. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- Dodington, D.W.; Desai, H.R.; Woo, M. JAK/STAT—Emerging Players in Metabolism. Trends Endocrinol. Metab. 2018, 29, 55–65. [Google Scholar] [CrossRef]
- Hong, S.; Song, W.; Zushin, P.J.H.; Liu, B.; Jedrychowski, M.P.; Mina, A.I.; Deng, Z.; Cabarkapa, D.; Hall, J.A.; Palmer, C.J.; et al. Phosphorylation of Beta-3 adrenergic receptor at serine 247 by ERK MAP kinase drives lipolysis in obese adipocytes. Mol. Metab. 2018, 12, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Deng, T.; Peterson, L.E.; Yu, R.; Lin, J.; Hamilton, D.J.; Reardon, P.R.; Sherman, V.; Winnier, G.E.; Zhan, M.; et al. Transcriptome analysis of human adipocytes implicates the NOD-like receptor pathway in obesity-induced adipose inflammation. Mol. Cell. Endocrinol. 2014, 394, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Febbraio, M.A. Role of interleukins in obesity: Implications for metabolic disease. Trends Endocrinol. Metab. 2014, 25, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keiser, M.J.; Roth, B.L.; Armbruster, B.N.; Ernsberger, P.; Irwin, J.J.; Shoichet, B.K. Relating protein pharmacology by ligand chemistry. Nat. Biotechnol. 2007, 25, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Heberle, H.; Meirelles, V.G.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Khanal, P.; Patil, B.M.; Chand, J.; Naaz, Y. Anthraquinone Derivatives as an Immune Booster and their Therapeutic Option Against COVID-19. Nat. Prod. Bioprospect. 2020, 10, 325–335. [Google Scholar] [CrossRef]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. Ligplot: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. Des. Sel. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Yang, H.; Lou, C.; Sun, L.; Li, J.; Cai, Y.; Wang, Z.; Li, W.; Liu, G.; Tang, Y. admetSAR 2.0: Web-service for prediction and optimization of chemical ADMET properties. Bioinformatics 2019, 35, 1067–1069. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | Pubchem ID | RT (mins) | Area (%) | Pharmacological Activities (Reference) |
|---|---|---|---|---|---|
| 1 | N-Acetylmannosamine | 11,096,158 | 3.520 | 0.49 | No reports |
| 2 | 1-Aminopropan-2-ol | 4 | 3.683 | 0.41 | Anti-malaria [23] |
| 3 | 2-Propenethioamide, 3-(acetyloxy)-N,N-dimethyl-, (E)- | 5,363,184 | 4.135 | 0.46 | No reports |
| 4 | 2-Hydroxy-3-methyl-4H-pyran-4-one | 54,681,620 | 4.318 | 2.34 | No reports |
| 5 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | 119,838 | 4.799 | 1.98 | No reports |
| 6 | Tetrahydrofuran-3,4-diol | 90,803 | 5.039 | 0.84 | No reports |
| 7 | 5-Hydroxymethylfurfural | 237,332 | 5.424 | 9.54 | No reports |
| 8 | Ascaridole | 10,545 | 5.943 | 0.34 | Anti-neoplasms (PCIDB) |
| 9 | 1-Nonene | 31,285 | 6.000 | 0.30 | No reports |
| 10 | Fumaramic acid | 5,364,140 | 6.539 | 0.22 | No reports |
| 11 | 2,1,3-Benzothiadiazole | 67,502 | 6.808 | 0.43 | No reports |
| 12 | Hypoxanthine | 135,398,638 | 6.856 | 0.32 | Anti-gout (PCIDB) |
| 13 | Guanidine, 1-ethyl-3-nitro- | 135,515,028 | 7.049 | 1.79 | No reports |
| 14 | Levoglucosan | 2,724,705 | 7.202 | 1.18 | No reports |
| 15 | Ethyl-α-d-glucopyranoside | 91,694,274 | 7.250 | 0.82 | No reports |
| 16 | 2-(4-Methylcyclohexyl)prop-2-en-1-ol | 543,946 | 7.606 | 0.45 | No reports |
| 17 | Octanoic acid | 379 | 8.020 | 12.91 | Antimicrobial [24] |
| 18 | 3,5-Dihydroxy-6-(hydroxymethyl)oxan-2-one | 541,561 | 8.145 | 5.95 | No reports |
| 19 | 2-Isopropenylthiophene | 121,729 | 8.385 | 2.82 | No reports |
| 20 | Sulfallate | 7216 | 8.481 | 0.55 | No reports |
| 21 | Crocetane | 136,331 | 8.568 | 0.94 | No reports |
| 22 | Diphenylmethane | 7580 | 8.731 | 0.67 | No reports |
| 23 | Palmitic acid | 985 | 8.914 | 0.87 | Anti-cancer |
| 24 | 1-(3-Butyn-2-yloxy)-1-methyl-1-silacyclohexane | 597,458 | 9.154 | 0.47 | No reports |
| 25 | Lactose | 6134 | 9.318 | 0.20 | Thyroid cancer(marker) (PCIDB) |
| 26 | Methyl linoleate | 5,284,421 | 9.424 | 0.45 | No reports |
| 27 | Phytol | 5,366,244 | 9.462 | 1.28 | Anti-necrosis (PCIDB) |
| 28 | cis,cis,cis-7,10,13-Hexadecatrienal | 5,367,366 | 9.616 | 0.82 | No reports |
| 29 | Linolenic acid | 5,280,934 | 9.654 | 0.32 | Anti-inflammation [23] |
| 30 | Phytone | 10,408 | 10.058 | 0.06 | No reports |
| 31 | 11-Dodecynyl acetate | 538,082 | 10.222 | 0.43 | No reports |
| 32 | 1-(Pyrrolidinocarbonylmethyl)piperazine | 100,614 | 10.327 | 0.26 | No reports |
| 33 | Amonafide | 50,515 | 10.693 | 0.14 | No reports |
| 34 | 2-Palmitoylglycerol | 123,409 | 10.904 | 1.02 | No reports |
| 35 | 7-Pentadecyne | 549,063 | 11.683 | 0.49 | No reports |
| 36 | cis,cis,cis-7,10,13-Hexadecatrienal | 5,367,366 | 11.722 | 1.41 | No reports |
| 37 | Squalene | 638,072 | 12.202 | 1.24 | Anti-leukemia |
| 38 | 2-Methyl-3-(3-methyl-but-2-enyl)-2-(4-methyl-pent-3-enyl)-oxetane | 550,119 | 12.827 | 0.23 | No reports |
| 39 | Stigmastan-3-ol | 6743 | 13.895 | 0.24 | No reports |
| 40 | Vitamin E | 14,985 | 14.577 | 1.28 | Anti-oxidant [23] |
| 41 | 4-Cyclohexene-1,2-dicarboximide, N-butyl-, cis- | 91,733,922 | 15.520 | 0.10 | No reports |
| 42 | 4-Dehydroxy-N-(4,5-methylenedioxy-2-nitrobenzylidene)tyramine | 610,062 | 16.000 | 0.08 | No reports |
| 43 | Clionasterol | 457,801 | 16.962 | 1.14 | Anti-ischemic (PCIDB) |
| 44 | 17-(1,5-Dimethylhexyl)-10,13-dimethyl-4-vinylhexadecahydrocyclopenta[a]phenanthren-3-ol | 537,099 | 17.318 | 0.24 | No reports |
| 45 | Pentanoic acid, 3-[(adamantan-1-ylmethyl)carbamoyl]-4-phenyl- | 4,920,612 | 17.500 | 0.67 | No reports |
| 46 | β-amyrenol | 225,689 | 17.712 | 2.47 | No reports |
| 47 | Ethyl 2-(2-chloroacetamido)-3,3,3-trifluoro-2-(3-fluoroanilino)propionate | 610,054 | 17.933 | 1.71 | No reports |
| 48 | Lupeol | 259,846 | 18.625 | 31.17 | Anti-carcinoma [25] |
| 49 | Ethyl 2-[(2-chloroacetyl)amino]-3,3,3-trifluoro-2-(4-fluoroanilino)propanoate | 610,053 | 21.020 | 0.24 | No reports |
| 50 | Lanosterol | 246,983 | 21.289 | 0.53 | Anti-osteosarcoma [26] |
| 51 | Cycloisolongifolene, 7-bromo- | 608,988 | 21.712 | 0.37 | No reports |
| 52 | Cycloeucalenyl acetate | 537,081 | 23.202 | 0.64 | No reports |
| No. | Compounds | Lipinski Rules | Lipinski’s Violations | Bioavailability Score | TPSA (Å2) | |||
|---|---|---|---|---|---|---|---|---|
| MW | HBA | HBD | MLog P | |||||
| <500 | <10 | ≤5 | ≤4.15 | ≤1 | >0.1 | <140 | ||
| 1 | N-Acetylmannosamine | 221.21 | 6 | 5 | −2.61 | 0 | 0.55 | 119.25 |
| 2 | 1-Aminopropan-2-ol | 75.11 | 2 | 2 | −0.63 | 0 | 0.55 | 46.25 |
| 3 | 2-Propenethioamide, 3-(acetyloxy)-N,N-dimethyl-, (E)- | 173.23 | 2 | 0 | 0.54 | 0 | 0.55 | 61.63 |
| 4 | 2-Hydroxy-3-methyl-4H-pyran-4-one | 126.11 | 3 | 1 | −0.03 | 0 | 0.55 | 50.44 |
| 5 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | 144.13 | 4 | 2 | −1.77 | 0 | 0.85 | 66.76 |
| 6 | Tetrahydrofuran-3,4-diol | 104.10 | 3 | 2 | −1.45 | 0 | 0.55 | 49.69 |
| 7 | 5-Hydroxymethylfurfural | 126.11 | 3 | 1 | −1.06 | 0 | 0.55 | 50.44 |
| 8 | Ascaridole | 168.23 | 2 | 0 | 2.19 | 0 | 0.55 | 18.46 |
| 9 | 1-Nonene | 126.24 | 0 | 0 | 4.38 | 1 | 0.55 | 0.00 |
| 10 | Fumaramic acid | 115.09 | 3 | 2 | −1.04 | 0 | 0.56 | 80.39 |
| 11 | 2,1,3-Benzothiadiazole | 136.17 | 2 | 0 | 0.63 | 0 | 0.55 | 54.02 |
| 12 | Hypoxanthine | 136.11 | 3 | 2 | −1.17 | 0 | 0.55 | 74.43 |
| 13 | Guanidine, 1-ethyl-3-nitro- | 132.12 | 3 | 2 | −0.94 | 0 | 0.55 | 96.23 |
| 14 | Levoglucosan | 162.14 | 5 | 3 | −1.94 | 0 | 0.55 | 79.15 |
| 15 | Ethyl-α-d-glucopyranoside | 208.21 | 6 | 4 | −2.07 | 0 | 0.55 | 99.38 |
| 16 | 2-(4-Methylcyclohexyl)prop-2-en-1-ol | 154.25 | 1 | 1 | 2.30 | 0 | 0.55 | 20.23 |
| 17 | Octanoic acid | 144.21 | 2 | 1 | 1.96 | 0 | 0.85 | 37.30 |
| 18 | 3,5-Dihydroxy-6-(hydroxymethyl)oxan-2-one | 162.14 | 5 | 3 | −1.68 | 0 | 0.55 | 86.99 |
| 19 | 2-Isopropenylthiophene | 124.20 | 0 | 0 | 2.17 | 0 | 0.55 | 28.24 |
| 20 | Sulfallate | 223.79 | 0 | 0 | 2.28 | 0 | 0.55 | 60.63 |
| 21 | Crocetane | 282.55 | 0 | 0 | 7.38 | 1 | 0.55 | 0.00 |
| 22 | Diphenylmethane | 168.23 | 0 | 0 | 5.06 | 1 | 0.55 | 0.00 |
| 23 | Palmitic acid | 256.42 | 2 | 1 | 4.19 | 1 | 0.85 | 37.30 |
| 24 | 1-(3-Butyn-2-yloxy)-1-methyl-1-silacyclohexane | 182.33 | 1 | 0 | 2.30 | 0 | 0.55 | 9.23 |
| 25 | Lactose | 342.30 | 11 | 8 | −4.37 | 2 | 0.17 | 189.53 |
| 26 | Methyl linoleate | 294.47 | 2 | 0 | 4.70 | 1 | 0.55 | 26.30 |
| 27 | Phytol | 296.53 | 1 | 1 | 5.25 | 1 | 0.55 | 20.23 |
| 28 | cis,cis,cis-7,10,13-Hexadecatrienal | 234.38 | 1 | 0 | 4.01 | 0 | 0.55 | 17.07 |
| 29 | Linolenic acid | 278.43 | 2 | 1 | 4.38 | 1 | 0.85 | 37.30 |
| 30 | PHYTONE | 268.48 | 1 | 0 | 4.79 | 1 | 0.55 | 17.07 |
| 31 | 11-Dodecynyl acetate | 224.34 | 2 | 0 | 3.58 | 0 | 0.55 | 26.30 |
| 32 | 1-(Pyrrolidinocarbonylmethyl)piperazine | 197.28 | 3 | 1 | 0.05 | 0 | 0.55 | 35.58 |
| 33 | Amonafide | 283.33 | 3 | 1 | 1.71 | 0 | 0.55 | 68.33 |
| 34 | 2-Palmitoylglycerol | 330.50 | 4 | 2 | 3.18 | 0 | 0.55 | 66.76 |
| 35 | 7-Pentadecyne | 208.38 | 0 | 0 | 6.04 | 1 | 0.55 | 0.00 |
| 36 | cis,cis,cis-7,10,13-Hexadecatrienal | 234.38 | 1 | 0 | 4.01 | 0 | 0.55 | 17.07 |
| 37 | Squalene | 410.72 | 0 | 0 | 7.93 | 1 | 0.55 | 0.00 |
| 38 | 2-Methyl-3-(3-methyl-but-2-enyl)-2-(4-methyl-pent-3-enyl)-oxetane | 222.37 | 1 | 0 | 3.56 | 0 | 0.55 | 9.23 |
| 39 | Stigmastan-3-ol | 416.72 | 1 | 1 | 6.88 | 1 | 0.55 | 20.23 |
| 40 | Vitamin E | 430.71 | 2 | 1 | 6.14 | 1 | 0.55 | 29.46 |
| 41 | 4-Cyclohexene-1,2-dicarboximide, N-butyl-, cis- | 207.27 | 2 | 0 | 1.68 | 0 | 0.55 | 37.38 |
| 42 | 4-Dehydroxy-N-(4,5-methylenedioxy-2-nitrobenzylidene)tyramine | 298.29 | 5 | 0 | 1.49 | 0 | 0.55 | 76.64 |
| 43 | Clionasterol | 414.71 | 1 | 1 | 6.73 | 1 | 0.55 | 20.23 |
| 44 | 17-(1,5-Dimethylhexyl)-10,13-dimethyl-4-vinylhexadecahydrocyclopenta[a]phenanthren-3-ol | 414.71 | 1 | 1 | 6.73 | 1 | 0.55 | 20.23 |
| 45 | Pentanoic acid, 3-[(adamantan-1-ylmethyl)carbamoyl]-4-phenyl- | 369.50 | 3 | 2 | 3.69 | 0 | 0.85 | 66.40 |
| 46 | beta-Amyrenol | 426.72 | 1 | 1 | 6.92 | 1 | 0.55 | 20.23 |
| 47 | Ethyl 2-(2-chloroacetamido)-3,3,3-trifluoro-2-(3-fluoroanilino)propionate | 356.70 | 7 | 2 | 2.60 | 0 | 0.55 | 67.43 |
| 48 | Lupeol | 426.72 | 1 | 1 | 6.92 | 1 | 0.55 | 20.23 |
| 49 | Ethyl 2-[(2-chloroacetyl)amino]-3,3,3-trifluoro-2-(4-fluoroanilino)propanoate | 356.70 | 7 | 2 | 2.60 | 0 | 0.55 | 67.43 |
| 50 | Lanosterol | 426.72 | 1 | 1 | 6.82 | 1 | 0.55 | 20.23 |
| 51 | Cycloisolongifolene, 7-bromo- | 283.25 | 0 | 0 | 5.26 | 1 | 0.55 | 0.00 |
| 52 | Cycloeucalenyl acetate | 468.75 | 2 | 0 | 7.08 | 1 | 0.55 | 26.30 |
| No. | Gene Symbol | Degree | No. | Gene Symbol | Degree |
|---|---|---|---|---|---|
| 1 | IL6 | 52 | 59 | CES1 | 9 |
| 2 | VEGFA | 47 | 60 | FABP2 | 9 |
| 3 | PTGS2 | 42 | 61 | HK2 | 9 |
| 4 | CASP3 | 36 | 62 | HSD11B1 | 9 |
| 5 | MAPK1 | 36 | 63 | NR1I3 | 9 |
| 6 | PPARG | 30 | 64 | ADA | 8 |
| 7 | CYP3A4 | 29 | 65 | ALDH1A1 | 8 |
| 8 | ESR1 | 29 | 66 | HTR2B | 8 |
| 9 | NR3C1 | 25 | 67 | MAOB | 8 |
| 10 | MPO | 23 | 68 | PTPN6 | 8 |
| 11 | CNR1 | 21 | 69 | SLC6A2 | 8 |
| 12 | FGF2 | 21 | 70 | TBXA2R | 8 |
| 13 | IL2 | 21 | 71 | BCHE | 7 |
| 14 | AR | 20 | 72 | FFAR4 | 7 |
| 15 | PPARA | 20 | 73 | GPBAR1 | 7 |
| 16 | PTPRC | 20 | 74 | KISS1R | 7 |
| 17 | CYP2C9 | 19 | 75 | MGEA5 | 7 |
| 18 | HIF1A | 19 | 76 | SI | 7 |
| 19 | HNF4A | 19 | 77 | SLC5A1 | 7 |
| 20 | NR0B2 | 19 | 78 | CES2 | 6 |
| 21 | ABCB1 | 18 | 79 | FABP3 | 6 |
| 22 | CYP19A1 | 18 | 80 | FFAR1 | 6 |
| 23 | ALOX5 | 16 | 81 | GRIN1 | 6 |
| 24 | HK1 | 16 | 82 | MME | 6 |
| 25 | MGLL | 16 | 83 | NAAA | 6 |
| 26 | NOS2 | 16 | 84 | NR1H2 | 6 |
| 27 | PLA2G4A | 16 | 85 | OGT | 6 |
| 28 | PTGS1 | 16 | 86 | SRD5A1 | 6 |
| 29 | ACHE | 15 | 87 | CHRM2 | 5 |
| 30 | TRPV1 | 15 | 88 | FGF1 | 5 |
| 31 | G6PD | 15 | 89 | KAT2B | 5 |
| 32 | CASP1 | 14 | 90 | NPC1L1 | 5 |
| 33 | MMP3 | 14 | 91 | PTPN2 | 5 |
| 34 | PRKCA | 14 | 92 | SLC22A1 | 5 |
| 35 | SREBF2 | 14 | 93 | ACP1 | 4 |
| 36 | AKR1B1 | 13 | 94 | CD81 | 4 |
| 37 | CYP2D6 | 13 | 95 | CYP26B1 | 4 |
| 38 | NR1H4 | 13 | 96 | ENPP2 | 4 |
| 39 | SCD | 13 | 97 | GABRA2 | 4 |
| 40 | SLC6A4 | 13 | 98 | P2RX7 | 4 |
| 41 | CYP17A1 | 12 | 99 | SLC16A1 | 4 |
| 42 | ESR2 | 12 | 100 | OXER1 | 4 |
| 43 | MGAM | 12 | 101 | TAAR1 | 4 |
| 44 | PLG | 12 | 102 | CA4 | 3 |
| 45 | SELP | 12 | 103 | FDFT1 | 3 |
| 46 | ALOX15 | 11 | 104 | HSD11B2 | 3 |
| 47 | CNR2 | 11 | 105 | PPARD | 3 |
| 48 | FAAH | 11 | 106 | SERPINA6 | 3 |
| 49 | FABP4 | 11 | 107 | SLC5A2 | 3 |
| 50 | LGALS3 | 11 | 108 | PIN1 | 3 |
| 51 | MAOA | 11 | 109 | TRPM8 | 3 |
| 52 | NOS1 | 10 | 110 | RORC | 3 |
| 53 | NR1H3 | 10 | 111 | HPSE | 2 |
| 54 | PTGES | 10 | 112 | SLC22A6 | 2 |
| 55 | PTPN1 | 10 | 113 | PHLPP1 | 2 |
| 56 | SHBG | 10 | 114 | PTPRF | 2 |
| 57 | VDR | 10 | 115 | GSTK1 | 1 |
| 58 | ALOX12 | 9 | 116 | HEXA | 1 |
| KEGG ID & Description | Target Proteins | False Discovery Rate |
|---|---|---|
| has03320:PPAR signaling pathway | PPARA,PPARG,FABP2,FABP3,FABP4,SCD,NR1H3 | 0.00000152 |
| hsa04370:VEGF signaling pathway | PLA2G4A,PRKCA,PTGS2,VEGFA,MAPK1 | 0.00058 |
| hsa04066:HIF-1 signaling pathway | PRKCA,VEGFA,MAPK1,IL6,NOS2,HIF1A,HK1,HK2 | 0.00000794 |
| hsa04664:Fc epsilon RI signaling pathway | ALOX5,PLA2G4A,PRKCA,MAPK1 | 0.0047 |
| hsa04917:Prolactin signaling pathway | MAPK1,ESR1,ESR2,CYP17A1 | 0.0051 |
| hsa04657:IL-17 signaling pathway | MMP3,MAPK1,IL6,PTGS2,CASP3 | 0.0025 |
| hsa04933:AGE-RAGE signaling pathway in diabetic complications | PRKCA,VEGFA,MAPK1,IL6,CASP3 | 0.0027 |
| hsa04668:TNF signaling pathway | MMP3,MAPK1,IL6,PTGS2,CASP3 | 0.0037 |
| hsa04020:Calcium signaling pathway | P2RX7,NOS1,NOS2,PRKCA,GRIN1,CHRM2,TBXA2R,HTR2B,PRKCA,MAPK1,HIF1A,ESR1,KAT2B | 0.00028 |
| hsa04919:Thyroid hormone signaling pathway | PRKCA,MAPK1,HIF1A,ESR1,KAT2B | 0.0044 |
| hsa04662:B cell receptor signaling pathway | MAPK1,PTPN6,CD81 | 0.0266 |
| hsa04660:T cell receptor signaling pathway | PTPN6,PTPRC,IL2,MAPK1 | 0.0124 |
| hsa04926:Relaxin signaling pathway | NOS1,PRKCA,VEGFA,MAPK1,NOS2 | 0.0062 |
| hsa04910:Insulin signaling pathway | MAPK1,PTPRF,PTPN1,HK1,HK2 | 0.0066 |
| hsa04015:Rap1 signaling pathway | FGF1,FGF2,VEGFA,MAPK1,PRKCA,GRIN1,CNR1 | 0.0024 |
| hsa04912:GnRH signaling pathway | PLA2G4A,PRKCA,MAPK1 | 0.0411 |
| hsa04014:RAS signaling pathway | FGF1,FGF2,PLA2G4A,VEGFA,MAPK1,PRKCA,GRIN1 | 0.0034 |
| hsa04921:Oxytocin signaling pathway | PLA2G4A,PRKCA,PTGS2,MAPK1 | 0.0342 |
| hsa04151:PI3K-Akt signaling pathway | FGF1,PHLPP1,FGF2,VEGFA,MAPK1,IL2,IL6,CHRM2,PRKCA | 0.0025 |
| hsa04630: Jak-STAT signaling pathway | PTPN2,PTPN6,IL2,IL6 | 0.0408 |
| hsa04010:MAPK signaling pathway | FGF1,FGF2,PLA2G4A,VEGFA,PRKCA,MAPK1,CASP3 | 0.0093 |
| hsa04621:NOD-like receptor signaling pathway | CASP1,P2RX7,MAPK1,IL6 | 0.0446 |
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
|---|---|---|---|---|---|
| Protein | Ligand | PubChem ID | Binding Energy (kcal/mol) | Amino Acid Residue | Amino Acid Residue |
| IL6 (PDB ID: 4NI9) | Linolenic acid | 5,280,934 | −4.1 | N/A | Tyr31, Asp34, Gln111 |
| Ethyl-α-d-glucopyranoside | 9,169,4274 | −6.1 | Arg16 | Arg15 | |
| 3,5-Dihydroxy-6-(hydroxymethyl)oxan-2-one | 541,561 | −6.3 | N/A | N/A | |
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
| Protein | Positive control | PubChem ID | Binding energy(kcal/mol) | Amino acid Residue | Amino acid Residue |
| IL6 (PDB ID: 4NI9) | Veratric acid [27] | 7121 | −6.1 | Arg16 | N/A |
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
|---|---|---|---|---|---|
| Protein | Ligand | PubChem ID | Binding Energy (kcal/mol) | Amino Acid Residue | Amino Acid Residue |
| MAPK1(PDB ID:4IZ5) | Palmitic acid | 985 | −4.4 | Lys28, Ala26, Cys27 | Glu29, Asp30, Tyr62 |
| Met13, Glu186, Val14 | |||||
| Leu28, Arg15, Lys54 | |||||
| Glu12 | |||||
| 4-Dehydroxy-N-(4,5-methylenedioxy-2-nitrobenzylidene)tyramine | 610,062 | −7.0 | Leu60, Arg83, Leu107 | Ser61,Ser41,Arg50 | |
| Glu109, Met108, Ile31 | Ser29 | ||||
| Linolenic acid | 5,280,934 | −4.6 | Asn144 | Lys330, Glu303, Thr206 | |
| Lys207, Ser43, Ser47 | |||||
| Ser142, Leu8 | |||||
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
| Protein | Positive control | PubChem ID | Binding energy (kcal/mol) | Amino acid Residue | Amino acid Residue |
| MAPK1(PDB ID:4IZ5) | CU Cpt 22 [28] | 71,503,400 | −6.0 | Thr12, Ile15 | Thr41, Pro319, Lys138 |
| Ile324, Asn13, Ser320 | |||||
| Gly42, Leu17, Leu20 | |||||
| Thr16 |
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
|---|---|---|---|---|---|
| Protein | Ligand | PubChem ID | Binding Energy (kcal/mol) | Amino Acid Residue | Amino Acid Residue |
| P2RX7(PDB ID: 5U2H) | Pentanoic acid, 3-[(adamantan-1-ylmethyl)carbamoyl]-4-phenyl- | 4,920,612 | −5.9 | N/A | Val37, Phe344, Tyr343 |
| Val340, Tyr336, Leu45 | |||||
| Ile41, Tyr40 | |||||
| Phytone | 10,408 | −3.3 | N/A | Leu45, Tyr336, Ser339 | |
| Ile41, Val340, Tyr343 | |||||
| Val37, Ala44 | |||||
| 4-Cyclohexene-1,2-dicarboximide, N-butyl-, cis- | 91,733,922 | −4.1 | N/A | Val37, Ile41, Tyr36 | |
| Leu45, Ala44, Val340 | |||||
| Ser339, Tyr343 | |||||
| cis,cis,cis-7,10,13-Hexadecatrienal | 5,367,366 | −3.1 | N/A | Val340, Ile341, Gly345 | |
| Thr348, His34, Phe38 | |||||
| Phe344, Ile337 | |||||
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
| Protein | Positive control | PubChem ID | Binding energy(kcal/mol) | Amino acid Residue | Amino acid Residue |
| P2RX7(PDB ID: 5U2H) | KN-62 [30] | 5,312,126 | −9.8 | N/A | Tyr40, Ile41, Ala44 |
| Leu45, Tyr336, Val340 | |||||
| Tyr343, Ala347, Leu346 |
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
|---|---|---|---|---|---|
| Protein | Ligand | PubChem ID | Binding Energy (kcal/mol) | Amino Acid Residue | Amino Acid Residue |
| CASP1 (PDB ID: 3D6F) | Pentanoic acid, 3-[(adamantan-1-ylmethyl)carbamoyl]-4-phenyl- | 4,920,612 | −7.3 | Asn259, Leu258, Arg286 | Arg391, Glu390, Ile282 |
| Gln257 | Cys331, Ile243, Gly242 | ||||
| Ile239, Glu241, Gln240 | |||||
| Hydrogen Bond Interactions | Hydrophobic Interactions | ||||
| Protein | Positive Control | PubChem ID | Binding Energy (kcal/mol) | Amino acid Residue | Amino acid Residue |
| CASP1 (PDB ID: 3D6F) | Belnacasan [31] | 11,398,092 | −7.0 | Arg286, Arg391 | Cys285, Gly242, Ile239 |
| Glu241, Gln240, Asn259 | |||||
| Leu258, Pro335, Ile282 | |||||
| Ala284, Ile243 | |||||
| Mulberroside A [32] | 6,443,484 | −7.2 | Val184, Asn132, Gln358 | Gly188, Ile354, Met345 | |
| Asp381, Arg383 | Arg352, Val348, Ile350 | ||||
| Gly351, Asp185 | |||||
| Q-VD-Oph [33] | 24,794,416 | −7.2 | Gln240, Leu258, Asn259 | Cys285, Ile239, Arg286 | |
| Ile243, Gln257, Arg391 | |||||
| Glu390, Gly242, Ile282 |
| Parameters. | Compound Name | |||
|---|---|---|---|---|
| Ethyl- α-d-glucopyranoside | 3,5-Dihydroxy-6-(hydroxymethyl)oxan-2-one | 4-Dehydroxy-N-(4,5-methylenedioxy-2-nitrobenzylidene)tyramine | Pentanoic acid, 3-[(adamantan-1-ylmethyl)carbamoyl]-4-phenyl- | |
| Ames toxicity | NAT | NAT | AT | NAT |
| Carcinogens | NC | NC | NC | NC |
| Acute oral toxicity | IV | Ⅲ | Ⅲ | Ⅲ |
| Rat acute toxicity | 0.9919 | 1.4924 | 2.6672 | 2.0497 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, K.-K.; Adnan, M.; Cho, D.-H. Network Pharmacology Study to Interpret Signaling Pathways of Ilex cornuta Leaves against Obesity. Processes 2021, 9, 1106. https://doi.org/10.3390/pr9071106
Oh K-K, Adnan M, Cho D-H. Network Pharmacology Study to Interpret Signaling Pathways of Ilex cornuta Leaves against Obesity. Processes. 2021; 9(7):1106. https://doi.org/10.3390/pr9071106
Chicago/Turabian StyleOh, Ki-Kwang, Md. Adnan, and Dong-Ha Cho. 2021. "Network Pharmacology Study to Interpret Signaling Pathways of Ilex cornuta Leaves against Obesity" Processes 9, no. 7: 1106. https://doi.org/10.3390/pr9071106
APA StyleOh, K.-K., Adnan, M., & Cho, D.-H. (2021). Network Pharmacology Study to Interpret Signaling Pathways of Ilex cornuta Leaves against Obesity. Processes, 9(7), 1106. https://doi.org/10.3390/pr9071106