
Biocatalytic Conversion of Short-Chain Fatty Acids to Corresponding Alcohols in Escherichia coli




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Construction
2.2. Fatty Alcohol Production
2.3. Analytic Methods
3. Results
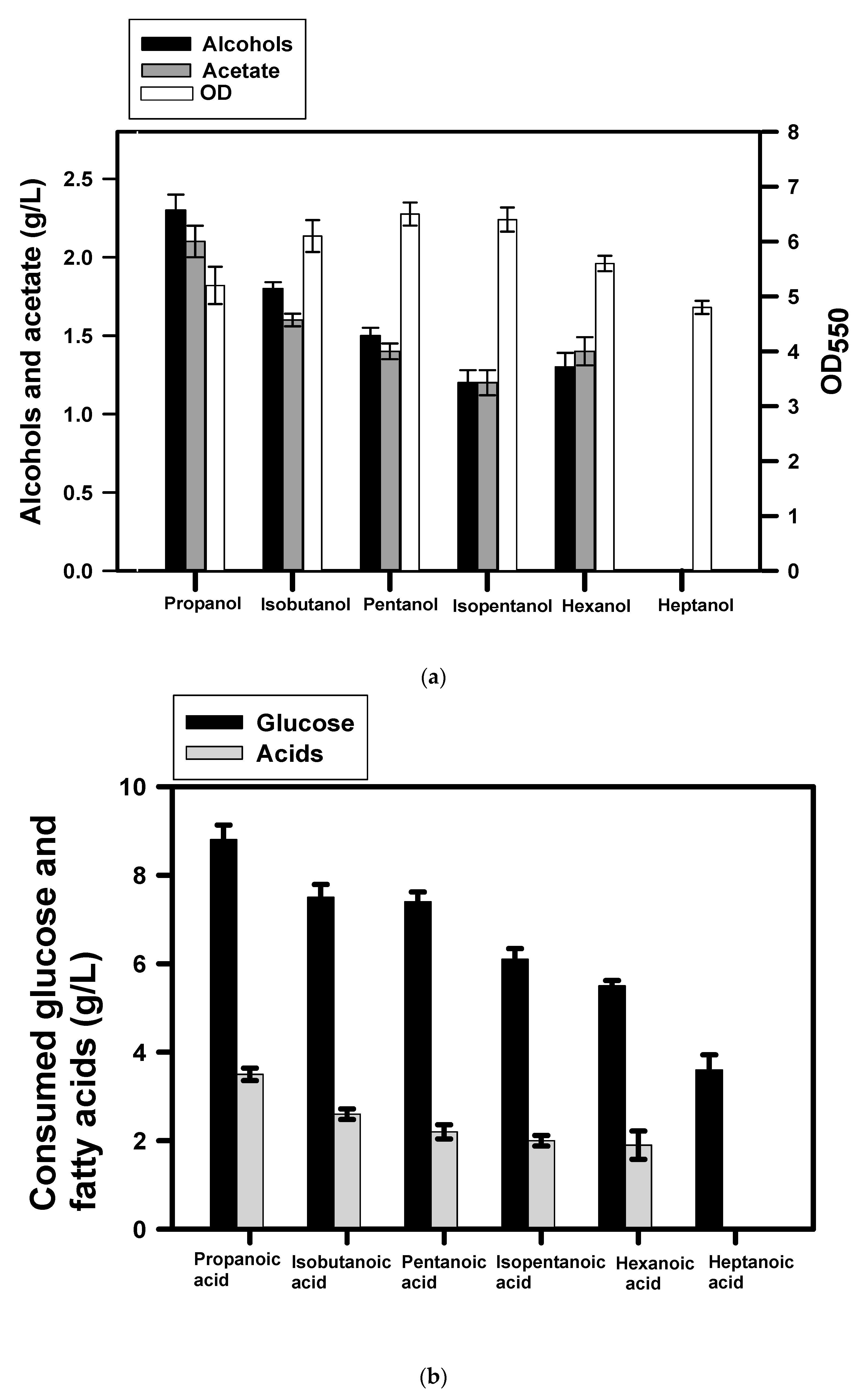
3.1. Production of Fatty Alcohols
3.2. Propanol Production by Substrate Feeding
3.3. Improvement of Propanol Production
3.4. n-Pentanol Production by Heavy Inoculum
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Schubert, C. Can biofuels finally take center stage? Nat. Biotechnol. 2006, 24, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.; Dragone, G.; Guimarães, P.M.; Silva, J.P.; Carneiro, L.M.; Roberto, I.C.; Vicentea, A.; Dominguesa, L.; Teixeira, J.A. Technological trends, global market, and challenges of bio-ethanol production. Biotechnol. Adv. 2010, 28, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Dürre, P. Biobutanol: An attractive biofuel. Biotechnol. J. 2007, 2, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Tsujimura, T.; Pitz, W.J.; Gillespie, F.; Curran, H.J.; Weber, B.W.; Zhang, Y.; Sung, C.J. Development of isopentanol reaction mechanism reproducing autoignition character at high and low temperatures. Energy Fuels 2012, 26, 4871–4886. [Google Scholar] [CrossRef]
- Wei, L.; Cheung, C.S.; Huang, Z. Effect of n-pentanol addition on the combustion, performance and emission characteristics of a direct-injection diesel engine. Energy 2014, 70, 172–180. [Google Scholar] [CrossRef]
- Masum, B.M.; Masjuki, H.H.; Kalam, M.A.; Palash, S.M.; Wakil, M.A.; Imtenan, S. Tailoring the key fuel properties using different alcohols (c2–c6) and their evaluation in gasoline engine. Energy Convers. Manag. 2014, 88, 382–390. [Google Scholar] [CrossRef]
- Walther, T.; François, J.M. Microbial production of propanol. Biotechnol. Adv. 2016, 34, 984–996. [Google Scholar] [CrossRef]
- Jones, D.T.; Woods, D.R. Acetone-butanol fermentation revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar] [CrossRef]
- Lee, S.Y.; Park, J.H.; Jang, S.H.; Nielsen, L.K.; Kim, J.; Jung, K.S. Fermentative butanol production by clostridia. Biotechnol. Bioeng. 2008, 101, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Wood, H.G. Metabolic cycles in the fermentation by propionic acid bacteria. Curr. Top. Cell. Regul. 1981, 18, 255–287. [Google Scholar]
- Tholozan, J.L.; Touzel, J.P.; Samain, E.; Grivet, J.P.; Prensier, G.; Albagnac, G. Clostridium neopropionicum sp. Nov., a strict anaerobic bacterium fermenting ethanol to propionate through acrylate pathway. Arch. Microbiol. 1992, 157, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Ramió-Pujol, S.; Ganigué, R.; Bañeras, L.; Colprim, J. Incubation at 25 °C prevents acid crash and enhances alcohol production in clostridium carboxidivorans p7. Bioresour. Technol. 2015, 192, 296–303. [Google Scholar] [CrossRef]
- Cheon, S.; Kim, H.M.; Gustavsson, M.; Lee, S.Y. Recent trends in metabolic engineering of microorganisms for the production of advanced biofuels. Curr. Opin. Chem. Biol. 2016, 35, 10–21. [Google Scholar] [CrossRef]
- Das, M.; Patra, P.; Ghosh, A. Metabolic engineering for enhancing microbial biosynthesis of advanced biofuels. Renew. Sust. Energy Rev. 2020, 119, 109562. [Google Scholar] [CrossRef]
- Saini, M.; Chen, M.H.; Chiang, C.J.; Chao, Y.P. Potential production platform of n-butanol in Escherichia coli. Metab. Eng. 2015, 27, 76–82. [Google Scholar] [CrossRef]
- Saini, M.; Li, S.Y.; Chiang, C.J.; Chao, Y.P. Systematic engineering of the central metabolism in Escherichia coli for effective production of n-butanol. Biotechnol. Biofuels 2016, 9, 69. [Google Scholar] [CrossRef]
- Sramek, S.J.; Frerman, F.E. Purification and properties of Escherichia coli coenzyme a-transferase. Arch. Biochem. Biophys. 1975, 171, 14–26. [Google Scholar] [CrossRef]
- Bell, G.H. Solubilities of normal aliphatic acids, alcohols and alkanes in water. Chem. Phys. Lipids 1973, 10, 1–10. [Google Scholar] [CrossRef]
- Roe, A.J.; O’Byrne, C.; McLaggan, D.; Booth, I.R. Inhibition of Escherichia coli growth by acetic acid: A problem with methionine biosynthesis and homocysteine toxicity. Microbiology 2002, 148, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Beatty, C.M.; Browning, D.F.; Busby, S.J.; Simel, E.J.; Hovel-Miner, G.; Wolfe, A.J. Regulation of acetyl coenzyme a synthetase in Escherichia coli. J. Bacteriol. 2000, 182, 4173–4179. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gu, J.; Wang, X.; Zhang, X.E.; Deng, J. Acs is essential for propionate utilization in Escherichia coli. Biochem. Biophys. Res. Commun. 2014, 449, 272–277. [Google Scholar] [CrossRef]
- Horswill, A.R.; Dudding, A.R.; Escalante-Semerena, J.C. Studies of propionate toxicity in Salmonella enterica identify 2-methylcitrate as a potent inhibitor of cell growth. J. Biol. Chem. 2001, 276, 19094–19101. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.R.; Liao, J.C. Synergy as design principle for metabolic engineering of 1-propanol production in Escherichia coli. Metab. Eng. 2013, 17. [Google Scholar] [CrossRef]
- Srirangan, K.; Liu, X.; Westbrook, A.; Akawi, L.; Pyne, M.E.; Moo-Young, M.; Chou, C.P. Biochemical, genetic, and metabolic engineering strategies to enhance coproduction of 1-propanol and ethanol in engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 9499–9515. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Yan, Y. Dehydratase mediated 1-propanol production in metabolically engineered Escherichia coli. Microb. Cell Factories 2011, 10, 97. [Google Scholar] [CrossRef]
- Jain, R.; Sun, X.; Yuan, Q.; Yan, Y. Systematically engineering Escherichia coli for enhanced production of 1,2-propanediol and 1-propanol. ACS Synth. Biol. 2014, 4, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, L.S.; Nunn, W.D. Genetic and molecular characterization of the genes involved in short-chain fatty acid degradation in Escherichia coli: The ato system. J. Bacteriol. 1987, 169, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Machado, H.B.; Dekishima, Y.; Luo, H.; Lan, E.I.; Liao, J.C. A selection platform for carbon chain elongation using the coa-dependent pathway to produce linear higher alcohols. Metab. Eng. 2012, 14, 504–511. [Google Scholar] [CrossRef]
- Chen, G.S.; Siao, S.W.; Shen, C.R. Saturated mutagenesis of ketoisovalerate decarboxylase V461 enabled specific synthesis of 1-pentanol via the ketoacid elongation cycle. Sci. Rep. 2017, 7, 11284. [Google Scholar] [CrossRef]
- Polen, T.; Rittmann, D.; Wendisch, V.F.; Sahm, H. DNA microarray analyses of the long-term adaptive response of Escherichia coli to acetate and propionate. Appl. Environ. Microbiol. 2003, 69, 1759–1774. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.-J.; Saini, M.; Chiang, C.-J.; Chao, Y.-P. Biocatalytic Conversion of Short-Chain Fatty Acids to Corresponding Alcohols in Escherichia coli. Processes 2021, 9, 973. https://doi.org/10.3390/pr9060973
Lin L-J, Saini M, Chiang C-J, Chao Y-P. Biocatalytic Conversion of Short-Chain Fatty Acids to Corresponding Alcohols in Escherichia coli. Processes. 2021; 9(6):973. https://doi.org/10.3390/pr9060973
Chicago/Turabian StyleLin, Li-Jen, Mukesh Saini, Chung-Jen Chiang, and Yun-Peng Chao. 2021. "Biocatalytic Conversion of Short-Chain Fatty Acids to Corresponding Alcohols in Escherichia coli" Processes 9, no. 6: 973. https://doi.org/10.3390/pr9060973
APA StyleLin, L.-J., Saini, M., Chiang, C.-J., & Chao, Y.-P. (2021). Biocatalytic Conversion of Short-Chain Fatty Acids to Corresponding Alcohols in Escherichia coli. Processes, 9(6), 973. https://doi.org/10.3390/pr9060973