
Enhancing Methanogenic Efficiency in Dry Anaerobic Digestion of Kitchen Waste Through Optimization of Total Solids Content

Abstract
1. Introduction
2. Materials and Methods
2.1. Substrate and Inoculum
2.2. Experimental Program
2.3. Analytical Methods
2.4. Microbial Analysis
3. Results and Discussion
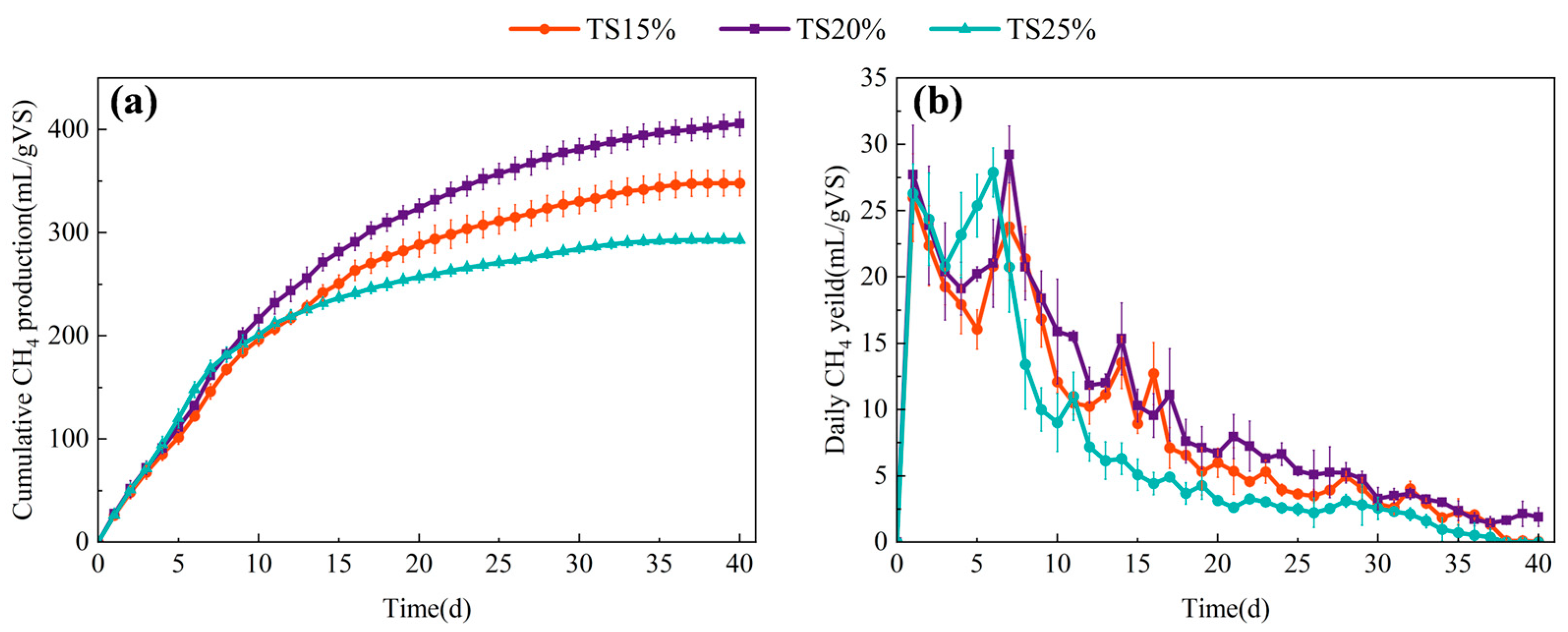
3.1. Effect of TS on the Methane Production Performance of Dry Anaerobic System
3.2. Effect of Increasing TS on the Change of System Stability
3.2.1. pH
3.2.2. TAN
3.2.3. VFAs and Their Compositions
3.2.4. Correlation Analysis
3.3. Effect of Increasing TS on Organic Matter Changes
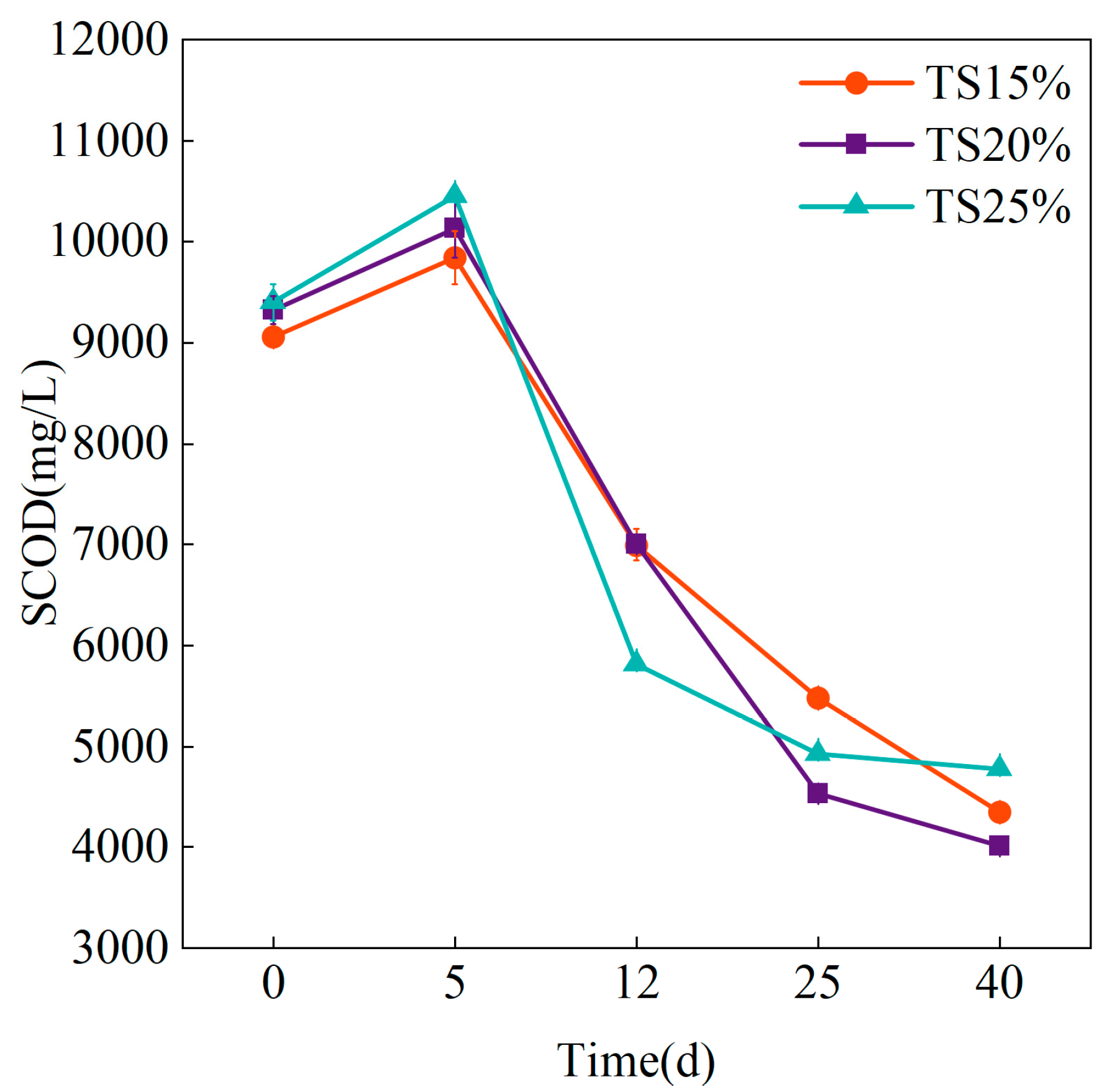
3.3.1. SCOD
3.3.2. SPS
3.3.3. SPN
3.4. Effect of TS on Microbial Community Structure
3.4.1. Bacterial Community Structure
3.4.2. Archaeal Community Structure
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| TS | Total Solids |
| VS | Volatile Solids |
| TAN | Total Ammonia Nitrogen |
| VFAs | Volatile Fatty Acids |
| SCOD | Soluble Chemical Oxygen Demand |
| SPS | Soluble Polysaccharides |
| SPN | Soluble Protein Nitrogen |
References
- Zhang, C.; Su, H.; Baeyens, J.; Tan, T. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sustain. Energy Rev. 2014, 38, 383–392. [Google Scholar] [CrossRef]
- Zhang, S.; Ma, X.; Sun, H.; Zhao, P.; Wang, Q.; Wu, C.; Gao, M. Response of semi-continuous anaerobic digestion of food waste to progressively increasing temperature: Methanogen community, correlation analysis, and energy balance. Ind. Crop. Prod. 2023, 192, 116066. [Google Scholar] [CrossRef]
- Du, M.; Liu, X.; Wang, D.; Yang, Q.; Duan, A.; Chen, H.; Liu, Y.; Wang, Q.; Ni, B.J. Understanding the fate and impact of capsaicin in anaerobic co-digestion of food waste and waste activated sludge. Water Res. 2021, 188, 116539. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Gnansounou, E.; Rebello, S.; Binod, P.; Varjani, S.; Thakur, I.S.; Nair, R.B.; Pandey, A. Conversion of food and kitchen waste to value-added products. J. Environ. Manag. 2019, 241, 619–630. [Google Scholar] [CrossRef]
- Chavan, S.; Yadav, B.; Tyagi, R.D.; Wong, J.W.C.; Drogui, P. Trends and challenges in the valorization of kitchen waste to polyhydroxyalkanoates. Bioresour. Technol. 2023, 369, 128323. [Google Scholar] [CrossRef]
- Liu, X.; Shen, J.; Guo, Y.; Wang, S.; Chen, B.; Luo, L.; Zhang, H. Technical progress and perspective on the thermochemical conversion of kitchen waste and relevant applications: A comprehensive review. Fuel 2023, 331, 125803. [Google Scholar] [CrossRef]
- Su, G.; Ong, H.C.; Fattah, I.M.R.; Ok, Y.S.; Jang, J.H.; Wang, C.T. State-of-the-art of the pyrolysis and co-pyrolysis of food waste: Progress and challenges. Sci. Total Environ. 2022, 809, 151170. [Google Scholar] [CrossRef]
- Zhang, P.; Chai, J.; Cao, J.; Qin, Y.; Dang, M.; Geng, K.; Wei, Y. Landfill leachate generation mechanism study: A review. Int. J. Environ. Sci. Technol. 2023, 20, 9271–9290. [Google Scholar] [CrossRef]
- Ajay, C.M.; Mohan, S.; Dinesha, P. Decentralized energy from portable biogas digesters using domestic kitchen waste: A review. Waste Manag. 2021, 125, 10–26. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Li, H.; Lu, Y.; Zhang, B.; Zhang, H.; Zhang, S. Synergistic effects of economic benefits, resource conservation and carbon mitigation of kitchen waste recycling from the perspective of carbon neutrality. Resour. Conserv. Recycl. 2023, 199, 107262. [Google Scholar] [CrossRef]
- Kim, M.-S.; Na, J.-G.; Lee, M.-K.; Ryu, H.; Chang, Y.-K.; Triolo, J.M.; Yun, Y.-M.; Kim, D.-H. More value from food waste: Lactic acid and biogas recovery. Water Res. 2016, 96, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiang, Y.; Wang, S.; Zhang, Y.; Hu, Y.; Hu, Z.-h.; Wu, G.; Zhan, X. Impact of total solids content on anaerobic co-digestion of pig manure and food waste: Insights into shifting of the methanogenic pathway. Waste Manag. 2020, 114, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Bollon, J.; Benbelkacem, H.; Gourdon, R.; Buffière, P. Measurement of diffusion coefficients in dry anaerobic digestion media. Chem. Eng. Sci. 2013, 89, 115–119. [Google Scholar] [CrossRef]
- Li, Y.; Park, S.Y.; Zhu, J. Solid-state anaerobic digestion for methane production from organic waste. Renew. Sustain. Energy Rev. 2011, 15, 821–826. [Google Scholar] [CrossRef]
- Hua, D.; Hou, X.; Zhao, Y.; Xu, H.; Jin, F.; Chen, L.; Meng, G.; Li, Y. Structural optimization and performance evaluation of high-solid anaerobic digestion reactors by CFD simulation at various scales. J. Water Process Eng. 2025, 70, 106999. [Google Scholar] [CrossRef]
- Peng, X.; Nges, I.A.; Liu, J. Improving methane production from wheat straw by digestate liquor recirculation in continuous stirred tank processes. Renew. Energy 2016, 85, 12–18. [Google Scholar] [CrossRef]
- Nie, H.; Jacobi, H.F.; Strach, K.; Xu, C.; Zhou, H.; Liebetrau, J. Mono-fermentation of chicken manure: Ammonia inhibition and recirculation of the digestate. Bioresour. Technol. 2015, 178, 238–246. [Google Scholar] [CrossRef]
- Song, Y.; Liu, J.; Chen, M.; Zheng, J.; Gui, S.; Wei, Y. Application of mixture design to optimize organic composition of carbohydrate, protein, and lipid on dry anaerobic digestion of OFMSW: Aiming stability and efficiency. Biochem. Eng. J. 2021, 172, 108037. [Google Scholar] [CrossRef]
- American Public Health Association; American Water Works Association; Water Environment Federation. Standard Methods for Examination of Water and Wastewater; American Public Health Association; American Water Works Association; Water Environment Federation: Washington, DC, USA, 2012; p. 185. [Google Scholar]
- Xu, M.; Gao, P.; Chen, H.-Q.; Shen, X.-X.; Xu, R.-Z.; Cao, J.-S. Metagenomic insight into the prevalence and driving forces of antibiotic resistance genes in the whole process of three full-scale wastewater treatment plants. J. Environ. Manag. 2023, 344, 118369. [Google Scholar] [CrossRef]
- Abbassi-Guendouz, A.; Brockmann, D.; Trably, E.; Dumas, C.; Delgenès, J.-P.; Steyer, J.-P.; Escudié, R. Total solids content drives high solid anaerobic digestion via mass transfer limitation. Bioresour. Technol. 2012, 111, 55–61. [Google Scholar] [CrossRef]
- Zhen, F.; Wu, D.; Sun, Y.; Qu, B.; Li, L.; Li, Y.; Li, Q.; Xing, T. Effect of different organic loads on the performance and microbial community mechanism of dry anaerobic digestion. Fuel 2024, 361, 130615. [Google Scholar] [CrossRef]
- Yao, Z.; Chunxing, L.; Zengwei, Y.; Ruming, W.; Irini, A.; Gefu, Z. Syntrophy mechanism, microbial population, and process optimization for volatile fatty acids metabolism in anaerobic digestion. Chem. Eng. J. 2023, 452, 139137. [Google Scholar]
- Mercado, J.V.; Koyama, M.; Nakasaki, K. Short-term changes in the anaerobic digestion microbiome and biochemical pathways with changes in organic load. Sci. Total Environ. 2022, 813, 152585. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Chen, R.; Huang, L.; Wang, X.; Chou, S.; Zhu, J. Acidogenic fermentation of potato peel waste for volatile fatty acids production: Effect of initial organic load. J. Biotechnol. 2023, 374, 114–121. [Google Scholar] [CrossRef]
- Rajagopal, R.; Massé, D.I.; Singh, G. A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresour. Technol. 2013, 143, 632–641. [Google Scholar] [CrossRef]
- Tian, H.; Fotidis, I.A.; Kissas, K.; Angelidaki, I. Effect of different ammonia sources on aceticlastic and hydrogenotrophic methanogens. Bioresour. Technol. 2018, 250, 390–397. [Google Scholar] [CrossRef]
- Yapeng, S.; Wanrong, H.; Wei, Q.; Maria, W.; Wandera, S.M.; Renjie, D. Upgrading the performance of high solids feeding anaerobic digestion of chicken manure under extremely high ammonia level. Renew. Energy 2022, 194, 13–20. [Google Scholar]
- Yang, J.; Zhang, J.; Du, X.; Gao, T.; Cheng, Z.; Fu, W.; Wang, S. Ammonia inhibition in anaerobic digestion of organic waste: A review. Int. J. Environ. Sci. Technol. 2024, 22, 3927–3942. [Google Scholar] [CrossRef]
- Niu, Q.; Qiao, W.; Qiang, H.; Li, Y.-Y. Microbial community shifts and biogas conversion computation during steady, inhibited and recovered stages of thermophilic methane fermentation on chicken manure with a wide variation of ammonia. Bioresour. Technol. 2013, 146, 223–233. [Google Scholar] [CrossRef]
- Zhuo, Y.; Wang, H.; Wang, X.; Jing, D.; Zhou, M.; Peng, D.; Han, Y. Performance of electroactive anaerobic granular sludge under ammonia stress: Performance, microbe and morphology. Bioresour. Technol. 2025, 424, 132295. [Google Scholar] [CrossRef]
- Zhu, C.; Cao, Z.; Wang, H.; Yuan, H.; Li, X. High-solid anaerobic digestion performance of municipal organic solid waste: Methane yield, microbial communities, enzymes, and key metabolic pathways. J. Environ. Chem. Eng. 2025, 13, 116297. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, H.; Guan, D.; Wang, Y.; Fu, Z.; Sun, Y.; Wang, D.; Zhang, H. New insights into mechanism of emerging pollutant polybrominated diphenyl ether inhibiting sludge dark fermentation. Bioresour. Technol. 2023, 368, 128358. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-W.; Feng, G.; Hong, X.; Wang, M.; Zhang, Q.; Sun, Z.-Y.; Chen, Y.-T.; Tang, Y.-Q. Effects of high solid content and straw proportion on volatile fatty acids production from straw, sludge and food wastes: Performance and microbial community characteristics. Appl. Biol. Chem. 2024, 67, 83. [Google Scholar] [CrossRef]
- Chen, X.; Liu, W.; Zhao, Y.; He, H.; Ma, J.; Cui, Z.; Yuan, X. Optimization of semi-continuous dry anaerobic digestion process and biogas yield of dry yellow corn straw: Based on “gradient anaerobic digestion reactor”. Bioresour. Technol. 2023, 389, 129804. [Google Scholar] [CrossRef]
- Liang, Y.; Yu, J.; Yao, Z.; Sun, Y.; Zhao, L. Performance, interaction, and metabolic pathway of novel dry–wet anaerobic digestion for treating high-solid agricultural waste. Energy 2024, 304, 132055. [Google Scholar] [CrossRef]
- Nam, J.-Y.; Yates, M.D.; Zaybak, Z.; Logan, B.E. Examination of protein degradation in continuous flow, microbial electrolysis cells treating fermentation wastewater. Bioresour. Technol. 2014, 171, 182–186. [Google Scholar] [CrossRef]
- Tiedong, L.; Tianming, S.; Xuelian, L.; Yuansong, W.; Junya, Z.; Tieguang, H. Dual character of methane production improvement and antibiotic resistance genes reduction by nano-Fe2O3 addition during anaerobic digestion of swine manure. J. Clean. Prod. 2022, 376, 134240. [Google Scholar]
- Ren, S.; Usman, M.; Tsang, D.C.W.; O-Thong, S.; Angelidaki, I.; Zhu, X.; Zhang, S.; Luo, G. Hydrochar-Facilitated Anaerobic Digestion: Evidence for Direct Interspecies Electron Transfer Mediated through Surface Oxygen-Containing Functional Groups. Environ. Sci. Technol. 2020, 54, 5755–5766. [Google Scholar] [CrossRef]
- Lee, J.; Hong, J.; Jeong, S.; Chandran, K.; Park, K.Y. Interactions between substrate characteristics and microbial communities on biogas production yield and rate. Bioresour. Technol. 2020, 303, 122934. [Google Scholar] [CrossRef]
- Zixin, W.; Cheng, Z.; Jamison, W.; Sharma, B.K.; Buchun, S.; Yuanhui, Z. Adsorption or direct interspecies electron transfer? A comprehensive investigation of the role of biochar in anaerobic digestion of hydrothermal liquefaction aqueous phase. Chem. Eng. J. 2022, 435, 135078. [Google Scholar]
- Zamorano-López, N.; Borrás, L.; Seco, A.; Aguado, D. Unveiling microbial structures during raw microalgae digestion and co-digestion with primary sludge to produce biogas using semi-continuous AnMBR systems. Sci. Total Environ. 2020, 699, 134365. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Wang, Y.; Dai, Y.; Zhou, S.; Wang, B.; Li, Y.; Li, J. Batch and semi–continuous experiments examining the sludge mesophilic anaerobic digestive performance with different varieties of rice straw. Bioresour. Technol. 2022, 346, 126651. [Google Scholar] [CrossRef] [PubMed]
- Ariesyady, H.D.; Ito, T.; Okabe, S. Functional bacterial and archaeal community structures of major trophic groups in a full-scale anaerobic sludge digester. Water Res. 2007, 41, 1554–1568. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.; Si, D.; Chen, Q. Evolution of microbial community along with increasing solid concentration during high-solids anaerobic digestion of sewage sludge. Bioresour. Technol. 2016, 216, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wang, Y.; Wang, Y.; Chin, F.Y.L.; Zhang, T. Cellular adhesiveness and cellulolytic capacity in Anaerolineae revealed by omics-based genome interpretation. Biotechnol. Biofuels 2016, 9, 111. [Google Scholar] [CrossRef]
- Kurth, J.M.; Op den Camp, H.J.M.; Welte, C.U. Several ways one goal—Methanogenesis from unconventional substrates. Appl. Microbiol. Biotechnol. 2020, 104, 6839–6854. [Google Scholar] [CrossRef]
- Conrad, R. Importance of hydrogenotrophic, aceticlastic and methylotrophic methanogenesis for methane production in terrestrial, aquatic and other anoxic environments: A mini review. Pedosphere 2020, 30, 25–39. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Inoculation Sludge | Kitchen Waste | Mixture |
|---|---|---|---|
| TS (%) | 24.73 | 52.3 | 25.41 |
| VS (%) | 8.75 | 51.8 | 11.36 |
| VS/TS (%) | 35.39 | 99.04 | 44.7 |
| pH | 8.46 | 5.02 | 8.14 |
| TAN/(mg/L) | 1589.76 | - | 2332.89 |
| SCOD/(mg/L) | 8202.5 | - | 9452.5 |
| SPN/(mg/L) | 1580.29 | - | 1951.62 |
| SPS/(mg/L) | 1309.03 | - | 1834.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Zheng, J.; Liu, J. Enhancing Methanogenic Efficiency in Dry Anaerobic Digestion of Kitchen Waste Through Optimization of Total Solids Content. Processes 2025, 13, 2332. https://doi.org/10.3390/pr13082332
Liu L, Zheng J, Liu J. Enhancing Methanogenic Efficiency in Dry Anaerobic Digestion of Kitchen Waste Through Optimization of Total Solids Content. Processes. 2025; 13(8):2332. https://doi.org/10.3390/pr13082332
Chicago/Turabian StyleLiu, Lan, Jiaxi Zheng, and Jianwei Liu. 2025. "Enhancing Methanogenic Efficiency in Dry Anaerobic Digestion of Kitchen Waste Through Optimization of Total Solids Content" Processes 13, no. 8: 2332. https://doi.org/10.3390/pr13082332
APA StyleLiu, L., Zheng, J., & Liu, J. (2025). Enhancing Methanogenic Efficiency in Dry Anaerobic Digestion of Kitchen Waste Through Optimization of Total Solids Content. Processes, 13(8), 2332. https://doi.org/10.3390/pr13082332