Abstract
Anaerobic ammonium oxidation (anammox) has shown success in past years for the treatment of municipal and industrial wastewater containing inorganic nutrients (i.e., nitrogen). However, the increase in polycyclic aromatic hydrocarbon (PAH)-contaminated matrices calls for new strategies for efficient and environmentally sustainable remediation. Therefore, the present review examined the literature on the connection between the anammox process and PAHs using VOSviewer to shed light on the mechanisms involved during PAH biodegradation and the key factors affecting anammox bacteria. The scientific literature thoroughly discussed here shows that PAHs can be involved in nitrogen removal by acting as electron donors, and their presence does not adversely affect the anammox bacteria. Anammox activity can be improved by regulating the operating parameters (e.g., organic load, dissolved oxygen, carbon-to-nitrogen ratio) and external supplementation (i.e., calcium nitrate) that promote changes in the microbial community (e.g., Candidatus Jettenia), favoring PAH degradation. The onset of a synergistic dissimilatory nitrate reduction to ammonium and partial denitrification can be beneficial for PAH and nitrogen removal.
1. Introduction
Anaerobic ammonium oxidation (anammox) is an autotrophic process underlying the biogeoclimatic nitrogen cycle mediated by anammox bacteria [1], in which the ammonium is oxidized to molecular nitrogen under anoxic conditions using nitrite as a terminal electron acceptor (TEA) [2,3,4]. This process is an attractive biological alternative to conventional nitrification and denitrification treatments for nitrogen removal, with efficiencies above 90% [5,6]. The advantages of the anammox process are the reduction in the oxygen demand for aeration (i.e., approximately 50–60%), the 100% decrease in external organic matter, lower sludge production, savings in energy costs, and reduced greenhouse gas emissions [7,8,9]. Anammox microbial communities may be abundant in harbor areas, marine sediments, oil reservoirs, mangrove soil, and urban wastewater and, thus, in sites contaminated by polycyclic aromatic hydrocarbons (PAHs) [4,10,11].
PAHs are a class of organic contaminants released into the environment from natural and anthropic sources [12]. The U.S. Environment and Protection Agency (EPA) recognized 28 PAHs as priority pollutants [13] that are toxic, carcinogenic, and mutagenic, posing a risk to human health [14,15]. PAHs have low solubility in water and high lipophilicity and are considered recalcitrant compounds [16,17,18]. Among the available remediation techniques, bioremediation is the most favored and environmentally sustainable [19]. However, the biological processes may have some limitations (e.g., bioavailability) and depend on different parameters (e.g., dissolved oxygen) [20]. The degradation of PAHs under anoxic conditions can be enhanced by employing supplementary TEAs, such as nitrate, as can occur during denitrification [21]. Thus, the anammox process converting nitrite produced during denitrification to N2 can be involved in PAH biodegradation.
The academic community has recently paid increased attention to PAH and nitrogen removal topics (e.g., nitrate reduction, anammox) (Figure 1a). However, previous reviews have not shed light on the mechanisms and impacts associated with PAH degradation during the anammox process. Furthermore, the identification of bacterial consortia with capacities and growth adaptive to PAHs is crucial for the bioremediation of polluted sites. Hence, the state of knowledge on PAH removal in the presence of anammox bacteria requires investigation.
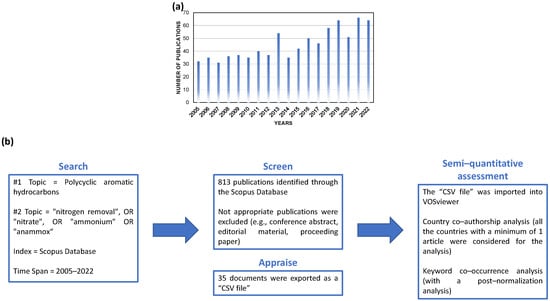
Figure 1.
(a) Publications over time (i.e., from 2005 to 2022) identified by analyzing the keywords “polycyclic aromatic hydrocarbons” with “nitrogen removal” OR “nitrate” OR “ammonium” OR “anammox” in the Scopus database. (b) Flow chart of the literature screening procedure. The literature search was conducted on 27 November 2022.
This review is the first to evaluate the degradation capabilities of anammox bacteria on matrices contaminated by PAHs and nitrogen compounds by focusing on the main anammox PAH-degrading phylotypes, identifying the primary issues in the anammox process and highlighting possible future development strategies to broaden the application of this process to a larger scale.
2. Data Collection and Extraction
For this study, semiquantitative evaluation was conducted using VOSviewer (https://app.vosviewer.com/, accessed on 27 November 2022) software. This software can develop maps of interconnected data [22]. The data were obtained from the Scopus database (https://www.scopus.com/, accessed on 27 November 2022). A total of 813 articles published between 2005 and 2022 were obtained from the Scopus database by searching for “polycyclic aromatic hydrocarbons” with “nitrogen removal” OR “nitrate” OR “ammonium” OR “anammox” (Figure 1b). The pertinent papers were manually screened to remove inconsistent references (Figure 1b) automatically found through the document search. Thus, 35 documents were exported as a CSV file and then imported into VOSviewer, which was executed with the settings for country co-authorship and keyword co-occurrence (Figure 1b).
The generated map is illustrated in Figure 2. Countries and keywords are marked with colored circles. The dimensions of the circles are associated with the co-presence of the item (e.g., keyword) in the title and abstract. Hence, the extent of a circle or line object was determined by its weight. An increase in object weight led to a more marked circle and line.
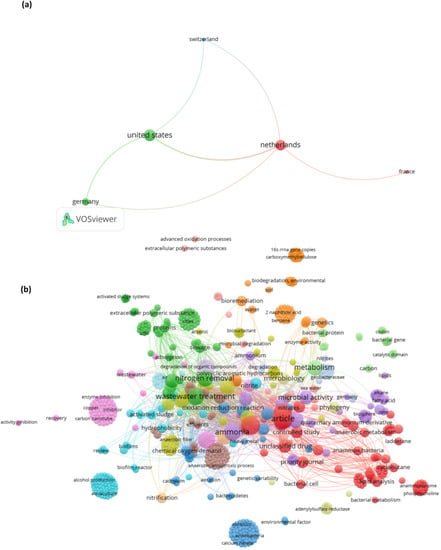
Figure 2.
Semiquantitative evaluation of the data collected from the Scopus database using VOSviewer software. The network map shows (a) the relations between various countries in the field and (b) keyword co-occurrences. The line thickness and circle size represent the correlations between the items. The colors of the lines or circles were obtained according to the colors of the items. Analogous item colors indicate correlation.
The articles were published in 14 different nations. All countries with a minimum of one article were considered for the analysis. Among the countries, the Netherlands, the United States, Germany, and China accounted for 8, 12, 4, and 12 publications, respectively. The network visualization of co-authorship relationships between nations suggested that the Netherlands and the United States are at the center of this research topic (Figure 2a). Furthermore, remarkable research collaborations were displayed between Germany and other countries (Figure 2a).
Regarding keyword co-occurrence, a total of 892 items grouped into 16 large clusters were identified (Figure 2b). For instance, the keywords “hydrocarbon”, “petroleum hydrocarbons”, and “total petroleum hydrocarbons” were grouped into the same cluster as “anammox bacteria” and “anammox process” (Figure 2b). After a normalization analysis performed via the association method, it was found that the clusters referring to PAH and anammox results were connected through the wastewater treatment cluster (Figure 2b). This indicated that the abovementioned keywords are highly correlated and are thus discussed in detail below.
3. Nitrogen Biogeochemistry and the Anammox Process
Nitrogen is the fifth most abundant element in our solar system and the basis for synthesizing nucleic acids and proteins, the two most essential life polymers [23]. The valence of nitrogen is variable since it can assume all oxidation numbers from a maximum of +5 to a minimum of −3, thus changing the oxidation state through microbial processes [24]. Although nitrogen is highly abundant in the Earth’s atmosphere (accounting for approximately 78%), the triple bond (N≡N) makes molecular nitrogen highly inert, and N2 has to be fixed by microorganisms during the so-called nitrogen cycle (N-cycle) before being readily useable by other organisms [25]. The N-cycle includes three main stages (Figure 3a) [26]: (i) the fixation of molecular nitrogen in compounds usable by living organisms, such as ammonium ions (NH4+) or ammonia (NH3); (ii) nitrification through the transformation of N–NH4+ to nitrite (NO2−) and then nitrate (NO3−); and (iii) denitrification through the reduction of nitrate to molecular nitrogen, which returns to the atmosphere and closes the N-cycle. These are the main pathways of the N-cycle, excluding the anammox process (Figure 3a), where nitrite acts as a TEA and ammonium as an electron donor [27].
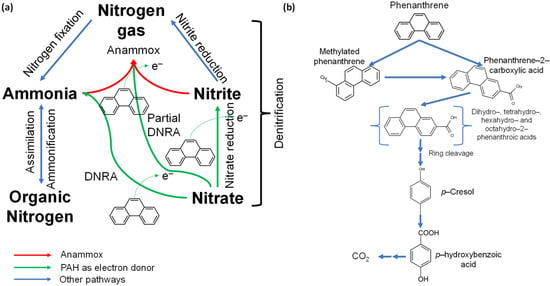
Figure 3.
(a) Nitrogen biogeochemistry and (b) biodegradation of polycyclic aromatic hydrocarbons [28,29].
In general, the anammox reaction can be divided into three steps. Firstly, (i) nitrite is reduced to nitric oxide, which (ii) reacts with ammonium to form hydrazine, and (iii) is then oxidized to nitrogen gas [30]. Anammox bacteria can also use ferrous and ferric iron, manganese, carboxylic acids (i.e., methanoic, ethanoic, and propanoic acid), nitrite, and nitrate as TEAs [30]. The use of nitrate is of interest because, during the denitrification process, NO3− is converted to nitrogen gas via quite a different route. Nitrate is firstly reduced to nitrite, which can be combined with ammonium to form N2 through the anammox mechanism; therefore, anammox bacteria can disguise themselves as denitrifying bacteria [3]. Metabolic versatility leads to bacterial survival in conditions that lack ammonium or nitrite [31].
However, a high concentration of N–NO3− (i.e., above 150 mg·L−1) can inhibit the anammox process [27]. The temperature of the process should be between 20 and 40 °C, with an optimal temperature of 35 °C. The process can be performed at lower (2.5 °C) or higher (up to 70 °C) temperatures in marine and hot environments (e.g., hot springs and hydrothermal areas), respectively. Moreover, it has been found that Candidatus Scalindua performances were successfully enhanced in a batch reactor using marine sediments at 15–20 °C. The pH range should be 6.5–8.3 [32]. The anammox process occurs under anoxic conditions, so an oxygen concentration above 0.016 mg·L−1 could inhibit the process. Although anammox bacteria develop in fresh water and saline environments, a salinity greater than 30 g L−1 should be avoided [33,34].
All anammox organisms belong to the same monophyletic cluster named Brocadiales and are related to the order of Planctomycetales. Brocadia anammoxidas was the first anammox bacterium microbiologically identified with the name Candidatus [24,35]. In general, five genera of anammox bacteria are recognized [3]. Four Candidatus anammox genera were enriched from activated sludge (Kuenenia, Brocadia, Anammoxoglobus, and Jettenia). The fifth anammox genus (Candidatus Scalindua) was detected in natural habitats, such as areas with marine deposits with low oxygen concentrations [3,24,36].
Anammox bacteria appear as small and irregular coccoid cells with a diameter of approximately 1 µm under the microscope [35]. A specific red color also characterizes these bacteria due to the heme C-group of the protein, which plays a vital role in their metabolism [31]. C-type cytochromes are involved in a variety of functions, such as electron transfer (cytochrome C), oxygen transport and storage (hemoglobin and myoglobin), catalysis (e.g., peroxidases, catalase, hydroxylamine oxidoreductase), gas sensing, and gene regulation. However, the most prevalent function of C-type cytochromes is electron conveyance, which occurs in a wide variety of energy transduction processes, including photosynthesis, various respiration pathways, and processes involved in the nitrogen and sulfur cycles.
Cells double only once per 11–20 d and reduce their doubling time in situ even further, and they are surrounded by a thin cell wall that connects to an outer (i.e., cytoplasmic) membrane. Moreover, cells contain a second (i.e., intracytoplasmic) and a third membrane, the latter surrounding a central vacuole (the anammoxosome). Furthermore, ladderane lipids are unique to anammox bacteria, but the function of the anammoxosome and the role of the ladderane lipids need further experimentation [3,32]. Anammox bacteria are considered chemolithotrophic organisms, and thus the chemiosmotic processes of anammox bacteria take place in the anammoxosome, similar to the mitochondria in eukaryotes. This implies that a chemiosmotic mechanism is the only way to synthesize ATP. Therefore, protons are translocated across a closed semipermeable membrane system comparable to conventional membranes (i.e., up to 1.5 and 1.0 kg·L−1) and connected through electron transfer processes in the central catabolism, thus establishing a proton motive force across the cell membrane [37]. The proton motive force is composed of a chemical gradient of protons (∆pH) and an electrical charge gradient (∆Ψ). The proton motive force drives ATP synthesis through the action of a membrane-bound proton-translocating enzyme complex: the F1 F0 ATP synthetase (ATPase). Moreover, the anammoxosome membrane contributes to the low energy dissipation of the anammox bacterium due to electron propagation outside the membrane and other substances, such as nitric oxide and hydrazine. This contributes to the limited growth of anammox bacteria [27,30,35].
Anammox bacteria can develop in different natural environments. Microbial activity was detected in the Black Sea and areas with low oxygen concentrations, such as Bengal and Peru. In other studies, anammox bacteria were detected in anoxic or suboxic zones of marine and freshwater ecosystems. Anammox bacteria have also been found in extreme saline-related environments [34,36,38,39]. The factors affecting anammox bacterial diversity and distribution have been recently investigated in habitat-specific studies—for example, organic carbon influences anammox diversity in freshwater habitats, estuary marine sediments, and soil. Ammonium and nitrite concentrations are also correlated with anammox diversity in mangrove sediments. The temperature can impact anammox communities in freshwater and estuary sediments. Depth affects anammox diversity in marine sediments [34,38,40].
4. Insights Regarding Anoxic PAH Degradation during the Anammox Process
Coastal areas, estuaries, mangrove soils, and sediments can be identified as the main locations with high eutrophication risk due to improper discharge of inorganic nutrients (e.g., nitrogen), which can lead to uncontrolled algal growth and greenhouse gas emissions (i.e., N2O), contributing to global warming and ozone depletion [41]. In this context, the indigenous microbial community plays a critical role in maintaining the correct balance of nutrients in the ecosystem through biological processes (Figure 3a,b), thus limiting the abovementioned environmental issues [42].
However, pollutants such as PAHs can compromise microbial metabolic functions in these ecosystems, such as sediments, by easily penetrating the cell membranes of microorganisms due to their lipophilicity [43]. Indeed, nitrate-reducing bacteria can lead to nitrite accumulation through the use of PAHs (e.g., phenanthrene) as electron donors under anoxic conditions (Equation (1)) instead of completing denitrification (Figure 3a) due to partial inhibition [16,44]. In this context, anammox bacteria, which are autotrophic bacteria detected in various matrices (e.g., wastewater and marine sediments) [45] and involved in ammonia oxidation to N2 (Figure 3a) through the conversion of inorganic carbon with nitrite as a TEA (Equation (2)) in hydrazine synthesis, can play a role during PAH biodegradation in the presence of nitrate (Equation (2)) [46,47].
Therefore, the injection of anammox sludge under denitrifying conditions can limit nitrite accumulation while degrading PAHs. On the other hand, anammox bacteria require ammonia nitrogen to perform their metabolic activities (Equation (2)). About environmental compartments such as mangrove ecosystems, continuous flooding can lead to alternating aerobic and anaerobic conditions, thus allowing the occurrence of nitrification, denitrification, and anammox (Figure 3a) [10]. Therefore, a good strategy may be regulating the aeration amount and mixing ratio during biological treatments [48] when PAH remediation is conducted in a controlled environment (e.g., ex–situ remediation). On the other hand, the employment of this approach can be complex with in situ treatment methods.
Therefore, ammonia nitrogen production can be induced by the dissimilatory nitrate reduction to ammonium (DNRA) process (Figure 3), as nitrate is used as a TEA for cellular metabolism in the denitrification process and the DNRA process [44,49]. DNRA can reduce nitrate to ammonium under anoxic conditions through the nrfA gene, while PAHs act as electron donors (Equation (3)) [41,50].
Thus, nitrate reduction coupled with an increase in the ammonia level should be a signal for the correct functioning of PAH biodegradation under anoxic conditions [51]. Furthermore, if a partial DNRA is considered, only the first step of the DNRA process takes place (Equation (3)) [52], and then the removal reaction can be written as follows (Equation (4)):
A possible drawback of DNRA is that N2O gas can be produced as an intermediate during nitrate reduction at high pH values [46]. However, the presence of PAHs can promote the conversion of N2O to nitrogen gas (Equation (5)), assuming there is no cell growth and that PAHs are fully converted to CO2 [53]:
Hence, various approaches can be used as a link between the N-cycle and PAH removal (Figure 3a,b). Furthermore, it should be remembered that more bioavailable forms of nitrogen (i.e., NH4+) are generally employed as amendments to improve the capacity of the bacterial community to metabolize PAHs (Equation (6)) by balancing the carbon-to-nitrogen (C/N) ratio:
This would suggest that, in the presence of PAH-degrading bacteria, an equilibrium with anammox bacteria should be achieved during DNRA to avoid ammonia competition. Furthermore, the growth of anammox bacteria is slow, and optimal retention of anammox biomass is critical for applying anammox in various wastewater treatment plants, especially at low temperatures [54]. Research on lab-scale and full-scale applications of anammox achieved high biomass retention using moving bed biofilm reactors (MBBRs) in which anammox cells were immobilized onto supporting carriers. MBBRs have been employed in previous studies for synthetic wastewater due to their high biomass content, prevention of biomass leaching, and lower toxicity towards microorganisms [55]. MBBR systems successfully maintained high concentrations of anammox bacteria and provided operating flexibility [54]. Other issues in anammox processes that can be fixed with MBBRs are the drawbacks during the start-up phase [56] and competition between anammox bacteria and nitrite-oxidizing bacteria, the latter of which can catalyze the second step of nitrification and grow faster than anammox bacteria [57].
5. Biodegradation Efficiencies of PAHs and Their Impact on Nitrogen Removal
The main studies and results referring to PAH biodegradation in the presence of anammox bacteria or systems are summarized in Table 1. Phenanthrene was the most commonly examined PAH in the scientific studies (Table 1) due to its average solubility and lipophilicity compared to other PAHs [22] and the fact that it contains both K-region and bay-region structures, which are typical of high-molecular-weight (HMW) PAHs [58]. Phenanthrene degradation was investigated alone or in the presence of other PAHs by considering several contaminated matrices (i.e., fresh medium, sediments, soil, and synthetic wastewater) (Table 1). Other studies investigated the effects of anammox systems in groundwater, constructed wetlands, and slurry (i.e., soil + water) polluted by naphthalene, pyrene, or benzo[a]pyrene (Table 1) alone due to these ecosystems’ high water solubility [59], soil abundance [60], and toxicity [22], respectively.

Table 1.
Summary of studies reporting polycyclic aromatic hydrocarbon (PAH) degradation coupled with anammox bacteria or processes. LMW = low molecular weight; HMW = high molecular weight; TN = total nitrogen.
Naphthalene, phenanthrene, fluoranthene, pyrene, and benzo[a]pyrene showed removal efficiencies of 7.94–95.54, 5–99.63, 30–50, >99.99, and 30–54% (Table 1), respectively. Low-molecular-weight (LMW) PAHs can show higher removal efficiencies, likely due to their less complicated chemical properties and greater bioavailability compared to HMW PAHs (Figure 4 and Table 1) [65]. For LMW PAHs (i.e., naphthalene and phenanthrene), abiotic degradation can be more affected by volatilization and photodegradation mechanisms compared to HMW PAHs (e.g., fluoranthene and benzo[a]pyrene) [66]. Ribeiro et al. [51] obtained a higher abiotic loss for fluoranthene than naphthalene (10 and 15%, respectively)—reaching about 30 and 35% of biodegradation, respectively—when coupled with nitrate reduction and DNRA metabolism in sediments. This was not valid for pyrene, which showed the highest removal efficiency (above 99.99%; Table 1) when coupled with an anammox bacterial group (i.e., Planctomycetales strains). Therefore, other factors can influence PAH removal.
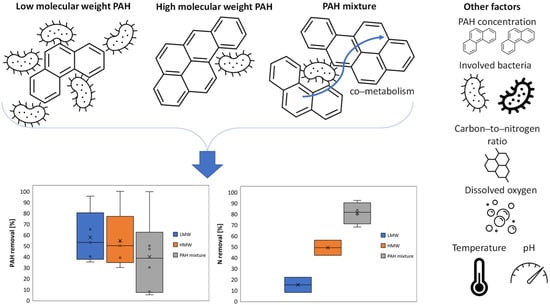
Figure 4.
Factors affecting polycyclic aromatic hydrocarbon (PAH) degradation and total nitrogen (TN) removal. The middle lines and both ends in the boxes represent the medians and 25th and 75th percentiles of PAH and N removal. The lower and upper whiskers are the minimum and maximum values of PAH and N removal. The inner points display the data points that lie between the lower whisker line and the upper whisker line. The cross represents the mean marker of the selected series.
Phenanthrene degradation increased from 53 to 99.63% (Table 1) alone and in the presence of naphthalene, respectively. This was most likely due to the co-metabolic effects induced by the co-occurrence of PAHs (Figure 4), where a more complex PAH can be degraded without being used as an energy source by bacteria while metabolizing a simpler PAH [67]. Furthermore, the half-lives (t1/2) of phenanthrene were significantly reduced from 6.37 to 0.19 d−1 (Table 1) in soil and synthetic wastewater, respectively. The use of synthetic solutions to simulate real polluted wastewater could not have affected the results since lower phenanthrene degradation was still achieved in both PAH-fortified soil and fresh medium (Table 1), thus affecting the mean and median values for the PAH mixture in Figure 4. One reason may be that pollutant degradation is more feasible when operating parameters (e.g., pH, temperature, lack of nutrients) are controlled in bioreactors, such as sequencing batch reactors (SBRs), compared to field conditions [68]. Different initial phenanthrene concentrations and greater PAH bioavailability when dissolved than when adsorbed into soils may be further factors affecting the gene copies of anammox that regulate the nitrogen cycle [61]. Finally, the microbial community involved during the biological process can affect PAH degradation (Figure 3b).
These studies (Table 1 and Figure 4) suggest that total nitrogen (TN) can be biodegraded at a rate of up to 92.46% by anammox bacteria in the presence of a PAH mixture through the reactions reported above (Equations (1)–(3)) in which PAHs can act as electron donors (Figure 3a). Proteobacteria, commonly found in anammox processes, can play a principal role in effective PAH biodegradation [19,46]. Among anammox bacteria, Candidatus Kuenenia and Parcubacteria (Figure 4) can boost PAH and nitrogen removal (Table 1) due to a further reaction in which they reduce NO to N2 [53]. In addition, these anammox bacteria can partially use the generated NO2− (Equation (2)) to return a fraction of the NO3− for PAH-degrading and nitrate-reducing bacteria [28]. Well-known anammox bacteria (i.e., Planctomycetales) can be successfully enriched when dissolved oxygen is maintained below 0.5 mg∙L−1 (Figure 4) [69]. A high organic load can also improve nitrate reduction by increasing the availability of nitrite for anammox bacteria [70]. The abundance of Candidatus Jettenia, which was associated with the highest nitrogen removal (Table 1), can be stimulated with calcium nitrate supplementation and a proper C/N ratio (i.e., above 6; Figure 4) [71,72]. Further operating parameters that can enhance the anammox process are pH levels and temperatures of 6.7–8.3 and 30–35 °C (Figure 4) [55], respectively, which are also suitable for anaerobic PAH metabolization (Equation (6)) [22].
With regard to the initial phase of anaerobic phenanthrene degradation (Equation (1)), carboxylation and methylation have been reported as essential reactions in which enzymes (i.e., ubiD-like carboxylase and ubiE-like methyltransferase) can be involved under nitrate-reducing conditions (Figure 3b) [28]. After this phase, a ring-reduction phase producing intermediates of reactions (e.g., dihydro-, tetrahydro-, hexahydro-, and octahydro-2-phenanthroic acids) can be followed by ring cleavage, which results in additional products, such as p-Cresol and p-hydroxybenzoic acid, before transformation into carbon dioxide (Figure 3b) [19].
The mentioned PAH metabolization pathway likely occurs due to the more negative standard Gibbs free energy change (ΔG0′) obtained with Equation (1) compared to Equation (6) (i.e., –163.2 and –62.8 kJ∙mol−1, respectively) [73], which can also follow the salicylate route. The ΔG0′ obtained with Equation (1) should be added to the ΔG0′ obtained during the anammox process (i.e., −357.8 kJ∙mol−1) [52] by accounting for a total ΔG0′ of –521 kJ∙mol−1. Coupling of partial DNRA and anammox activity (Equation (4)) can lead to a further decrease in ΔG0′ [52], making PAH and nitrogen removal even more feasible. In each case, this latter removal mechanism can be involved in the late stage, where DNRA can play a role with a high C/N ratio [29].
6. Conclusions and Future Directions
This article provides a comprehensive overview of PAH biodegradation in anammox systems by highlighting various critical aspects, such as mechanisms, PAH removal efficiencies, nitrogen biogeochemistry, and the involved microbial community. A high PAH removal efficiency (i.e., above 99.99%) can be achieved in anammox processes due to the nitrate reduction and DNRA mechanisms, in which a single PAH or a mixture can serve as the electron donor(s) and be metabolized through carboxylation and methylation. The produced nitrite and ammonia nitrogen can be successfully converted at an efficiency of up to 92.46% by anammox bacteria (e.g., Candidatus Jettenia, Candidatus Kuenenia, Parcubacteria). The abundance of these bacteria can be improved by regulating operating parameters, such as the dissolved oxygen and C/N ratio (<0.5 mg∙L−1 and >6, respectively). Future work should be aimed at investigating the use of continuous-flow bioreactors for enhancing nitrogen removal in real contaminated wastewater. The employment of MBBRs should help facilitate anammox biomass growth during the start-up phase in full-scale applications and avoid the drawbacks due to bacterial competition. Additional investigations should focus on: (i) verifying the actual role of anammox in the PAH biodegradation process; (ii) exploring the synergy between anammox bacteria and PAH-degrading bacteria since knowledge of the microbial population and metabolic and genetic capabilities is essential to establish optimal operating conditions; (iii) identifying the microbial communities capable of effectively removing HMW PAHs; (iv) investigating other PAH biodegradation pathways; and (v) performing further tests on naturally contaminated matrices with variable ratios of pollutants.
Author Contributions
Conceptualization, F.B., A.A.S.A.-G. and M.R.; methodology, F.B.; software, F.B.; data curation, F.B.; writing—original draft preparation, F.B.; writing—review and editing, F.B., A.A.S.A.-G. and M.R.; supervision, M.R.; project administration, M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pereira, A.D.; Cabezas, A.; Etchebehere, C.; Chernicharo, C.A.D.L.; de Araújo, J.C. Microbial Communities in Anammox Reactors: A Review. Environ. Technol. Rev. 2017, 6, 74–93. [Google Scholar] [CrossRef]
- Thamdrup, B. New Pathways and Processes in the Global Nitrogen Cycle. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 407–428. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Van Niftrik, L.; Strous, M.; Kartal, B.; Keltjens, J.T.; Op Den Camp, H.J.M. Biochemistry and Molecular Biology of Anammox Bacteria. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 65–84. [Google Scholar] [CrossRef]
- Wang, S.; Peng, Y.; Ma, B.; Wang, S.; Zhu, G. Anaerobic Ammonium Oxidation in Traditional Municipal Wastewater Treatment Plants with Low-Strength Ammonium Loading: Widespread but Overlooked. Water Res. 2015, 84, 66–75. [Google Scholar] [CrossRef]
- Du, R.; Cao, S.; Li, B.; Wang, S.; Peng, Y. Simultaneous Domestic Wastewater and Nitrate Sewage Treatment by DEnitrifying AMmonium OXidation (DEAMOX) in Sequencing Batch Reactor. Chemosphere 2017, 174, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Khin, T.; Annachhatre, A.P. Novel Microbial Nitrogen Removal Processes. Biotechnol. Adv. 2004, 22, 519–532. [Google Scholar] [CrossRef]
- Liu, S.; Yang, F.; Meng, F.; Chen, H.; Gong, Z. Enhanced Anammox Consortium Activity for Nitrogen Removal: Impacts of Static Magnetic Field. J. Biotechnol. 2008, 138, 96–102. [Google Scholar] [CrossRef]
- Monballiu, A.; Desmidt, E.; Ghyselbrecht, K.; De Clippeleir, H.; Van Hulle, S.W.H.; Verstraete, W.; Meesschaert, B. Enrichment of Anaerobic Ammonium Oxidizing (Anammox) Bacteria from OLAND and Conventional Sludge: Features and Limitations. Sep. Purif. Technol. 2013, 104, 130–137. [Google Scholar] [CrossRef]
- Kampschreur, M.J.; Poldermans, R.; Kleerebezem, R.; Van Der Star, W.R.L.; Haarhuis, R.; Abma, W.R.; Jetten, M.S.M.; Van Loosdrecht, M.C.M. Emission of Nitrous Oxide and Nitric Oxide from a Full-Scale Single-Stage Nitritation-Anammox Reactor. Water Sci. Technol. 2009, 60, 3211–3217. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, H.C.; Pan, Y.; Shyla, F.S.; Tam, N.F.Y. Effects of Polybrominated Diphenyl Ethers and Plant Species on Nitrification, Denitrification and Anammox in Mangrove Soils. Sci. Total Environ. 2016, 553, 60–70. [Google Scholar] [CrossRef]
- Lawal, A.T. Polycyclic Aromatic Hydrocarbons. A Review. Cogent Environ. Sci. 2017, 3, 1339841. [Google Scholar] [CrossRef]
- Soclo, H.H.; Garrigues, P.; Ewald, M. Origin of Polycyclic Aromatic Hydrocarbons (PAHs) in Coastal Marine Sediments: Case Studies in Cotonou (Benin) and Aquitaine (France) Areas. Mar. Pollut. Bull. 2000, 40, 387–396. [Google Scholar] [CrossRef]
- Munoz, B.; Albores, A. DNA Damage Caused by Polycyclic Aromatic Hydrocarbons: Mechanisms and Markers. In Selected Topics in DNA Repair; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Habe, H.; Omori, T. Genetics of Polycyclic Aromatic Hydrocarbon Metabolism in Diverse Aerobic Bacteria. Biosci. Biotechnol. Biochem. 2003, 67, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Ambrosoli, R.; Petruzzelli, L.; Minati, J.L.; Marsan, F.A. Anaerobic PAH Degradation in Soil by a Mixed Bacterial Consortium under Denitrifying Conditions. Chemosphere 2005, 60, 1231–1236. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; He, Z.; Zhang, Q.; Liu, J.; Guo, J.; Sun, G.; Zhou, J. Responses of Aromatic-Degrading Microbial Communities to Elevated Nitrate in Sediments. Environ. Sci. Technol. 2015, 49, 12422–12431. [Google Scholar] [CrossRef]
- Duran, R.; Cravo-Laureau, C. Role of Environmental Factors and Microorganisms in Determining the Fate of Polycyclic Aromatic Hydrocarbons in the Marine Environment. FEMS Microbiol. Rev. 2016, 40, 814–830. [Google Scholar] [CrossRef]
- Semple, K.T.; Morriss, A.W.J.; Paton, G.I. Bioavailability of Hydrophobic Organic Contaminants in Soils: Fundamental Concepts and Techniques for Analysis. Eur. J. Soil Sci. 2003, 54, 809–818. [Google Scholar] [CrossRef]
- Bianco, F.; Race, M.; Papirio, S.; Oleszczuk, P.; Esposito, G. Coupling of Desorption of Phenanthrene from Marine Sediments and Biodegradation of the Sediment Washing Solution in a Novel Biochar Immobilized–Cell Reactor. Environ. Pollut. 2022, 308, 119621. [Google Scholar] [CrossRef]
- Bianco, F.; Marcińczyk, M.; Race, M.; Papirio, S.; Esposito, G.; Oleszczuk, P. Low Temperature-Produced and VFA-Coated Biochar Enhances Phenanthrene Adsorption and Mitigates Toxicity in Marine Sediments. Sep. Purif. Technol. 2022, 296, 121414. [Google Scholar] [CrossRef]
- Mu, J.; Chen, Y.; Song, Z.; Liu, M.; Zhu, B.; Tao, H.; Bao, M.; Chen, Q. Effect of Terminal Electron Acceptors on the Anaerobic Biodegradation of PAHs in Marine Sediments. J. Hazard. Mater. 2022, 438, 129569. [Google Scholar] [CrossRef]
- Bianco, F.; Race, M.; Papirio, S.; Esposito, G. A Critical Review of the Remediation of PAH- Polluted Marine Sediments: Current Knowledge and Future Perspectives. Resour. Environ. Sustain. 2022, 11, 100101. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The Evolution and Future of Earth ’ s Nitrogen Cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef]
- Kartal, B.; Van Niftrik, L.; Rattray, J.; Van De Vossenberg, J.L.C.M.; Schmid, M.C.; Sinninghe Damstà ©, J.; Jetten, M.S.M.; Strous, M. Candidatus ‘Brocadia fulgida’: An autofluorescent anaerobic ammonium oxidizing bacterium. FEMS Microbiol. Ecol. 2008, 63, 46–55. [Google Scholar] [CrossRef]
- Francis, C.A.; Beman, J.M.; Kuypers, M.M.M. New Processes and Players in the Nitrogen Cycle: The Microbial Ecology of Anaerobic and Archaeal Ammonia Oxidation. ISME J. 2007, 1, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G. Earth’ s Biogeochemical Cycles. Science 2012, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Huy Quoc Anh, D.; Tantayotai, P.; Cheenkachorn, K.; Sriariyanun, M. Anammox Process: The Principle, the Technological Development and Recent Industrial Applications. Appl. Sci. Eng. Prog. 2015, 8, 237–244. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, H.; Sun, J.; Wang, H. Investigation of Anaerobic Phenanthrene Biodegradation by a Highly Enriched Co-Culture, PheN9, with Nitrate as an Electron Acceptor. J. Hazard. Mater. 2020, 383, 121191. [Google Scholar] [CrossRef]
- Urakawa, H.; Rajan, S.; Feeney, M.E.; Sobecky, P.A.; Mortazavi, B. Ecological Response of Nitrification to Oil Spills and Its Impact on the Nitrogen Cycle. Environ. Microbiol. 2019, 21, 18–33. [Google Scholar] [CrossRef]
- Kartal, B.; Maalcke, W.J.; De Almeida, N.M.; Cirpus, I.; Gloerich, J.; Geerts, W.; Op Den Camp, H.J.M.; Harhangi, H.R.; Janssen-Megens, E.M.; Francoijs, K.J.; et al. Molecular Mechanism of Anaerobic Ammonium Oxidation. Nature 2011, 479, 127–130. [Google Scholar] [CrossRef]
- Kartal, B.; Keltjens, J.T.; Jetten, M.S.M. Metabolism and Genomics of Anammox Bacteria. In Nitrification; ASM Press: Washington, DC, USA, 2014; pp. 179–200. [Google Scholar]
- Rattray, J.E.; van de Vossenberg, J.; Jaeschke, A.; Hopmans, E.C.; Wakeham, S.G.; Lavik, G.; Kuypers, M.M.M.; Strous, M.; Jetten, M.S.M.; Schouten, S.; et al. Impact of Temperature on Ladderane Lipid Distribution in Anammox Bacteria. Appl. Environ. Microbiol. 2010, 76, 1596–1603. [Google Scholar] [CrossRef]
- Liu, C.; Yamamoto, T.; Nishiyama, T.; Fujii, T.; Furukawa, K. Effect of Salt Concentration in Anammox Treatment Using Non Woven Biomass Carrier. J. Biosci. Bioeng. 2009, 107, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Sonthiphand, P.; Hall, M.W.; Neufeld, J.D. Biogeography of Anaerobic Ammonia-Oxidizing (Anammox) Bacteria. Front. Microbiol. 2014, 5, 1–15. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Strous, M.; van de Pas-Schoonen, K.T.; Schalk, J.; van Dongen, U.G.J.M.; van de Graaf, A.A.; Logemann, S.; Muyzer, G.; van Loosdrecht, M.C.M.; Kuenen, J.G. The Anaerobic Oxidation of Ammonium. FEMS Microbiol. Rev. 1998, 22, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Woebken, D.; Lam, P.; Kuypers, M.M.M.; Naqvi, S.W.A.; Kartal, B.; Strous, M.; Jetten, M.S.M.; Fuchs, B.M.; Amann, R. A Microdiversity Study of Anammox Bacteria Reveals a Novel Candidatus Scalindua Phylotype in Marine Oxygen Minimum Zones. Environ. Microbiol. 2008, 10, 3106–3119. [Google Scholar] [CrossRef]
- Van Niftrik, L.A.; Fuerst, J.A.; Sinninghe Damsté, J.S.; Kuenen, J.G.; Jetten, M.S.M.; Strous, M. The Anammoxosome: An Intracytoplasmic Compartment in Anammox Bacteria. FEMS Microbiol. Lett. 2004, 233, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Luan, X.; Dang, H.; Zhou, H.; Zhao, Y.; Liu, H.; Zhang, Y.; Dai, L.; Ye, Y.; Klotz, M.G. Deep-Sea Methane Seep Sediments in the Okhotsk Sea Sustain Diverse and Abundant Anammox Bacteria. FEMS Microbiol. Ecol. 2014, 87, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.L.; Shen, L.D.; Xu, X.Y.; Zheng, P. Anaerobic Ammonium Oxidation (Anammox) in Different Natural Ecosystems. Biochem. Soc. Trans. 2011, 39, 1811–1816. [Google Scholar] [CrossRef] [PubMed]
- Osaka, T.; Kimura, Y.; Otsubo, Y.; Suwa, Y.; Tsuneda, S.; Isaka, K. Temperature Dependence for Anammox Bacteria Enriched from Freshwater Sediments. J. Biosci. Bioeng. 2012, 114, 429–434. [Google Scholar] [CrossRef]
- Cao, W.; Yang, J.; Li, Y.; Liu, B.; Wang, F.; Chang, C. Dissimilatory Nitrate Reduction to Ammonium Conserves Nitrogen in Anthropogenically Affected Subtropical Mangrove Sediments in Southeast China. Mar. Pollut. Bull. 2016, 110, 155–161. [Google Scholar] [CrossRef]
- Sun, P.; Bai, J.; Li, K.; Zhao, Y.; Tian, W.; Bai, X.; Tian, Y. Impact of Phenanthrene on Denitrification Activity and Transcription of Related Functional Genes in Estuarine and Marine Sediments. J. Ocean Univ. China 2020, 19, 124–134. [Google Scholar] [CrossRef]
- Suszek-Łopatka, B.; Maliszewska-Kordybach, B.; Klimkowicz-Pawlas, A.; Smreczak, B. The Drought and High Wet Soil Condition Impact on PAH (Phenanthrene) Toxicity towards Nitrifying Bacteria. J. Hazard. Mater. 2019, 368, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhang, L.; Zhang, D.; Lu, P.; Zhang, X.; He, Q. Denitrification Synergized with ANAMMOX for the Anaerobic Degradation of Benzene: Performance and Microbial Community Structure. Appl. Microbiol. Biotechnol. 2017, 101, 4315–4325. [Google Scholar] [CrossRef]
- Ali, M.; Chai, L.-Y.; Tang, C.-J.; Zheng, P.; Min, X.-B.; Yang, Z.-H.; Xiong, L.; Song, Y.-X. The Increasing Interest of ANAMMOX Research in China: Bacteria, Process Development, and Application. Biomed Res. Int. 2013, 2013, 134914. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Niu, J.; Liu, Y.; Zhao, J.; Yin, Z. Effects of Polycyclic Aromatic Hydrocarbons on Sludge Performance for Denitrification and Phosphorus Removal. Chem. Eng. J. 2020, 397, 125552. [Google Scholar] [CrossRef]
- Ribeiro, H.; Mucha, A.P.; Azevedo, I.; Salgado, P.; Teixeira, C.; Almeida, C.M.R.; Joye, S.B.; Magalhães, C. Differential Effects of Crude Oil on Denitrification and Anammox, and the Impact on N2O Production. Environ. Pollut. 2016, 216, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Lu, H.; Zhang, J.; Zhu, S.; Wang, Y.; Lei, Y.; Zhang, R.; Song, L. Simultaneous Heterotrophic Nitrification and Aerobic Denitrification (SND) for Nitrogen Removal: A Review and Future Perspectives. Environ. Adv. 2022, 9, 100254. [Google Scholar] [CrossRef]
- Han, X.; Peng, S.; Zhang, L.; Lu, P.; Zhang, D. The Co-Occurrence of DNRA and Anammox during the Anaerobic Degradation of Benzene under Denitrification. Chemosphere 2020, 247, 125968. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, F.; Ma, Z.; Wan, Y. Nitrogen Biogeochemistry of Anaerobic Biodegradation of Naphthalene. Water Air Soil Pollut. 2019, 230, 222. [Google Scholar] [CrossRef]
- Ribeiro, H.; de Sousa, T.; Santos, J.P.; Sousa, A.G.G.; Teixeira, C.; Monteiro, M.R.; Salgado, P.; Mucha, A.P.; Almeida, C.M.R.; Torgo, L.; et al. Potential of Dissimilatory Nitrate Reduction Pathways in Polycyclic Aromatic Hydrocarbon Degradation. Chemosphere 2018, 199, 54–67. [Google Scholar] [CrossRef]
- Castro-Barros, C.M.; Jia, M.; van Loosdrecht, M.C.M.; Volcke, E.I.P.; Winkler, M.K.H. Evaluating the Potential for Dissimilatory Nitrate Reduction by Anammox Bacteria for Municipal Wastewater Treatment. Bioresour. Technol. 2017, 233, 363–372. [Google Scholar] [CrossRef]
- Zhang, D.; Han, X.; Zhou, S.; Yuan, S.; Lu, P.; Peng, S. Nitric Oxide-Dependent Biodegradation of Phenanthrene and Fluoranthene: The Co-Occurrence of Anaerobic and Intra-Aerobic Pathways. Sci. Total Environ. 2021, 760, 144032. [Google Scholar] [CrossRef] [PubMed]
- Trinh, H.P.; Lee, S.-H.; Jeong, G.; Yoon, H.; Park, H.-D. Recent Developments of the Mainstream Anammox Processes: Challenges and Opportunities. J. Environ. Chem. Eng. 2021, 9, 105583. [Google Scholar] [CrossRef]
- Cho, S.; Kambey, C.; Nguyen, V. Performance of Anammox Processes for Wastewater Treatment: A Critical Review on Effects of Operational Conditions and Environmental Stresses. Water 2019, 12, 20. [Google Scholar] [CrossRef]
- Xu, G.; Zhou, Y.; Yang, Q.; Lee, Z.M.-P.; Gu, J.; Lay, W.; Cao, Y.; Liu, Y. The Challenges of Mainstream Deammonification Process for Municipal Used Water Treatment. Appl. Microbiol. Biotechnol. 2015, 99, 2485–2490. [Google Scholar] [CrossRef]
- Laureni, M.; Weissbrodt, D.G.; Villez, K.; Robin, O.; de Jonge, N.; Rosenthal, A.; Wells, G.; Nielsen, J.L.; Morgenroth, E.; Joss, A. Biomass Segregation between Biofilm and Flocs Improves the Control of Nitrite-Oxidizing Bacteria in Mainstream Partial Nitritation and Anammox Processes. Water Res. 2019, 154, 104–116. [Google Scholar] [CrossRef]
- Agrawal, N.; Kumar, V.; Shahi, S.K. Biodegradation and Detoxification of Phenanthrene in in Vitro and in Vivo Conditions by a Newly Isolated Ligninolytic Fungus Coriolopsis Byrsina Strain APC5 and Characterization of Their Metabolites for Environmental Safety. Environ. Sci. Pollut. Res. 2022, 29, 61767–61782. [Google Scholar] [CrossRef]
- Kumar, A.; Gupta, H. Activated Carbon from Sawdust for Naphthalene Removal from Contaminated Water. Environ. Technol. Innov. 2020, 20, 101080. [Google Scholar] [CrossRef]
- Yu, C.C.; Chang, T.C.; Liao, C.S.; Chang, Y.T. A Comparison of the Microbial Community and Functional Genes Present in Free-Living and Soil Particle-Attached Bacteria from an Aerobic Bioslurry Reactor Treating High-Molecular-Weight PAHs. Sustainability 2019, 11, 1088. [Google Scholar] [CrossRef]
- Khandelwal, A.; Sugavanam, R.; Ramakrishnan, B.; Dutta, A.; Varghese, E.; Banerjee, T.; Nain, L.; Singh, S.B.; Singh, N. Bio-Polysaccharide Composites Mediated Degradation of Polyaromatic Hydrocarbons in a Sandy Soil Using Free and Immobilized Consortium of Kocuria Rosea and Aspergillus Sydowii. Environ. Sci. Pollut. Res. 2022, 29, 80005–80020. [Google Scholar] [CrossRef]
- Lu, J.; Guo, Z.; He, M.; Hu, Z.; Wu, H.; Zhuang, L.; Kong, Q.; Zhang, J. Highly Enhanced Removal of Nutrients and Benzo[a]Pyrene in a Siphon Constructed Wetland with Magnetite: Performance and Mechanisms. Chem. Eng. J. 2022, 446, 136895. [Google Scholar] [CrossRef]
- Liang, L.; Song, X.; Kong, J.; Shen, C.; Huang, T.; Hu, Z. Anaerobic Biodegradation of High-Molecular-Weight Polycyclic Aromatic Hydrocarbons by a Facultative Anaerobe Pseudomonas Sp. JP1. Biodegradation 2014, 25, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Weng, R.; He, Y.; Wang, J.; Zhang, Z.; Wei, Z.; Yang, Y.; Huang, M.; Zhou, G. Quantitative Characterization and Genetic Diversity Associated with N-Cycle Pathways in Urban Rivers with Different Remediation Techniques. Sci. Total Environ. 2022, 804, 150235. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, S.; Persson, Y.; Frankki, S.; van Bavel, B.; Lundstedt, S.; Haglund, P.; Tysklind, M. Degradation of Polycyclic Aromatic Hydrocarbons (PAHs) in Contaminated Soils by Fenton’s Reagent: A Multivariate Evaluation of the Importance of Soil Characteristics and PAH Properties. J. Hazard. Mater. 2007, 149, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.-R.; Cang, L.; Wang, Q.-Y.; Zhou, D.-M.; Cheng, J.-M.; Xu, H. Roles of Abiotic Losses, Microbes, Plant Roots, and Root Exudates on Phytoremediation of PAHs in a Barren Soil. J. Hazard. Mater. 2010, 176, 919–925. [Google Scholar] [CrossRef]
- Nzila, A. Update on the Cometabolism of Organic Pollutants by Bacteria. Environ. Pollut. 2013, 178, 474–482. [Google Scholar] [CrossRef]
- Struk-Sokołowska, J.; Kotowska, U.; Piekutin, J.; Laskowski, P.; Mielcarek, A. Analysis of 1H-Benzotriazole Removal Efficiency from Wastewater in Individual Process Phases of a Sequencing Batch Reactor SBR. Water Resour. Ind. 2022, 28, 100182. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, B.; Gong, X.; Liu, X.; Peng, Y. Anammox Bacteria Enrich Naturally in Suspended Sludge System during Partial Nitrification of Domestic Sewage and Contribute to Nitrogen Removal. Sci. Total Environ. 2021, 787, 147658. [Google Scholar] [CrossRef]
- Hu, B.; Shen, L.; Du, P.; Zheng, P.; Xu, X.; Zeng, J. The Influence of Intense Chemical Pollution on the Community Composition, Diversity and Abundance of Anammox Bacteria in the Jiaojiang Estuary (China). PLoS ONE 2012, 7, e33826. [Google Scholar] [CrossRef]
- Zhi, W.; Ji, G. Quantitative Response Relationships between Nitrogen Transformation Rates and Nitrogen Functional Genes in a Tidal Flow Constructed Wetland under C/N Ratio Constraints. Water Res. 2014, 64, 32–41. [Google Scholar] [CrossRef]
- Sheng, H.; Weng, R.; Zhu, J.; He, Y.; Cao, C.; Huang, M. Calcium Nitrate as a Bio-Stimulant for Anaerobic Ammonium Oxidation Process. Sci. Total Environ. 2021, 760, 143331. [Google Scholar] [CrossRef]
- Dhar, K.; Subashchandrabose, S.R.; Venkateswarlu, K.; Krishnan, K.; Megharaj, M. Anaerobic Microbial Degradation of Polycyclic Aromatic Hydrocarbons: A Comprehensive Review. In Reviews of Environmental Contamination and Toxicology; Springer: New York, NY, USA, 2019; Volume 251, pp. 25–108. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).