Abstract
Coronaviruses have caused devastation in both human and animal populations, affecting both health and the economy. Amidst the emergence and re-emergence of coronaviruses, humans need to surmount the health and economic threat of coronaviruses through science and evidence-based approaches. One of these approaches is through biotechnology, particularly the heterologous production of biopharmaceutical proteins. This review article briefly describes the genome, general virion morphology, and key structural proteins of different coronaviruses affecting animals and humans. In addition, this review paper also presents the different systems in recombinant protein technology such as bacteria, yeasts, plants, mammalian cells, and insect/insect cells systems used to express key structural proteins in the development of countermeasures such as diagnostics, prophylaxis, and therapeutics in the challenging era of coronaviruses.
1. Introduction
Coronaviruses or CoVs are enveloped, positive-sensed, single-stranded RNA viruses belonging to the order Nidovirales, family Coronaviridae, and subfamily Coronavirinae. Coronaviruses were originally grouped based on serology but are now classified based on phylogenetic relationships. This system of classification divided Coronavirinae into four genera, namely, alpha-, beta-, gamma-, and deltacoronavirus [1]. Generally, alpha- and betacoronaviruses are associated with humans and other mammals, while gamma- and deltacoronaviruses are linked to avians and, to some extent, marine mammals [2]. Their mode of transmission is usually through respiratory droplets, which are particles less than 5–10 μm in size, or alternatively, through fecal–oral routes. However, emerging evidence shows that airborne transmission or inhalation of droplet nuclei (aerosols) that may remain suspended in the air for some time, especially in occluded spaces, is possible [3]. Coronaviruses may not cause apparent infection in their original vertebrate hosts, but once they jump to livestock and humans from wildlife, they may cause mild to severe infection, even reaching epidemic and pandemic proportions if unabated [4,5]. Usually, coronaviruses cause mild to severe respiratory infection, kidney failure, enteric, including hepatic disease, and to some extent, neurological involvement depending on the type of coronavirus and animal host.
The world has seen the devastation of different coronaviruses in agriculture and human populations with grave implications for health systems, the food chain, and the economies of countries affected. The very first known coronavirus that was described and isolated in the 1930s is the infectious bronchitis virus (IBV) [6]. Infectious bronchitis virus belongs to the Genus Gammacoronavirus, which primarily infects the respiratory tract of chickens but also shows tissue tropism for other organs such as the kidney, oviducts, testes, and the alimentary tract [7]. Another avian coronavirus is the turkey coronavirus (TCoV) which also belongs to the Genus Gammacoronavirus and can cause gastrointestinal disease in both young poults and adult turkeys [8]. In turkey poults, TCoV may cause mortality, while it can cause stunting and underperformance in terms of meat and egg yield in adult birds [9]. Both IBV and TCoV are imminent threats to the poultry industry worldwide, which captured the attention of scientists early on due to the economic importance of the poultry industry, as reviewed by Cavanagh [9]. Another coronavirus that has devastated the agriculture sector is the porcine epidemic diarrhea virus (PEDV). The first outbreak of PEDV was reported in England in 1971 among growing and fattening pigs that manifested watery diarrhea [10]. Pensaert and de Bouck (1978) reported that a new coronavirus-like particle was detected in the intestinal contents of pigs during an outbreak of diarrhea in four swine breeding farms in Belgium [11]. Their team replicated Koch’s postulate and designated the novel coronavirus as PEDV CV777, which was unique from the other two coronaviruses, transmissible gastroenteritis virus (TGEV) and porcine hemagglutinating encephalomyelitis virus (PHEV), that also infect hogs [12,13]. Although it does not cause diseases in agricultural and livestock animals, murine hepatitis coronavirus (MHCoV) is of interest to scientists since it can be used as a model to study hepatitis, demyelinating diseases such as multiple sclerosis in humans, and possibly other infections caused by other coronaviruses [14].
Humans have not been spared from the ravages of coronaviruses. To date, seven coronaviruses have been found to circulate within the human population. Of these, four of the seven human coronaviruses (HCoV), namely 229E, NL63, OC43, and HKU1, cause annual mild to moderate upper respiratory tract illnesses, as reviewed elsewhere by Hulse [15]. However, the other three remaining known HCoV have grievously affected humans in recent history, and one is still occurring as of the writing of this review. These HCoVs are the severe acute respiratory syndrome coronavirus (SARS-CoV), Middle East respiratory syndrome coronavirus (MERS-CoV), and severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), all of which cause severe and life-threatening respiratory tract infections, which sometimes lead to the death of the infected individuals [16]. The 2002–2003 SARS epidemic originated in Guangzhou, Guangdong Province, People’s Republic of China, and spread to Hong Kong, Singapore, Taiwan, and Canada, wherein the main suspected animal reservoir host was a civet cat (Paguma larvata) [17]. Another coronavirus that caused an epidemic within this decade is the MERS-CoV, which originated in Saudi Arabia [18]. The 2012 MERS-CoV most probably jumped from bats and dromedary camels to humans [19] and affected several countries in the Middle East, Southeast Asia, East Asia, Europe, North Africa, and North America [20]. Lastly, the most damaging of all the infections caused by a coronavirus in human history is Coronavirus Disease-19, or COVID-19, which has already infected around 328 million people and killed more than 5.5 million people worldwide (as of 19 January 2022). Coronavirus Disease-19 is caused by a novel coronavirus, originally named 2019-nCoV but renamed SARS-CoV-2, which originated in the city of Wuhan, in Hubei province, central China [21,22]. The prime suspected animal reservoirs of SARS-CoV-2 are horseshoe bats (Rhinolopus affinis) or Malayan pangolins (Manis javanica) [23]. Table 1 shows the list of coronaviruses, their animal hosts and host receptors, and the diseases they cause in their host.

Table 1.
Veterinary and medically important coronaviruses, their hosts and host receptors, and diseases they cause.
Countries around the world are implementing non-pharmaceutical control measures such as border restrictions and travel bans, community lockdowns, mass testing, contact tracing, isolating and quarantining exposed and suspected individuals, wearing face masks, respiratory etiquette, hand hygiene, and social distancing in their efforts to control the transmission of SARS-CoV-2 [41]. Additionally, pharmacotherapies such as Remdesivir, Hydroxychloroquine, Lopinavir, and Interferon were also tested under the WHO SOLIDARITY clinical trials, but these regimens showed little or no effect on hospitalized COVID-19 patients [42]. Only recently, the US FDA granted Emergency Use Authorization (EUA) to two oral antiviral agents against COVID-19, Molnupiravir and Paxlovid, which were found to reduce the risk of COVID-19-related hospitalization by 50% and 89%, respectively [43,44]. While the non-pharmaceutical strategies are effective in preventing transmission and antiviral drugs may reduce hospitalization and death among infected individuals, the only way to avert the further health and socioeconomic impacts of the COVID-19 pandemic and establish long-term prevention and control measures is to develop a vaccine and a safe and effective vaccination program [45]. As of 10 December 2020, 214 vaccine candidates were under development for COVID-19 [46]. Of these, 52 were in clinical trials, while the remaining 162 were in pre-clinical evaluation. Several vaccine candidates have completed their Phase III clinical trial, and their manufacturers have obtained Emergency Use Authorization (EUA) from regulatory authorities. Vaccine candidates for coronaviruses and other viral pathogens are developed based on different techniques. One of the strategies in vaccine development against coronaviruses is the use of protein subunits, which may be composed of just one type of viral antigen or structural proteins that have the ability to assemble into repeated arrays, also known as virus-like particles (VLPs), produced through recombinant protein technology in various culture systems. Recombinant protein subunit vaccines are produced by the heterologous expression of an epitope-carrying immunogen protein subunit of a target pathogen in a competent host [47], while VLPs are protein subunits that mimic the structure of authentic virus particles that can be presented to the immune system in a more native conformation in the absence of infectious genetic material [48,49]. The use of recombinant protein technology has expanded beyond therapeutic proteins to vaccine engineering and the development of diagnostic platforms for different diseases due to the flexibility of its expression system and proven safety record. This review briefly discusses the key structural proteins of coronaviruses and highlights the different expression systems currently used to produce these recombinant key structural proteins of coronaviruses and their various applications in the rational design of prophylactic vaccines, diagnostics, and therapeutics.
2. Coronaviruses: Genome, Virion Morphology, and Key Structural Proteins
Understanding the genomic make-up and the major key structural proteins of coronaviruses is fundamental to the rational design of any measures against the virus, such as vaccines, diagnostic platforms, and antiviral agents. In this section, the genome and basic structural architecture of coronaviruses will be briefly described. Coronaviruses make up a large family of non-segmented, positive-sense, single-stranded RNA viruses (++ssRNA) [50]. This group of viruses possesses the largest genomes among all known RNA viruses, ranging from 26 kb to 32 kb [51]. Having such large genomes increases their plasticity for genetic accommodation and modification. Their genomes share a commonality, wherein both 5′ and 3′ ends contain short untranslated regions. The typical arrangement of the genome from 5′-3′ starts with ORF1ab occupying a third of the genome, which encodes for the viral replicase polyproteins made up of Papain-like protease 1 and 2 (PL1 and PL2), chymotrypsin-like protease (3CL), RNA-dependent RNA polymerase (RdRp), helicase (Hel), and hemagglutinin esterase (HE) found in subgroup A of Betacoronavirus that is believed to have been acquired from Influenza C virus through heterologous recombination. The downstream portion of the genome encodes for the coronaviruses’ key structural proteins such as spike (S), envelope (E), membrane (M), and nucleocapsid (N), respectively [52,53,54] (See Figure 1A,B).
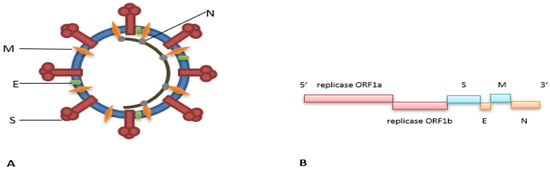
Figure 1.
Schematic diagram of a coronavirus’ (A) virion and (B) genome.
2.1. Key Structural Proteins of Coronaviruses
Spike (S) Protein
S glycoprotein, or simply spike protein, is the protein protruding from the surface of coronaviruses, giving them the name-sake corona or sun-like appearance under an electron microscope. These spikes have a club-like shape with a stalk and rough globular head [55]. The S glycoprotein is a homotrimer with an N terminus that binds to varying host cell receptors (Table 1) and is crucial for viral entry into the cell [40]. Additionally, spike proteins have a short transmembrane domain at the C terminus followed by a short cytoplasmic tail that is cysteine-rich, which interacts with the virion membrane [56]. The S protein is cleaved into S1 and S2 domains at a consensus cleavage site, wherein S1 domain sequences are much more variable than S2 domain sequences. The S1 domain functions as the receptor-binding domain (RBD), while S2 acts as the membrane fusion domain [57]. Millet et al. (2020) and Li et al. (2015) wrote comprehensive reviews on the diversity and receptor recognition mechanisms of different coronaviruses [58,59]. Aside from the spike protein’s function in cell entry and fusion, it plays an important role in tissue tropism, host specificity [60], and the induction of immune response [61]. Moreover, recent evidence shows that S proteins may also play a role in host jumping or cross-species transmission [62].
2.2. Membrane (M) Protein
Membrane, or M, glycoprotein is the most abundant of all the structural proteins [63]. It is an important component of the virus, which maintains the overall morphology and assembly of the virion [64]. Membrane proteins contain three transmembrane segments. The ectodomain of the M protein is the least conserved portion that undergoes glycosylation in the Golgi body [64,65,66]. The post-translational modification of the M protein is important for the fusion of the virion to its target cell and in the protein’s antigenicity [67,68]. The M protein also interacts with nucleocapsid (N) protein to envelop the viral RNA genome [69]. There may be variations in M proteins across different coronaviruses, but the structural similarity is generally the same or moderately conserved [70,71].
2.3. Envelope (E) Protein
The envelope (E) glycoprotein of coronaviruses is a small integral transmembrane protein ranging from 8.4 to 12 kDa or 74–109 amino acid residues in length that forms an ion channel when it oligomerizes [72]. Its translation is cap-independent and relies on an internal ribosome entry site (IRES) [73]. Despite being a small membrane protein, E protein performs a crucial role in viral assembly and release [74]. It is also an important protein in viral pathogenesis and in the downregulation of antiviral immune response [75].
2.4. Nucleocapsid (N) Protein
Nucleocapsid (N) protein is one of the viral structural proteins with a size ranging from 43–50 kDa. This viral phosphoprotein binds with the helical RNA genome of coronaviruses and provides stability [76]. The N-terminal domain of the nucleocapsid protein is believed to be responsible for the proteins’ ability to bind to the nucleic acid of the virus, while the C-terminal domain may be involved in N protein oligomerization [77]. Furthermore, N protein is important in viral assembly and the maintenance of virion structure. In addition, it is an essential protein in coronavirus replication and transcription since it is confined to both the replication/transcriptional region of the coronaviruses and to the Endoplasmic reticulum-Golgi intermediate Compartment (ERGIC), where the virus is collected and transported [78]. McBride and co-authors comprehensively reviewed the multifunction of N protein in SARS-CoV’s life cycle and cellular response [79]. In terms of antigenicity, N proteins of SARS-CoV induced a strong humoral and cellular immune response. Antibodies against N protein were found to be longer-lasting and more abundant in SARS-CoV patients than for other major structural proteins, which may be attributed to the higher expression of this protein over the rest [80,81].
3. Recombinant Protein Expression Systems and the Production of Key Structural Proteins of Coronaviruses
One of the technologies explored by scientists who work in coronavirus research is the use of recombinant protein technology to produce key structural proteins of coronaviruses. This section presents the different platforms used in the production of recombinant proteins and their different applications to combat coronaviruses afflicting infection and death to animals and humans, as summarized in Table 2.
Table 2.
Recombinant protein expression system and its various applications on different coronaviruses.
3.1. Bacterial Systems
Bacterial cells are one of the most commonly used systems in recombinant protein production. The earliest and most widely exploited bacterial species to serve this purpose was Escherichia coli. E coli is the most popular expression platform as a cell protein factory for several reasons, as reviewed by Rosano and Ceccarelli (2004), i.e., (i) a very short doubling time, which may only take around 20 min; (ii) high density of biomass in culture; (iii) inexpensive and readily available culture media for cultivation; and (iv) fast and easy transformation of heterologous DNA in as little as 5 min [127]. The first-ever in vitro edited plasmid shown to be biologically functional when transformed into E. coli was described by Cohen and colleagues in 1973 when they constructed bacterial antibiotic resistance plasmids by joining EcoRI-treated plasmids or plasmid DNA fragments [128].
This advancement in plasmid technology as a means of transportation and manipulation of foreign DNA into a host cell, particularly in prokaryotic cells, gave rise to a plethora of possibilities in genetic engineering and recombinant DNA and protein technologies. Although it is not a recombinant protein, mRNA technology used to produce drug products and other biopharmaceuticals is emerging and shares a mutual step with recombinant protein expression, which is the use of plasmids [129]. This is because plasmids are easy to replicate and reliably contain the target gene sequence. In 1982, Genentech developed the first-ever recombinant human insulin expressed in E. coli (Humulin®) and was the first-ever recombinant peptide drug hormone marketed by Eli Lilly [130]. However, E. coli may have some drawbacks in terms of secretion of protein products into the culture medium, problems in glycosylation and other post-translational modification, and safety due to the presence of lipopolysaccharide endotoxin. Hence, other Gram-negative bacteria are gaining traction as protein expression systems. Pseudomonas, for example, grows rapidly and has the ability to secrete proteins [131]. Vibrio natriegens is a non-pathogenic Vibrio with a doubling time of <10 min at optimal growth conditions and has been used as a host for the expression of multisubunit membrane protein complexes [132]. Gram-positive bacterial expression systems are also gaining popularity, such as lactic acid bacteria (LAB), Streptomyces, Corynebacterium [133], and Bacillus [134,135]. Lactic acid bacteria, as compared to E. coli, are generally recognized as safe (GRAS) and do not induce anaphylaxis due to the absence of endotoxins, making it an ideal protein factory for food-grade protein and biopharmaceutical production [136]. Gram-positive bacterial expression systems are also known to secrete large amounts of soluble proteins into growth medium, indicating a very efficient secretion system [137,138]. Recently, when highly infectious strains of coronaviruses emerged in human populations and brought pestilence to domesticated animals, prokaryotic systems of protein expression were utilized for various purposes. Much of the work in diagnostics and prophylaxis development was focused on the S protein, which is essential in the binding and fusion of the virus to the host cell.
There are several reports of prokaryotic systems of recombinant protein production that were used to develop vaccines for human coronavirus infections. In 2005, Woo et al. showed that among all the combinations of SARS-CoV vaccines they examined in their pre-clinical studies in mice, those primed with spike polypeptide DNA and boosted with spike polypeptide heterogeneously produced in E. coli with pQE30 as the expression vector generated the highest titer of neutralizing antibody [82]. Aside from E. coli, a lactic acid bacteria platform was also harnessed to produce recombinant spike protein. Wang and colleagues recently reported that they successfully produced a recombinant Lactobacillus plantarum expressing SARS-CoV-2 S protein using pLP-tS as an expression plasmid. The recombinant S protein produced is an oral vaccine candidate that is tolerant to the hostile environment of the gastrointestinal system [83]. Lactic acid bacteria as food-grade bacteria are good candidates in producing oral vaccines that will effectively express target protein subunit antigens and elicit mucosal immune responses and provide protection against viral pathogens of the gastrointestinal and respiratory tract in a manner that is cost-effective and easy to store and administer.
Aside from vaccines, bacterial expression systems were also exploited to produce recombinant protein for the diagnostic development of human coronavirus infections. An indirect ELISA that used recombinant ectodomain and S1 subunit of the spike of MERS-CoV as a coating antigen produced from E. coli BL21 cells using pQE2 expression plasmid was developed by the group of Hashem and co-researchers [84]. The ELISA method they developed maintained a high level of sensitivity and specificity of around ≥90%, indicating that this could be a suitable platform of choice for seroepidemiological testing and surveillance in regions endemic to MERS-CoV. Recombinant N protein was also used in the development of an ELISA based-detection system for the diagnosis of HCoV-OC43 [85] and SARS-CoV without cross-reactivity with HCoVs OC43 and 229E [86]. The former reported the use of E. coli BL21 (DE3) cells with pET-28 as the expression plasmid vector, while the latter used E. coli strain Origami B (DE3) pLysS competent cells pET21a as the expression vector.
In addition to the application of bacterial systems in the production of recombinant proteins for human coronaviruses, similar systems were used to target coronaviruses affecting animals. Zou and co-workers (2015) described monoclonal antibodies (MAbs) developed against a recombinant S1 protein subunit of avian (IBV), heterologously expressed in Escherichia coli BL21 (DE3) competent cells using pET-32a (+) as the expression vector [87]. The same technology was utilized by Wang et al., where they produced MAbs against a truncated S1 protein subunit of porcine epidemic diarrhea virus (PEDV) also produced through E. coli BL21(DE3) competent cells using pET32a(+)/tS1 as the expression vector [88]. The antibodies and antigens they developed may be valuable in the development of enzyme link immunosorbent assay (ELISA) test for the detection of antigen and antibody or therapeutic antibody for PEDV in swine.
Another CoV protein chosen to be heterologously expressed in a bacterial system for diagnostic platforms to detect animal coronaviruses is M protein. M protein is the most abundant protein in coronaviruses and is important in eliciting immune protection against CoVs. It may also be an ideal candidate for the development of serodiagnostic devices. In 2017, Luo and co-workers developed an indirect ELISA using a recombinant M protein produced by E. coli BL21 with the use of pET-32a plasmid DNA expression vector to detect anti-porcine deltacoronavirus M protein IgG with diagnostic sensitivity and specificity of >90% [89]. Another ELISA-based method was developed using the recombinant M protein of canine coronavirus (CCoV) in E. coli M15(pREP4) employing the pQE30 expression plasmid [90]. The sensitivity and specificity of the recombinant M protein-based ELISA were unchanged compared to Western blot and whole CCoV ELISA. Therefore, the recombinant M protein-based ELISA represents an alternative and valid serodiagnostic tool for CCoV.
Recombinant N protein was also expressed in E. coli for the development of immunodiagnosis platforms for several coronaviruses affecting animals. Using pET-32a and pGEX-6P-1 as expression vectors in E. coli BL21(DE3) competent cells, Su et al. (2016) heterologously expressed N protein to detect antibodies against PDCoV through indirect ELISA [91]. Their data demonstrated that the rPDCoV-N-ELISA could be used for epidemiological investigations of PDCoV. In two different studies where N protein was used to develop IBV immunodiagnosis, Finger and co-workers (2018) expressed N protein in E. coli BL21(DE3) Star competent cells using pAE/n vector [92], while Pradhan and colleagues (2014) expressed N protein in E. coli BL21 (DE3) Rosetta cells using pET32a (+) as the expression vector [93]. An enzyme-linked immunosorbent assay to detect antibodies to TCoV was also developed by Abdelwahab et al. (2015), wherein the recombinant plasmid containing the entire N protein gene was inserted in pTri-N and transformed into competent Escherichia coli strain Tuner (DE3) pLacI or Origami (DE3) pLacI [94].
3.2. Yeast Systems
Yeasts are microscopic unicellular fungal hosts that have emerged as indispensable tools in recombinant protein technology. Since the 1980s, the most widely used protein expression system among eukaryotes has been the yeast Saccharomyces cerevisiae. The complete genome sequence of S. cerevisiae in 1996, the first whole-genome sequence ever published for any eukaryotic organism, gave rise to a better understanding and utilization of yeasts as cellular protein factories [139]. Saccharomyces cerevisiae offers many advantages as a producer of recombinant biopharmaceutical proteins, including the production and proper folding of many human proteins, secretion of proteins to extracellular medium facilitating isolation and purification, proper post-translational modification of proteins such as glycosylation, and ease in propagation just like the unicellular prokaryotes [140]. Despite these advantages, S. cerevisiae may also have some drawbacks, such as its production of hypermannosylated glycan structures, hyperglycosylation of proteins, low protein yield, and plasmid instability [141]. Lately, other species of yeasts such as Pichia pastoris, Hansenula polymorpha, Yarrowia lipolytica, Kluyveromyces lactis, Talaromyces emersonii, and others were considered as biofactories of recombinant proteins to overcome the disadvantages of S. cerevisiae as a heterologous host for protein production [142,143]. The methylotrophic yeast Pichia pastoris, first described by Koichi Ogata and colleagues in 1969 [144] and now reclassified as Komagataella pastoris, is becoming mainstream in biotechnology, particularly as a heterologous host in protein production. The use of P. pastoris began in the 1970s when the Philips Petroleum Company developed media and a protocol for its culture as a source of a single-cell protein animal feed additive. Later on, it was used as a heterologous protein expression organism by Salk Institute Biotechnology/Industrial Associates, Inc. [145]. In 1993, Philips Petroleum sold its patent to Research Corporation Technologies, which currently holds the patent, and licensed the Invitrogen Corporation to sell components of the system [128]. As with S. cerevisiae, P. pastoris has advantages and disadvantages, as reviewed by Karbalaei and co-authors (2020). To name a few, P. pastoris is inexpensive, has relatively rapid expression time, co-translational and posttranslational processing, and direct secretion of its recombinant protein into the culture medium [146]. However, there are also some drawbacks such as high plasmid requirements of its competent cells, protein production dependence on the concentration of methanol (toxic to cell viability at high concentration), presence of very few selectable markers, and secretion of proteases that may hydrolize and destroy secreted protein [146]. Despite these disadvantages, P. pastoris still offers additional advantages over S. cerevisiae, including the production of more complex secondary metabolites and complex reduced molecules [147]. Moreover, its secretory pathway, including the Golgi structure, was shown to resemble higher eukaryotic cells, including humans, which may be the reason why P. pastoris secretes recombinant proteins more efficiently than S. cerevisiae [147]. In 2017, Tran et al. reported that P. pastoris has emerged as an alternative host in the production of higher yield and more active recombinant granulocyte-macrophage colony-stimulating factor (GM-CSF) [148].
Chen et al. (2014) produced a vaccine candidate expressed in Pichia pastoris X-33 with the use of expression vector pPICZαA containing the gene encoding for the receptor-binding domain (RBD) of the spike protein of SARS-CoV [95]. The recombinant protein they created from P. pastoris exhibited lower glycosylation levels with a higher expression yield and induced an RBD-specific neutralizing antibody response against both the pseudovirus and live SARS-CoV, making it an ideal protein subunit vaccine candidate for SARS-CoV. In 2017, the same authors, Chen, and co-workers, improved the process of recombinant SARS-CoV RBD protein production in P. pastoris, rendering the process highly reproducible with high yields and high recovery [96]. Chen and colleagues also reported in 2021 that both the RBD of the spike protein of SARS-CoV that they produced in P. pastoris and the homologous RBD of SARS-CoV-2 were recognized by a neutralizing monoclonal antibody directed to an epitope outside the receptor-binding motif (RBM) [149]. This observation can be attributed to the substantial amino acid sequence similarity of the RBD of SARS-CoV-2 and the recombinant SARS-CoV RBD, suggesting that the recombinant subunit RBD protein vaccine produced in P. pastoris against SARS-CoV may offer partial protection against SARS-CoV-2 [149]. Chuck and co-workers (2009) also published a paper describing how they used P. pastoris to produce pure and biologically active RBD of the S protein of SARS-CoV [97]. They demonstrated that P. pastoris KM71H with the use of expression vector pPICZα-A was an excellent alternative to express active viral antigens with extensive posttranslational modifications such as glycosylation, which is important in protein folding and functionality. The recombinant RBD protein they produced may subsequently be used in diagnostic, prophylactic, receptor identification, and other functional studies. Aside from the spike protein subunit, nucleocapsid protein was also successfully expressed in the methylotrophic yeast, P. pastoris, as reported by Liu et al. [98]. Preliminary results indicate that the conformation of the heterologously expressed N protein using pPIC3.5K DNA vector transformed in P. pastoris GS115 (his-mut+) is almost the same as the natural SARS-CoV N protein. The authors reported that the recombinant N protein has high specificity with a SARS-CoV N protein mAb and SARS-CoV positive sera, underscoring its potential use in developing an immunodiagnostic assay.
3.3. Plant Expression Systems
The use of plants or plant cell cultures to produce diagnostic reagents, biopharmaceuticals, and other medically important proteins is called plant molecular farming. Using plants as heterologous competent hosts expressing foreign proteins started in 1986 when chimeric human growth hormone was produced in transformed tobacco and sunflower callous tissues [150]. Following this, in 1989, monoclonal antibodies were produced in transgenic tobacco [151], and human serum albumin was expressed in transgenic tobacco and cell cultures [152]. In the succeeding years, several vaccine antigens were expressed in plants and plant cell cultures, such as the hepatitis B surface antigen expressed in transgenic tobacco in 1992 [153], followed by an epitope derived from the VP1 of foot-and-mouth disease virus expressed on cowpea through cowpea plant mosaic virus [154]. Plants as a biopharmaceutical production platform offer a wide range of advantages [155], including rapid and affordable production, optimized growth conditions, absence of pathogen and toxin contaminants, affordability, and a capacity to perform post-translational modification. Nevertheless, some issues remain, such as limitations in glycosylation and some other regulatory compliance problems. There are two types of expression methods in plants, namely, stable and transient expressions. Either type can be used depending on the applications that use whole or minimally processed plants or plant parts. Stable transformation involves Agrobacterium-mediated transformation or particle bombardment with the nucleus or chloroplast, while transient expression is performed by using a plant virus or by infiltration with Agrobacterium [156]. Once plants or plant parts are transformed, they can be cultivated and produced in two ways, i.e., open-field cultivation, meaning outdoor plantation, or a closed-indoor system using greenhouses, vertical farming units, plant cell bioreactors, and hydroponic systems [156].
A study using plants as a biofactory for coronavirus protein was investigated by Pogrebnyak and co-authors (2005), wherein they successfully expressed the S1 domain of the SARS-CoV spike protein in tomato (Lycopersicon esculentum L. cv. Money Maker) and in low-nicotine tobacco plants (Nicotiana tabacum cv. LAMD-609), both transformed by the Agrobacterium-mediated method [99]. When the authors fed mice with the tomato fruit expressing S1 protein, a significant increase in SARS-CoV-specific IgA was observed. Western blot and ELISA revealed that sera of mice parenterally primed with tobacco-derived S1 protein exhibited SARS-CoV-specific IgG. In the following year, a different team led by Li (2006) stably expressed the S1 subunit of the SARS-CoV S protein to high levels in the cytosol (nuclear-transformed) and chloroplasts (plastid-transformed) of tobacco and lettuce plants through Agrobacterium-mediated transformation [100]. Their work further supports the idea that a safe recombinant protein subunit vaccine against coronaviruses can be manufactured and administered orally through edible plants. Similarly, Zheng and colleagues expressed another SARS-CoV immunogenic recombinant protein, the nucleocapsid protein, in tobacco leaves using Agrobacterium-mediated transformation [101]. Their results were considered the very first demonstration that plant-expressed recombinant SARS-CoV N protein induces a robust humoral and cellular immune response in mice. Recombinant proteins were also produced for animal coronaviruses.
Plant expression systems were also used to produce recombinant proteins intended for animal coronaviruses. Transgenic potato plants created by Gomez et al. (2000) expressed the N terminal domain of the S protein of the swine-transmissible gastroenteritis coronavirus (TGEV) through infection with transformed Agrobacterium tumefaciens [102]. When potato tuber extracts from the transgenic potato crops were intraperitoneally injected into mice, the test animals developed an anti-TGEV antibody. When the same potato tubers were fed directly to mice, the mice developed a serum antibody specific to the heterologously expressed S protein of TGEV. In a separate study with another swine coronavirus, PEDV, Tien et al. (2019) successfully expressed in higher amounts a second neutralizing epitope of the S protein of PEDV called S1D in the chloroplast of Nicotiana tabacum when fused with the B subunit of cholera toxin (CTB) [103]. The leaf tissue of N. tabacum was transformed using bombardment with gold particle-DNA plasmid complexes. This recent strategy of a chloroplast-derived fusion protein of CTB-S1D proves to be a promising oral vaccine candidate in the prevention of PEDV infections in the hog industry.
Additional evidence that plant-derived SARS-CoV proteins have potential as diagnostic, prophylactic, and therapeutic tools that can be rapidly manufactured at low cost with minimal risk was provided by Demurtas and co-workers (2016) who demonstrated that SARS-CoV N and M proteins could be transiently expressed in Nicotiana benthamiana, a close relative of tobacco [104]. The M protein was expressed through Agrobacterium infiltration. This is the very first description of SARS-CoV M protein expression in plants. On the other hand, the N protein was expressed through a virus-derived vector and recognized by sera derived from the 2003 SARS-CoV patients in Hong Kong.
3.4. Mammalian Expression Systems
Another very useful expression system for recombinant protein production is the mammalian system. The very first human cell to be cultured in vitro in 1953 was the HeLa cell line derived from cervical cancer and gave rise to the use of mammalian cells in many different applications, such as the production of recombinant biopharmaceutical products [157,158]. Heterologous genes were first successfully transfected in vitro into mammalian cells in 1978 [159]. In 1996, Genentech produced a human tissue-derived plasminogen activator (TPA, Activase®) for thrombolysis therapy, which was the very first mammalian cell (CHO cells)-derived biopharmaceutical to receive regulatory approval [160]. Mammalian cell systems for recombinant protein production offer diverse advantages, such as good protein folding, absence of potentially immunogenic post-translational modification products because of a humanized glycosylation pattern, good secretion capacity, absence of pyrogens, medium to high product yield, high product quality, and amenability to several transfection methods [161,162]. Despite these advantages, there are also disadvantages in terms of its use in recombinant protein production; namely, slow growth rate, high overall cost, high production time, difficult propagation, very low scale-up capacity, high purification cost, and high risk of contamination with viruses, prions, and oncogenic DNA [161,162]. Khan and co-authors (2013) and Hacker and Wurn (2007) presented comprehensive reviews on the different appropriate cell lines and expression vectors required for the transfection of mammalian cells [160,163].
Numerous works have already been reported on the use of mammalian cell systems to study and combat coronaviruses. Special interest in this technology intensified when HCoVs infections exploded to epidemic and pandemic proportions, including COVID-19 caused by SARS-CoV-2. A study published by Tai et al. (2020) characterized the RBD of SARS-CoV-2 recombinantly expressed in human embryonic kidney (HEK) 293T cells with the use of pFUSE-hIgG1-Fc2 as the expression vector [105]. They reported that SARS-CoV-2 RBD protein produced in 293T cells exhibited strong binding affinity to human and bat cell-associated and soluble ACE2 receptors. The authors also showed that the recombinant RBD protein could block S protein-mediated SARS-CoV-2 and SARS-CoV pseudovirus entry to its host cell through the ACE2 receptor. In addition, the recombinant RBD protein induced antibodies that neutralized both SARS-CoV-2 and SARS-CoV, attributed to the high sequence homology of their RBDs. These results highlight RBD’s role as a viral attachment inhibitor and possible use as a protein subunit vaccine. The same group of Tai et al. (2017) previously presented that recombinant RBD proteins of several MERS-CoVs produced in 293T cells using pFUSE-hIgG1-Fc2 elicited cross-neutralizing antibodies against divergent human and camel MERS-CoVs [106]. The recombinant RBD proteins are also bound to dipeptidyl-peptidase 4 (DPP4), the human receptor for the MERS-CoVs spike protein. The authors confirmed that recombinant RBD protein constitutes a safe, effective, broad-spectrum vaccine that can even trigger cross-neutralizing antibody production against infection with current and future divergent MERS-CoVs. Another protein subunit vaccine was described by Kim and colleagues [107] based on the expression of the S1 subunit of S protein of MERS-CoV and SARS-CoV-2 in HEK-293 cells through an adenovirus vectors pAd and pmax. The recombinant S1 protein subunit vaccine for MERS-CoV and SARS-CoV-2 were inoculated subcutaneously in test animals through a dissolvable microneedle array (MNA). This delivery strategy elicited robust antigen-specific antibodies, underscoring its effectiveness as a recombinant protein subunit vaccine delivery system. A different strategy was reported by Elshabrawy et al. in 2012. Instead of recombinantly producing the S1 protein subunit to raise human monoclonal antibodies, their team produced Heptad Repeat 1 and Heptad Repeat 2 (HR1 and HR2) of the S2 subunit of the S protein in 293FT cells using pcDNA3.1 and S1Ig plasmid [108]. This strategy was undertaken by the authors because RBD of the S1 domain of the S protein of SARS-CoV has a high mutation rate, which may give way to neutralization escape while maintaining the ability to infect cells. On the contrary, the highly conserved S2 domain of the S protein of different clinical isolates of SARS-CoV is more amenable to neutralization with antibodies specific to this region, thus conferring protection to broader isolates of SARS-CoV [108]. This hypothesis is congruent to the results of this study, where human monoclonal antibodies against the highly conserved region of HR1 and HR2 of the S2 domain of S protein are more broadly neutralizing. In a different approach, Xu et al. (2020) assembled SARS-CoV-2 VLPs, instead of just one singular protein, through the expression of two SARS-CoV-2 structural proteins, M and E proteins, in Vero E6 cells [109]. They demonstrated that M and E proteins are basic requirements for the efficient assembly and release of SARS-CoV-2 VLPs. This report reiterates that VLPs presenting native conformation of SARS-CoV-2 virion expressed through the use of pEAK13 plasmid vector can be used as a potential vaccine for SARS-CoV-2 and other virological investigations involving SARS-CoV-2. Although contrasting with the claims of Xu and co-workers in 2020, Siu and colleagues (2008) underscored in their work that M, E, and N proteins of SARS-CoV are required for the efficient assembly, trafficking, and release of VLPs expressed in Vero E6 African green monkey kidney cells [115]. One of the most successful protein subunit vaccine candidates against SARS-CoV-2 is MVC-COV1901 of Medigen Vaccine Biologics Corporation, recombinantly expressed in a mammalian culture system. MVC-COV1901 comprises the SARS-CoV-2 prefusion spike protein expressed in ExpiCHO-S cells transfected with the plasmid encoding for S-2P protein ectodomains [110]. MVC-COV1901 adjuvanted with CpG 1018 and aluminum hydroxide was safe and immunogenic after a Phase 1 dose-escalation study [111].
A mammalian system of recombinant protein expression was also used in the development of diagnostic platforms to detect coronaviruses affecting animals and humans. Human embryonic kidney (HEK)-293T cells were employed by Thachil and co-workers to express the putative S1 portion of the spike protein of PDCoV, also known as porcine coronavirus HKU15, using pcDNA3.1 as the expression vector in the development of an indirect ELISA that detects IgG against the said virus [112]. Their work illustrates the use of mammalian systems for protein expression in developing diagnostic platforms for coronavirus affecting livestock. In addition to the work of Thachil, Esposito and colleagues (2020) heterologously expressed trimeric SARS-CoV-2 S protein in Expi293F cells for serology assays to diagnose SARs-CoV-2. Their optimized method consistently yielded 5 mg of protein per liter of expression culture for two commonly used forms of SARS-CoV-2 spike protein [113]. In a separate study by Mehalko and co-workers (2020), several improvements were made in the production of SARS-CoV-2 RBD recombinant protein to increase protein yield, such as the use of a strong CMV51 promoter, which provided high-level protein production in HEK293 cells. They also tweaked their incubation temperature and time to 32 °C for 96 h to maximize production while maintaining high levels of cell viability. In addition, they developed a simplified and robust purification strategy that resulted in higher protein yield. Lastly, they reported that the serology assay improved when a caboxy-terminal streptavidin-binding protein (SBP) tag was added to the recombinant protein [114].
Finally, the recent work of Hoffman and co-authors transfected 293T cells to produce a pseudotyped virus expressing SARS-CoV and SARS-CoV-2 S protein to investigate whether SARS-CoV and SARS-CoV-2 S protein was recognized by human angiotensin-converting enzyme 2 (ACE2) and human cellular serine protease TMPRSS2, and whether this interaction was blocked by a clinically proven protease inhibitor [40]. Taken together, scientists exploited mammalian cell expression systems to study the biology and host–pathogen interaction of coronaviruses with their various hosts, in addition to their use in the production of protein subunits to develop diagnostics, preventive vaccines, and therapeutic arsenal against the virus.
3.5. Insect/Insect Cell Systems
Insect and insect cell systems, generally using baculovirus expression vector systems (BEVS), have been widely used in recombinant protein production, especially for biopharmaceuticals. Baculoviruses form a diverse group of large rod-shaped enveloped viruses (30–60 nm × 250–300 nm) containing supercoiled double-stranded genomes of varying size (80–180 kb) encoding 90–180 genes [164]. Baculoviridae is currently divided into 4 genera, alpha-, beta-, gamma-, and deltabaculovirus. Alphabaculovirus, Gammabaculovirus, and Deltacoronivirus are nucleopolyhedroviruses that infect insects in the orders Lepidoptera, Hymenoptera, and Diptera, respectively, while Betabaculoviruses are the granuloviruses that infect only Lepidopterans [165]. No infection outside of the Phylum Arthropoda has yet been observed [166]. Baculoviruses play a major role in the natural control of the insect population, and much of the attention in earlier years was due to their threat to the silk industry. Because of their capacity to infect a wide variety of insect species, baculoviruses were developed as biopesticides as well as an efficient tool for heterologous protein production in insect cells and insects [167]. Insect cell expression systems offer an array of advantages in the production of recombinant proteins, including high yield of the desired product, ability to produce complex proteins including secreted, membrane, and intracellular proteins, proper protein folding and post-translational modification, and no risk of contamination with prions and oncogenic DNA [155,161,162]. While advantageous, insect cell expression systems remain imperfect due to some disadvantages, such as the high cost of materials (particularly the culture media to maintain the cells) and time-consuming procedures. When BEVS were invented and used to produce human interferon in insect cells, insect cells were revolutionized as a system to produce therapeutic proteins [168]. To date, several commercially available cell lines are used for protein expression through BEVS, including Sf21 and its clone Sf9, High 5 (Hi5) cells, and others that were comprehensively reviewed by van Oers et al. [169]. Aside from the use of insect cells to produce the desired heterologously expressed proteins, several groups have kicked it up a notch and produced protein subunits directly in insect larvae. For example, Lin and co-workers immunized chickens against a lethal dose of H5N1 pathogenic avian influenza with hemagglutinin produced in Trichoplusia ni larvae [170]. The very first biopharmaceutical for animal use derived from insect cells through baculovirus expression vector technology came in 2000 (the Porcilis® Pesti) [171], followed by the first-ever insect cell-derived human cervical cancer VLP vaccine, Cervarix, by GlaxoSmithKline in 2007 [172]. In an attempt to counter the devastating effects of coronaviruses infections in livestock and humans, several studies have used insect cell expression systems to produce protein subunits in the development of diagnostics, prophylaxis, and therapeutics.
One application of insect/insect cell expression system for human use is the development of protein subunit vaccines. Li and colleagues (2013) used Sf9 and High 5 insect cells that were co-transfected with pAcGP67 and linearized baculovirus DNA to produce trimeric and monomeric SARS-CoV S protein as a protein subunit vaccine candidate [116]. This recombinant S protein produced neutralizing antibodies and protected the vaccinated mice in a viral challenge. Similarly, S trimer protein micellular nanoparticles of SARS-CoV and MERS-CoV were produced through recombinant baculovirus-infected Sf9 cells under the transcriptional control of the Autographa californica Multiple Nuclear Polyhedrosis Virus (AcM-NPV) polyhedrin promoter, as reported by Coleman and co-workers (2014) [117]. Their approach stimulated the production of neutralizing antibodies in mice. Instead of using insect cells, Fujita and company (2020) made use of the baculovirus-silk worm expression system for the efficient production of recombinant SARS-CoV-2 S glycoprotein, which would be useful in the development of serological detection kits, antigen for immunization for antibody production, and protein subunit vaccine [118]. Their group made use of pFastBac1 transfer vector and BmNPV bacmid to create the recombinant virus used to infect the fourth instar larvae of silkworm (Bombyx mori). One of the most successful examples of the use of insect/insect cell-baculovirus expression vector systems against coronaviruses is the SARS-CoV-2 recombinant S protein nanoparticle vaccine candidate by Novavax called NVX-CoV2373 [119]. This full-length S protein nanoparticle adjuvanted with plant-based saponin called Matrix M elicited virus-neutralizing antibodies, anti-S IgG that blocks the binding with human angiotensin-converting enzyme 2 (hAce2) receptor, and antigen-specific T cells both in mice and baboon models. Keech et al. recently reported the encouraging results of the first-in-human trial of the said vaccine candidate against SARS-CoV-2, wherein the adjuvanted recombinant protein subunit vaccine induced a level of anti-S protein IgG and neutralizing antibodies that even exceeded the mean response in convalescent sera of naturally infected SARS-CoV-2 patients [120]. The Phase I clinical trial also showed high levels of antigen-specific T cells with a largely T helper 1 (Th1) phenotype [120].
Aside from vaccine candidates for human use, insect cell-BEVS were also used by Hsu et al. (2020) to produce a VLP vaccine candidate against PEDV [121]. Hsu and colleagues made use of a novel polycistronic baculovirus expression vector developed by the laboratory of the authors of this review wherein the S gene was codon-optimized in an insect cell system and made use of honeybee melittin as a signal peptide and the addition of 6xHis-tag. The M gene was linked by the 2A-like sequence driven by the polyhedrin promoter. The E gene was translated through an internal ribosome entry site (IRES) of Rhopalosiphum padi virus (RhPV). The enhanced green fluorescent protein (EGFP) gene was inserted in the plasmid and expressed through another IRES, Perina nuda picorna-like virus (PnV339 IRES), resulting in a plasmid called pFastBac1-HM6H-PEDV- S-2A-M-Pnv339- eGFPRhir-E [121]. The post-weaning pigs that were immunized with the VLP preparation with mucosal adjuvants chemokines CCL25 and CCL28 had enhanced anti-PEDV S IgG, mucosal IgA, and cellular immunity, and superior immune protection against PEDV. These results make the BEVS a potential platform for the production of a coronavirus VLP.
In addition to the use of insect cell protein expression systems for human and animal vaccine development, many reports have been published on their use in recombinant protein production for diagnostics purposes. Their use in poultry animals was explored by Yilmaz and colleagues (2018) when they expressed nucleocapsid protein of avian coronavirus-infectious bronchitis virus in a baculovirus expression system in Sf9 cells for ELISA diagnostic test development [122]. They concluded that the baculovirus expressed IBV N protein could serve as a diagnostic antigen for the detection of IBV in chickens due to the high agreement of the ELISA they developed with commercially available IBV ELISA test kits. In a similar study wherein coronavirus N protein was expressed using BEVS in Trichoplusia ni (High 5) insect cells, Severance et al. (2008) developed a solid-phase ELISA to detect IgG specific to 4 coronavirus strains (229E, HKU1, NL63, and OC43) and a feline coronavirus [123]. This ELISA assay detected high rates of exposure to 229E, NL63, and OC43 and a moderate rate of exposure to HKU1 among individuals tested in a metropolitan population. Kato et al. (2019) successfully expressed MERS-CoV spike protein without its transmembrane and cytoplasmic domain in the larvae of silkworm and recombinant S protein-displaying nanovesicles in Bombyx mori (Bm5) cells which may be a lead nanoparticle-based vaccine and diagnostic detection for MERS-CoV [124]. Another diagnostic system based on BEVS and insect cell expression system was reported by Lee et al. (2018) to detect MERS-CoV antibodies and antigens [125]. Using the recombinant full-length MERS-CoV S protein harvested from baculovirus-infected Sf9 cells, their group developed indirect and sandwich ELISA to detect MERS-CoV induced antibodies and S protein antigens, respectively.
Insect/insect cell systems using BEVS were also employed to elucidate viral entry of murine hepatitis coronavirus (MHCoV) into its host cell through the CEACAM1a receptor. Shang et al. (2020) recombinantly expressed mouse coronavirus spike protein in Sf9 cells using BEVS. The S protein produced was used to show how the spike protein complexes to its receptor and provided mechanistic insights into the diversity of mechanisms used by different coronaviruses to gain access to their host cells [126].
4. Concluding Remarks
We live in an interconnected ecosystem and in a One Health web where humans, animals, and the environment always interact. As long as viruses continue to evolve and adapt to new hosts, humans will be a receptive species to viral pathogens harbored by wildlife, such as coronaviruses. The preservation of the human species through adaptation and survival against viral ravages may be aided by technologies, including biotechnology, such as the production of biopharmaceuticals through recombinant protein expression systems. Today, there are a variety of expression systems to choose from in the development of vaccines, diagnostics, and therapeutics for diseases caused by coronaviruses. Each has its own advantages and disadvantages; thus, the choice of a competent host cell that will serve as a factory for the desired protein will all depend on the protein that ought to be expressed. However, based on the reports presented in this review, the two most successful platforms in delivering recombinant proteins used as protein subunit vaccines, particularly for COVID-19, are the mammalian expression system and the insect cell/insect expression system, which may be attributed to their good secretion capacity, high product yield, and ability to produce complex proteins and high-quality proteins. As more and more proteins are produced and enter clinical trials for diagnostics, prophylaxis, and therapeutic purposes, recombinant protein technology involving different expression systems also becomes better and more sophisticated, especially in the fight against viral pathogens of global public health importance challenging our time and humanity such as coronaviruses.
Author Contributions
Conceptualization, A.B.S.C. and T.-Y.W.; writing—original draft preparation, A.B.S.C.; writing—review and editing, T.-Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Technology (MOST), Taiwan, grant number MOST-109-2321-B-033-001.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to express our sincere appreciation to Marco Vignuzzi of Viral Population and Pathogenesis Unit, Institute Pasteur, Paris, France, for his invaluable help in ironing out some English grammar errors in this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carstens, E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2009). Arch. Virol. 2010, 155, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Milek, J.; Blicharz-Domanska, K. Coronaviruses in Avian Species—Review with Focus on Epidemiology and Diagnosis in Wild Birds. J. Vet. Res. 2018, 62, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.D.; Wang, Z.Y.; Zhang, S.F.; Li, X.; Li, L.; Li, C.; Cui, Y.; Fu, R.B.; Dong, Y.Z.; Chi, X.Y.; et al. Aerosol and Surface Distribution of Severe Acute Respiratory Syndrome Coronavirus 2 in Hospital Wards, Wuhan, China, 2020. Emerg. Infect. Dis. 2020, 26, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and Sources of Endemic Human Coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [CrossRef]
- Ye, Z.W.; Yuan, S.; Yuen, K.S.; Fung, S.Y.; Chan, C.P.; Jin, D.Y. Zoonotic origins of human coronaviruses. Int. J. Biol. Sci. 2020, 16, 1686–1697. [Google Scholar] [CrossRef]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef]
- Raj, G.D.; Jones, R.C. Infectious bronchitis virus: Immunopathogenesis of infection in the chicken. Avian Pathol. 1997, 26, 677–706. [Google Scholar] [CrossRef]
- Ismail, M.; Tang, Y.; Saif, Y. Pathogenicity of turkey coronavirus in turkeys and chickens. Avian Dis. 2003, 47, 515–522. [Google Scholar] [CrossRef][Green Version]
- Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol. J. 2015, 12, 193. [Google Scholar] [CrossRef]
- Pensaert, M.; De Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef]
- Liu, Q.; Gerdts, V. Transmissible Gastroenteritis Virus of Pigs and Porcine Epidemic Diarrhea Virus (Coronaviridae). Encycl. Virol. 2021, 2, 850–853. [Google Scholar] [CrossRef]
- Mora-Diaz, J.C.; Pineyro, P.E.; Houston, E.; Zimmerman, J.; Gimenez-Lirola, L.G. Porcine Hemagglutinating Encephalomyelitis Virus: A Review. Front. Vet. Sci. 2019, 6, 53. [Google Scholar] [CrossRef]
- Korner, R.W.; Majjouti, M.; Alcazar, M.A.A.; Mahabir, E. Of Mice and Men: The Coronavirus MHV and Mouse Models as a Translational Approach to Understand SARS-CoV-2. Viruses 2020, 12, 880. [Google Scholar] [CrossRef]
- Hulse, J.D. Human coronaviruses: The deadly seven. ACTA Sci. Microbiol. 2020, 3, 86–89. [Google Scholar] [CrossRef]
- Chen, B.; Tian, E.K.; He, B.; Tian, L.; Han, R.; Wang, S.; Xiang, Q.; Zhang, S.; El Arnaout, T.; Cheng, W. Overview of lethal human coronaviruses. Signal Transduct. Target Ther. 2020, 5, 89. [Google Scholar] [CrossRef]
- Anderson, R.M.; Fraser, C.; Ghani, A.C.; Donnelly, C.A.; Riley, S.; Ferguson, N.M.; Leung, G.M.; Lam, T.H.; Hedley, A.J. Epidemiology, transmission dynamics and control of SARS: The 2002–2003 epidemic. Philos. Trans. R Soc. Lond B Biol. Sci. 2004, 359, 1091–1105. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Mohd, H.A.; Al-Tawfiq, J.A.; Memish, Z.A. Middle East Respiratory Syndrome Coronavirus (MERS-CoV) origin and animal reservoir. Virol. J. 2016, 13, 87. [Google Scholar] [CrossRef]
- Al Mutair, A.; Ambani, Z. Narrative review of Middle East respiratory syndrome coronavirus (MERS-CoV) infection: Updates and implications for practice. J. Int. Med. Res. 2020, 48, 300060519858030. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef]
- Liu, C.; Tang, J.; Ma, Y.; Liang, X.; Yang, Y.; Peng, G.; Qi, Q.; Jiang, S.; Li, J.; Du, L.; et al. Receptor usage and cell entry of porcine epidemic diarrhea coronavirus. J. Virol 2015, 89, 6121–6125. [Google Scholar] [CrossRef] [PubMed]
- Schultze, B.; Krempl, C.; Ballesteros, M.L.; Shaw, L.; Schauer, R.; Enjuanes, L.; Herrler, G. Transmissible gastroenteritis coronavirus, but not the related porcine respiratory coronavirus, has a sialic acid (N-glycolylneuraminic acid) binding activity. J. Virol. 1996, 70, 5634–5637. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Gao, W.; Lu, H.; Zhao, K.; Ding, N.; Liu, W.; Zhao, J.; Lan, Y.; Tang, B.; Jin, Z.; et al. A small region of porcine hemagglutinating encephalomyelitis virus spike protein interacts with the neural cell adhesion molecule. Intervirology 2015, 58, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Dveksler, G.; Pensiero, M.; Cardellichio, C.; Williams, R.; Jiang, G.; Holmes, K.; Dieffenbach, C. Cloning of the mouse hepatitis virus (MHV) receptor: Expression in human and hamster cell lines confers susceptibility to MHV. J. Virol. 1991, 65, 6881–6891. [Google Scholar] [CrossRef] [PubMed]
- Ambepitiya Wickramasinghe, I.N.; de Vries, R.P.; Weerts, E.A.; van Beurden, S.J.; Peng, W.; McBride, R.; Ducatez, M.; Guy, J.; Brown, P.; Eterradossi, N.; et al. Novel Receptor Specificity of Avian Gammacoronaviruses That Cause Enteritis. J. Virol 2015, 89, 8783–8792. [Google Scholar] [CrossRef]
- Promkuntod, N.; van Eijndhoven, R.E.; de Vrieze, G.; Grone, A.; Verheije, M.H. Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 2014, 448, 26–32. [Google Scholar] [CrossRef]
- Zhang, Y.; Buckles, E.; Whittaker, G.R. Expression of the C-type lectins DC-SIGN or L-SIGN alters host cell susceptibility for the avian coronavirus, infectious bronchitis virus. Vet. Microbiol. 2012, 157, 285–293. [Google Scholar] [CrossRef]
- Ji, W.; Peng, Q.; Fang, X.; Li, Z.; Li, Y.; Xu, C.; Zhao, S.; Li, J.; Chen, R.; Mo, G.; et al. Structures of a deltacoronavirus spike protein bound to porcine and human receptors. Nat. Commun. 2022, 13, 1467. [Google Scholar] [CrossRef]
- Li, W.; Hulswit, R.J.G.; Kenney, S.P.; Widjaja, I.; Jung, K.; Alhamo, M.A.; van Dieren, B.; van Kuppeveld, F.J.M.; Saif, L.J.; Bosch, B.J. Broad receptor engagement of an emerging global coronavirus may potentiate its diverse cross-species transmissibility. Proc. Natl. Acad. Sci. USA 2018, 115, E5135–E5143. [Google Scholar] [CrossRef]
- Li, Z.; Tomlinson, A.C.; Wong, A.H.; Zhou, D.; Desforges, M.; Talbot, P.J.; Benlekbir, S.; Rubinstein, J.L.; Rini, J.M. The human coronavirus HCoV-229E S-protein structure and receptor binding. eLife 2019, 8, e51230. [Google Scholar] [CrossRef]
- Hofmann, H.; Pyrc, K.; Van Der Hoek, L.; Geier, M.; Berkhout, B.; Pöhlmann, S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc. Natl. Acad. Sci. USA 2005, 102, 7988–7993. [Google Scholar] [CrossRef]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; van Kuppeveld, F.J.M.; Boons, G.J.; Bosch, B.J.; et al. Human coronaviruses OC43 and HKU1 bind to 9-O-acetylated sialic acids via a conserved receptor-binding site in spike protein domain A. Proc. Natl. Acad. Sci. USA 2019, 116, 2681–2690. [Google Scholar] [CrossRef]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef]
- Li, W.; Hulswit, R.J.G.; Widjaja, I.; Raj, V.S.; McBride, R.; Peng, W.; Widagdo, W.; Tortorici, M.A.; van Dieren, B.; Lang, Y.; et al. Identification of sialic acid-binding function for the Middle East respiratory syndrome coronavirus spike glycoprotein. Proc. Natl. Acad. Sci. USA 2017, 114, E8508–E8517. [Google Scholar] [CrossRef]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Muller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Baker, A.N.; Richards, S.J.; Guy, C.S.; Congdon, T.R.; Hasan, M.; Zwetsloot, A.J.; Gallo, A.; Lewandowski, J.R.; Stansfeld, P.J.; Straube, A.; et al. The SARS-COV-2 Spike Protein Binds Sialic Acids and Enables Rapid Detection in a Lateral Flow Point of Care Diagnostic Device. ACS Cent. Sci. 2020, 6, 2046–2052. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef]
- Flaxman, S.; Mishra, S.; Gandy, A.; Unwin, H.J.T.; Mellan, T.A.; Coupland, H.; Whittaker, C.; Zhu, H.; Berah, T.; Eaton, J.W.; et al. Estimating the effects of non-pharmaceutical interventions on COVID-19 in Europe. Nature 2020, 584, 257–261. [Google Scholar] [CrossRef]
- Consortium, W.H.O.S.T.; Pan, H.; Peto, R.; Henao-Restrepo, A.M.; Preziosi, M.P.; Sathiyamoorthy, V.; Abdool Karim, Q.; Alejandria, M.M.; Hernandez Garcia, C.; Kieny, M.P.; et al. Repurposed Antiviral Drugs for Covid-19—Interim WHO Solidarity Trial Results. N. Engl. J. Med. 2021, 384, 497–511. [Google Scholar] [CrossRef]
- Jayk Bernal, A.; Gomes da Silva, M.M.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; Delos Reyes, V.; Martin-Quiros, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L.; et al. Molnupiravir for Oral Treatment of Covid-19 in Nonhospitalized Patients. N. Engl. J. Med. 2022, 386, 509–520. [Google Scholar] [CrossRef]
- Pfizer’s Novel. Available online: https://www.pfizer.com/news/press-release/press-release-detail/pfizers-novel-covid-19-oral-antiviral-treatment-candidate#.YnIDOcZfvMY.link<EUA (accessed on 30 December 2021).
- Schaffer DeRoo, S.; Pudalov, N.J.; Fu, L.Y. Planning for a COVID-19 Vaccination Program. JAMA 2020, 323, 2458–2459. [Google Scholar] [CrossRef]
- Novel Coronavirus Landscape COVID-19. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 8 January 2021).
- Hansson, M.; Nygren, P.A.k.; Ståhl, S. Design and production of recombinant subunit vaccines. Biotechnol. Appl. Biochem. 2000, 32, 95–107. [Google Scholar] [CrossRef]
- Noad, R.; Roy, P. Virus-like particles as immunogens. Trends Microbiol. 2003, 11, 438–444. [Google Scholar] [CrossRef]
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Tok, T.T.; Tatar, G. Structures and functions of coronavirus proteins: Molecular modeling of viral nucleoprotein. Int. J. Virol. Infect. Dis. 2017, 2, 001–007. [Google Scholar]
- Woo, P.C.; Huang, Y.; Lau, S.K.; Yuen, K.Y. Coronavirus genomics and bioinformatics analysis. Viruses 2010, 2, 1804–1820. [Google Scholar] [CrossRef]
- Ziebuhr, J. The coronavirus replicase. Curr. Top. Microbiol. Immunol. 2005, 287, 57–94. [Google Scholar]
- Bárcena, M.; Oostergetel, G.T.; Bartelink, W.; Faas, F.G.; Verkleij, A.; Rottier, P.J.; Koster, A.J.; Bosch, B.J. Cryo-electron tomography of mouse hepatitis virus: Insights into the structure of the coronavirion. Proc. Natl. Acad. Sci. USA 2009, 106, 582–587. [Google Scholar] [CrossRef]
- Song, H.C.; Seo, M.Y.; Stadler, K.; Yoo, B.J.; Choo, Q.L.; Coates, S.R.; Uematsu, Y.; Harada, T.; Greer, C.E.; Polo, J.M.; et al. Synthesis and characterization of a native, oligomeric form of recombinant severe acute respiratory syndrome coronavirus spike glycoprotein. J. Virol. 2004, 78, 10328–10335. [Google Scholar] [CrossRef]
- Beniac, D.R.; Andonov, A.; Grudeski, E.; Booth, T.F. Architecture of the SARS coronavirus prefusion spike. Nat. Struct. Mol. Biol. 2006, 13, 751–752. [Google Scholar] [CrossRef]
- Millet, J.K.; Jaimes, J.A.; Whittaker, G.R. Molecular diversity of coronavirus host cell entry receptors. FEMS Microbiol. Rev. 2021, 45, fuaa057. [Google Scholar] [CrossRef]
- Li, F. Receptor recognition mechanisms of coronaviruses: A decade of structural studies. J. Virol. 2015, 89, 1954–1964. [Google Scholar] [CrossRef]
- Li, F. Structure, Function, and Evolution of Coronavirus Spike Proteins. Annu Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, M.L.; Chien, C.S.; Yarmishyn, A.A.; Yang, Y.P.; Lai, W.Y.; Luo, Y.H.; Lin, Y.T.; Chen, Y.J.; Chang, P.C.; et al. Highlight of Immune Pathogenic Response and Hematopathologic Effect in SARS-CoV, MERS-CoV, and SARS-Cov-2 Infection. Front. Immunol. 2020, 11, 1022. [Google Scholar] [CrossRef]
- Lu, G.; Wang, Q.; Gao, G.F. Bat-to-human: Spike features determining ‘host jump’ of coronaviruses SARS-CoV, MERS-CoV, and beyond. Trends Microbiol. 2015, 23, 468–478. [Google Scholar] [CrossRef]
- De Haan, C.A.; Kuo, L.; Masters, P.S.; Vennema, H.; Rottier, P.J. Coronavirus particle assembly: Primary structure requirements of the membrane protein. J. Virol. 1998, 72, 6838–6850. [Google Scholar] [CrossRef]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef]
- Niemann, H.; Geyer, R.; Klenk, H.; Linder, D.; Stirm, S.; Wirth, M. The carbohydrates of mouse hepatitis virus (MHV) A59: Structures of the O-glycosidically linked oligosaccharides of glycoprotein E1. EMBO J. 1984, 3, 665–670. [Google Scholar] [CrossRef]
- Holmes, K.V.; Doller, E.W.; Sturman, L.S. Tunicamycin resistant glycosylation of a coronavirus glycoprotein: Demonstration of a novel type of viral glycoprotein. Virology 1981, 115, 334–344. [Google Scholar] [CrossRef]
- Wissink, E.H.J.; Kroese, M.V.; Maneschijn-Bonsing, J.G.; Meulenberg, J.J.M.; van Rijn, P.A.; Rijsewijk, F.A.M.; Rottier, P.J.M. Significance of the oligosaccharides of the porcine reproductive and respiratory syndrome virus glycoproteins GP2a and GP5 for infectious virus production. J. Gen. Virol. 2004, 85, 3715–3723. [Google Scholar] [CrossRef] [PubMed]
- de Haan, C.A.M.; de Wit, M.; Kuo, L.; Montalto-Morrison, C.; Haagmans, B.L.; Weiss, S.R.; Masters, P.S.; Rottier, P.J.M. The glycosylation status of the murine hepatitis coronavirus M protein affects the interferogenic capacity of the virus in vitro and its ability to replicate in the liver but not the brain. Virology 2003, 312, 395–406. [Google Scholar] [CrossRef]
- Narayanan, K.; Makino, S. Characterization of nucleocapsid-M protein interaction in murine coronavirus. Adv. Exp. Med. Biol. 2001, 494, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Cagliani, R.; Forni, D.; Clerici, M.; Sironi, M. Computational Inference of Selection Underlying the Evolution of the Novel Coronavirus, Severe Acute Respiratory Syndrome Coronavirus 2. J. Virol. 2020, 94, e00411-20. [Google Scholar] [CrossRef]
- Hu, Y.; Wen, J.; Tang, L.; Zhang, H.; Zhang, X.; Li, Y.; Wang, J.; Han, Y.; Li, G.; Shi, J.; et al. The M Protein of SARS-CoV: Basic Structural and Immunological Properties. Genom. Proteom. Bioinform. 2003, 1, 118–130. [Google Scholar] [CrossRef]
- Raamsman, M.J.; Locker, J.K.; De Hooge, A.; De Vries, A.A.; Griffiths, G.; Vennema, H.; Rottier, P.J. Characterization of the coronavirus mouse hepatitis virus strain A59 small membrane protein E. J. Virol. 2000, 74, 2333–2342. [Google Scholar] [CrossRef]
- Thiel, V.; Siddell, S.G. Internal ribosome entry in the coding region of murine hepatitis virus mRNA 5. J. Gen. Virol. 1994, 75, 3041–3046. [Google Scholar] [CrossRef]
- Ruch, T.R.; Machamer, C.E. The coronavirus E protein: Assembly and beyond. Viruses 2012, 4, 363–382. [Google Scholar] [CrossRef]
- DeDiego, M.L.; Nieto-Torres, J.L.; Jimenez-Guardeno, J.M.; Regla-Nava, J.A.; Alvarez, E.; Oliveros, J.C.; Zhao, J.; Fett, C.; Perlman, S.; Enjuanes, L. Severe acute respiratory syndrome coronavirus envelope protein regulates cell stress response and apoptosis. PLoS Pathog. 2011, 7, e1002315. [Google Scholar] [CrossRef]
- Grunewald, M.E.; Fehr, A.R.; Athmer, J.; Perlman, S. The coronavirus nucleocapsid protein is ADP-ribosylated. Virology 2018, 517, 62–68. [Google Scholar] [CrossRef]
- Zuwala, K.; Golda, A.; Kabala, W.; Burmistrz, M.; Zdzalik, M.; Nowak, P.; Kedracka-Krok, S.; Zarebski, M.; Dobrucki, J.; Florek, D.; et al. The nucleocapsid protein of human coronavirus NL63. PLoS ONE 2015, 10, e0117833. [Google Scholar] [CrossRef]
- Stertz, S.; Reichelt, M.; Spiegel, M.; Kuri, T.; Martinez-Sobrido, L.; Garcia-Sastre, A.; Weber, F.; Kochs, G. The intracellular sites of early replication and budding of SARS-coronavirus. Virology 2007, 361, 304–315. [Google Scholar] [CrossRef]
- McBride, R.; van Zyl, M.; Fielding, B.C. The coronavirus nucleocapsid is a multifunctional protein. Viruses 2014, 6, 2991–3018. [Google Scholar] [CrossRef]
- Chen, Z.; Pei, D.; Jiang, L.; Song, Y.; Wang, J.; Wang, H.; Zhou, D.; Zhai, J.; Du, Z.; Li, B.; et al. Antigenicity analysis of different regions of the severe acute respiratory syndrome coronavirus nucleocapsid protein. Clin. Chem. 2004, 50, 988–995. [Google Scholar] [CrossRef]
- Hu, H.; Tao, L.; Wang, Y.; Chen, L.; Yang, J.; Wang, H. Enhancing immune responses against SARS-CoV nucleocapsid DNA vaccine by co-inoculating interleukin-2 expressing vector in mice. Biotechnol. Lett. 2009, 31, 1685–1693. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Tsoi, H.W.; Chen, Z.W.; Wong, B.H.; Zhang, L.; Chan, J.K.; Wong, L.P.; He, W.; Ma, C.; et al. SARS coronavirus spike polypeptide DNA vaccine priming with recombinant spike polypeptide from Escherichia coli as booster induces high titer of neutralizing antibody against SARS coronavirus. Vaccine 2005, 23, 4959–4968. [Google Scholar] [CrossRef]
- Wang, M.; Fu, T.; Hao, J.; Li, L.; Tian, M.; Jin, N.; Ren, L.; Li, C. A recombinant Lactobacillus plantarum strain expressing the spike protein of SARS-CoV-2. Int. J. Biol. Macromol. 2020, 160, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.M.; Al-Amri, S.S.; Al-Subhi, T.L.; Siddiq, L.A.; Hassan, A.M.; Alawi, M.M.; Alhabbab, R.Y.; Hindawi, S.I.; Mohammed, O.B.; Amor, N.S.; et al. Development and validation of different indirect ELISAs for MERS-CoV serological testing. J. Immunol. Methods 2019, 466, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, E.G.; Miao, C.; Haupt, T.E.; Anderson, L.J.; Haynes, L.M. Development of a recombinant truncated nucleocapsid protein based immunoassay for detection of antibodies against human coronavirus OC43. J. Virol. Methods 2011, 177, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Lee, B.H.; Seok, S.H.; Baek, M.W.; Lee, H.Y.; Kim, D.J.; Na, Y.R.; Noh, K.J.; Park, S.H.; Kumar, D.N.; et al. Production of specific antibodies against SARS-coronavirus nucleocapsid protein without cross reactivity with human coronaviruses 229E and OC43. J. Vet. Sci. 2010, 11, 165–167. [Google Scholar] [CrossRef]
- Zou, N.; Wang, F.; Duan, Z.; Xia, J.; Wen, X.; Yan, Q.; Liu, P.; Cao, S.; Huang, Y. Development and characterization of neutralizing monoclonal antibodies against the S1 subunit protein of QX-like avian infectious bronchitis virus strain Sczy3. Monoclon. Antib. Immunodiagn. Immunother. 2015, 34, 17–24. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Wei, F.; Dong, Y.; Zhu, L.; Han, W.; Wang, L.; Shen, Z. Prokaryotic Expression of Truncated S1 Protein of Porcine Epidemic Diarrhea Virus and Production of Monoclonal Antibodies to Recombinant Protein. Monoclon. Antib. Immunodiagn. Immunother. 2015, 34, 327–333. [Google Scholar] [CrossRef]
- Luo, S.X.; Fan, J.H.; Opriessnig, T.; Di, J.M.; Liu, B.J.; Zuo, Y.Z. Development and application of a recombinant M protein-based indirect ELISA for the detection of porcine deltacoronavirus IgG antibodies. J. Virol. Methods 2017, 249, 76–78. [Google Scholar] [CrossRef]
- Elia, G.; Fiermonte, G.; Pratelli, A.; Martella, V.; Camero, M.; Cirone, F.; Buonavoglia, C. Recombinant M protein-based ELISA test for detection of antibodies to canine coronavirus. J. Virol. Methods 2003, 109, 139–142. [Google Scholar] [CrossRef]
- Su, M.; Li, C.; Guo, D.; Wei, S.; Wang, X.; Geng, Y.; Yao, S.; Gao, J.; Wang, E.; Zhao, X.; et al. A recombinant nucleocapsid protein-based indirect enzyme-linked immunosorbent assay to detect antibodies against porcine deltacoronavirus. J. Vet. Med. Sci. 2016, 78, 601–606. [Google Scholar] [CrossRef]
- Finger, P.F.; Pepe, M.S.; Dummer, L.A.; Magalhaes, C.G.; de Castro, C.C.; de Oliveira Hubner, S.; Leite, F.P.L.; Ritterbusch, G.A.; Esteves, P.A.; Conceicao, F.R. Combined use of ELISA and Western blot with recombinant N protein is a powerful tool for the immunodiagnosis of avian infectious bronchitis. Virol. J. 2018, 15, 189. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Kamble, N.M.; Pillai, A.S.; Gaikwad, S.S.; Khulape, S.A.; Reddy, M.R.; Mohan, C.M.; Kataria, J.M.; Dey, S. Recombinant nucleocapsid protein based single serum dilution ELISA for the detection of antibodies to infectious bronchitis virus in poultry. J. Virol. Methods 2014, 209, 1–6. [Google Scholar] [CrossRef]
- Abdelwahab, M.; Loa, C.C.; Wu, C.C.; Lin, T.L. Recombinant nucleocapsid protein-based enzyme-linked immunosorbent assay for detection of antibody to turkey coronavirus. J. Virol. Methods 2015, 217, 36–41. [Google Scholar] [CrossRef]
- Chen, W.H.; Du, L.; Chag, S.M.; Ma, C.; Tricoche, N.; Tao, X.; Seid, C.A.; Hudspeth, E.M.; Lustigman, S.; Tseng, C.T.; et al. Yeast-expressed recombinant protein of the receptor-binding domain in SARS-CoV spike protein with deglycosylated forms as a SARS vaccine candidate. Hum. Vaccin. Immunother. 2014, 10, 648–658. [Google Scholar] [CrossRef]
- Chen, W.H.; Chag, S.M.; Poongavanam, M.V.; Biter, A.B.; Ewere, E.A.; Rezende, W.; Seid, C.A.; Hudspeth, E.M.; Pollet, J.; McAtee, C.P.; et al. Optimization of the Production Process and Characterization of the Yeast-Expressed SARS-CoV Recombinant Receptor-Binding Domain (RBD219-N1), a SARS Vaccine Candidate. J. Pharm. Sci. 2017, 106, 1961–1970. [Google Scholar] [CrossRef]
- Chuck, C.P.; Wong, C.H.; Chow, L.M.; Fung, K.P.; Waye, M.M.; Tsui, S.K. Expression of SARS-coronavirus spike glycoprotein in Pichia pastoris. Virus Genes 2009, 38, 1–9. [Google Scholar] [CrossRef]
- Liu, R.-S.; Yang, K.-Y.; Lin, J.; Lin, Y.-W.; Zhang, Z.-H.; Zhang, J.; Xia, N.-S. High-yield expression of recombinant SARS coronavirus nucleocapsid protein in methylotrophic yeast Pichia pastoris. World J. Gastroenterol. WJG 2004, 10, 3602. [Google Scholar] [CrossRef]
- Pogrebnyak, N.; Golovkin, M.; Andrianov, V.; Spitsin, S.; Smirnov, Y.; Egolf, R.; Koprowski, H. Severe acute respiratory syndrome (SARS) S protein production in plants: Development of recombinant vaccine. Proc. Natl. Acad. Sci. USA 2005, 102, 9062–9067. [Google Scholar] [CrossRef]
- Li, H.-Y.; Ramalingam, S.; Chye, M.-L. Accumulation of recombinant SARS-CoV spike protein in plant cytosol and chloroplasts indicate potential for development of plant-derived oral vaccines. Exp. Biol. Med. 2006, 231, 1346–1352. [Google Scholar] [CrossRef]
- Zheng, N.; Xia, R.; Yang, C.; Yin, B.; Li, Y.; Duan, C.; Liang, L.; Guo, H.; Xie, Q. Boosted expression of the SARS-CoV nucleocapsid protein in tobacco and its immunogenicity in mice. Vaccine 2009, 27, 5001–5007. [Google Scholar] [CrossRef]
- Gomez, N.; Wigdorovitz, A.; Castanon, S.; Gil, F.; Ordá, R.; Borca, M.; Escribano, J. Oral immunogenicity of the plant derived spike protein from swine-transmissible gastroenteritis coronavirus. Arch. Virol. 2000, 145, 1725–1732. [Google Scholar] [CrossRef]
- Tien, N.Q.; Huy, N.X.; Kim, M.Y. Improved expression of porcine epidemic diarrhea antigen by fusion with cholera toxin B subunit and chloroplast transformation in Nicotiana tabacum. Plant Cell Tissue Organ Cult. 2019, 137, 213–223. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Massa, S.; Illiano, E.; De Martinis, D.; Chan, P.K.; Di Bonito, P.; Franconi, R. Antigen Production in Plant to Tackle Infectious Diseases Flare Up: The Case of SARS. Front. Plant Sci. 2016, 7, 54. [Google Scholar] [CrossRef]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef]
- Tai, W.; Wang, Y.; Fett, C.A.; Zhao, G.; Li, F.; Perlman, S.; Jiang, S.; Zhou, Y.; Du, L. Recombinant Receptor-Binding Domains of Multiple Middle East Respiratory Syndrome Coronaviruses (MERS-CoVs) Induce Cross-Neutralizing Antibodies against Divergent Human and Camel MERS-CoVs and Antibody Escape Mutants. J. Virol. 2017, 91, e01651-16. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Erdos, G.; Huang, S.; Kenniston, T.W.; Balmert, S.C.; Carey, C.D.; Raj, V.S.; Epperly, M.W.; Klimstra, W.B.; Haagmans, B.L.; et al. Microneedle array delivered recombinant coronavirus vaccines: Immunogenicity and rapid translational development. EBioMedicine 2020, 55, 102743. [Google Scholar] [CrossRef] [PubMed]
- Elshabrawy, H.A.; Coughlin, M.M.; Baker, S.C.; Prabhakar, B.S. Human monoclonal antibodies against highly conserved HR1 and HR2 domains of the SARS-CoV spike protein are more broadly neutralizing. PLoS ONE 2012, 7, e50366. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Shi, M.; Li, J.; Song, P.; Li, N. Construction of SARS-CoV-2 Virus-Like Particles by Mammalian Expression System. Front. Bioeng. Biotechnol. 2020, 8, 862. [Google Scholar] [CrossRef]
- Kuo, T.Y.; Lin, M.Y.; Coffman, R.L.; Campbell, J.D.; Traquina, P.; Lin, Y.J.; Liu, L.T.; Cheng, J.; Wu, Y.C.; Wu, C.C.; et al. Development of CpG-adjuvanted stable prefusion SARS-CoV-2 spike antigen as a subunit vaccine against COVID-19. Sci. Rep. 2020, 10, 20085. [Google Scholar] [CrossRef]
- Hsieh, S.M.; Liu, W.D.; Huang, Y.S.; Lin, Y.J.; Hsieh, E.F.; Lian, W.C.; Chen, C.; Janssen, R.; Shih, S.R.; Huang, C.G.; et al. Safety and immunogenicity of a Recombinant Stabilized Prefusion SARS-CoV-2 Spike Protein Vaccine (MVC-COV1901) Adjuvanted with CpG 1018 and Aluminum Hydroxide in healthy adults: A Phase 1, dose-escalation study. EClinicalMedicine 2021, 38, 100989. [Google Scholar] [CrossRef]
- Thachil, A.; Gerber, P.F.; Xiao, C.T.; Huang, Y.W.; Opriessnig, T. Development and application of an ELISA for the detection of porcine deltacoronavirus IgG antibodies. PLoS ONE 2015, 10, e0124363. [Google Scholar] [CrossRef]
- Esposito, D.; Mehalko, J.; Drew, M.; Snead, K.; Wall, V.; Taylor, T.; Frank, P.; Denson, J.P.; Hong, M.; Gulten, G.; et al. Optimizing high-yield production of SARS-CoV-2 soluble spike trimers for serology assays. Protein Expr. Purif. 2020, 174, 105686. [Google Scholar] [CrossRef]
- Mehalko, J.; Drew, M.; Snead, K.; Denson, J.P.; Wall, V.; Taylor, T.; Sadtler, K.; Messing, S.; Gillette, W.; Esposito, D. Improved production of SARS-CoV-2 spike receptor-binding domain (RBD) for serology assays. Protein Expr. Purif. 2021, 179, 105802. [Google Scholar] [CrossRef]
- Siu, Y.L.; Teoh, K.T.; Lo, J.; Chan, C.M.; Kien, F.; Escriou, N.; Tsao, S.W.; Nicholls, J.M.; Altmeyer, R.; Peiris, J.S.; et al. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J. Virol. 2008, 82, 11318–11330. [Google Scholar] [CrossRef]
- Li, J.; Ulitzky, L.; Silberstein, E.; Taylor, D.R.; Viscidi, R. Immunogenicity and protection efficacy of monomeric and trimeric recombinant SARS coronavirus spike protein subunit vaccine candidates. Viral. Immunol. 2013, 26, 126–132. [Google Scholar] [CrossRef]
- Coleman, C.M.; Liu, Y.V.; Mu, H.; Taylor, J.K.; Massare, M.; Flyer, D.C.; Smith, G.E.; Frieman, M.B. Purified coronavirus spike protein nanoparticles induce coronavirus neutralizing antibodies in mice. Vaccine 2014, 32, 3169–3174. [Google Scholar] [CrossRef]
- Fujita, R.; Hino, M.; Ebihara, T.; Nagasato, T.; Masuda, A.; Lee, J.M.; Fujii, T.; Mon, H.; Kakino, K.; Nagai, R.; et al. Efficient production of recombinant SARS-CoV-2 spike protein using the baculovirus-silkworm system. Biochem. Biophys. Res. Commun. 2020, 529, 257–262. [Google Scholar] [CrossRef]
- Tian, J.H.; Patel, N.; Haupt, R.; Zhou, H.; Weston, S.; Hammond, H.; Logue, J.; Portnoff, A.D.; Norton, J.; Guebre-Xabier, M.; et al. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice. Nat. Commun. 2021, 12, 372. [Google Scholar] [CrossRef]
- Keech, C.; Glenn, G.M.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; et al. First-in-Human Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef]
- Hsu, C.W.; Chang, M.H.; Chang, H.W.; Wu, T.Y.; Chang, Y.C. Parenterally Administered Porcine Epidemic Diarrhea Virus-Like Particle-Based Vaccine Formulated with CCL25/28 Chemokines Induces Systemic and Mucosal Immune Protectivity in Pigs. Viruses 2020, 12, 1122. [Google Scholar] [CrossRef]
- Yilmaz, H.; Faburay, B.; Turan, N.; Cotton-Caballero, M.; Cetinkaya, B.; Gurel, A.; Yilmaz, A.; Cizmecigil, U.Y.; Aydin, O.; Tarakci, E.A.; et al. Production of Recombinant N Protein of Infectious Bronchitis Virus Using the Baculovirus Expression System and Its Assessment as a Diagnostic Antigen. Appl. Biochem. Biotechnol. 2019, 187, 506–517. [Google Scholar] [CrossRef]
- Severance, E.G.; Bossis, I.; Dickerson, F.B.; Stallings, C.R.; Origoni, A.E.; Sullens, A.; Yolken, R.H.; Viscidi, R.P. Development of a nucleocapsid-based human coronavirus immunoassay and estimates of individuals exposed to coronavirus in a U.S. metropolitan population. Clin. Vaccine Immunol. 2008, 15, 1805–1810. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Takami, Y.; Kumar Deo, V.; Park, E.Y. Preparation of virus-like particle mimetic nanovesicles displaying the S protein of Middle East respiratory syndrome coronavirus using insect cells. J. Biotechnol. 2019, 306, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Ko, H.L.; Lee, E.Y.; Park, H.J.; Kim, Y.S.; Kim, Y.S.; Cho, N.H.; Park, M.S.; Lee, S.M.; Kim, J.; et al. Development of a diagnostic system for detection of specific antibodies and antigens against Middle East respiratory syndrome coronavirus. Microbiol. Immunol. 2018, 62, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Liu, C.; Yount, B.; Gully, K.; Yang, Y.; Auerbach, A.; Peng, G.; Baric, R.; Li, F. Structure of mouse coronavirus spike protein complexed with receptor reveals mechanism for viral entry. PLoS Pathog. 2020, 16, e1008392. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef]
- Cohen, S.N.; Chang, A.C.; Boyer, H.W.; Helling, R.B. Construction of biologically functional bacterial plasmids in vitro. Proc. Natl. Acad. Sci. USA 1973, 70, 3240–3244. [Google Scholar] [CrossRef]
- Ouranidis, A.; Davidopoulou, C.; Tashi, R.-K.; Kachrimanis, K. Pharma 4.0 Continuous MRNA Drug Products Manufacturing. Pharmaceutics 2021, 13, 1371. [Google Scholar] [CrossRef]
- Baeshen, N.A.; Baeshen, M.N.; Sheikh, A.; Bora, R.S.; Ahmed, M.M.M.; Ramadan, H.A.; Saini, K.S.; Redwan, E.M. Cell factories for insulin production. Microb. Cell Factories 2014, 13, 1–9. [Google Scholar] [CrossRef]
- Krzeslak, J.; Braun, P.; Voulhoux, R.; Cool, R.H.; Quax, W.J. Heterologous production of Escherichia coli penicillin G acylase in Pseudomonas aeruginosa. J. Biotechnol. 2009, 142, 250–258. [Google Scholar] [CrossRef]
- Schleicher, L.; Muras, V.; Claussen, B.; Pfannstiel, J.; Blombach, B.; Dibrov, P.; Fritz, G.; Steuber, J. Vibrio natriegens as Host for Expression of Multisubunit Membrane Protein Complexes. Front. Microbiol. 2018, 9, 2537. [Google Scholar] [CrossRef]
- Chen, R. Bacterial expression systems for recombinant protein production: E. coli and beyond. Biotechnol. Adv. 2012, 30, 1102–1107. [Google Scholar] [CrossRef]
- Cai, D.; Rao, Y.; Zhan, Y.; Wang, Q.; Chen, S. Engineering Bacillus for efficient production of heterologous protein: Current progress, challenge and prospect. J. Appl. Microbiol. 2019, 126, 1632–1642. [Google Scholar] [CrossRef]
- Cui, W.; Han, L.; Suo, F.; Liu, Z.; Zhou, L.; Zhou, Z. Exploitation of Bacillus subtilis as a robust workhorse for production of heterologous proteins and beyond. World J. Microbiol. Biotechnol. 2018, 34, 145. [Google Scholar] [CrossRef]
- Morello, E.; Bermudez-Humaran, L.G.; Llull, D.; Sole, V.; Miraglio, N.; Langella, P.; Poquet, I. Lactococcus lactis, an efficient cell factory for recombinant protein production and secretion. J. Mol. Microbiol. Biotechnol. 2008, 14, 48–58. [Google Scholar] [CrossRef]
- Yzturk, S.; Yalik, P.; Yzdamar, T.H. Fed-Batch Biomolecule Production by Bacillus subtilis: A State of the Art Review. Trends Biotechnol. 2016, 34, 329–345. [Google Scholar] [CrossRef]
- Kang, Z.; Yang, S.; Du, G.; Chen, J. Molecular engineering of secretory machinery components for high-level secretion of proteins in Bacillus species. J. Ind. Microbiol. Biotechnol. 2014, 41, 1599–1607. [Google Scholar] [CrossRef]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef]
- Nielsen, J. Production of biopharmaceutical proteins by yeast: Advances through metabolic engineering. Bioengineered 2013, 4, 207–211. [Google Scholar] [CrossRef]
- Baghban, R.; Farajnia, S.; Rajabibazl, M.; Ghasemi, Y.; Mafi, A.; Hoseinpoor, R.; Rahbarnia, L.; Aria, M. Yeast Expression Systems: Overview and Recent Advances. Mol. Biotechnol. 2019, 61, 365–384. [Google Scholar] [CrossRef]
- Vieira Gomes, A.M.; Souza Carmo, T.; Silva Carvalho, L.; Mendonca Bahia, F.; Parachin, N.S. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef]
- Kim, H.; Yoo, S.J.; Kang, H.A. Yeast synthetic biology for the production of recombinant therapeutic proteins. FEMS Yeast Res. 2015, 15, 1–16. [Google Scholar] [CrossRef]
- Ogata, K.; Nishikawa, H.; Ohsugi, M. A Yeast Capable of Utilizing Methanol. Agric. Biol. Chem. 2014, 33, 1519–1520. [Google Scholar] [CrossRef]
- Cereghino, J.L.; Cregg, J.M. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Gasser, B.; Mattanovich, D. A yeast for all seasons—Is Pichia pastoris a suitable chassis organism for future bioproduction? FEMS Microbiol. Lett. 2018, 365, fny181. [Google Scholar] [CrossRef]
- Tran, A.M.; Nguyen, T.T.; Nguyen, C.T.; Huynh-Thi, X.M.; Nguyen, C.T.; Trinh, M.T.; Tran, L.T.; Cartwright, S.P.; Bill, R.M.; Tran-Van, H. Pichia pastoris versus Saccharomyces cerevisiae: A case study on the recombinant production of human granulocyte-macrophage colony-stimulating factor. BMC Res. Notes 2017, 10, 148. [Google Scholar] [CrossRef]
- Chen, W.H.; Hotez, P.J.; Bottazzi, M.E. Potential for developing a SARS-CoV receptor-binding domain (RBD) recombinant protein as a heterologous human vaccine against coronavirus infectious disease (COVID)-19. Hum. Vaccin Immunother. 2020, 16, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Barta, A.; Sommergruber, K.; Thompson, D.; Hartmuth, K.; Matzke, M.A.; Matzke, A.J. The expression of a nopaline synthase—human growth hormone chimaeric gene in transformed tobacco and sunflower callus tissue. Plant Mol. Biol. 1986, 6, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P. Plant-based vaccines against viruses. Virol. J. 2014, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sijmons, P.C.; Dekker, B.M.; Schrammeijer, B.; Verwoerd, T.C.; Van Den Elzen, P.J.; Hoekema, A. Production of correctly processed human serum albumin in transgenic plants. Bio/Technology 1990, 8, 217–221. [Google Scholar] [CrossRef]
- Mason, H.S.; Lam, D.; Arntzen, C.J. Expression of hepatitis B surface antigen in transgenic plants. Proc. Natl. Acad. Sci. USA 1992, 89, 11745–11749. [Google Scholar] [CrossRef]
- Usha, R.; Rohll, J.B.; Spall, V.E.; Shanks, M.; Maule, A.J.; Johnson, J.E.; Lomonossoff, G.P. Expression of an animal virus antigenic site on the surface of a plant virus particle. Virology 1993, 197, 366–374. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Malla, A.; Phoolcharoen, W. Emergence of Novel Coronavirus 2019-nCoV: Need for Rapid Vaccine and Biologics Development. Pathogens 2020, 9, 148. [Google Scholar] [CrossRef]
- Moon, K.B.; Park, J.S.; Park, Y.I.; Song, I.J.; Lee, H.J.; Cho, H.S.; Jeon, J.H.; Kim, H.S. Development of Systems for the Production of Plant-Derived Biopharmaceuticals. Plants 2019, 9, 30. [Google Scholar] [CrossRef]
- Scherer, W.F.; Syverton, J.T.; Gey, G.O. Studies on the propagation in vitro of poliomyelitis viruses: IV. Viral. multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. J. Exp. Med. 1953, 97, 695–710. [Google Scholar] [CrossRef]
- Dumont, J.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R. Human cell lines for biopharmaceutical manufacturing: History, status, and future perspectives. Crit. Rev. Biotechnol. 2016, 36, 1110–1122. [Google Scholar] [CrossRef]
- Pellicer, A.; Wigler, M.; Axel, R.; Silverstein, S. The transfer and stable integration of the HSV thymidine kinase gene into mouse cells. Cell 1978, 14, 133–141. [Google Scholar] [CrossRef]
- Hacker, D.L.; Wurn, F.M. Protein Production in Mammalian Cells. eLS 2007, 1–5. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Shrivastava, A. Recent Developments in Bioprocessing of Recombinant Proteins: Expression Hosts and Process Development. Front. Bioeng. Biotechnol. 2019, 7, 420. [Google Scholar] [CrossRef]
- Owczarek, B.; Gerszberg, A.; Hnatuszko-Konka, K. A Brief Reminder of Systems of Production and Chromatography-Based Recovery of Recombinant Protein Biopharmaceuticals. Biomed. Res. Int. 2019, 2019, 4216060. [Google Scholar] [CrossRef]
- Khan, K.H. Gene expression in Mammalian cells and its applications. Adv. Pharm. Bull. 2013, 3, 257–263. [Google Scholar] [CrossRef]
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R.; et al. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef]
- Ikeda, M.; Hamajima, R.; Kobayashi, M. Baculoviruses: Diversity, evolution and manipulation of insects. Entomol. Sci. 2015, 18, 1–20. [Google Scholar] [CrossRef]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]
- Summers, M.D. Milestones Leading to the Genetic Engineering of Baculoviruses as Expression Vector Systems and Viral. Pesticides. Insect Viruses Biotechnol. Appl. 2006, 68, 3–73. [Google Scholar]
- Smith, G.E.; Summers, M.D.; Fraser, M. Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol. Cell. Biol. 1983, 3, 2156–2165. [Google Scholar] [PubMed]
- van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus-insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-J.; Jinn, T.R.; Chen, Y.-J.; Deng, M.-C.; Hwang, C.-S.; Hsieh, F.-C.; Kao, S.-S.; Chen, Y.-J.; Tung, K.-L.; Wu, T.Y.; et al. Vaccination with hemagglutinin produced in Trichoplusia ni larvae protects chickens against lethal H5N1 challenge. J. Taiwan Inst. Chem. Eng. 2011, 42, 223–227. [Google Scholar] [CrossRef]
- Ahrens, U.; Kaden, V.; Drexler, C.; Visser, N. Efficacy of the classical swine fever (CSF) marker vaccine Porcilis® Pesti in pregnant sows. Vet. Microbiol. 2000, 77, 83–97. [Google Scholar] [CrossRef]
- Harper, D.M. Impact of vaccination with Cervarix (trade mark) on subsequent HPV-16/18 infection and cervical disease in women 15-25 years of age. Gynecol. Oncol. 2008, 110, S11–S17. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).