1. Introduction
Membrane computing (MC) is an important branch of bio-inspired computing, which is initiated by Păun [
1], and the computing models of membrane computing are also called membrane systems or P systems. P systems focus on abstracting some fundamental concept from the structure and functioning of the living cells, cell tissues or colonies of cells. Research shows that some P systems have the same computing power as Turing machines, or are more efficient, to some extent [
2]. There are three classic computing models of P systems according to the structure of the membrane or cell arrangement in previous studies and researches, including cell-like P systems, tissue-like P systems and neural-like P systems [
3]. Many variants of P systems based on biological facts, mathematical biological cells, theoretical computer science or application motivations have been presented for solving difficult optimization problems in real life [
4,
5].
In general, several variants of P systems on the theoretical development of MC that are mostly based on three classic P systems that recruit various ingredients, including energy, catalysts and mitosis, have been reported in previous studies and related works [
6]. Many kinds of extended cell-like P systems have been reported in the literature, such as cell-like P systems with active membranes inspired in the mitosis process [
7,
8,
9], cell-like P systems with evolutional symport/antiport rules inspired by the conservation law [
10,
11], and so on. Similarly, tissue-like P systems with evolutional symport/antiport rules are a class of computing modes based on cell inter-communication in tissues [
6,
12,
13]. The interchange of objects between regions are determined by the existence of promoters or inhibitors, it is simply called tissue-like P systems with a promoter/inhibitor [
14,
15,
16]. The multiset rewriting rules are introduced to tissue-like P systems based on the energy assumption, which names homeostasis tissue-like P systems [
17,
18]. Spiking neural-like P systems (SN P) are an important computing model of neural-like P systems [
19]. Many variants of SN P systems have emerged, such as SN P systems with rules on synapses [
20], SN P systems with multiple channels [
21], SN P systems with anti-spikes [
22], and other made changes on communication rules [
23]. The analysis of computing power and computational efficiency for P systems and their variants is also an important part of studies and works [
24].
Evolutionary computation (EC) based on the fundamental principles of biological evolution is an important branch of natural computing [
25], which generally divided into two broad areas, evolutionary algorithms (EAs) and swarm intelligence (SI) [
26]. Especially, EAs are inspired by natural selection and molecular genetics, consisting of classic EAs and recently developed EAs [
27]. Classic EAs included genetic algorithms (GAs) [
28], evolution strategies (ES) [
29], evolutionary programming (EP) [
30] and genetic programming (GP) [
31]. Recently developed EAs contained a family of optimization algorithms, such as quantum-inspired evolutionary algorithms (QIEAS) [
32], simulated annealing (SA) [
33], differential evolution (DE) [
34], and tabu search (TS) [
35]. SI involved many kinds of meta-heuristic techniques, including particle swarm optimization (PSO) [
36], ant colony optimization (ACO) [
37], an artificial bee colony algorithm (ABC) [
38], biogeography-based optimization (BBO) [
39], and an artificial fish swarm algorithm [
40]; these techniques have been widely used to solve many complicated problems [
41].
The combination of MC and EC is an important application in studies of MC and EC [
42], which is named evolutionary membrane computing (EMC) [
43]. The parallel-distributed framework of MC and computing characteristics of EC, such as easy implementation, robust performance, are utilized to EMC in order to improve computation performance [
44]. In this respect, two wide research areas of EMC have been studied, including membrane-inspired evolutionary algorithms (MIEAs), also called membrane algorithms (MAs), and the automated design of membrane computing models (ADMCM) [
27]. Specially, MIEAs or MAs are outstanding instances in MC for approaching real-word applications [
45], which can be classified into two categories according to different membrane structure: hierarchical membrane structure from cell-like P systems and network membrane structure from tissue-like P systems and neural-like P systems [
46].
Furthermore, hierarchical structure based MIEAs, including a nested membrane structure (NMS) [
47], one level membrane structure (OLMS) [
48], hybrid membrane structure (HMS) [
49] and dynamic membrane structure (DMS) [
50], are the first kind of EMC. NMS-based MIEAs are designed to integrate cell-like P systems with different meta-heuristic techniques, such as GA [
51], DE [
52,
53], ACO [
54], and so on [
55,
56]. OLMS-based MIEAs are proposed to combine cell-like P systems and different algorithms in a membrane (AIM), such as GA [
57], DE [
58], PSO [
59,
60], ACO [
61], and QIEAS [
62]. Due to combination possibilities of a hybrid membrane structure, HMS-based MIEAs based on the membrane structure of NMS and OLMS are presented. However, the complexity and difficulty of a hybrid structure are limited to the development of HMS-based MIEAs [
63,
64]. The dynamic membrane structure of DMS-based MIEAs is different from the static membrane structure of NMS-based MIEAs, OLMS-based MIEAs and HMS-based MIEAs. Moreover, the hierarchical membrane structure of DMS-based MIEAs can be changed with active membranes during the evolution process [
65,
66].
The network structure based MIEAs, including the statistical network structure (SNS) and dynamical network structure (DNS), are the second kind of EMC [
67]. SNS-based MIEAs are presented by using tissue-like P systems or neural-like P systems with various network topologies [
68]. Various meta-heuristic algorithms, such as GA [
69], DE [
70] and its variants [
71,
72], PSO [
73,
74], ABC [
75], and BBO [
76], are usually introduced to SNS-based MIEAs as the basic evolutionary operation in the cell or neural [
77,
78,
79,
80,
81]. The membrane structure in DNS-based MIEAs can be dynamically changed according to communication channel rules, and this class of MIEAs, with an extended membrane structure, has great potential for solving complex problems [
82,
83]. For another kind of EMC, ADMCMs are designed to overcome the complex programmability of membrane-based modes, and the automated synthesis of computing models by applying various meta-heuristic methods can be obtained through this class of EMC [
84,
85,
86]. Three important research topics of ADMCM have been investigated, including abstract rewriting systems on multisets (ARSM) [
87], parameter optimization of P system models (POPSM) [
88] and automatic design of P systems (ADPS) [
89,
90,
91].
Quantum-behaved particle swarm optimization (QPSO) is a variant of PSO which was initiated by Sun in 2004 [
92], and the sampling space of QPSO covers the whole search space due to the probabilities of potential well models [
93]. It has been proved that the global search ability of QPSO has been improved more than classic PSO [
94]. Although QPSO has been shown a greater potential than traditional EC [
95], it still has some limitations in the sample test set, such as being easily trapped into local optima and exhibited prematurity [
96]. Tissue-like P systems are a kind of classic computing model in MC, and the underlying membrane structure of tissue-like P systems can be abstracted to an arbitrary graph in mathematics compared with cell-like P systems [
97]. The parallel-distributed framework, particularly the membrane structure, and evolution–communication mechanism of tissue-like P systems can be used to improve the performance of EC. Therefore, the tissue-like P system with evolutional symport/antiport rules and a promoter/inhibitor is introduced to enhance the optimization performance of QPSO and to overcome the limitation as we mentioned above.
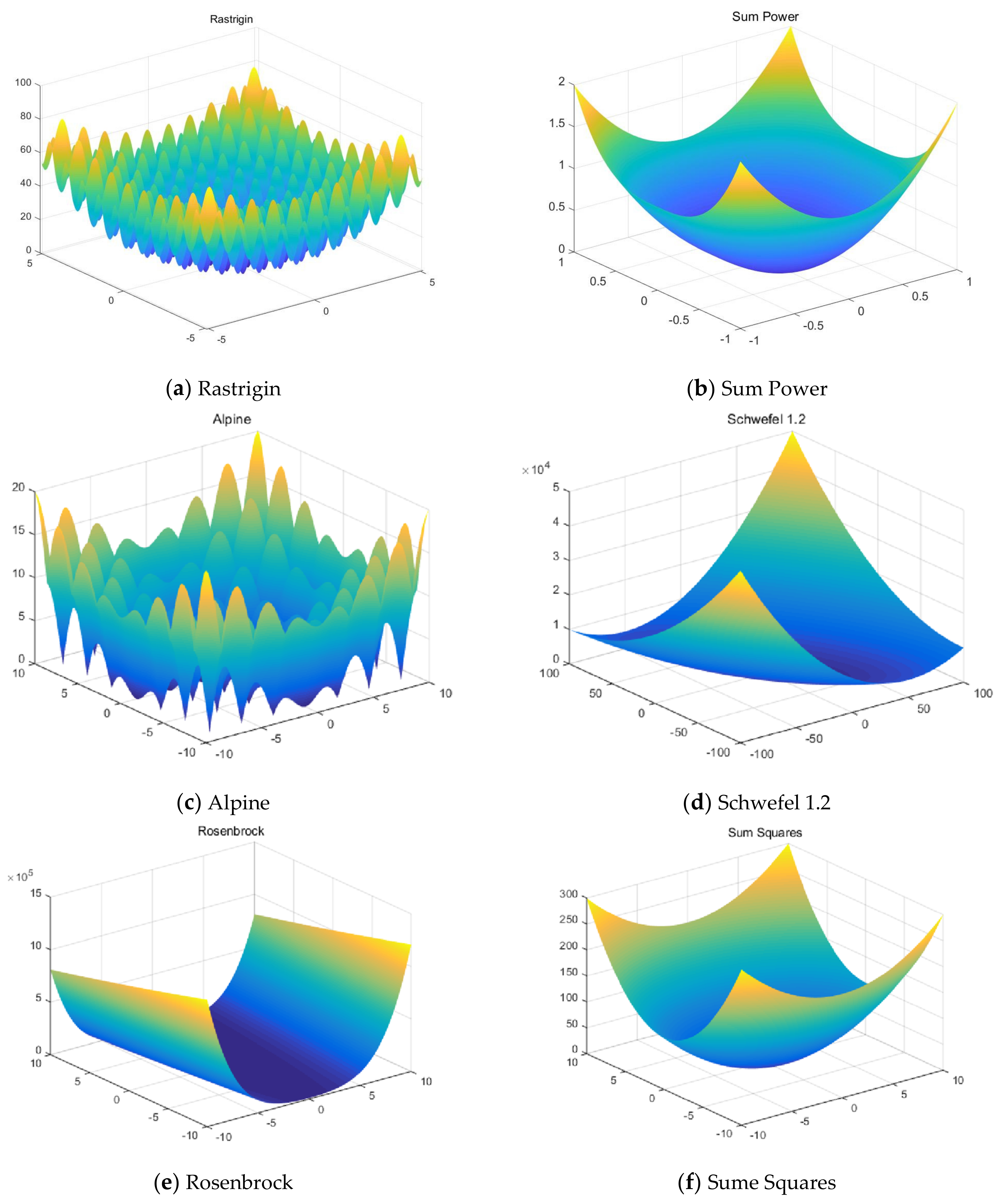
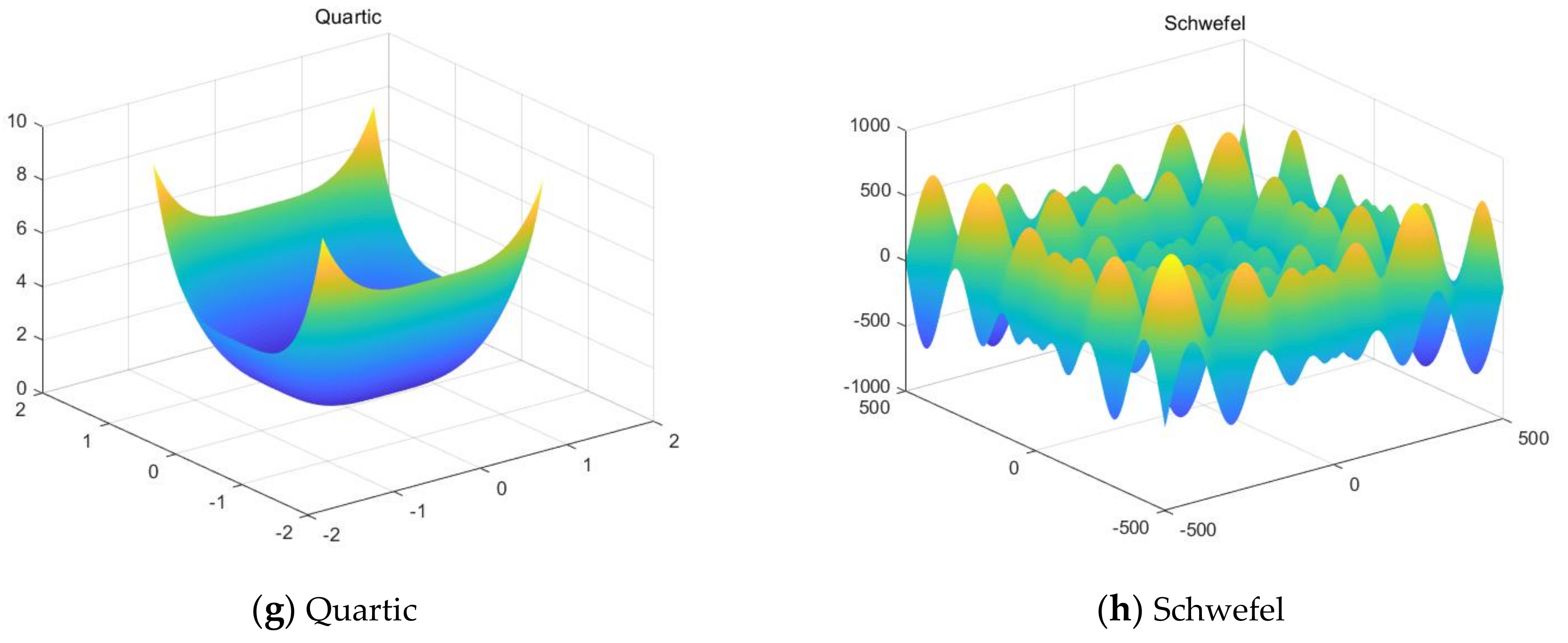
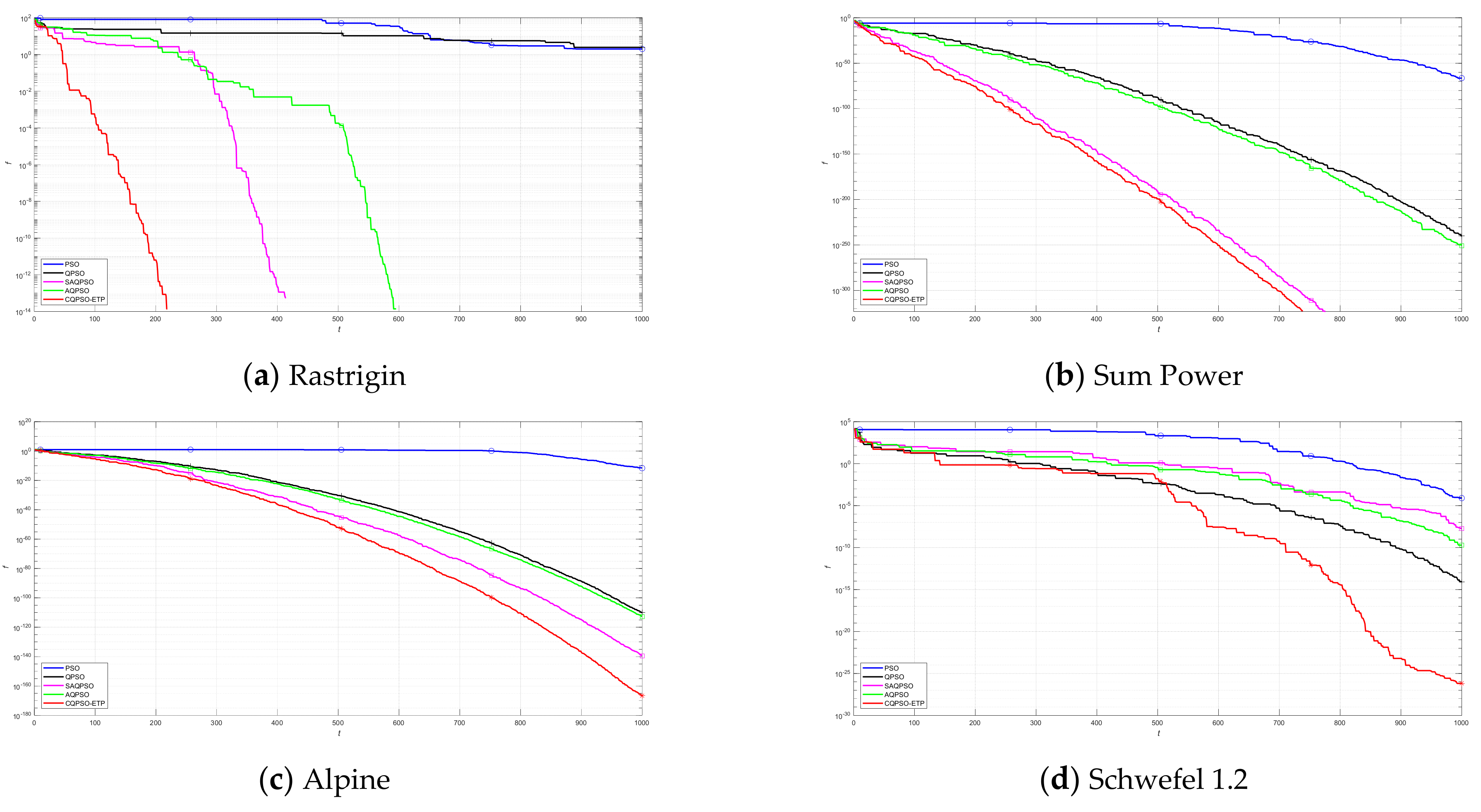
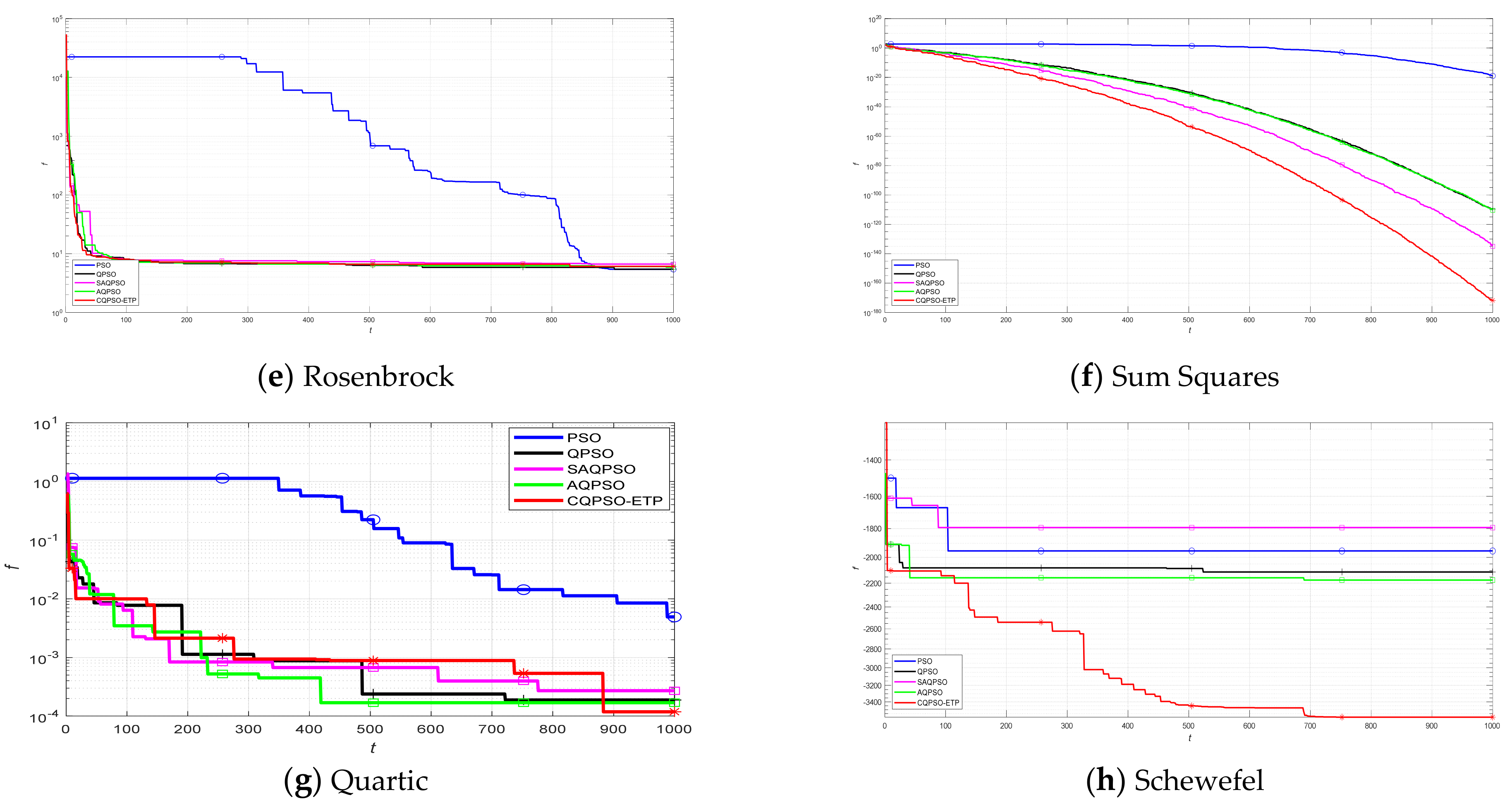
This work focuses on the development of a membrane computing model in SNS-based MIEAs to solve some optimization problems and image segmentation problems, which is based on the evolution mechanism of QPSO and improved QPSO, and the evolution–communication mechanism of the tissue-like P system. An extended tissue-like P system based on the evolution–communication mechanism of the tissue-like P system, with evolutional symport/antiport rules and a promoter/inhibitor, and an evolutionary mechanism of QPSO and improved QPSO is proposed. It is one kind of SNS-based MIEA, and simply named CQPSO-ETP. Evolutional symport/antiport rules and a promoter/inhibitor are introduced to this extended P system in order to adjust the executing sequence of evolution–communication rules. In CQPSO-ETP, two kinds of evolution rules with a promoter and inhibitor for objects based on different generating strategies for local attractor are described, one of the evolution rules with a promoter is based on the evolutionary mechanism of the standard QPSO technique. Another evolution rule with an inhibitor is based on the evolutionary mechanism of an improved QPSO technique using self-adaptive selection, cooperative evolution and a logistic chaotic mapping method. The communication for objects in different membranes or regions is achieved by the execution of communication rules with a promoter and inhibitor for objects. At last, the proposed CQPSO-ETP is evaluated on eight classic numerical benchmark functions and compared with PSO, QPSO, and two existing improved QPSO optimization approaches to verify the validity and performance. Furthermore, this extended P system is compared with three classic existing clustering techniques, and comparison experiments on eight tested images from image segmentation problems are performed to validate the clustering efficiency of this proposed CQPSO-ETP.
The rest of this paper is organized as follows: the basic framework of the tissue-like P system with evolutional symport/antiport rules and a promoter/inhibitor are described in
Section 2. More details about the evolutionary mechanism of QPSO and improved QPSO are given in
Section 3. The extended tissue-like P system based on the tissue-like P system with evolutional symport/antiport rules and a promoter/inhibitor, and QPSO and improved QPSO are proposed in
Section 4, and evolution and communication rules with a promoter/inhibitor for objects are described in more details in this section. Experimental results and analysis on eight classic numerical benchmark functions with four comparative approaches are reported in
Section 5.
Section 6 gives the experimental results and discussion which are made on eight tested images with three classic existing clustering approaches to evaluate the clustering efficiency of this proposed extended tissue-like P system.
Section 7 provides some conclusions and outlines future research directions.
4. The Proposed CQPSO-ETP
In this section, an extended tissue-like P system based on the evolution–communication mechanism of ETP and the evolution mechanism of QPSO and improved QPSO is proposed, and simply named CQPSO-ETP. The evolutionary mechanism for objects and communication mechanism for global objects are introduced in this extended P system. The rest of this section is organized in the following. Firstly, the general framework of this extended tissue-like P system is described, and the basic membrane structure is given in more details. Secondly, the evolution mechanism of QPSO and CQPSO and the evolution-communication mechanism of a tissue-like P system with evolutional symport/antiport rules and a promoter/inhibitor are introduced in the CQPSO-ETP to improve the performance of the extended P system. The computation of the proposed CQPSO-ETP is given in the following content. The complexity analysis of the CQPSO-ETP is described in the last parts.
4.1. The General Framework of CQPSO-ETP
The general framework of CQPSO-ETP is similar to the tissue-like P system with evolutional symport/antiport rules and a promoter/inhibitor. However, the rules for objects in the CQPSO-ETP are divided into two kinds of rules for objects, including the evolution rules for objects and communication rules for global objects. The membranes in the CQPSO-ETP system, are labeled from
to
, and simply denoted by
. Respectively, CQPSO-ETP is a tuple which can be formally described in the following,
where
- (1)
is a finite non-empty alphabet of objects;
- (2)
is the membrane structure of CQPSO-ETP consisting of membranes;
- (3)
are finite multisets of initial objects in the membranes, with , for ;
- (4)
represents finite sets of evolution rules in the CQPSO-ETP, . Where () represents a finite set of evolution rules for objects associated with membrane . Furthermore, the evolution rules of membrane are of the form: , for . When the evolution rule is applied, object is evolved to object with the promoter/inhibitor in the same membrane. In particular, is the promoter or inhibitor of the form and , where represents the promoter and represents the inhibitor.
- (5)
represents finite sets of communication rules in the CQPSO-ETP, . () represents a finite set of commutation rules for global objects associated with membrane to membrane . The form of the communication rules is also the same as the form of the evolutional symport/antiport rules, which can be described in the form: or , for , , where is the promoter or inhibitor of the form and , represents the promoter, and represents the inhibitor; note that .
- (6)
is the output membrane in the CQPSO-ETP, where
or
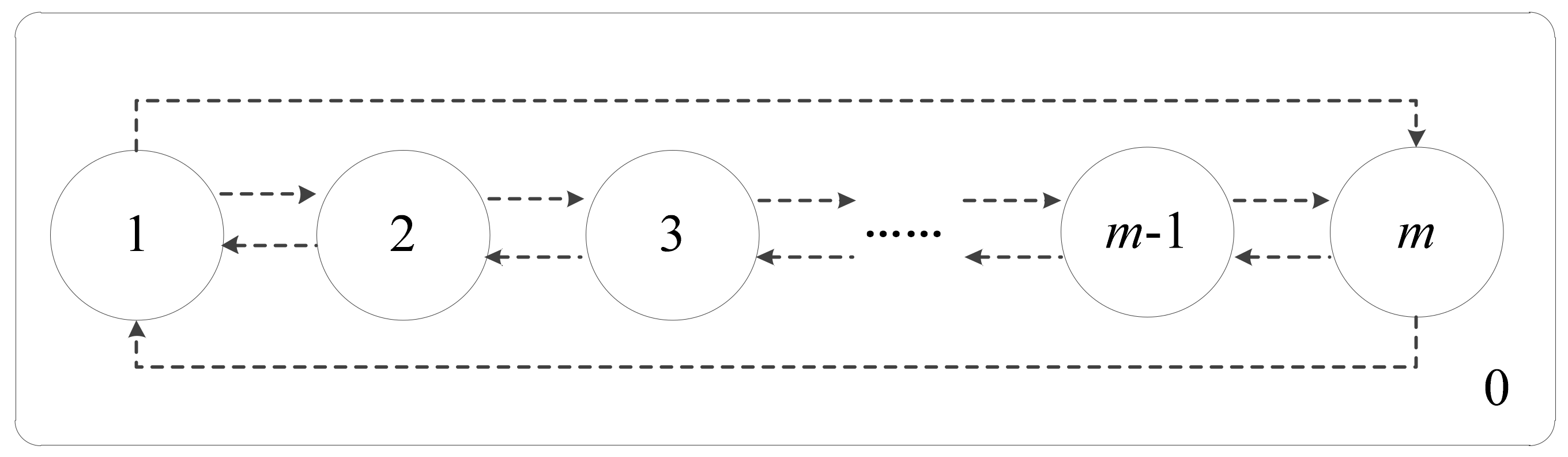
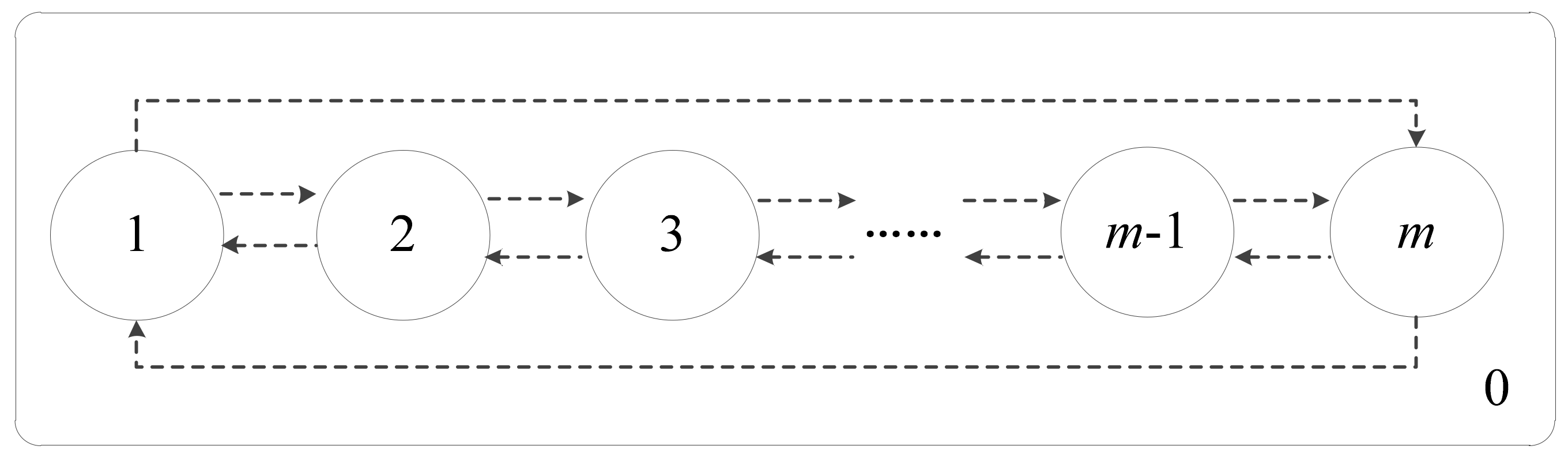
. Once the computation is completed, the computational results or objects in the output membrane will be transported to the environment. The membrane structure of CQPSO-ETP is especially graphically depicted in
Figure 2.
In
Figure 2, the CQPSO-ETP contains
membranes, which are simply denoted by
.
(
) is called the elementary membrane and does not contain any other membrane, and
represents the environment that is around the elementary membrane. All membranes work in a parallel way, and each membrane can be viewed as a parallel computing unit. The elementary membrane only communicated with its neighboring membranes, which are the adjacent membranes of the current membrane in the graphical structure of
Figure 2. This communication relationship is defined by the dotted lines, the exchange and sharing of information in different membranes is connected with the direction of the arrow in
Figure 2. All initial objects of the system are contained in elementary membranes from
to
in the CQPSO-ETP. Therefore, in the proposed CQPSO-ETP,
(
) represents the input membrane
, consisting of all initial objects for the system, and
represents the output region
, which is used to store computational results or objects of the system for each iteration.
4.2. Evolution Rules
There are two kinds of evolution rules based on different updating mechanisms of QPSO and improved QPSO for objects in the proposed CQPSO-ETP, including the evolution rules with the promoter and the evolution rules with the inhibitor . The evolution rules with the promoter/inhibitor are normally executed only on the elementary membrane in order to generate the position of objects.
4.2.1. The Evolution Rules with Promoter
In this work, the mechanism of QPSO is adopted to generate the position of the local attractor and objects in the elementary membrane
(
). The evolution rules with promoter
are of the form:
. At iteration
, the position
of the
-th object
(
) in the elementary membrane
is determined by (12) in the following,
where
is the position of object
in
at iteration
, for
.
is the total number of objects in
.
is iteration counter.
is a random parameter of
at iteration
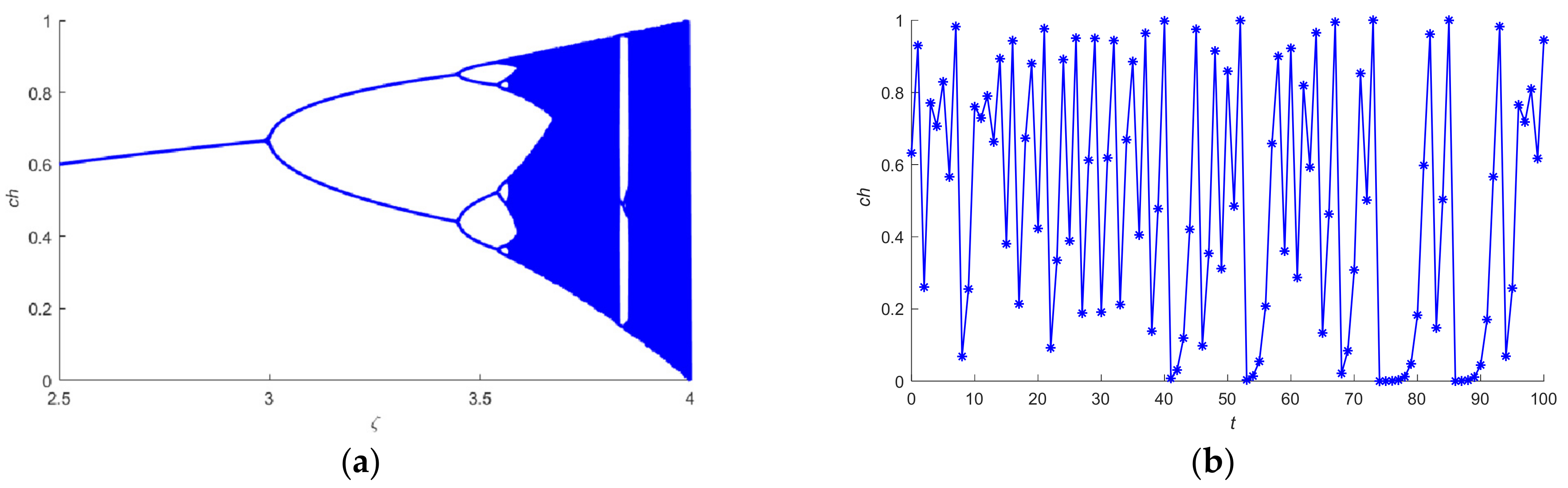
, which is based on the logistic chaotic mapping method according to Equation (11).
is the contraction–expansion coefficient of
at iteration
according to Equation (2). The values of contraction–expansion coefficient are dynamically updated according to a linear decreasing method for each iteration, and are given by (13) in the following,
where
is the maximum number of iterations. In Equation (12),
is the mean position of the local best for all objects in
at iteration
according to Equation (3), and it is given by (14) in the following,
where
(
) is the position of the local best in
at iteration
.
is the local attractor of
in
at iteration
, which is determined by (15) in the following,
where
is regulation parameter at iteration
, and its values are dynamically updated according to Equation (5).
is the position of the global best for all particles in the population of
at iteration
.
4.2.2. The Evolution Rules with Inhibitor
The mechanism of CQPSO is adopted to generate the position of the local attractor and objects in the elementary membrane
(
). The evolution rules with inhibitor
are of the form:
. Updating the position for objects in the evolution rules with the inhibitor is determined by Equation (12). A perturbation vector is introduced to the updating method of the local attractor for the objects, which is different from the updating method based on the QPSO technique in the evolution rules with a promoter through Equation (15). The local attractor
of object
in the elementary membrane
at iteration
is given by (16) in the following,
where
is the position of the local best for object
in
at iteration
; note that
.
is a random object in
which is randomly selected from the population of
.
is a perturbation vector, which is determined by (17) according to Equation (9) in the following,
where
is the adjustment parameter of the local attractor
,
.
is a uniform random number of
, which is distributed on the interval from 0 to 1.
is the position of the local best for object
in
at iteration
; note that
, and
is a random object which is randomly selected from the population of the elementary membrane
.
4.3. Communication Rules
In the proposed CQPSO-ETP, the evolutional antiport rules, as we mentioned above, are introduced to the communication rules to improve the convergence speed and accuracy. The exchange and sharing of information for different membranes or regions is achieved by the execution of the evolutional antiport rules for the objects. Two kinds of communication rules with a promoter/inhibitor are adopted to the CQPSO-ETP, including the communication rules with promoter and the communication rules with inhibitor .
4.3.1. The Communication Rules with Promoter
In the CQPSO-ETP, the execution of communication rules with promoter depends on the relationship between the elementary membrane and its adjacent membrane ; note that , which are of the form: , for or . It only can be executed on a moment if there is an elementary membrane in a configuration which contains a multiset of promoter objects, . When the communication rule with a promoter is applied, the position of the global best for all objects in the adjacent membrane of is evolved to the position of the local best for a random object, , which is randomly selected from the population of , and is send to at iteration . At the same time, the position of the global best for all the objects and promoter object in the elementary membrane are consumed during this evolutional antiport process. Especially, if , or , or else if , or .
4.3.2. The Communication Rules with Inhibitor
The execution of communication rules with inhibitor depends on the relationship between the elementary membrane and its adjacent membrane ; note that , which are of the form: , for or . It only can be executed on a moment if there is an elementary membrane in a configuration which contains a multiset of inhibitor objects . When the communication rule with an inhibitor is applied, the position of the global best for all objects in the elementary membrane is evolved to the position of the local best for a random object, , which is randomly selected from the population of , and is sent to the adjacent membrane at iteration . At the same time, the position of the global best for all objects in the adjacent membrane and inhibitor object in the elementary membrane are consumed during this evolutional antiport process. Especially, if , or , or else if , or .
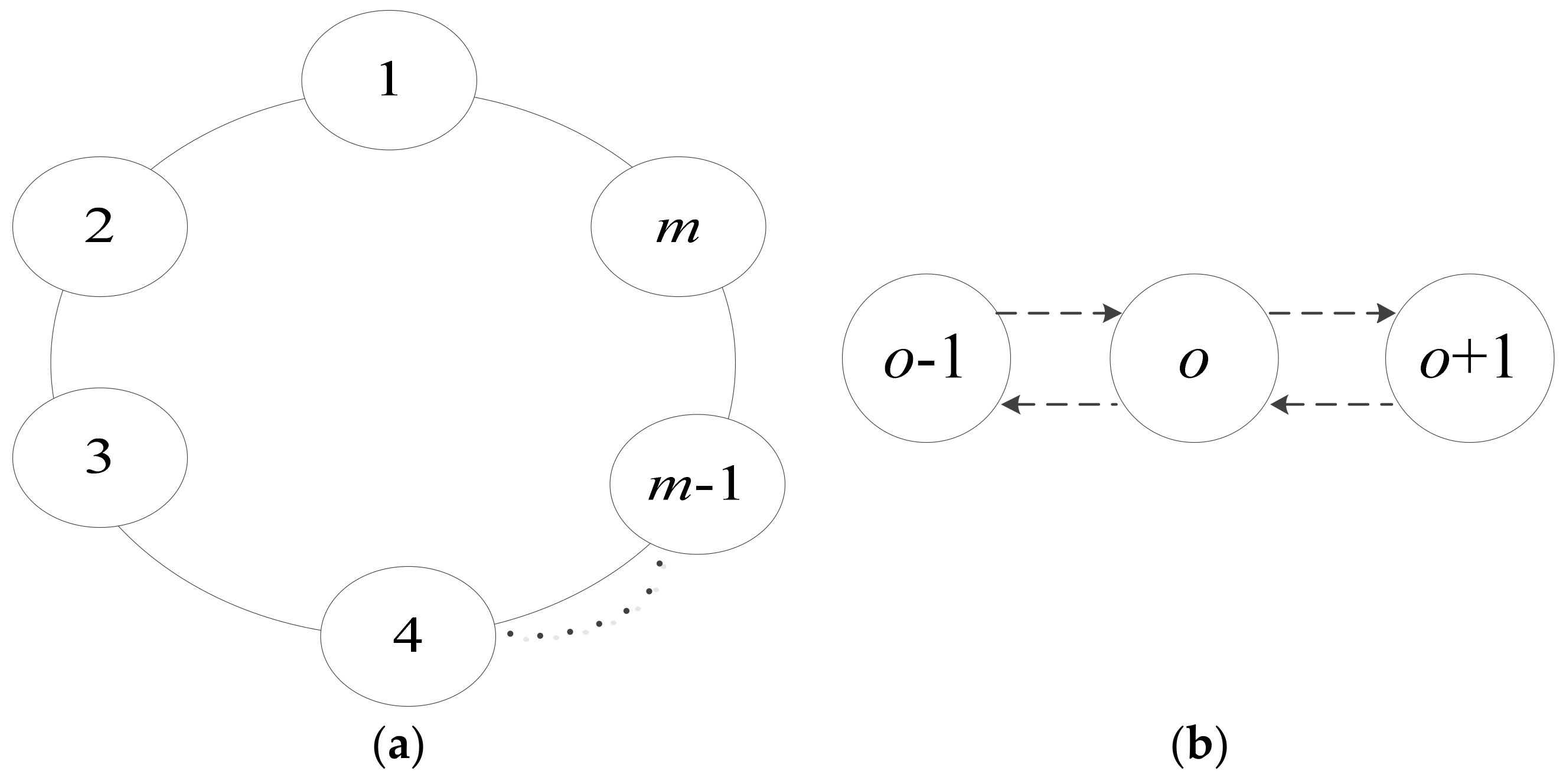
The commutation relationship is established from the execution of communication rules with a promoter/inhibitor between the elementary membrane and its adjacent membrane, which is depicted by a loop topology structure in mathematics, as shown in
Figure 3a. Besides this, the communication relationship based on the communication rules with a promoter/inhibitor also constructed the neighborhood structure of the elementary membranes. The exchange and sharing of information only performed on the elementary membrane and its neighboring membranes through communication rules with a promoter/inhibitor, as shown in
Figure 3b.
Especially, the communication rules of the elementary membrane () and environment are also based on the evolutional symport rules for objects, which are of the form: . When the communication rule is applied, the position of the global best for all objects in the elementary membrane is evolved to the position of the local best for the object , and is sent to the environment at iteration . Then the position of the global best which is selected from the position of the local best for all transformation objects is always stored in the environment as the global best of the system at iteration .
4.4. Compuatation of CQPSO-ETP
- (1)
Initialization
Step1: Parameter initialization
In the proposed CQPSO-ETP, the elementary membrane () contains all initial objects, and the number of objects in each membrane is the same, which is denoted by as we mentioned above. The total number of objects in this system is denoted by , where ;
Step2: Position initialization
The position of the objects is randomly initialized in the search space. The membrane structure of this extended P system is shown in
Figure 2. In general, the optimization problem is considered as the minimum optimization problem;
Step3: Update the position of the local best and global best
Update the position of the local best and global best for all objects in each elementary membrane. Note that the promoter and inhibitor will not simultaneously exist in the same elementary membrane. The evolution and communication rules with the promoter/inhibitor are only executed on a configuration at a moment when the promoter/inhibitor objects have appeared in the elementary membrane.
- (2)
Evolution rules for objects
The promoter and inhibitor in the evolution rules are described as some restricted conditions of the system, including the comparison condition and stagnation condition. The comparison condition is adopted to compare the fitness values of the global best in the elementary membrane and its neighboring membranes, and the stagnation condition is adopted to evaluate whether the position of the global best for all objects in the elementary membrane cannot be further improved for iterations.
Step1: if , for , the evolution rules with promoter for the objects are adopted to update the position of the local best and global best in each elementary membrane, according to Equations (12)–(15); if , for , the evolution rules with inhibitor for the objects are adopted to update the position of the local best and global best in each elementary membrane, according to Equations (12)–(14), (16) and (17);
Step2: Update the position of the local best and global best in each elementary membrane.
- (3)
Communication rules for objects
The promoter and inhibitor in the communication rules are considered as some comparison conditions. The comparison condition is introduced to compare the fitness values of the global best for all objects in the elementary membrane and its neighboring membranes.
Step1: if , for , the evolutional antiport rules with promoter are adopted to transport the position of the global best in the neighboring membrane to the elementary membrane; if , for , the evolutional antiport rules with inhibitor are adopted to transport the position of the global best in the elementary membrane to its neighboring membranes;
Step2: The position of the global best for object in each elementary membrane is sent to the environment through the execution of communication rules, , and is evolved to the position of the local best for object in the environment. Then the environment stored the best position among these transformation local bests as the global best in the system, and it also represented the computational results of the system for each iteration.
- (4)
Halting and output
The evolution and communication rules for objects in the proposed CQPSO-ETP will be implemented repeatedly with an iterative form until the termination criterion is satisfied. The termination criterion of CQPSO-ETP is stopped when the maximum number of iterations is reached. When the system halts, the position of the last global best, which is stored in the environment, is regarded as the final computational results for the system. Algorithm 1 depicted the main pseudo code of the computation for proposed CQPSO-ETP.
| Algorithms1 CQPSO-ETP: The pseudo code of computation. |
| Input: ,; |
| (1) Initialization |
| Update the local and global best; |
| to |
| for to , |
| = object(i). Local best. Position; |
| = object(i). Local best. Fitness; |
| if |
| = object(i). Global best. Position; |
| = object(i). Global best. Fitness; |
| end if |
| end |
| end |
| for to |
| (2) Evolution rules for objects |
| for to |
| for to |
| Step 1: if , |
| ; |
| ; |
| else , |
| ; |
| ; |
| end |
| Step2: update the local and global best; |
| end |
| end |
| (3) Communication rules for the objects |
| for to |
| Step 1: if , |
| ,; |
| else , |
| , |
| end |
| Step 2: ; |
| Update the global best in the system |
| end |
| end |
| (4) Halting and output |
| if |
| =Best Position; |
| =Best Fitness; |
| end |
| Output: Best Position, Best Fitness. |
4.5. Complexity Analysis
The complexity of the proposed CQPSO-ETP is analyzed in this subsection. As defined earlier, is the total number of objects in the system. () is the initial number of objects in the elementary membrane , where . is the number of the elementary membranes in the system. is the maximum number of iterations. is the dimension of the search space. The computation of the proposed CQPSO-ETP consists of three steps. In the first step of initialization, the computational time of fitness function for the initial objects in each elementary membrane is . Due to the parallel working manner, the complexity of the initialization for the system is . In the second step, the computational time for the execution of the evolution rules with a promoter for the objects is the same as the computational time for the execution of the evolution rules with an inhibitor for the objects. Moreover, the computing time needed by executing one evolution rule with a promoter or inhibitor for an object is . Hence, the total computing time needed by executing one evolution rule with a promoter/inhibitor for the objects in each elementary membrane is . Therefore, the complexity of the evolution process for all objects in the system is . In the third step, the communication time for the execution of the communication rules with a promoter is same as the communication time for the execution of the communication rules with an inhibitor, which contains the exchange time for objects between the membrane and its neighboring membranes. The communication time needed by executing one communication rule is 2. Therefore, the complexity of the communication process for all elementary membranes in the system is . As a result, the computational time at each iteration for the system is . Thus, the total computational time needed for the computing process of CQPSO-ETP is , and the complexity of the system is . As the number of objects in each elementary membrane is the same and will not change in the computing process, which is denoted by , then . The complexity of the proposed CQPSO-ETP is also described as , which can be simplified to .
7. Conclusions
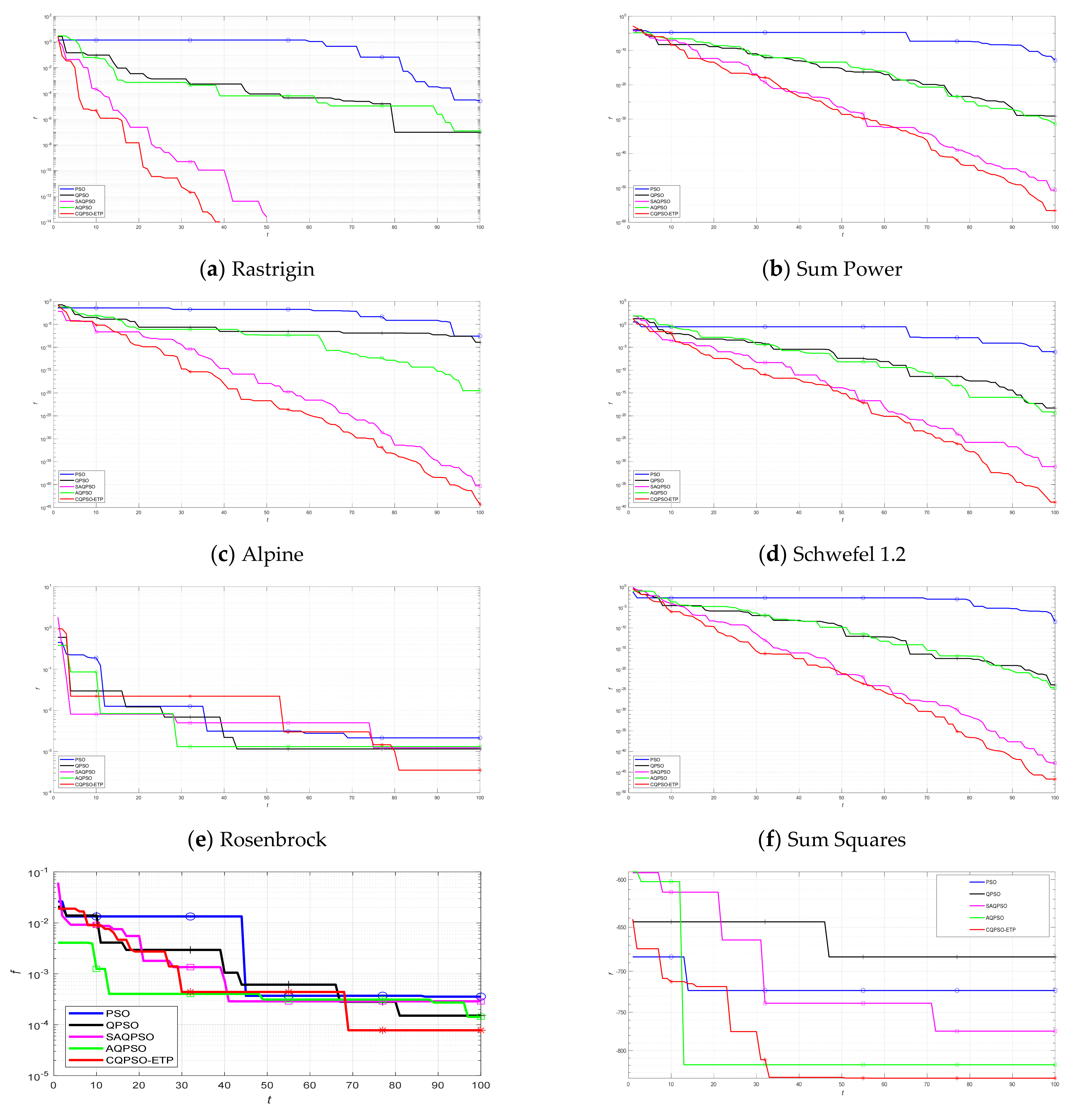
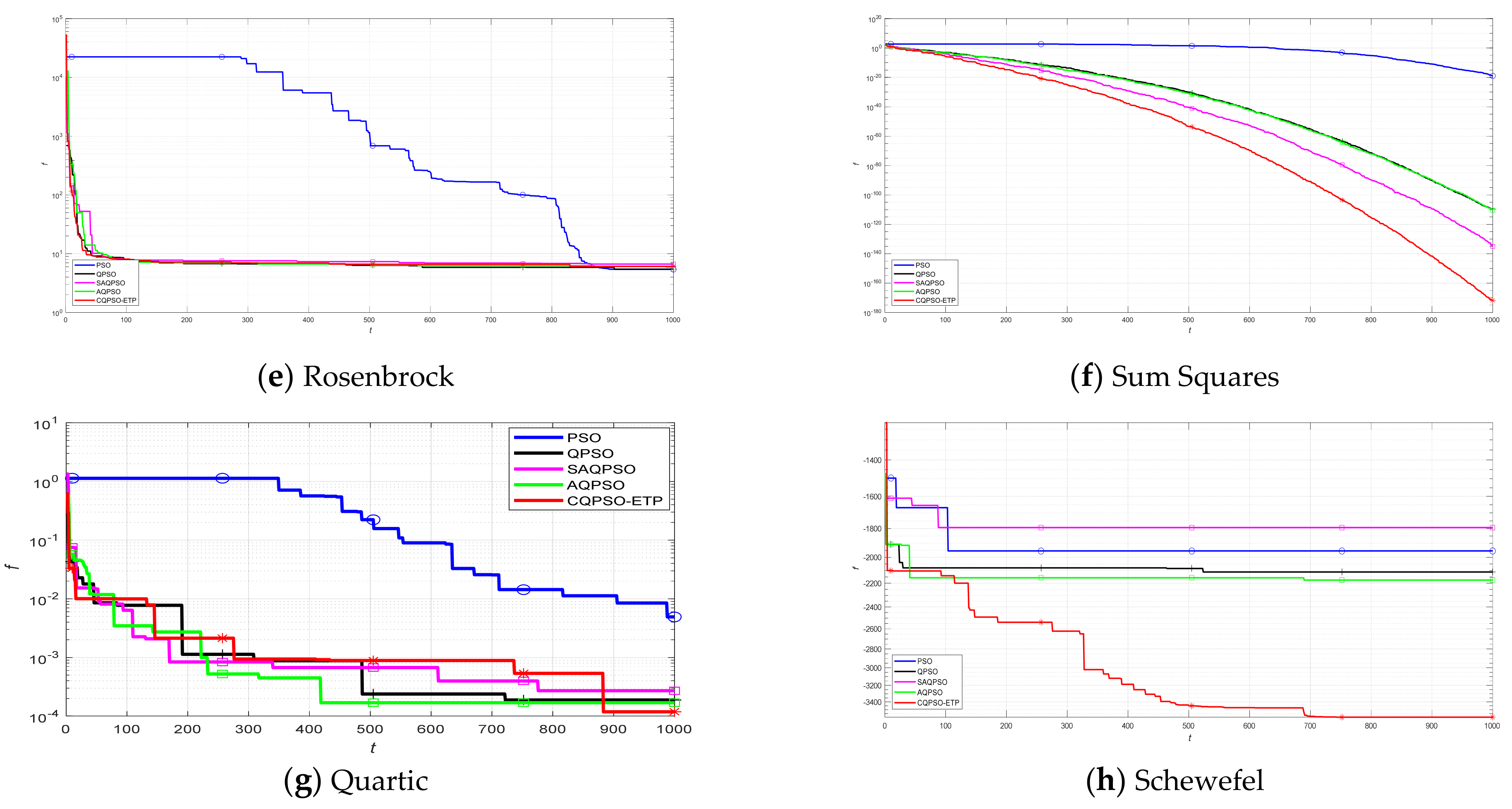
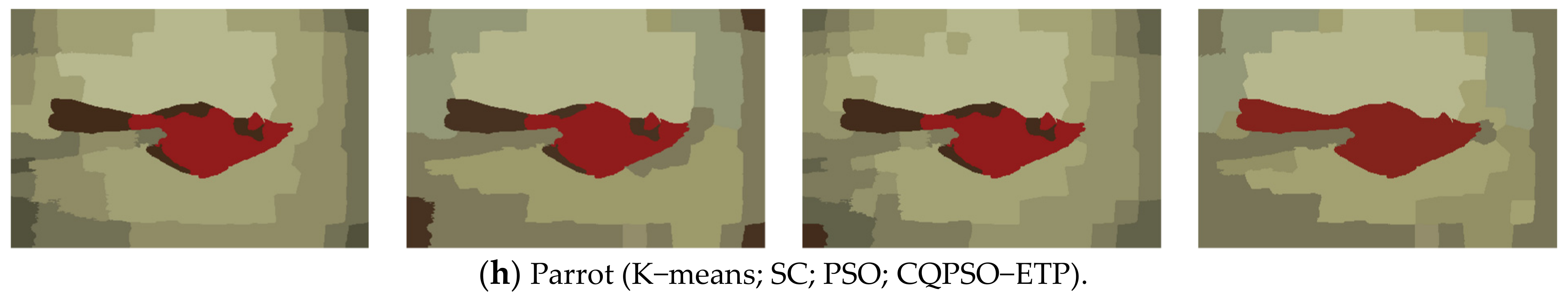
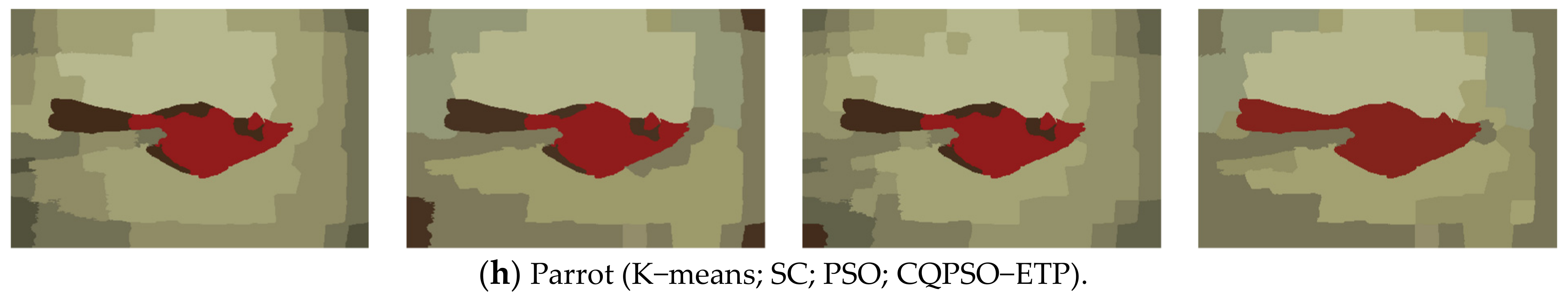
An extended tissue−like P system combining the evolutionary mechanism of QPSO and improved QPSO and the evolution–communication mechanism of tissue−like P systems is proposed, called the CQPSO−ETP, to solve optimization problems and image segmentation problems. This extended tissue−like P system under the framework of membrane computing using the tissue−like P system with evolutional symport/antiport rules and a promoter/inhibitor for objects, and the distributed parallel computing model of tissue−like P systems is adopted to enhance the global search ability of the CQPSO−ETP. Different from the existing SNS−based MIEAs, the CQPSO−ETP has two kinds of evolution rules with a promoter and inhibitor for objects. The evolution rules with a promoter are based on the basic position updating strategy for particles in a standard QPSO. Other evolution rules with an inhibitor are based on the position updating strategy for particles in an improved QPSO using self−adaptive selection, and cooperative evolutionary and logistic chaotic mapping methods, to accomplish the evolution of the objects in the system. The evolution mechanism for objects based on the QPSO and improved QPSO is used to improve the optimization performance of the CQPSO−ETP. The communication rules with promoter and inhibitor for objects are adopted to transfer the local and global best positions of objects to achieve the exchange and sharing of information between different membranes. The communication mechanism for objects based on communication rules in tissue−like P systems is introduced to improve the convergence speed and accuracy. The promoter and inhibitor are also introduced to control the exchanged direction of information between different membranes. The computational experiments, which are compared with PSO, QPSO and two improved QPSO approaches, are evaluated on eight classic numerical benchmark functions from previous studies and researches, and the results clearly exhibit the optimization effectiveness of this proposed extended tissue−like P system. Furthermore, the comparison experiments which are made on eight tested images from the image segmentation datasets are conducted to verify the clustering performance of the CQPSO−ETP as compared with three classic clustering techniques, including K−means, SC, and PSO, and the computational results show the validity of this extended tissue−like P system.
The computing model of tissue−like P systems is the parallel computing model, which is highly effective and more efficient in solving optimization problems with linear or polynomial complexity. However, the application of tissue−like P systems has been limited by the incomplete fundamental operation and implementation difficulties. The evolution mechanism based on the evolution computing techniques provides a new way to achieve the evolutionary process for objects in P systems. Then, the computation of tissue−like P systems in biology, which contain the execution of the evolution and communication rules for objects, is converted to the update and exchange of potential solutions in mathematics. The proposed CQPSO−ETP takes the tissue−like P system with evolutional symport/antiport rules and a promoter/inhibitor as the basic computational structure, and the communication rules for objects will establish the communication relationship between different membranes and regions in P systems. These relation links between different membranes and regions are bidirectional, which are simple and easy to implement. However, these static relation links in future studies are going to be replaced by dynamic relation links in order to further accelerate convergence and improve the population diversity. Besides, a more complicated membrane computing structure in P systems may be introduced in future studies to improve the optimization performance of the SNS−based MIEAs. Furthermore, the computing experiments were only executed on a low dimensional search space for classic numerical benchmark functions, and the proposed CQPSO−ETP may have some limitations in a high dimensional space. Therefore, future studies may test the effectiveness of CQPSO−ETP using high dimensional benchmark functions, and may also focus on MIEAs based on the extended neural−like P systems and other bio−inspired computing models. More works are also needed to balance the local and global search abilities of this proposed extended P system.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}