

The Immunological Mechanisms Involved in the Pathophysiology of Allergic Proctocolitis †
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Study Enrolment
2.2. Analysis of Immune Subsets and Treg Cells
2.3. Statistical Analysis
3. Results
3.1. Clinical Characteristics of Enrolled Infants
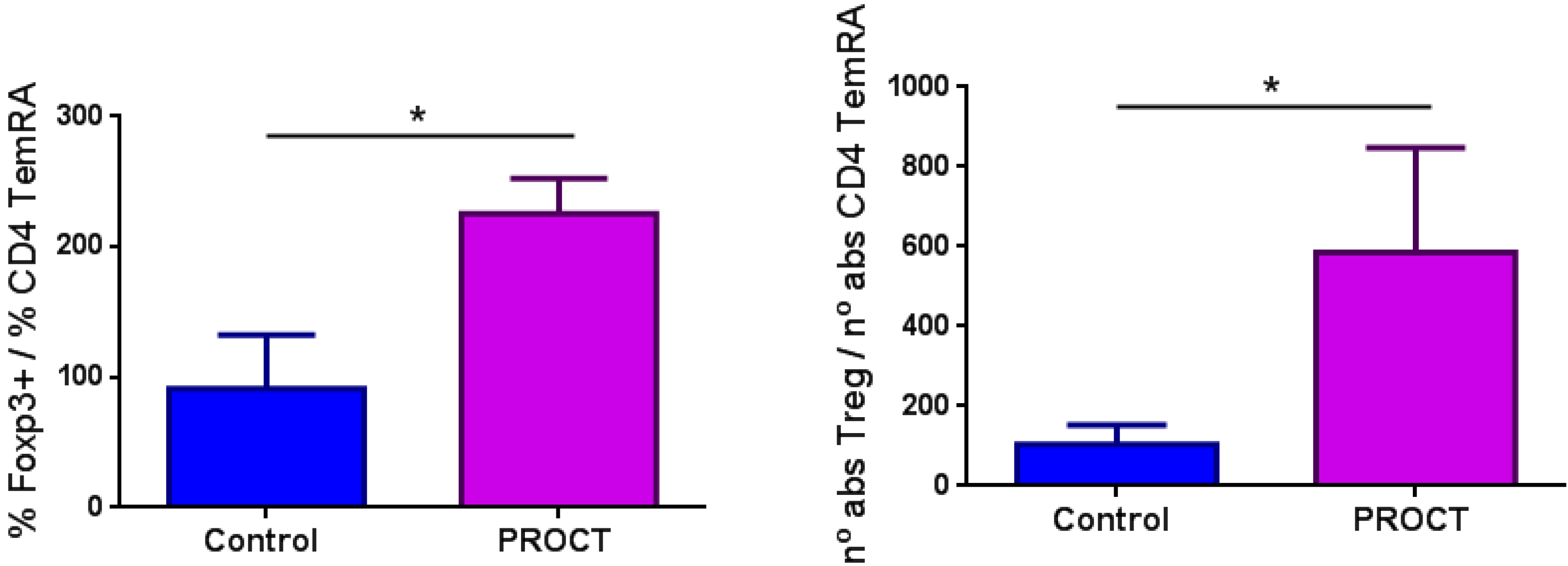
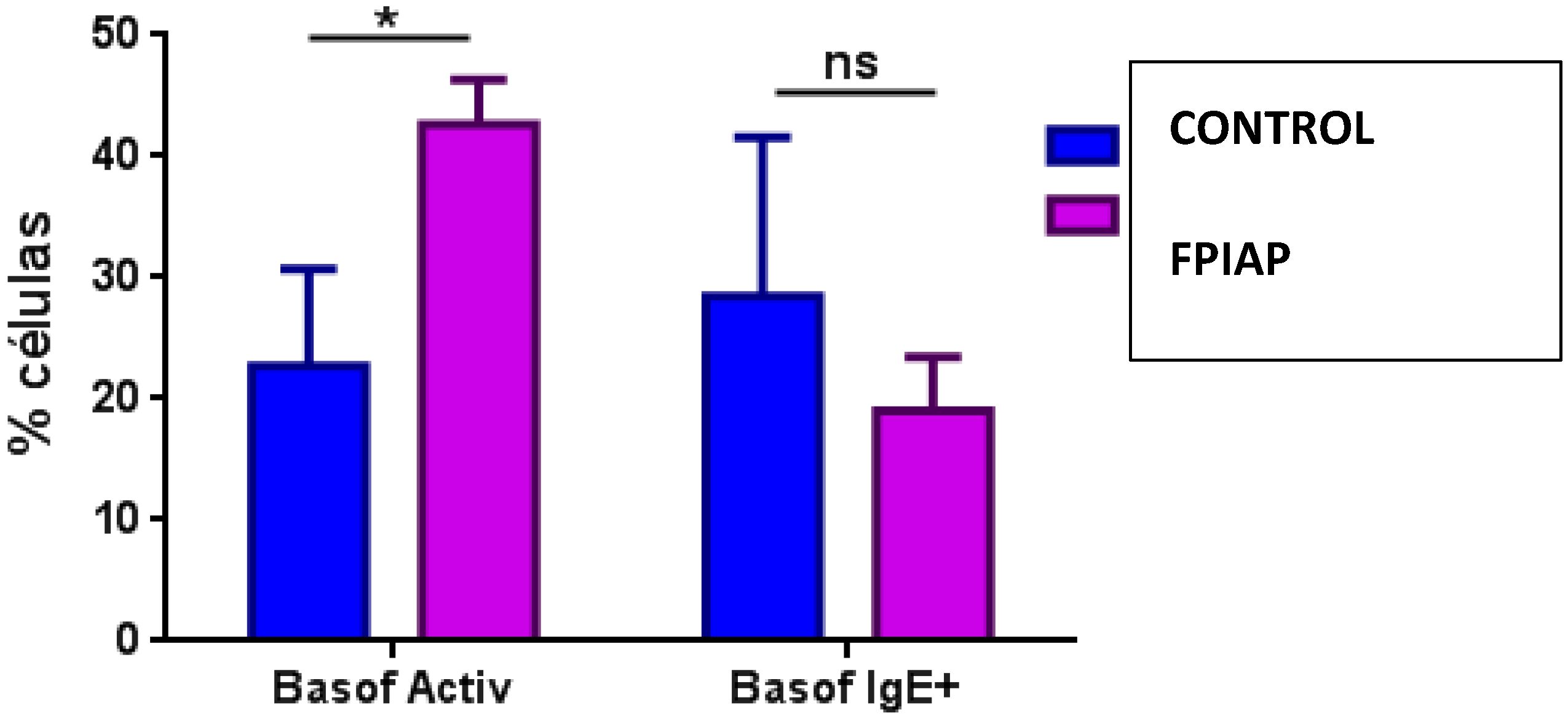
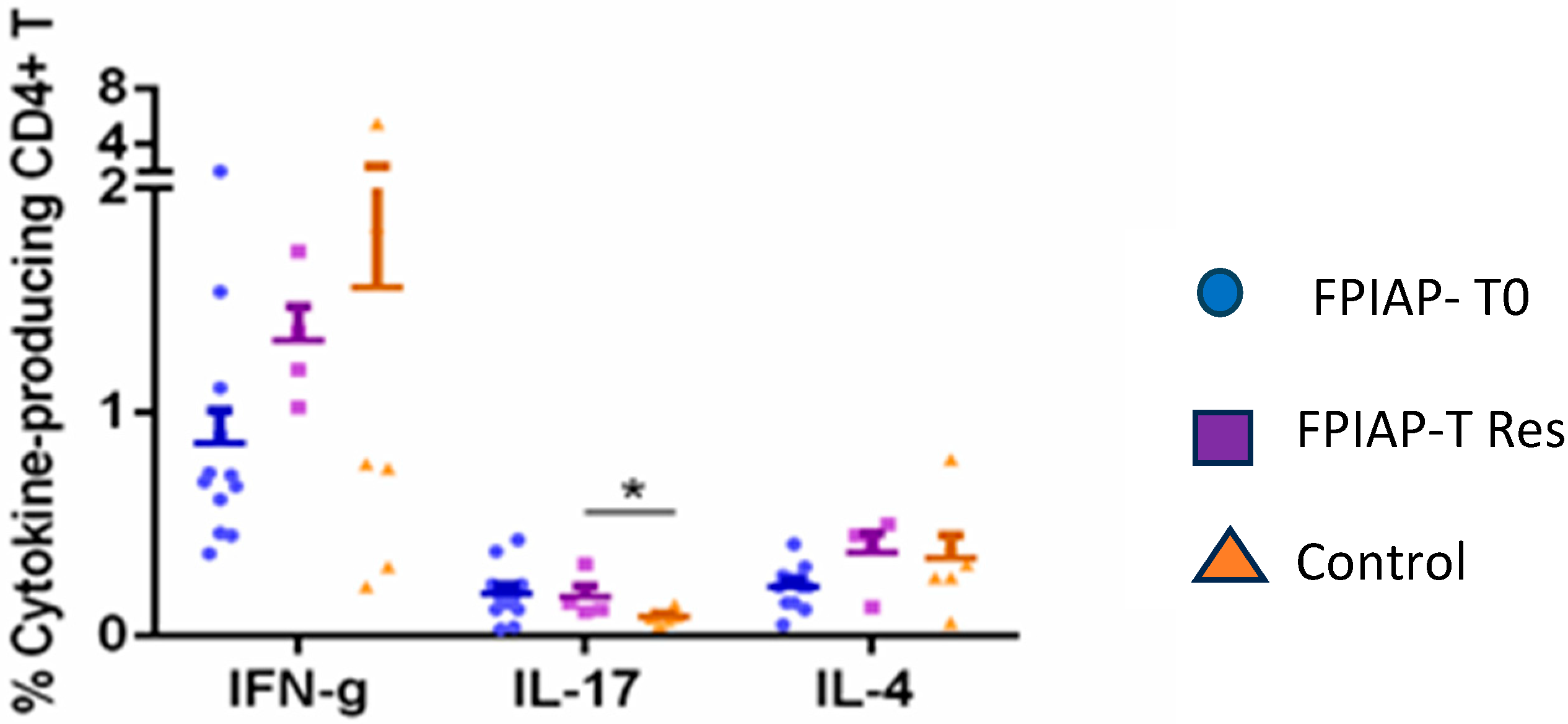
3.2. Results from the Analysis of Immune Subsets and Treg Cells
4. Discussion
5. Key Message
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schoemaker, A.A.; Sprikkelman, A.B.; Grimshaw, K.E.; Roberts, G.; Grabenhenrich, L.; Rosenfeld, L.; Siegert, S.; Dubakiene, R.; Rudzeviciene, O.; Reche, M.; et al. Incidence and natural history of challenge-proven cow’s milk allergy in European children—EuroPrevall birth cohort. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, S.; Heine, R.G. Non-IgE mediated cow’s milk allergy in EuroPrevall. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 1679–1680. [Google Scholar] [CrossRef] [PubMed]
- Martorell-Aragonés, A.; Echeverría-Zudaire, L.; Alonso-Lebrero, E.; Boné-Calvo, J.; Martín-Muñoz, M.F.; Nevot-Falcó, S.; Piquer-Gibert, M.; Valdesoiro-Navarrete, L.; Food Allergy Committee of SEICAP (Spanish Society of Pediatric Allergy, Asthma and Clinical Immunology). Position document: IgE-mediated cow’s milk allergy. Allergol. Immunopathol. 2015, 43, 507–526. [Google Scholar] [CrossRef]
- Nowak-Węgrzyn, A.; Katz, Y.; Mehr, S.S.; Koletzko, S. Non–IgE-mediated gastrointestinal food allergy. J. Allergy Clin. Immunol. 2015, 135, 1114–1124. [Google Scholar] [CrossRef]
- Connors, L.; O’Keefe, A.; Rosenfield, L.; Kim, H. Non-IgE-mediated food hypersensitivity. Allergy Asthma Clin. Immunol. 2018, 14, 56. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Węgrzyn, A.; Chehade, M.; Groetch, M.E.; Spergel, J.M.; Wood, R.A.; Allen, K.; Atkins, D.; Bahna, S.; Barad, A.V.; Berin, C.; et al. International consensus guidelines for the diagnosis and management of food protein–induced enterocolitis syndrome: Executive summary—Workgroup Report of the Adverse Reactions to Foods Committee, American Academy of Allergy, Asthma & Immunology. J. Allergy Clin. Immunol. 2017, 139, 1111–1126.e4. [Google Scholar]
- Espín Jaime, B.; Díaz Martín, J.J.; Blesa Baviera, L.C.; Claver Monzón, Á.; Hernández Hernández, A.; García Burriel, J.I.; Mérida, M.J.G.; Fernández, C.P.; Rodríguez, C.C.; Riechmann, E.R.; et al. Non-IgE-mediated cow’s milk allergy: Consensus document of the Spanish Society of Paediatric Gastroenterology, Hepatology, and Nutrition (SEGHNP), the Spanish Association of Paediatric Primary Care (AEPAP), the Spanish Society of Extra-hospital Paediatrics and Primary Health Care (SEPEAP), and the Spanish Society of Paediatric ClinicaL Immunology, Allergy, and Asthma (SEICAP). An. Pediatr. 2019, 90, 193.e1–193.e11. [Google Scholar]
- Nowak-Węgrzyn, A. Food protein-induced enterocolitis syndrome and allergic proctocolitis. Allergy Asthma Proc. 2015, 36, 172–184. [Google Scholar] [CrossRef]
- Salvatore, S.; Folegatti, A.; Ferrigno, C.; Pensabene, L.; Agosti, M.; D’Auria, E. To Diet or Not to Diet This Is the Question in Food-Protein-Induced Allergic Proctocolitis (FPIAP)-A Comprehensive Review of Current Recommendations. Nutrients 2024, 16, 589. [Google Scholar] [CrossRef]
- Meyer, R.; Venter, C.; Bognanni, A.; Szajewska, H.; Shamir, R.; Nowak-Wegrzyn, A.; Fiocchi, A.; Vandenplas, Y. World Allergy Organization (WAO) Diagnosis and Rationale for Action against Cow’s Milk Allergy (DRACMA) Guideline update—VII—Milk elimination and reintroduction in the diagnostic process of cow’s milk allergy. World Allergy Organ. J. 2023, 16, 100785. [Google Scholar] [CrossRef]
- Fox, A.; Brown, T.; Walsh, J.; Venter, C.; Meyer, R.; Nowak-Wegrzyn, A.; Levin, M.; Spawls, H.; Beatson, J.; Lovis, M.-T.; et al. An update to the Milk Allergy in Primary Care guideline. Clin. Transl. Allergy 2019, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Senocak, N.; Ertugrul, A.; Ozmen, S.; Bostanci, I. Clinical Features and Clinical Course of Food Protein—Induced Allergic Proctocolitis: 10-Year Experience of a Tertiary Medical Center. J. Allergy Clin. Immunol. Pract. 2022, 10, 1608–1613. [Google Scholar] [CrossRef] [PubMed]
- Mennini, M.; Fiocchi, A.G.; Cafarotti, A.; Montesano, M.; Mauro, A.; Villa, M.P.; Di Nardo, G. Food protein-induced allergic proctocolitis in infants: Literature review and proposal of a management protocol. World Allergy Organ. J. 2020, 13, 100471. [Google Scholar] [CrossRef]
- Barni, S.; Mori, F.; Giovannini, M.; Liotti, L.; Mastrorilli, C.; Pecoraro, L.; Saretta, F.; Castagnoli, R.; Arasi, S.; Caminiti, L.; et al. Allergic Proctocolitis: Literature Review and Proposal of a Diagnostic–Therapeutic Algorithm. Life 2023, 13, 1824. [Google Scholar] [CrossRef]
- Boné, J.; Claver, Á.; Guallar, I.; Plaza, A.M. Allergic proctocolitis, food-induced enterocolitis: Immune mechanisms, diagnosis and treatment. Allergol. Immunopathol. 2009, 37, 36–42. [Google Scholar] [CrossRef]
- Noval Rivas, M.C.T. Regulatory T cells in Allergic Diseases Magali. J. Allergy Clin. Immunol. 2016, 138, 639–652. [Google Scholar] [CrossRef]
- Perezabad, L.; López-Abente, J.; Alonso-Lebrero, E.; Seoane, E.; Pion, M.C.-R.R. The establishment of cow’s milk protein allergy in infants is related with a deficit of regulatory T cells (Treg) and vitamin D. Pediatr. Res. 2017, 81, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Aparicio, V.; Alonso-Lebrero, E.; Zapatero, L.; Infante, S.; Lorente, R.; Muñoz-Fernández, M.Á.; Correa-Rocha, R. Induction of Treg cells after oral immunotherapy in hen’s egg-allergic children. Pediatr. Allergy Immunol. 2014, 25, 103–106. [Google Scholar] [CrossRef]
- Carucci, L.; Nocerino, R.; Coppola, S.; Bedogni, G.; Capasso, P.; Giglio, V.; Berni Canani, R. Factors influencing the natural history of non-IgE-mediated gastrointestinal food allergies in paediatric age: A prospective multicentre cohort study. BMJ Paediatr. Open 2025, 9, e003203. [Google Scholar] [CrossRef] [PubMed]
- Barni, S.; Pessina, B.; Fioretti, L.; Scarallo, L.; Di Siena, A.; Bramuzzo, M.; Liccioli, G.; Sarti, L.; Tomei, L.; Giovannini, M.; et al. Food Protein-Induced Allergic Proctocolitis: Real-World Experience from an Italian Cohort. Nutrients 2024, 17, 98. [Google Scholar] [CrossRef]
- Vallianatou, G.N.; Douladiris, N.; Mageiros, L.; Manousakis, E.; Zisaki, V.; Galani, M.; Xepapadaki, P.; Taka, S.; Papadopoulos, N.G. Duration of food protein-induced allergic proctocolitis (FPIAP) and the role of intestinal microbiota. Pediatr. Allergy Immunol. 2024, 35, e70008. [Google Scholar] [CrossRef] [PubMed]
- Caubet, J.C.; Nowak-Wȩgrzyn, A. Current understanding of the immune mechanisms of food protein-induced enterocolitis syndrome. Expert Rev. Clin. Immunol. 2011, 7, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Nocerino, R.; Terrin, G.; Frediani, T.; Lucarelli, S.; Cosenza, L.; Passariello, A.; Leone, L.; Granata, V.; Di Costanzo, M.; et al. Formula selection for management of children with cow’s milk allergy influences the rate of acquisition of tolerance: A prospective multicenter study. J. Pediatr. 2013, 163, 771–777.e1. [Google Scholar] [CrossRef]
- Hernández-Martín, A.; Kennedy-Batalla, R.; Cañedo, E.; Bernaldo-de-Quirós, E.; Carazo-Gallego, B.; Vera, A.; Torrelo, A.; Noguera-Morel, L.; González-Sarmiento, R.; Bolling, M.; et al. Imbalance in T-Helper 17 Cells and Targeted Therapy in an Infant with SAM-like Syndrome. N. Engl. J. Med. 2019, 381, 2176–2178. [Google Scholar] [CrossRef] [PubMed]
- López-Abente, J.; Bernaldo-de-Quirós, E.; Camino, M.; Gil, N.; Panadero, E.; Campos-Domínguez, M.; Seoane-Reula, E.; Gil-Jaurena, J.M.; Pion, M.; Correa-Rocha, R. Immune dysregulation and Th2 polarization are associated with atopic dermatitis in heart-transplant children: A delicate balance between risk of rejection or atopic symptoms. Am. J. Transplant. 2019, 19, 1536–1544. [Google Scholar] [CrossRef]
- Caubet, J.C.; Ford, L.S.; Sickles, L.; Järvinen, K.M.; Sicherer, S.H.; Sampson, H.A.; Nowak-Węgrzyn, A. Clinical features and resolution of food protein-induced enterocolitis syndrome: 10-year experience. J. Allergy Clin. Immunol. 2014, 134, 382–389. [Google Scholar] [CrossRef]
- Goswami, R.; Blazquez, A.B.; Kosoy, R.; Berin, M.C.; Sciences, G.; Core, I.M. Systemic innate immune activation in food protein–induced enterocolitis syndrome. J. Allergy Clin. Immunol. 2017, 139, 1885–1896. [Google Scholar] [CrossRef]
- Martin, V.M.; Virkud, Y.V.; Dahan, E.; Seay, H.L.; Itzkovits, D.; Vlamakis, H.; Xavier, R.; Shreffler, W.G.; Yuan, Q.; Yassour, M. Longitudinal disease-associated gut microbiome differences in infants with food protein-induced allergic proctocolitis. Microbiome 2022, 10, 154. [Google Scholar] [CrossRef]
- Akdis, M. Immune Responses in Healthy and Allergic Individuals Are Characterized by a Fine Balance between Allergen-specific T Regulatory 1 and T Helper 2 Cells. J. Exp. Med. 2004, 199, 1567–1575. [Google Scholar] [CrossRef]
- Dang, T.D.; Allen, K.J.; Martino, D.J.; Koplin, J.J.; Licciardi, P.V.; Tang, M.L.K. Food-allergic infants have impaired regulatory T-cell responses following in vivo allergen exposure. Pediatr. Allergy Immunol. 2016, 27, 35–43. [Google Scholar] [CrossRef]
- Wawrzyniak, M.; Mahony, L.O.; Akdis, M. Role of Regulatory Cells in Oral Tolerance. Allergy Asthma Immunol. Res. 2017, 9, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Palomares, O.; Martín-Fontecha, M.; Lauener, R.; Traidl-Hoffmann, C.; Cavkaytar, O.; Akdis, M.; Akdis, C.A. Regulatory T cells and immune regulation of allergic diseases: Roles of IL-10 and TGF-β. Genes Immun. 2014, 15, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.R.; Rugtveit, J.; Brandtzaeg, P. Allergen-responsive CD4+CD25+ regulatory T cells in children who have outgrown cow’s milk allergy. J. Exp. Med. 2004, 199, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Cseh, A.; Molnár, K.; Pintér, P.; Szalay, B.; Szebeni, B.; Treszl, A.; Arató, A.; Vásárhelyi, B.; Veres, G. Regulatory T cells and T helper subsets in breast-fed infants with hematochezia caused by allergic colitis. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 675–677. [Google Scholar] [CrossRef]
- Kosnai, I.; Kuitunen, P.; Savilahti, E.S.P. Mast cells and eosinophils in the jejunal mucosa of patients with intestinal cow’s milk allergy and celiac disease of childhood. J. Pediatr. Gastroenterol. Nutr. 1984, 3, 368–372. [Google Scholar] [CrossRef]
- Wada, T.; Toma, T.; Muraoka, M.; Matsuda, Y.; Yachie, A. Elevation of fecal eosinophil-derived neurotoxin in infants with food protein-induced enterocolitis syndrome. Pediatr. Allergy Immunol. 2014, 25, 617–619. [Google Scholar] [CrossRef]
- Belizón, C.T.; Páez, E.O.; Claros, A.F.M.; Sánchez, I.R.; González, A.R.; Medialdea, R.V.; Salguero, J.M.R. Calprotectina fecal como apoyo al diagnóstico en la alergia a las proteínas de leche de vaca no IgE mediada. An. Pediatr. 2016, 84, 318–323. [Google Scholar] [CrossRef]
- Khan, S. Testing for fecal calprotectin in food protein–induced enterocolitis syndrome. J. Investig. Allergol. Clin. Immunol. 2018, 28, 287–288. [Google Scholar] [CrossRef]
- Koksal, B.T.; Barıs, Z.; Sencelikel, T.; Ozcay, F.; Ozbek, O.Y. Food protein-induced allergic proctocolitis in infants is associated with low serum levels of macrophage inflammatory protein-3a. J. Pediatr. Gastroenterol. Nutr. 2024, 78, 211–216. [Google Scholar] [CrossRef]
- Sekerkova, A.; Fuchs, M.; Cecrdlova, E.; Svachova, V.; Kralova Lesna, I.; Striz, I.; Tlaskalova-Hogenova, H. High Prevalence of Neutrophil Cytoplasmic Autoantibodies in Infants with Food Protein-Induced Proctitis/Proctocolitis: Autoimmunity Involvement? J. Immunol. Res. 2015, 2015, 902863. [Google Scholar] [CrossRef]
- Schmidt-Weber, C.B.; Akdis, M.; Akdis, C.A. TH17 cells in the big picture of immunology. J. Allergy Clin. Immunol. 2007, 120, 247–254. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune Parameter (%) | FPIAP | FPIAP | CONTROL | p |
|---|---|---|---|---|
| T0 (n = 22) | Tres (n = 10) | (n = 10) | ||
| Total lymphocyte count (CD3+) | 61.27 ± 8.4 | 61.7 ± 8.39 | 62.7 ± 7.44 | 0.85 |
| T-DP CD4+,CD8+ | 0.14 ± 0.19 | 0.07 ± 0.09 | 0.05 ± 0.07 | 0.014 * |
| TCD4+ cells | 46.22 ± 7.33 | 45.33 ± 8.06 | 47.72 ± 6.94 | 0.889 |
| CD4_38DR | 0.72 ± 0.35 | 0.72 ± 0.26 | 0.59 ± 0.25 | 0.388 |
| CD4_38 | 82.33 ± 3.6 | 80.7 ± 5.67 | 82.66 ± 4.0 | 0.646 |
| -naïve | 85.89 ± 3.07 | 86.20 ± 4.90 | 87.19 ± 3.19 | 0.305 |
| -central memory (CM) | 10.68 ± 2.50 | 10.01 ± 4.35 | 9.08 ± 2.19 | 0.204 |
| -effector (TemRA) | 0.07 ± 0.06 | 0.08 ± 0.08 | 0.18 ± 0.22 | 0.345 |
| -activated (Act) | 0.54 ± 0.43 | 0.56 ± 0.36 | 0.51 ± 0.38 | 0.952 |
| -RTE | 72.23 ± 4.98 | 72.76 ± 8.35 | 79.4 ± 5.58 | 0.007 * |
| -effector memory (EM) | 0.36 ± 0.22 | 0.43 ± 0.22 | 0.40 ± 0.47 | 0.251 |
| Treg cells | 7.6 ± 1.52 | 7.00 ± 1.52 | 7.49 ± 1.33 | 0.67 |
| Treg_38DR | 4.89 ± 2.9 | 4.80 ± 2.88 | 1.6 ± 1.03 | 0.003 * |
| Treg_38 | 55.53 ± 6.5 | 51.38 ± 5.33 | 56.86 ± 5.6 | 0.366 |
| -naïve | 69.75 ± 5.38 | 72.28 ± 8.56 | 73.83 ± 6.46 | 0.058 |
| -central memory (CM) | 25.35 ± 4.99 | 22.47 ± 7.83 | 20.81 ± 5.35 | 0.031 * |
| -effector (TemRA) | 0.09 ± 0.06 | 0.07 ± 0.04 | 0.33 ± 0.58 | 0.345 |
| -activated (Act) | 2.96 ± 2.39 | 2.58 ± 1.18 | 2.76 ± 1.67 | 0.984 |
| -RTE | 53.79 ± 4.08 | 55.87 ± 8.84 | 63.18 ± 8.04 | 0.007 * |
| -effector memory (EM) | 0.75 ± 0.36 | 0.94 ± 0.44 | 1.09 ± 1.19 | 0.857 |
| Treg Foxp3 | 7.03 ± 1.39 | 5.89 ± 1.22 | 6.68 ± 1.43 | 0.588 |
| Foxp3 activated | 5.06 ± 3 | 4.7 ± 3 | 4.9 ± 3.3 | 0.887 |
| Foxp3_CD39 | 1.8 ± 1.37 | 3.6 ± 3.3 | 3.6 ± 6.7 | 0.661 |
| Foxp3_memory | 41.23 ± 19.2 | 37.86 ± 17.24 | 33.62 ± 8.4 | 0.288 |
| Foxp3_naïve | 58.2 ± 19 | 61.38 ± 17 | 65.57 ± 8.1 | 0.288 |
| Foxp3_RTE | 41.07 ± 14.2 | 44.94 ± 14 | 51.3 ± 7.6 | 0.043 * |
| TCD8+ cells | 13.88 ± 4.25 | 13.72 ± 4.53 | 12.8 ± 2.64 | 0.589 |
| -naïve | 88.74 ± 9.05 | 87.15 ± 10.82 | 87.16 ± 10.76 | 0.857 |
| -central memory (CM) | 7.78 ± 5.23 | 5.89 ± 3.10 | 7.68 ± 5.05 | 0.889 |
| -effector (TemRA) | 0.95 ± 2.41 | 3.78 ± 7.75 | 1.77 ± 2.75 | 0.388 |
| -activated (Act) | 1.07 ± 2.25 | 1.22 ± 2.31 | 1.38 ± 2.32 | 0.535 |
| -effector memory (EM) | 0.5 ± 1.6 | 0.7 ± 1.22 | 0.81 ± 1.59 | 0.562 |
| B CD19+ cells | 22.88 ± 7.21 | 21.04 ± 8.29 | 22.62 ± 9.17 | 0.826 |
| Naïve | 93.47 ± 2.06 | 92.34 ± 3.63 | 93.57 ± 1.53 | 0.92 |
| Memory switch | 0.87 ± 0.56 | 10.7 ± 0.63 | 0.75 ± 0.52 | 0.482 |
| Memory non-switch | 2.68 ± 1.41 | 2.13 ± 0.68 | 2.84 ± 1.34 | 0.704 |
| B regulatory (BR1) | 13.08 ± 6.46 | 8.83 ± 2.51 | 7.19 ± 3.71 | 0.010 * |
| BR1 Real | 0.09 ± 0.037 | 0.08 ± 0.04 | 0.04 ± 0.01 | 0.002 * |
| Breg I | 0.65 ± 0.25 | 0.83 ± 0.33 | 0.63 ± 0.20 | 0.974 |
| Breg II | 31.76 ± 7.24 | 27.23 ± 10 | 29.32 ± 8.9 | 0.671 |
| Plasmablasts | 0.71 ± 0.44 | 0.96 ± 0.94 | 0.68 ± 0.71 | 0.341 |
| MYELOID CELL POPULATION (%) | FPIAP | FPIAP | CONTROLS | p |
|---|---|---|---|---|
| T0 (n = 16) | Tres (n = 9) | (n = 7) | ||
| Granulocytes | 26.92 ± 10.67 | 25.57 ± 8.84 | 17.8 ± 4.6 | 0.03 * |
| Basophils | 0.80 ± 0.41 | 0.87 ± 0.33 | 0.85 ± 0.33 | 0.646 |
| Activated basophils | 36.13 ± 16.73 | 38.67 ± 21.14 | 30.43 ± 12.37 | 0.562 |
| IgE basophils | 13.37 ± 16.5 | 9.59 ± 8.9 | 13.3 ± 20 | 0.743 |
| Neutrophils | 66.70 ± 19.44 | 71.42 ± 28.82 | 74.71 ± 7.51 | 0.013 * |
| Eosinophils | 30.22 ± 18.55 | 26.6 ± 28.87 | 21.94 ± 6.41 | 0.009 * |
| Dendritic cells | 2.03 ± 0.78 | 1.65 ± 0.47 | 2.03 ± 0.71 | 0.891 |
| mDC2 | 6.28 ± 8.2 | 4.7 ± 6.66 | 0.7 ± 0.91 | 0.011 * |
| mDC1 | 16.90 ± 7.99 | 18.8 ± 5.44 | 18.9 ± 6.11 | 0.237 |
| -myeloid cells | 75.67 ± 11 | 73.1 ± 6.4 | 75.38 ± 8.68 | 0.680 |
| -plasma cells | 45.89 ± 19.7 | 42.85 ± 26.09 | 48.15 ± 19 | 0.731 |
| All NK cells | 21.38 ± 11.26 | 21.69 ± 13.42 | 18.73 ± 5.1 | 0.914 |
| NK16+56++ | 12.56 ± 7.86 | 9.94 ± 4.2 | 13.03 ± 5.07 | 0.494 |
| NK16+56+ | 58.68 ± 12.1 | 67.7 ± 19.2 | 77.25 ± 6.1 | 0.001 * |
| NK16+56- | 24.8 ± 8.3 | 18.68 ± 12.4 | 8.67 ± 6.7 | 0.001 * |
| Monocytes | 7.4 ± 2.4 | 6.83 ± 2.68 | 6.05 ± 1.16 | 0.261 |
| Classical monocytes | 87.26 ± 5.53 | 86.77 ± 8.4 | 90.68 ± 2.47 | 0.203 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Moreno, J.; Bernaldo-de-Quirós, E.; Tolín Hernani, M.; Álvarez-Calatayud, G.; Perezábad, L.; Sánchez Sánchez, C.; Correa-Rocha, R. The Immunological Mechanisms Involved in the Pathophysiology of Allergic Proctocolitis. Children 2025, 12, 688. https://doi.org/10.3390/children12060688
Pérez-Moreno J, Bernaldo-de-Quirós E, Tolín Hernani M, Álvarez-Calatayud G, Perezábad L, Sánchez Sánchez C, Correa-Rocha R. The Immunological Mechanisms Involved in the Pathophysiology of Allergic Proctocolitis. Children. 2025; 12(6):688. https://doi.org/10.3390/children12060688
Chicago/Turabian StylePérez-Moreno, Jimena, Esther Bernaldo-de-Quirós, Mar Tolín Hernani, Guillermo Álvarez-Calatayud, Laura Perezábad, César Sánchez Sánchez, and Rafael Correa-Rocha. 2025. "The Immunological Mechanisms Involved in the Pathophysiology of Allergic Proctocolitis" Children 12, no. 6: 688. https://doi.org/10.3390/children12060688
APA StylePérez-Moreno, J., Bernaldo-de-Quirós, E., Tolín Hernani, M., Álvarez-Calatayud, G., Perezábad, L., Sánchez Sánchez, C., & Correa-Rocha, R. (2025). The Immunological Mechanisms Involved in the Pathophysiology of Allergic Proctocolitis. Children, 12(6), 688. https://doi.org/10.3390/children12060688