
The Interplay Between Body Weight and the Onset of Puberty
Abstract
1. Introduction
2. The Timing of Puberty
2.1. Genetic Factors
2.2. Adrenarche
3. Body Weight and Puberty
3.1. Childhood Obesity
3.2. EDCs and Puberty
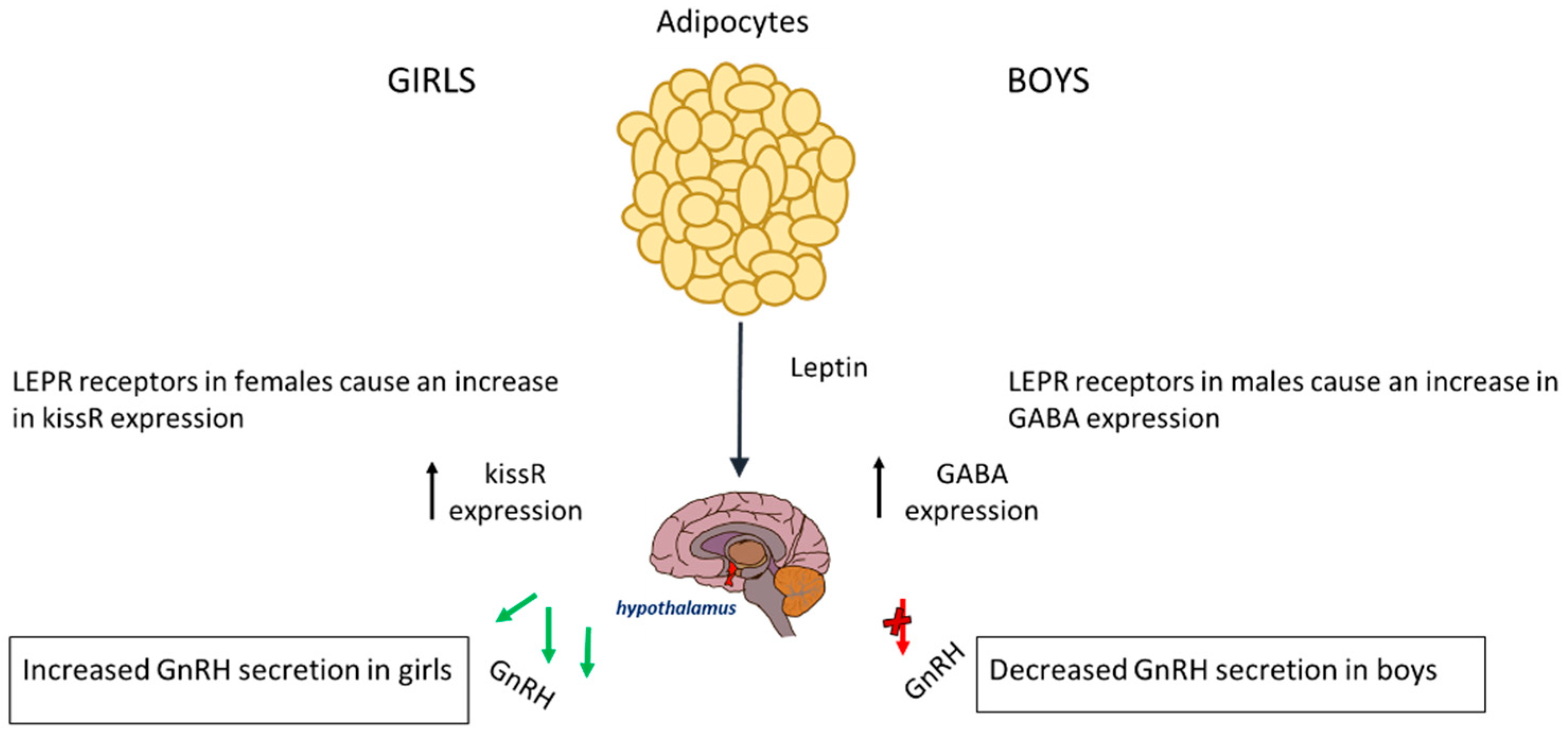
4. Adipokines (Leptin and Adiponectin)
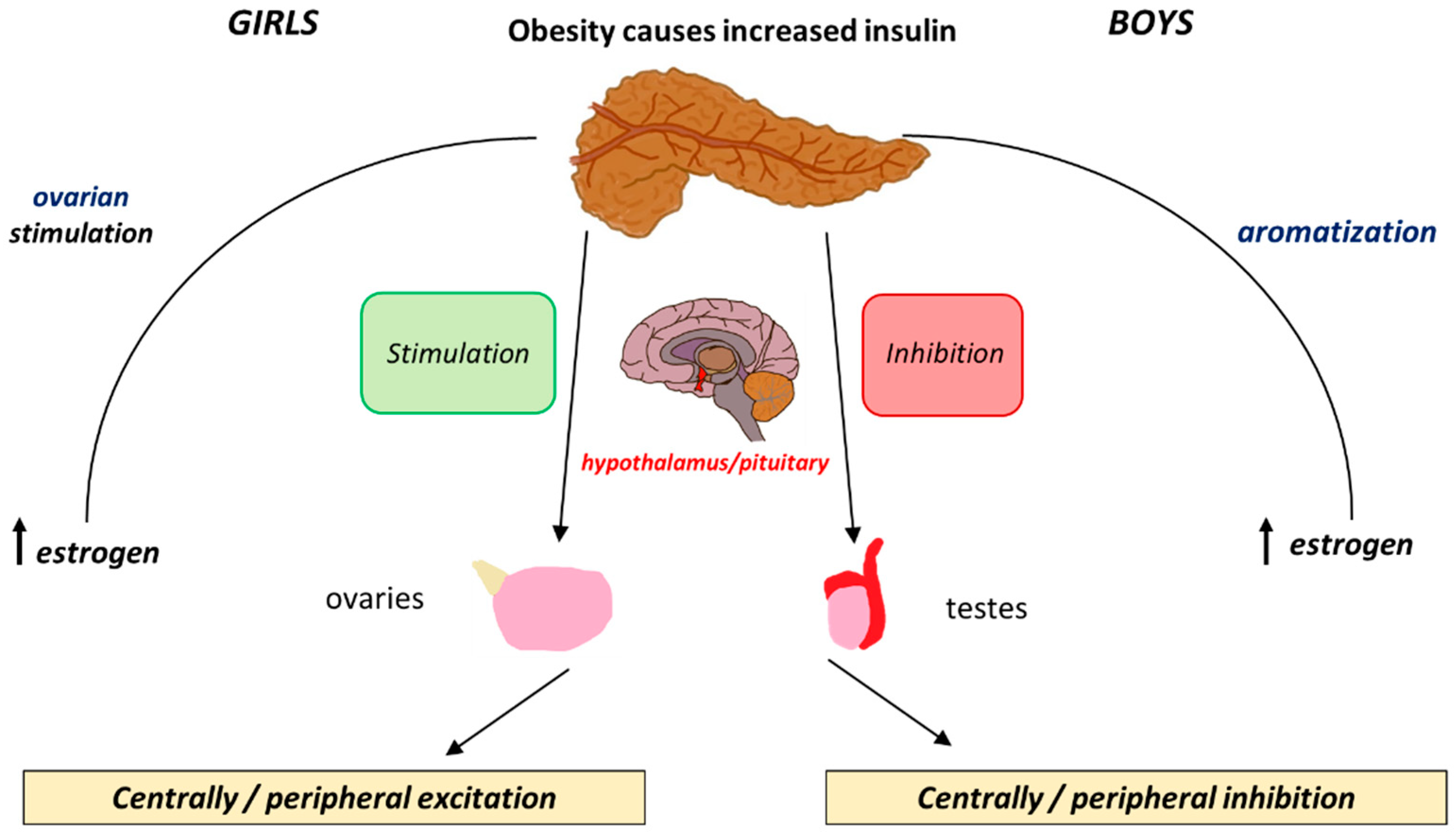
5. Insulin
6. SGA, Obesity, and Premature Gonadarche
7. Conclusions
8. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| HPG | Hypothalamic–Pituitary–Gonadal |
| KISS1 | Kisspeptin 1 |
| MKRN3 | Makorin Ring Finger Protein 3 |
| EDCs | Endocrine-Disrupting Chemicals |
| GnRH | Gonadotropin-Releasing Hormone |
| BMI | Body Mass Index |
| FSH | Follicle Stimulating Hormone |
| LH | Luteinising Hormone |
| PA | Puberty-Activating |
| PI | Puberty-Inhibiting |
| NPY | Neuropeptide Y |
| LEP | Leptin |
| GABA | Gamma-Aminobutyric Acid |
| DLK1 | Delta-Like Non-Canonical Notch Ligand 1 |
| KISS1R | Kisspeptin Receptor |
| GPR54 | G Protein-Coupled Receptor 54 |
| DHEA | Dehydroepiandrosterone |
| DHEAS | Dehydroepiandrosterone Sulphate |
| HPA | Hypothalamic–Pituitary–Adrenal |
| WHO | World Health Organization |
| UNICEF | United Nations Children’s Fund |
| NCD-RisC | NCD Risk Factor Collaboration |
| DEHP | Di(2-ethylhexyl) phthalate |
| BBB | Blood–Brain Barrier |
| LEPR | Leptin Receptor |
| GH | Growth Hormone |
| CYP19A1 | Cytochrome P450 19A1 |
| AdipoR1 | Adiponectin Receptor 1 |
| AdipoR2 | Adiponectin Receptor 2 |
| IL-6 | Interleukin-6 |
| TNFα | Tumor Necrosis Factor-Alpha |
| SHBG | Sex Hormone-Binding Globulin |
| SGA | Small for Gestational Age |
| BAI | Body Adiposity Index |
References
- Breehl, L.; Caban, O. Physiology, Puberty; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK534827 (accessed on 29 May 2024).
- Vijayakumar, N.; de Macks, Z.O.; Shirtcliff, E.A.; Pfeifer, J.H. Puberty and the Human Brain: Insights into Adolescent Development. Neurosci. Biobehav. Rev. 2018, 92, 417–436. [Google Scholar] [CrossRef]
- Manotas, M.C.; González, D.M.; Céspedes, C.; Forero, C.; Moreno, A.P.R. Genetic and Epigenetic Control of Puberty. Sex. Dev. 2021, 16, 1–10. [Google Scholar] [CrossRef]
- Joseph, D.N.; Whirledge, S. Stress and the HPA Axis: Balancing Homeostasis and Fertility. Int. J. Mol. Sci. 2017, 18, 2224. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.P.; Kaiser, U.B. Pubertal development and regulation. Lancet Diabetes Endocrinol. 2016, 4, 254–264. [Google Scholar] [CrossRef]
- Cunningham, S.A.; Hardy, S.T.; Jones, R.; Ng, C.; Kramer, M.R.; Narayan, K.V. Changes in the Incidence of Childhood Obesity. Pediatrics 2022, 150, e2021053708. [Google Scholar] [CrossRef]
- Eckert-Lind, C.; Busch, A.S.; Petersen, J.H.; Biro, F.M.; Butler, G.; Bräuner, E.V.; Juul, A. Worldwide Secular Trends in Age at Pubertal Onset Assessed by Breast Development Among Girls. JAMA Pediatr. 2020, 174, e195881. [Google Scholar] [CrossRef]
- Wohlfahrt-Veje, C.; Tinggaard, J.; Juul, A.; Toppari, J.; Skakkebæk, N.E.; Main, K.M. Pubarche and Gonadarche Onset and Progression Are Differently Associated with Birth Weight and Infancy Growth Patterns. J. Endocr. Soc. 2021, 5, bvab108. [Google Scholar] [CrossRef] [PubMed]
- Beccuti, G.; Ghizzoni, L. Normal and Abnormal Puberty; Endotext. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279024/ (accessed on 1 June 2024).
- Yang, D.; Zhang, W.; Zhu, Y.; Liu, P.; Tao, B.; Fu, Y.; Chen, Y.; Zhou, L.; Liu, L.; Gao, X.; et al. Initiation of the Hypothalamic–Pituitary–Gonadal Axis in Young Girls Undergoing Central Precocious Puberty Exerts Remodeling Effects on the Prefrontal Cortex. Front. Psychiatry 2019, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Spaziani, M.; Tarantino, C.; Tahani, N.; Gianfrilli, D.; Sbardella, E.; Lenzi, A.; Radicioni, A.F. Hypothalamo-Pituitary Axis and Puberty. Mol. Cell. Endocrinol. 2021, 520, 111094. [Google Scholar] [CrossRef]
- Blair, J.A.; McGee, H.; Bhatta, S.; Palm, R.; Casadesus, G. Hypothalamic–Pituitary–Gonadal Axis Involvement in Learning and Memory and Alzheimer’s Disease: More than ‘Just’ Estrogen. Front. Endocrinol. 2015, 6, 45. [Google Scholar] [CrossRef]
- Guercio, G.; Saraco, N.; Costanzo, M.; Marino, R.; Ramirez, P.; Berensztein, E.; Rivarola, M.A.; Belgorosky, A. Estrogens in Human Male Gonadotropin Secretion and Testicular Physiology from Infancy to Late Puberty. Front. Endocrinol. 2020, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Shaw, N.D.; Histed, S.N.; Srouji, S.S.; Yang, J.; Lee, H.; Hall, J.E. Estrogen Negative Feedback on Gonadotropin Secretion: Evidence for a Direct Pituitary Effect in Women. J. Clin. Endocrinol. Metab. 2010, 95, 1955–1961. [Google Scholar] [CrossRef] [PubMed]
- Lodish, M.B.; Ramnitz, M.S. Racial Disparities in Pubertal Development. Semin. Reprod. Med. 2013, 31, 333–339. [Google Scholar] [CrossRef]
- Grgic, O.; Shevroja, E.; Dhamo, B.; Uitterlinden, A.G.; Wolvius, E.B.; Rivadeneira, F.; Medina-Gomez, C. Skeletal Maturation in Relation to Ethnic Background in Children of School Age: The Generation R Study. Bone 2020, 132, 115180. [Google Scholar] [CrossRef]
- Chan, Y.-M.; Lippincott, M.F.; Barroso, P.S.; Alleyn, C.; Brodsky, J.; Granados, H.; Roberts, S.A.; Sandler, C.; Srivatsa, A.; Seminara, S.B. Using Kisspeptin to Predict Pubertal Outcomes for Youth with Pubertal Delay. J. Clin. Endocrinol. Metab. 2020, 105, e2717–e2725. [Google Scholar] [CrossRef]
- Christoforidis, A.; Skordis, N.; Fanis, P.; Dimitriadou, M.; Sevastidou, M.; Phelan, M.M.; Neocleous, V.; Phylactou, L.A. A Novel MKRN3 Nonsense Mutation Causing Familial Central Precocious Puberty. Endocrine 2017, 56, 446–449. [Google Scholar] [CrossRef]
- Howard, S.R. The Genetic Basis of Delayed Puberty. Front. Endocrinol. 2019, 10, 423. [Google Scholar] [CrossRef] [PubMed]
- Faienza, M.F.; Urbano, F.; Moscogiuri, L.A.; Chiarito, M.; De Santis, S.; Giordano, P. Genetic, Epigenetic and Enviromental Influencing Factors on the Regulation of Precocious and Delayed Puberty. Front. Endocrinol. 2022, 13, 1019468. [Google Scholar] [CrossRef]
- Pagani, S.; Calcaterra, V.; Acquafredda, G.; Montalbano, C.; Bozzola, E.; Ferrara, P.; Gasparri, M.; Villani, A.; Bozzola, M. MKRN3 and KISS1R Mutations in Precocious and Early Puberty. Ital. J. Pediatr. 2020, 46, 39. [Google Scholar] [CrossRef]
- Neocleous, V.; Shammas, C.; Phelan, M.M.; Nicolaou, S.; Phylactou, L.A.; Skordis, N. In Silico Analysis of a Novel MKRN3 Missense Mutation in Familial Central Precocious Puberty. Clin. Endocrinol. 2015, 84, 80–84. [Google Scholar] [CrossRef]
- Fanis, P.; Skordis, N.; Toumba, M.; Papaioannou, N.; Makris, A.; Kyriakou, A.; Neocleous, V.; Phylactou, L.A. Central Precocious Puberty Caused by Novel Mutations in the Promoter and 5′-UTR Region of the Imprinted MKRN3 Gene. Front. Endocrinol. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Rosenfield, R.L. Normal and Premature Adrenarche. Endocr. Rev. 2021, 42, 783–814. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.A.; Bales, N.J.; Myers, S.A.; Bautista, A.I.; Roueinfar, M.; Hale, T.M.; Handa, R.J. The Hypothalamic-Pituitary-Adrenal Axis: Development, Programming Actions of Hormones, and Maternal-Fetal Interactions. Front. Behav. Neurosci. 2021, 14, 601939. [Google Scholar] [CrossRef] [PubMed]
- Heck, A.L.; Handa, R.J. Sex Differences in the Hypothalamic–Pituitary–Adrenal Axis’ Response to Stress: An Important Role for Gonadal Hormones. Neuropsychopharmacology 2018, 44, 45–58. [Google Scholar] [CrossRef]
- Stephens, M.A.C.; Mahon, P.B.; McCaul, M.E.; Wand, G.S. Hypothalamic–Pituitary–Adrenal Axis Response to Acute Psychosocial Stress: Effects of Biological Sex and Circulating Sex Hormones. Psychoneuroendocrinology 2016, 66, 47–55. [Google Scholar] [CrossRef]
- WHO. Obesity and Overweight. World Health Organization. 7 May 2025. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 7 June 2024).
- World Obesity Federation. Prevalence of Obesity. World Obesity Federation. 2024. Available online: https://www.worldobesity.org/about/about-obesity/prevalence-of-obesity (accessed on 7 June 2024).
- Daley, S.F.; Balasundaram, P. Obesity in Pediatric Patients; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK570626 (accessed on 7 June 2024).
- Nicolaou, M.; Toumba, M.; Kythreotis, A.; Daher, H.; Skordis, N. Obesogens in Adolescence: Challenging Aspects and Prevention Strategies. Children 2024, 11, 602. [Google Scholar] [CrossRef]
- Lee, J.E.; Jung, H.W.; Lee, Y.J.; Lee, Y.A. Early-Life Exposure to Endocrine-Disrupting Chemicals and Pubertal Development in Girls. Ann. Pediatr. Endocrinol. Metab. 2019, 24, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Uldbjerg, C.S.; Koch, T.; Lim, Y.-H.; Gregersen, L.S.; Olesen, C.S.; Andersson, A.-M.; Frederiksen, H.; Coull, B.A.; Hauser, R.; Juul, A.; et al. Prenatal and postnatal exposures to endocrine disrupting chemicals and timing of pubertal onset in girls and boys: A systematic review and meta-analysis. Hum. Reprod. Update 2022, 28, 687–716. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Nah, W.H.; Park, M.J.; Gye, M.C. Effects of Early Prepubertal Exposure to Bisphenol A on the Onset of Puberty, Ovarian Weights, and Estrous Cycle in Female Mice. Clin. Exp. Reprod. Med. 2011, 38, 75–81. [Google Scholar] [CrossRef]
- Tassinari, R.; Tait, S.; Busani, L.; Martinelli, A.; Narciso, L.; Valeri, M.; Gastaldelli, A.; Deodati, A.; La Rocca, C.; Maranghi, F.; et al. Metabolic, Reproductive and Thyroid Effects of Bis(2-Ethylhexyl) phthalate (DEHP) Orally Administered to Male and Female Juvenile Rats at Dose Levels Derived from Children Biomonitoring Study. Toxicology 2021, 449, 152653. [Google Scholar] [CrossRef] [PubMed]
- Shalitin, S.; Gat-Yablonski, G. Associations of Obesity with Linear Growth and Puberty. Horm. Res. Paediatr. 2021, 95, 120–136. [Google Scholar] [CrossRef] [PubMed]
- López-Rodríguez, D.; Aylwin, C.F.; Delli, V.; Sevrin, E.; Campanile, M.; Martin, M.; Franssen, D.; Gérard, A.; Blacher, S.; Tirelli, E.; et al. Multi- and Transgenerational Outcomes of an Exposure to a Mixture of Endocrine-Disrupting Chemicals (EDCs) on Puberty and Maternal Behavior in the Female Rat. Environ. Health Perspect. 2021, 129, 87003. [Google Scholar] [CrossRef]
- Lucaccioni, L.; Trevisani, V.; Marrozzini, L.; Bertoncelli, N.; Predieri, B.; Lugli, L.; Berardi, A.; Iughetti, L. Endocrine-Disrupting Chemicals and Their Effects during Female Puberty: A Review of Current Evidence. Int. J. Mol. Sci. 2020, 21, 2078. [Google Scholar] [CrossRef]
- Adamska-Patruno, E.; Ostrowska, L.; Goscik, J.; Pietraszewska, B.; Kretowski, A.; Gorska, M. The Relationship between the Leptin/Ghrelin Ratio and Meals with Various Macronutrient Contents in Men with Different Nutritional Status: A Randomized Crossover Study. Nutr. J. 2018, 17, 118. [Google Scholar] [CrossRef]
- Picó, C.; Palou, M.; Pomar, C.A.; Rodríguez, A.M.; Palou, A. Leptin as a Key Regulator of the Adipose Organ. Rev. Endocr. Metab. Disord. 2021, 23, 13–30. [Google Scholar] [CrossRef]
- Li, W.; Liu, Q.; Deng, X.; Chen, Y.; Liu, S.; Story, M. Association between Obesity and Puberty Timing: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2017, 14, 1266. [Google Scholar] [CrossRef] [PubMed]
- Dunger, D.B.; Ahmed, M.L.; Ong, K.K. Effects of Obesity on Growth and Puberty. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 375–390. [Google Scholar] [CrossRef]
- Myers, M.G., Jr.; Leibel, R.L.; Seeley, R.J.; Schwartz, M.W. Obesity and Leptin Resistance: Distinguishing Cause from Effect. Trends Endocrinol. Metab. 2010, 21, 643–651. [Google Scholar] [CrossRef]
- Cheng, H.L.; Sainsbury, A.; Garden, F.; Sritharan, M.; Paxton, K.; Luscombe, G.; Hawke, C.; Steinbeck, K. Ghrelin and Peptide YY Change during Puberty: Relationships with Adolescent Growth, Development, and Obesity. J. Clin. Endocrinol. Metab. 2018, 103, 2851–2860. [Google Scholar] [CrossRef]
- Huang, A.; Reinehr, T.; Roth, C.L. Connections between Obesity and Puberty. Curr. Opin. Endocr. Metab. Res. 2020, 14, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Jiang, Z.; Zhang, L. Childhood Obesity and Central Precocious Puberty. Front. Endocrinol. 2022, 13, 1056871. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 12, 585887. [Google Scholar] [CrossRef]
- Odle, A.K.; Akhter, N.; Syed, M.M.; Allensworth-James, M.L.; Beneš, H.; Castillo, A.I.M.; MacNicol, M.C.; MacNicol, A.M.; Childs, G.V. Leptin Regulation of Gonadotrope Gonadotropin-Releasing Hormone Receptors as a Metabolic Checkpoint and Gateway to Reproductive Competence. Front. Endocrinol. 2018, 8, 367. [Google Scholar] [CrossRef]
- Machinal-Quélin, F.; Dieudonné, M.-N.; Pecquery, R.; Leneveu, M.-C.; Giudicelli, Y. Direct In Vitro Effects of Androgens and Estrogens on Ob Gene Expression and Leptin Secretion in Human Adipose Tissue. Endocrine 2002, 18, 179–184. [Google Scholar] [CrossRef]
- Kaplowitz, P. Delayed Puberty in Obese Boys: Comparison with Constitutional Delayed Puberty and Response to Testosterone Therapy. J. Pediatr. 1998, 133, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Zuure, W.A.; Roberts, A.L.; Quennell, J.H.; Anderson, G.M. Leptin Signaling in GABA Neurons, but Not Glutamate Neurons, Is Required for Reproductive Function. J. Neurosci. 2013, 33, 17874–17883. [Google Scholar] [CrossRef]
- Ahima, R.S. No Kiss1ng by Leptin during Puberty? J. Clin. Investig. 2011, 121, 34–36. [Google Scholar] [CrossRef]
- Nieuwenhuis, D.; Pujol-Gualdo, N.; Arnoldussen, I.A.; Kiliaan, A.J. Adipokines: A Gear Shift in Puberty. Obes. Rev. 2020, 21, e13005. [Google Scholar] [CrossRef]
- Song, Y.; Kong, Y.; Xie, X.; Wang, Y.; Wang, N. Association between Precocious Puberty and Obesity Risk in Children: A Systematic Review and Meta-Analysis. Front. Pediatr. 2023, 11, 1226933. [Google Scholar] [CrossRef]
- Ahl, S.; Guenther, M.; Zhao, S.; James, R.; Marks, J.; Szabo, A.; Kidambi, S. Adiponectin Levels Differentiate Metabolically Healthy vs. Unhealthy among Obese and Nonobese White Individuals. J. Clin. Endocrinol. Metab. 2015, 100, 4172–4180. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. Adiponectin-Resistance in Obesity. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2017; pp. 415–441. [Google Scholar] [CrossRef]
- Wu, Y.; Zhong, L.; Li, G.; Han, L.; Fu, J.; Li, Y.; Li, L.; Zhang, Q.; Guo, Y.; Xiao, X.; et al. Puberty Status Modifies the Effects of Genetic Variants, Lifestyle Factors and Their Interactions on Adiponectin: The BCAMS Study. Front. Endocrinol. 2021, 12, 737459. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, M.; Yin, J.; Cheng, H.; Yu, M.; Zhao, X.; Xiao, X.; Mi, J. Change of Body Composition and Adipokines and Their Relationship with Insulin Resistance across Pubertal Development in Obese and Nonobese Chinese Children: The BCAMS Study. Int. J. Endocrinol. 2012, 2012, 389108. [Google Scholar] [CrossRef] [PubMed]
- Barbe, A.; Bongrani, A.; Mellouk, N.; Estienne, A.; Kurowska, P.; Grandhaye, J.; Elfassy, Y.; Levy, R.; Rak, A.; Froment, P.; et al. Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. Int. J. Mol. Sci. 2019, 20, 1526. [Google Scholar] [CrossRef]
- Ohman-Hanson, R.A.; Cree-Green, M.; Kelsey, M.M.; Bessesen, D.H.; Sharp, T.A.; Pyle, L.; Pereira, R.I.; Nadeau, K.J. Ethnic and Sex Differences in Adiponectin: From Childhood to Adulthood. J. Clin. Endocrinol. Metab. 2016, 101, 4808–4815. [Google Scholar] [CrossRef]
- Kelsey, M.M.; Zeitler, P.S. Insulin Resistance of Puberty. Curr. Diabetes Rep. 2016, 16, 64. [Google Scholar] [CrossRef]
- Gołacki, J.; Matuszek, M.; Matyjaszek-Matuszek, B. Link between Insulin Resistance and Obesity—From Diagnosis to Treatment. Diagnostics 2022, 12, 1681. [Google Scholar] [CrossRef]
- Saleh, F.L.; Joshi, A.A.; Tal, A.; Xu, P.; Hens, J.R.; Wong, S.L.; Flannery, C.A. Hyperinsulinemia Induces Early and Dyssynchronous Puberty in Lean Female Mice. J. Endocrinol. 2022, 254, 121–135. [Google Scholar] [CrossRef]
- Sliwowska, J.H.; Fergani, C.; Gawałek, M.; Skowronska, B.; Fichna, P.; Lehman, M.N. Insulin: Its Role in the Central Control of Reproduction. Physiol. Behav. 2014, 133, 197–206. [Google Scholar] [CrossRef]
- Solorzano, C.M.B.; McCartney, C.R. Obesity and the Pubertal Transition in Girls and Boys. Reproduction 2010, 140, 399–410. [Google Scholar] [CrossRef]
- Mauras, N.; Ross, J.; Mericq, V. Management of Growth Disorders in Puberty: GH, GnRHa, and Aromatase Inhibitors: A Clinical Review. Endocr. Rev. 2022, 44, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nassar, G.N.; Leslie, S.W. Physiology, Testosterone; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK526128/ (accessed on 4 July 2024).
- Osuchukwu, O.O.; Reed, D.J. Small for Gestational Age; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK563247/ (accessed on 20 July 2024).
- Hong, Y.H.; Chung, S. Small for Gestational Age and Obesity Related Comorbidities. Ann. Pediatr. Endocrinol. Metab. 2018, 23, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhu, J.; Wang, X.; Shi, H.; Huo, Y.; Liu, M.; Sun, F.; Lan, H.; Guo, C.; Liu, H.; et al. Rapid BMI Increases and Persistent Obesity in Small-for-Gestational-Age Infants. Front. Pediatr. 2021, 9, 625853. [Google Scholar] [CrossRef]
- Nam, H.-K.; Lee, K.-H. Small for Gestational Age and Obesity: Epidemiology and General Risks. Ann. Pediatr. Endocrinol. Metab. 2018, 23, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Verkauskiene, R.; Petraitiene, I.; Wikland, K.A. Puberty in Children Born Small for Gestational Age. Horm. Res. Paediatr. 2013, 80, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, T.; Roth, C.L. Is There a Causal Relationship between Obesity and Puberty? Lancet Child Adolesc. Health 2019, 3, 44–54. [Google Scholar] [CrossRef]
- Jebeile, H.; Kelly, A.S.; O’Malley, G.; Baur, L.A. Obesity in Children and Adolescents: Epidemiology, Causes, Assessment, and Management. Lancet Diabetes Endocrinol. 2022, 10, 351–365. [Google Scholar] [CrossRef]
- Li, W.; Liu, Q.; Deng, X.; Chen, Y.; Yang, B.; Huang, X.; Østbye, T. Association of Prepubertal Obesity with Pubertal Development in Chinese Girls and Boys: A Longitudinal Study. Am. J. Hum. Biol. 2018, 30, e23195. [Google Scholar] [CrossRef]
- Lian, Q.; Mao, Y.; Luo, S.; Zhang, S.; Tu, X.; Zuo, X.; Lou, C.; Zhou, W. Puberty Timing Associated with Obesity and Central Obesity in Chinese Han Girls. BMC Pediatr. 2019, 19, 1. [Google Scholar] [CrossRef]
- Lazzeri, G.; Tosti, C.; Pammolli, A.; Troiano, G.; Vieno, A.; Canale, N.; Dalmasso, P.; Lemma, P.; Borraccino, A.; Petraglia, F.; et al. Overweight and Lower Age at Menarche: Evidence from the Italian HBSC Cross-Sectional Survey. BMC Women’s Health 2018, 18, 168. [Google Scholar] [CrossRef]
- Biro, F.M.; Pajak, A.; Wolff, M.S.; Pinney, S.M.; Windham, G.C.; Galvez, M.P.; Greenspan, L.C.; Kushi, L.H.; Teitelbaum, S.L. Age of Menarche in a Longitudinal US Cohort. J. Pediatr. Adolesc. Gynecol. 2018, 31, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Benidis, K.D.; Tzortzaki, E.; Georgiou, A.; Zachariadou, T.; Adamidi, T.; Zannetos, S.; Bakakos, P.; Koulouris, N.G.; Rovina, N. Prevalence and Characteristics of Self-Reported Adult Asthma in Cyprus: A Population-Based Observational Study. J. Asthma Allergy 2023, 16, 215–226. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Insulin | Adipokines (Leptin and Adiponectin) | |
|---|---|---|
| Obesity in Boys | Delay in initiation of puberty secondary to the suppression of the HPG axis | Increased inhibition of the HPG axis that further contributes to the delay in the initiation of puberty |
| Obesity in Girls | Acceleration in initiation of puberty secondary to the positive feedback loop exerted on the HPG axis | Earlier activation of the HPG axis that further contributes to the acceleration of the initiation of puberty |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kythreotis, A.K.; Nicolaou, M.; Mitsinga, E.; Daher, H.; Skordis, N. The Interplay Between Body Weight and the Onset of Puberty. Children 2025, 12, 679. https://doi.org/10.3390/children12060679
Kythreotis AK, Nicolaou M, Mitsinga E, Daher H, Skordis N. The Interplay Between Body Weight and the Onset of Puberty. Children. 2025; 12(6):679. https://doi.org/10.3390/children12060679
Chicago/Turabian StyleKythreotis, Alexandros K., Marina Nicolaou, Eirini Mitsinga, Habib Daher, and Nicos Skordis. 2025. "The Interplay Between Body Weight and the Onset of Puberty" Children 12, no. 6: 679. https://doi.org/10.3390/children12060679
APA StyleKythreotis, A. K., Nicolaou, M., Mitsinga, E., Daher, H., & Skordis, N. (2025). The Interplay Between Body Weight and the Onset of Puberty. Children, 12(6), 679. https://doi.org/10.3390/children12060679