
In Vivo Lentiviral Gene Delivery of HLA-DR and Vaccination of Humanized Mice for Improving the Human T and B Cell Immune Reconstitution

 , ,
, ,  , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Primary Cells
2.3. Construction, Production, and Titering of Lentiviral Vectors
2.4. Confirmation of Transgene Expression
2.5. In Vitro Activation of 58-T Cells with DR4+ 3T3 Cells Generated with LV-DR4/fLuc
2.6. Generation of Humanized NRG Mice
2.7. In Vivo Administration of LVs into Mice and Longitudinal BLI Analyses
2.8. Flow Cytometry Analyses of Human T and B Cells
2.9. ELISA Analyses for Detection of Anti-gB IgG in Mouse Plasma
2.10. Statistical Analyses
3. Results
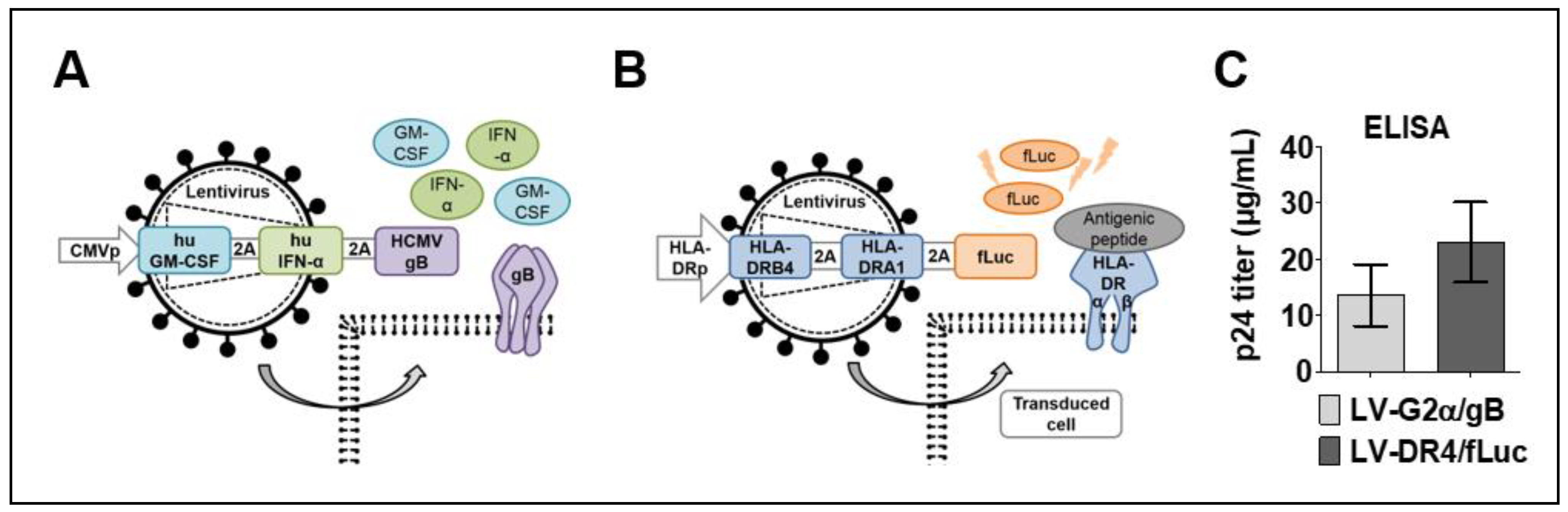
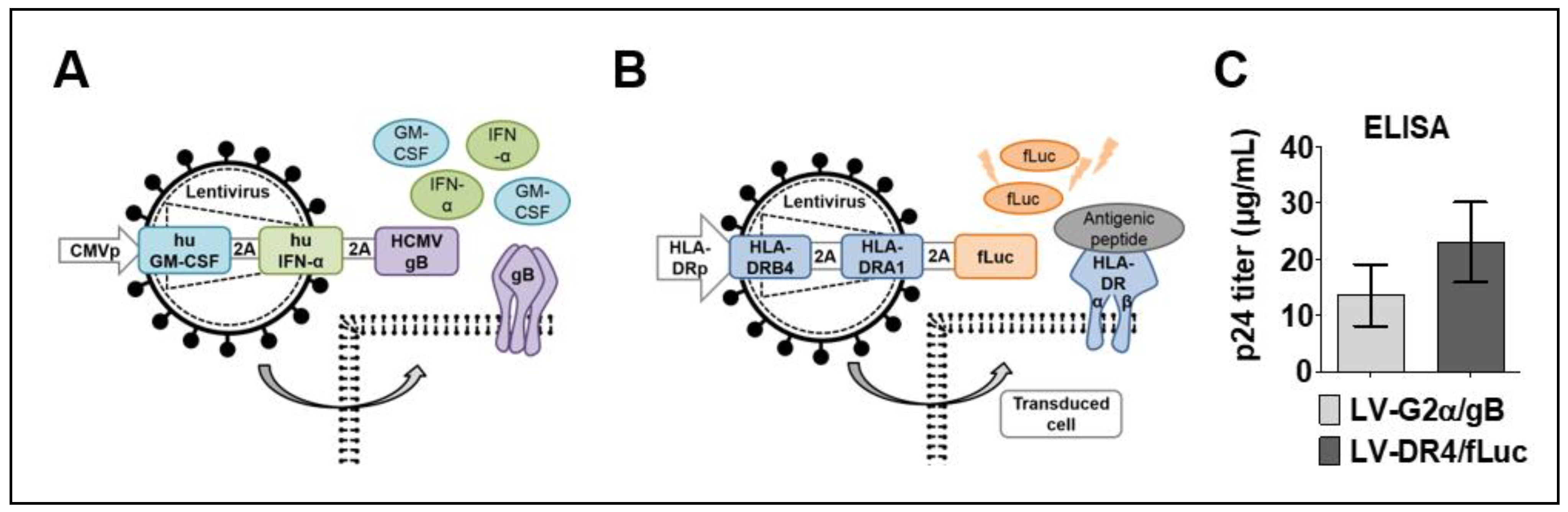
3.1. Lentiviral Vector Designs and Titer
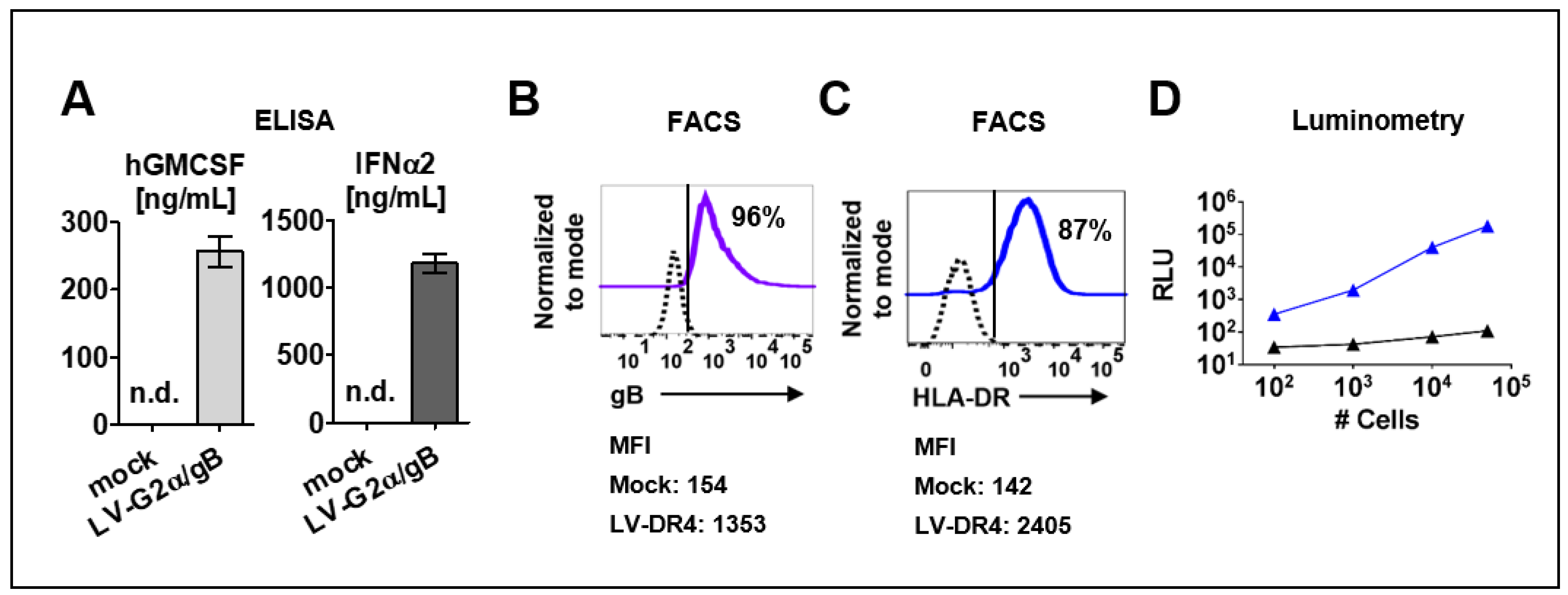
3.2. Detection of Transgenes in Transduced 293T and 3T3 Cells
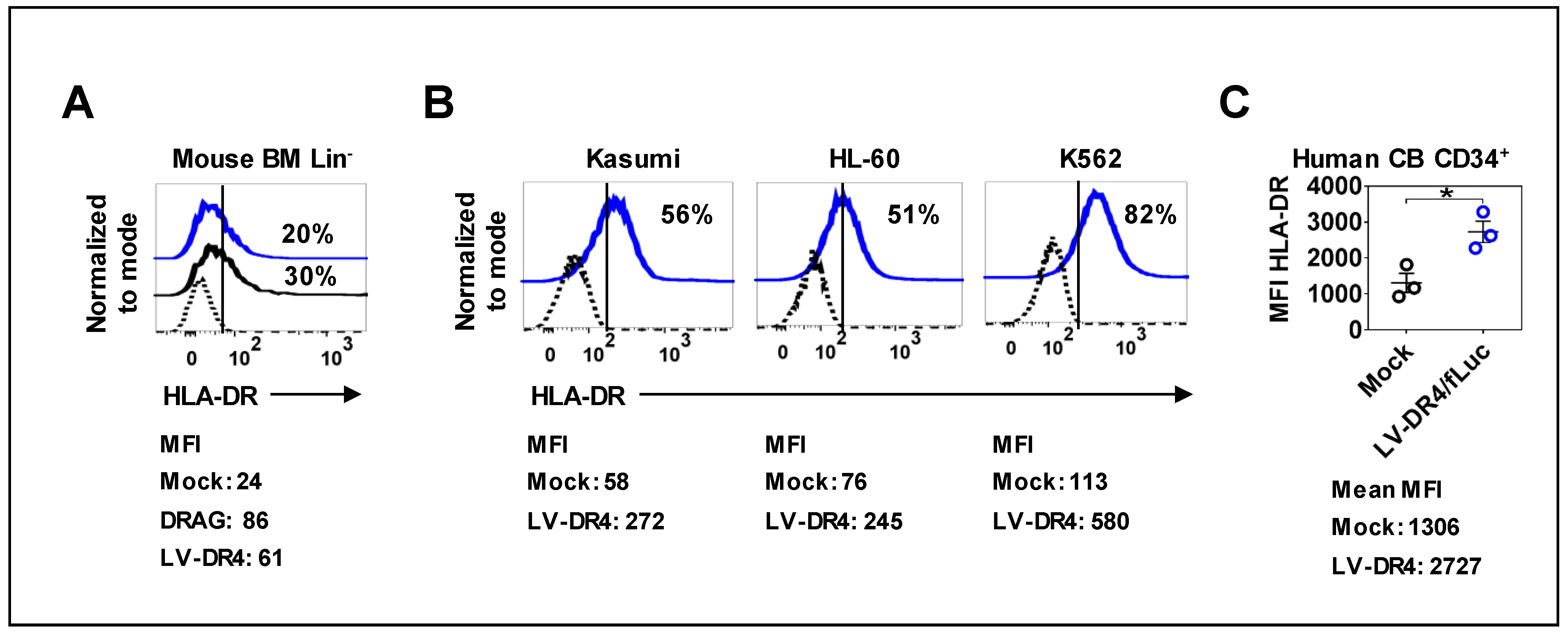
3.3. Mouse and Human Hematopoietic Cells Express DR4 after Transduction with LV-DR4/fLuc
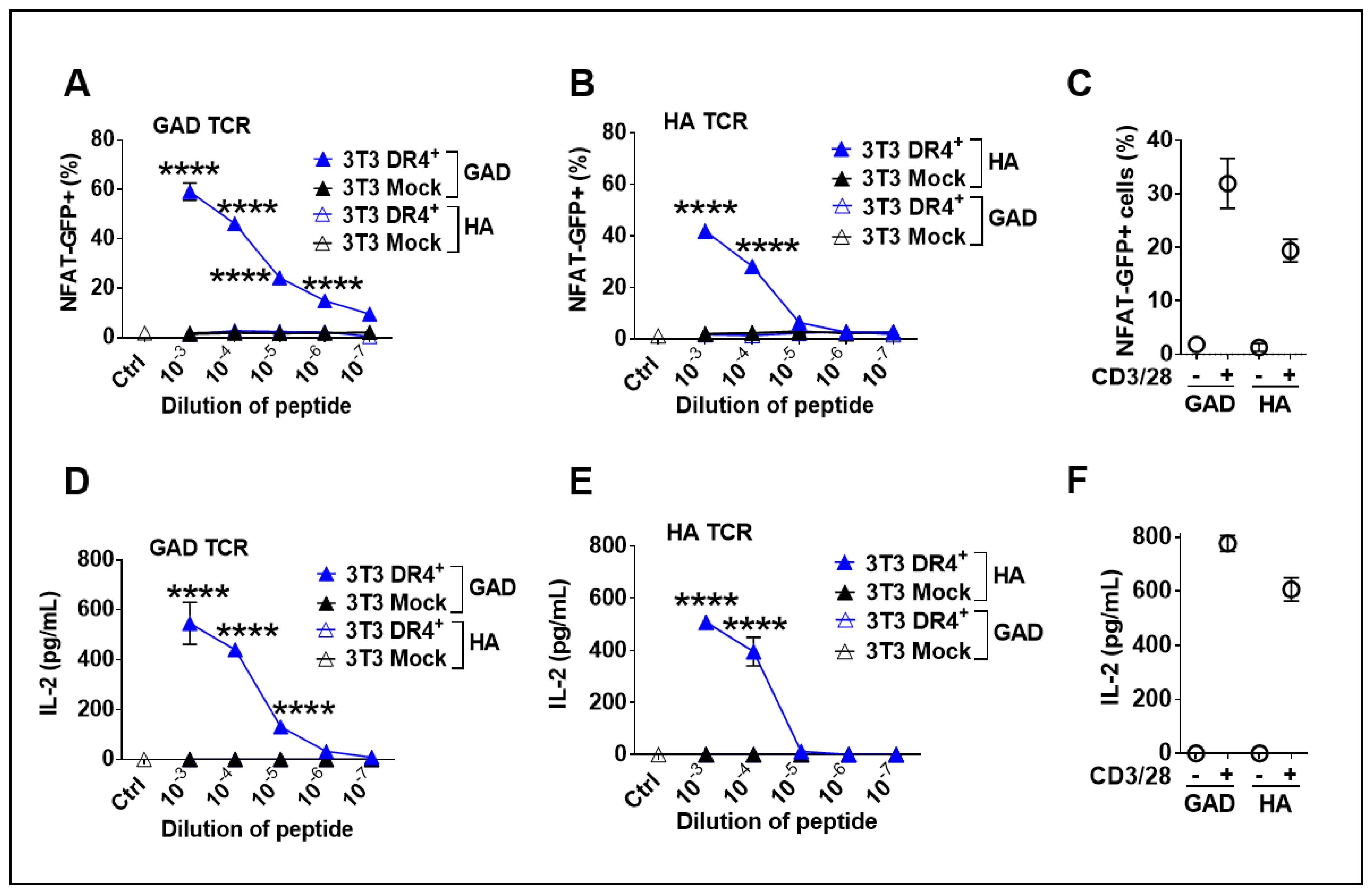
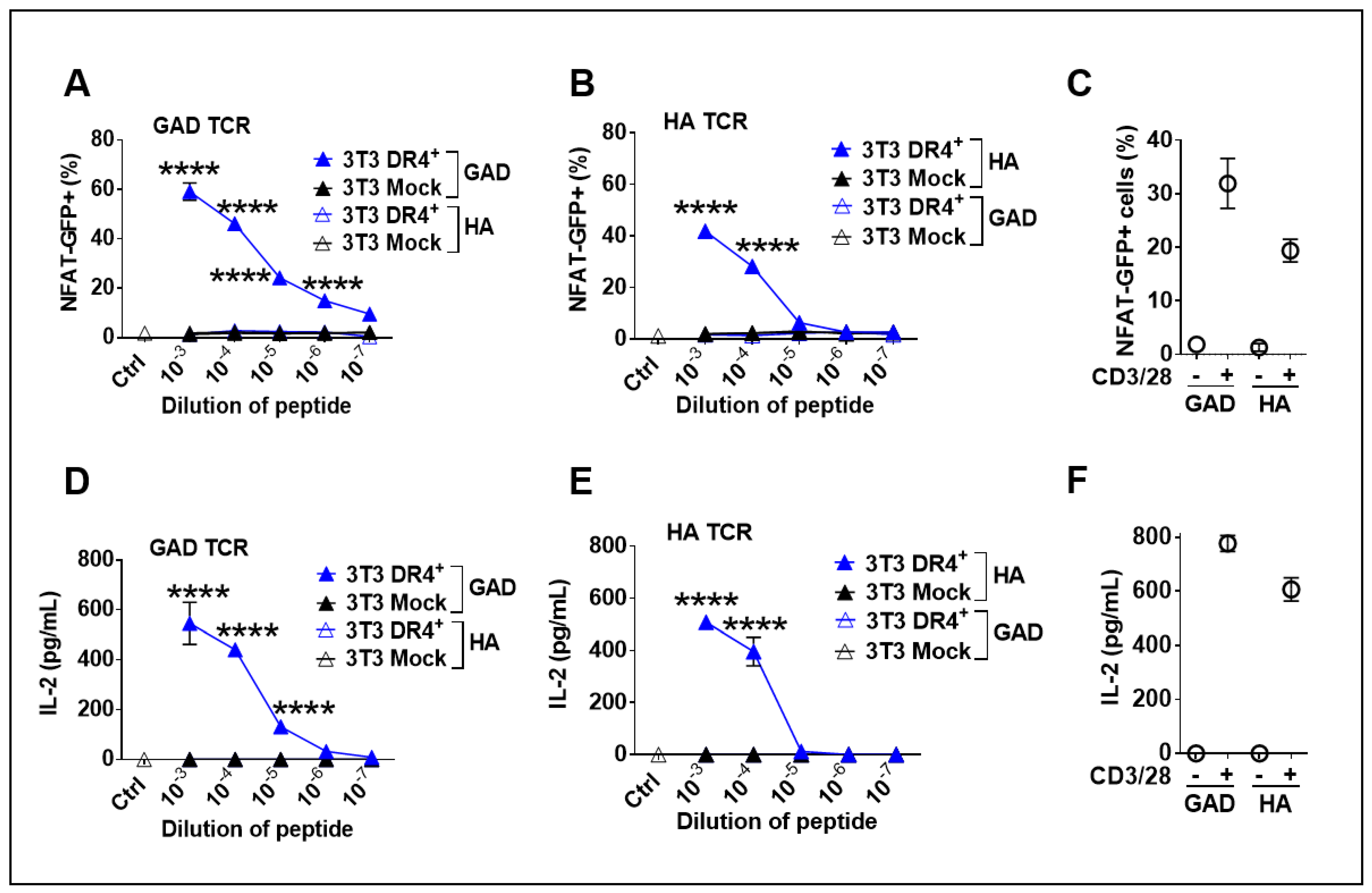
3.4. Functionality of Transgenic DR4 to Present Peptides to DR4-Restricted T Cells
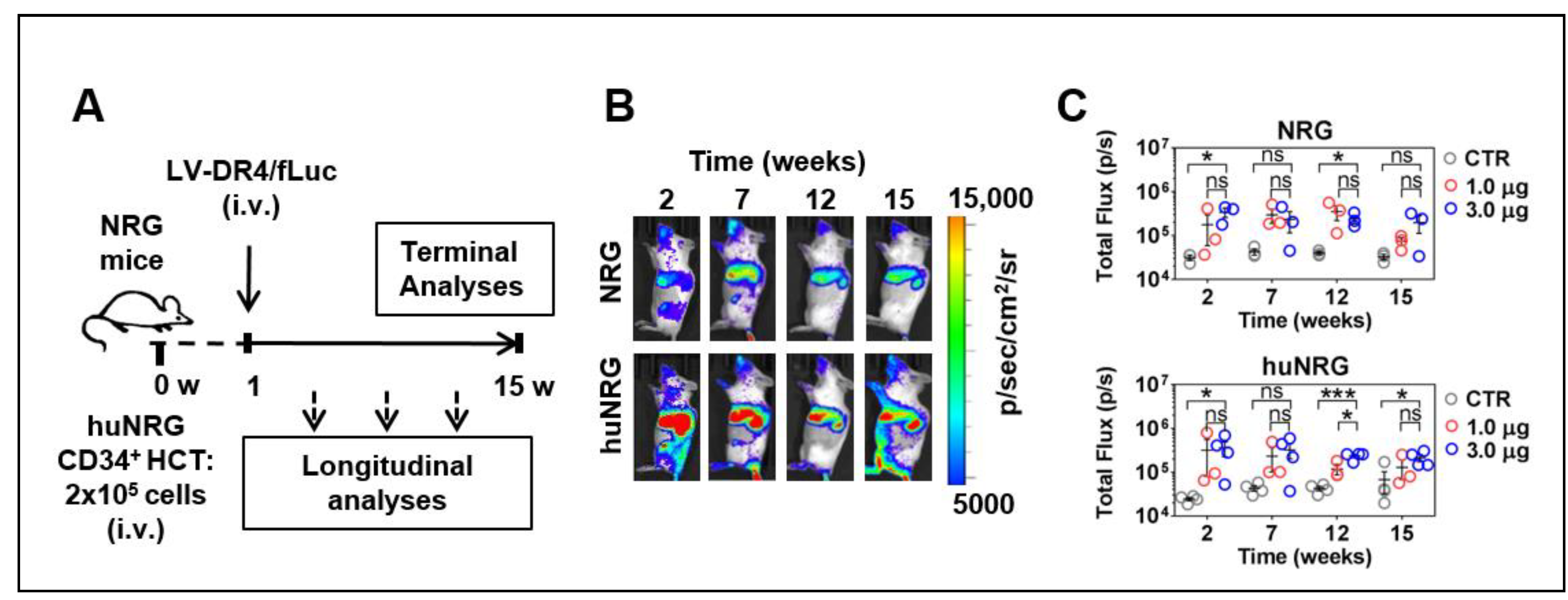
3.5. LV-DR4/fLuc Applied i.v. into NRG and huNRG Mice and In Vivo Transduction
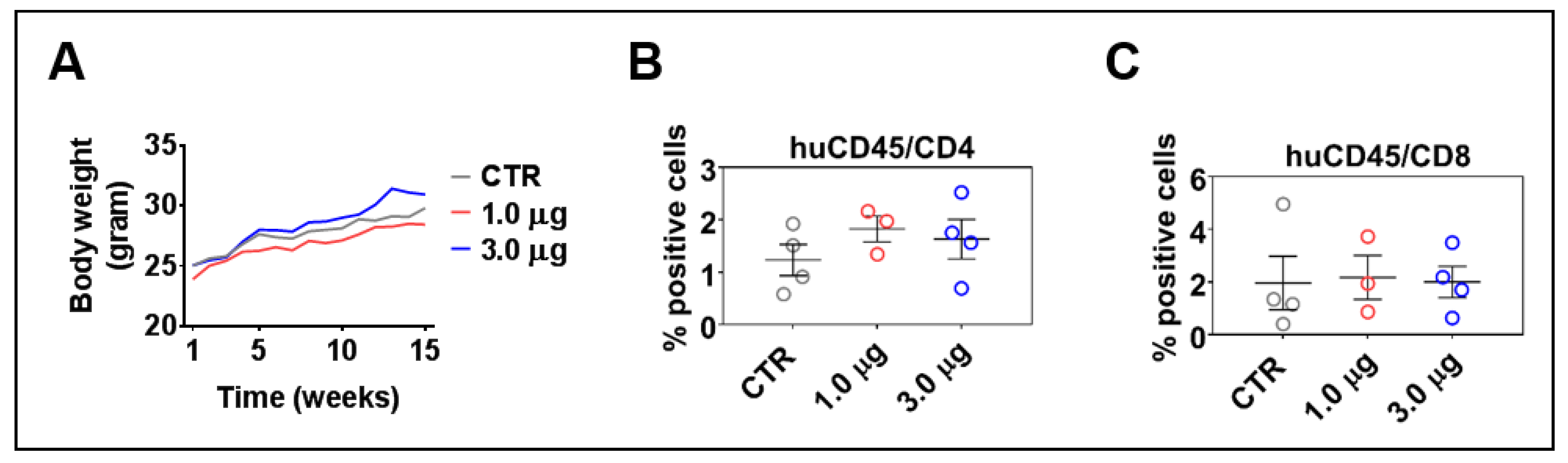
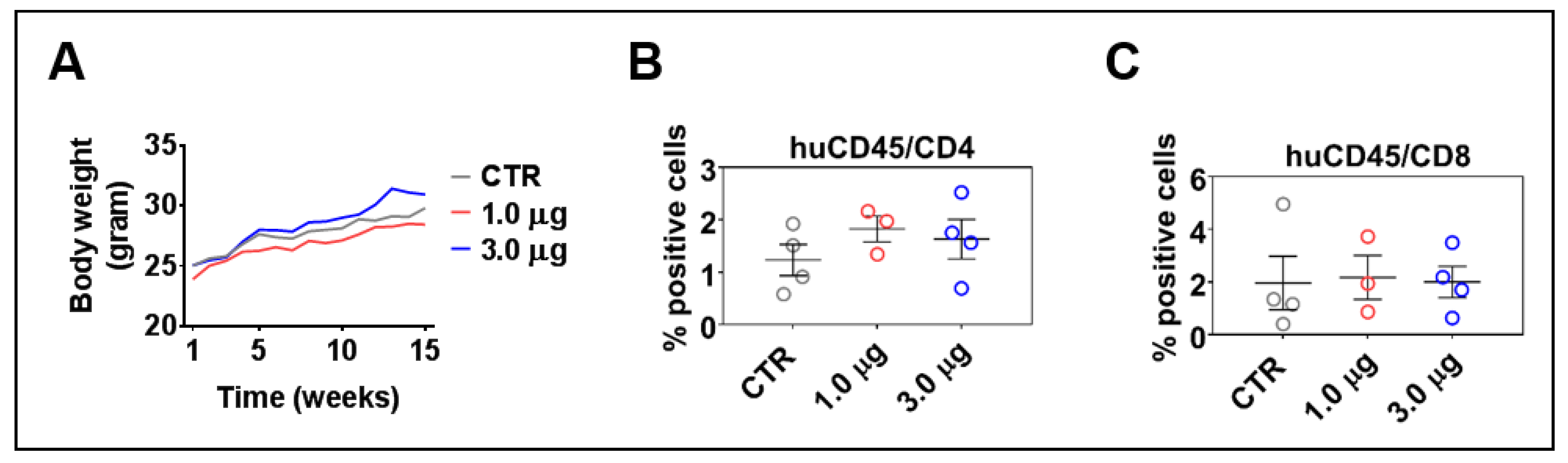
3.6. LV-DR4/fLuc i.v. Delivery Does Not Lead to Acute or Chronic Graft-versus-Host Disease
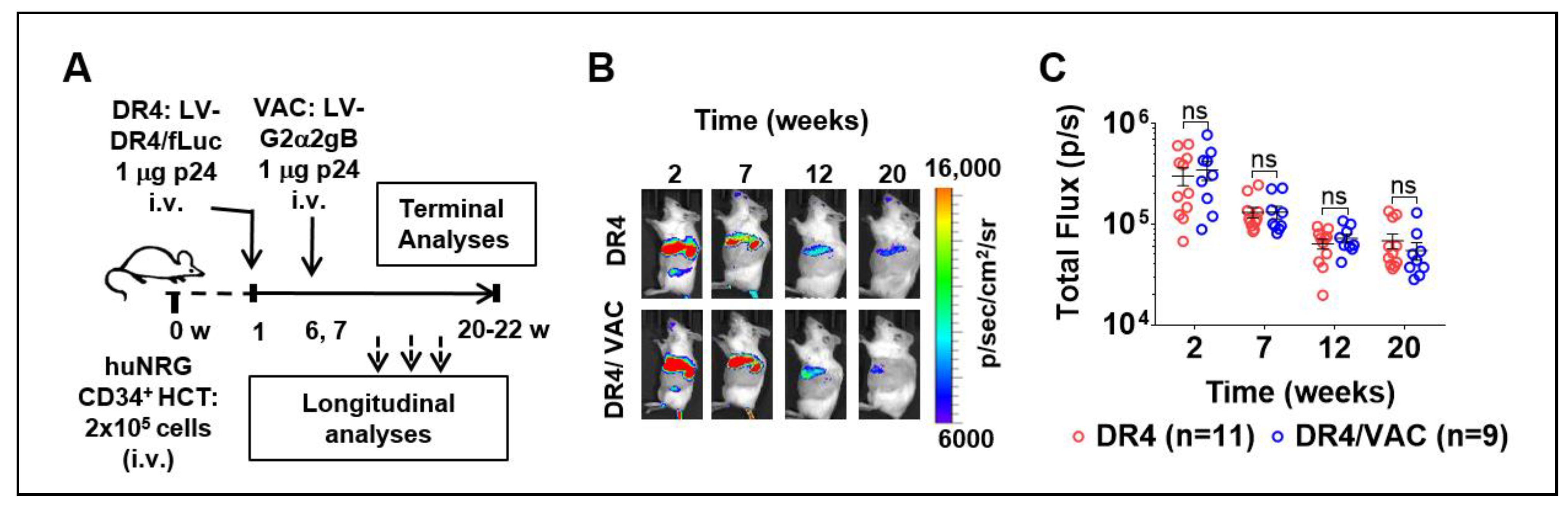
3.7. Feasibility of LV-DR4/fLuc i.v. Application (DR4) Combined with LV-G2α/gB (VAC)
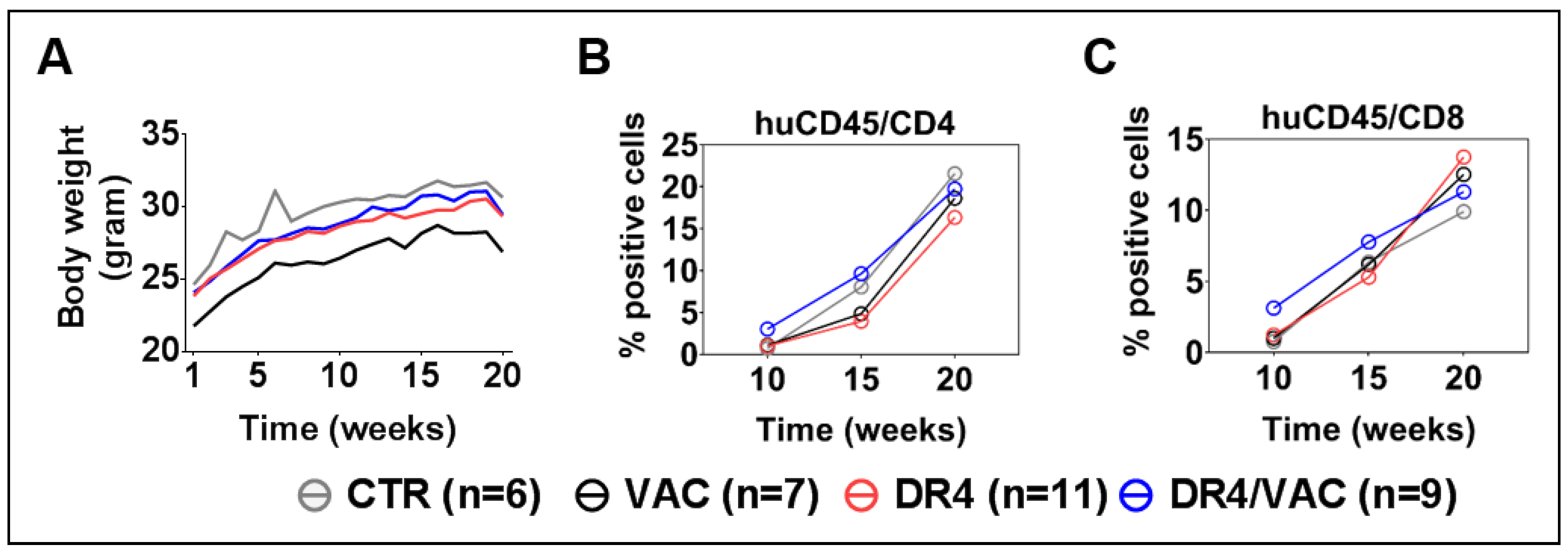
3.8. DR4 Combined with VAC Does Not Cause GvHD but Results in a Transient Increase in T Cell Frequencies
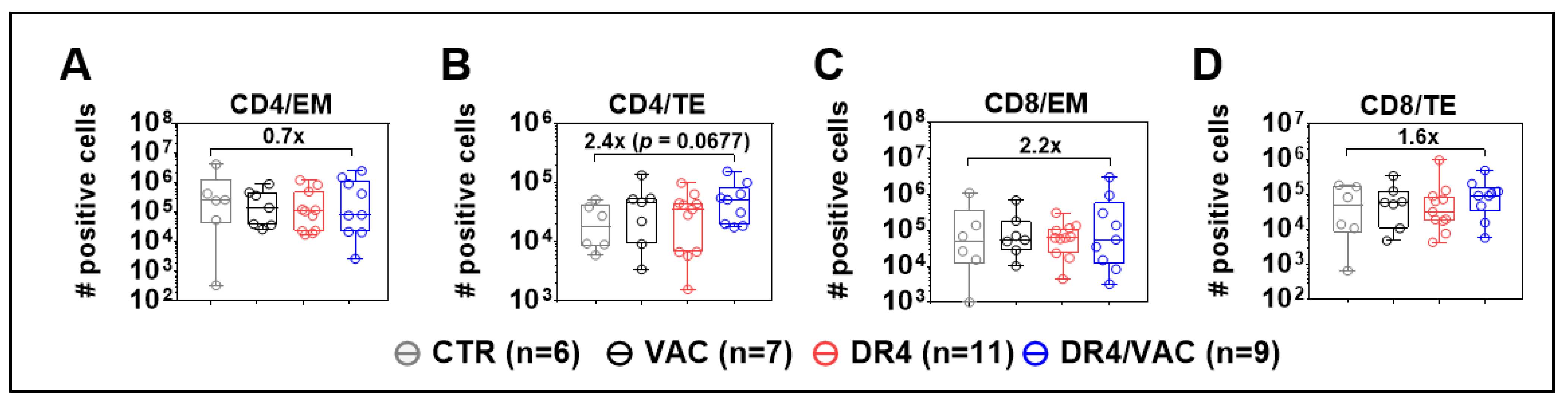
3.9. DR4 Combined with VAC Promotes CD4+ and CD8+ T Cell Maturation
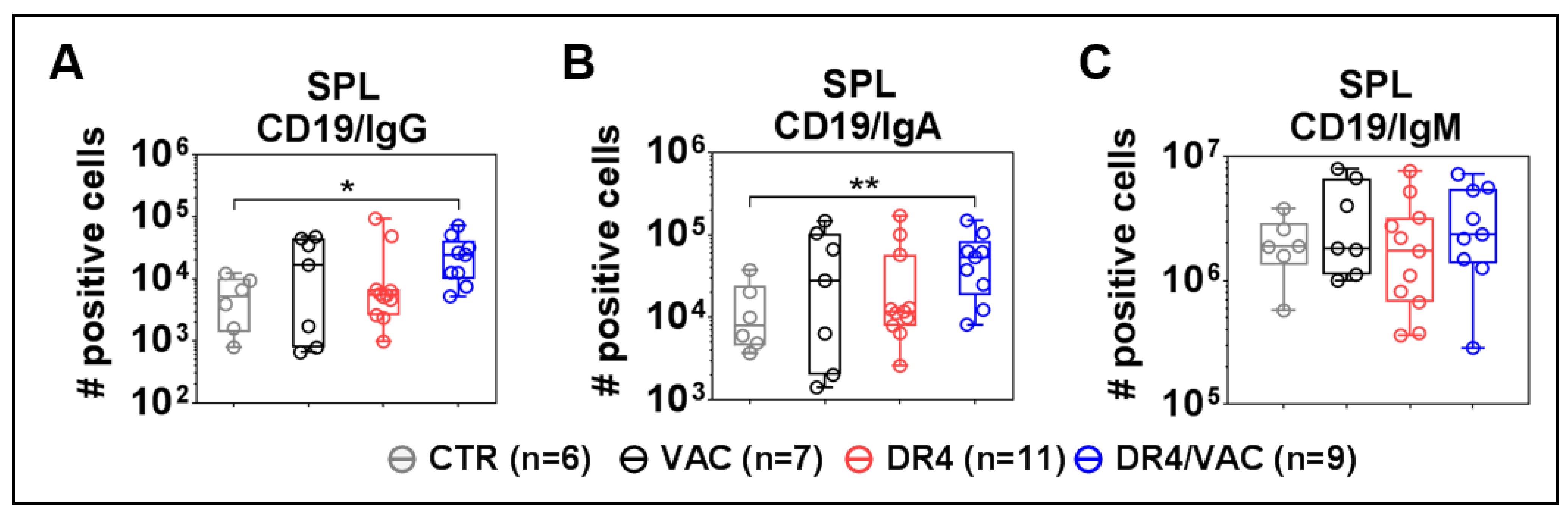
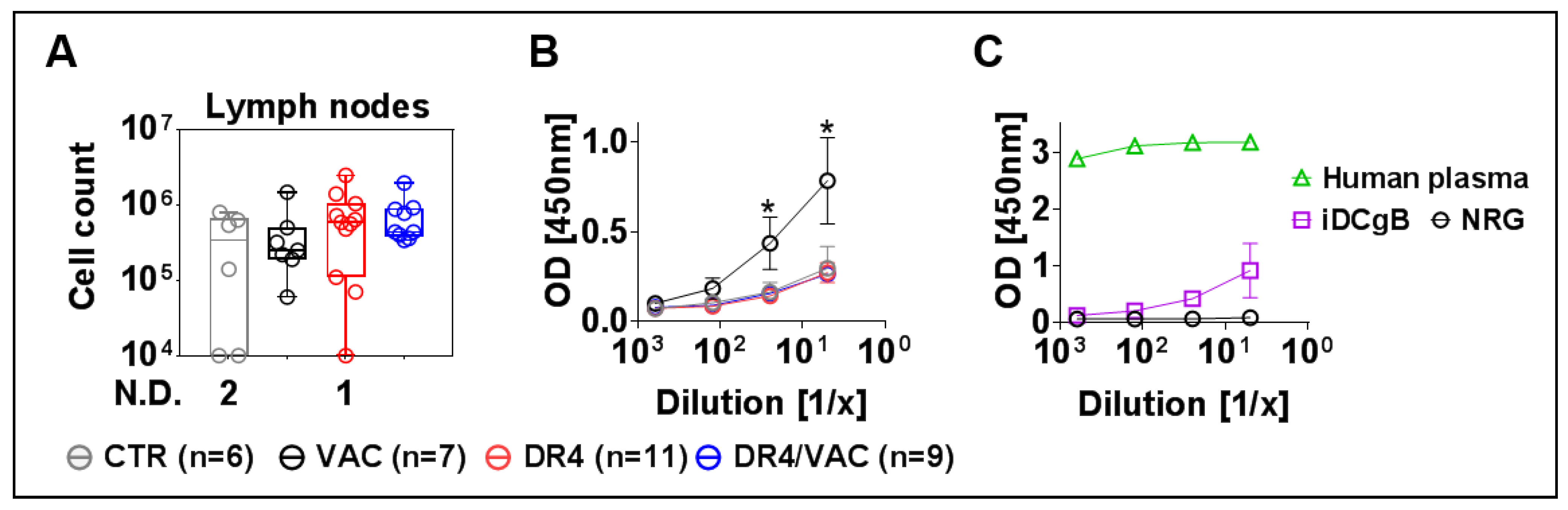
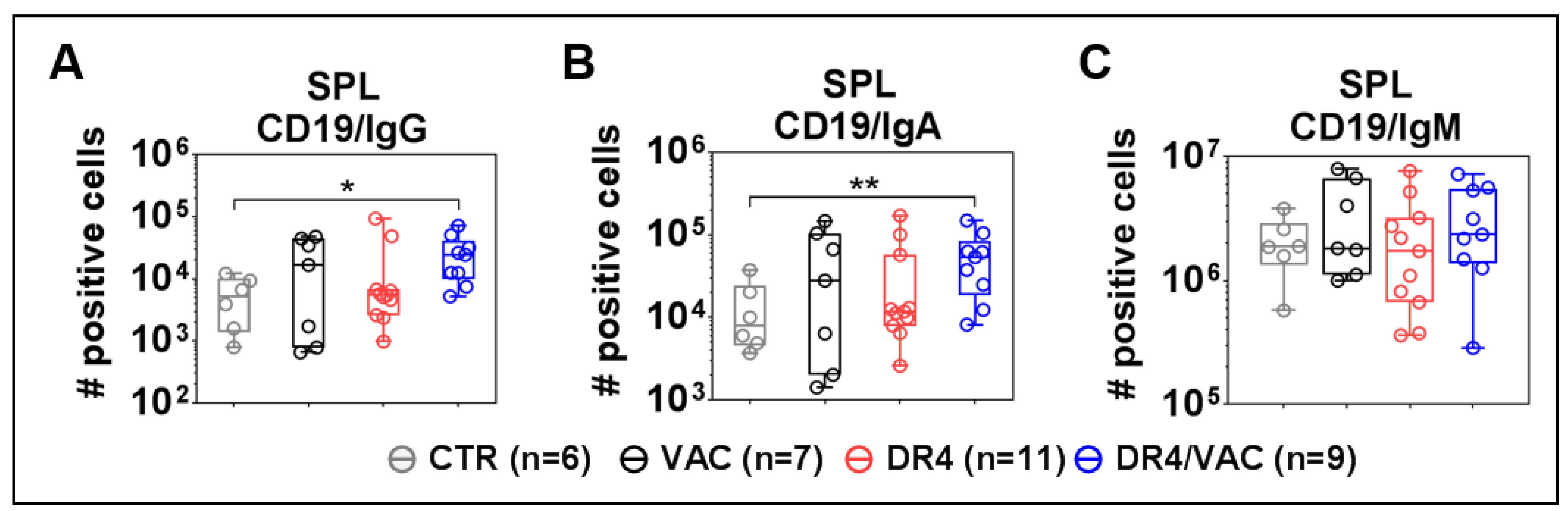
3.10. DR4 Combined with VAC Promote B Cell Class Switch
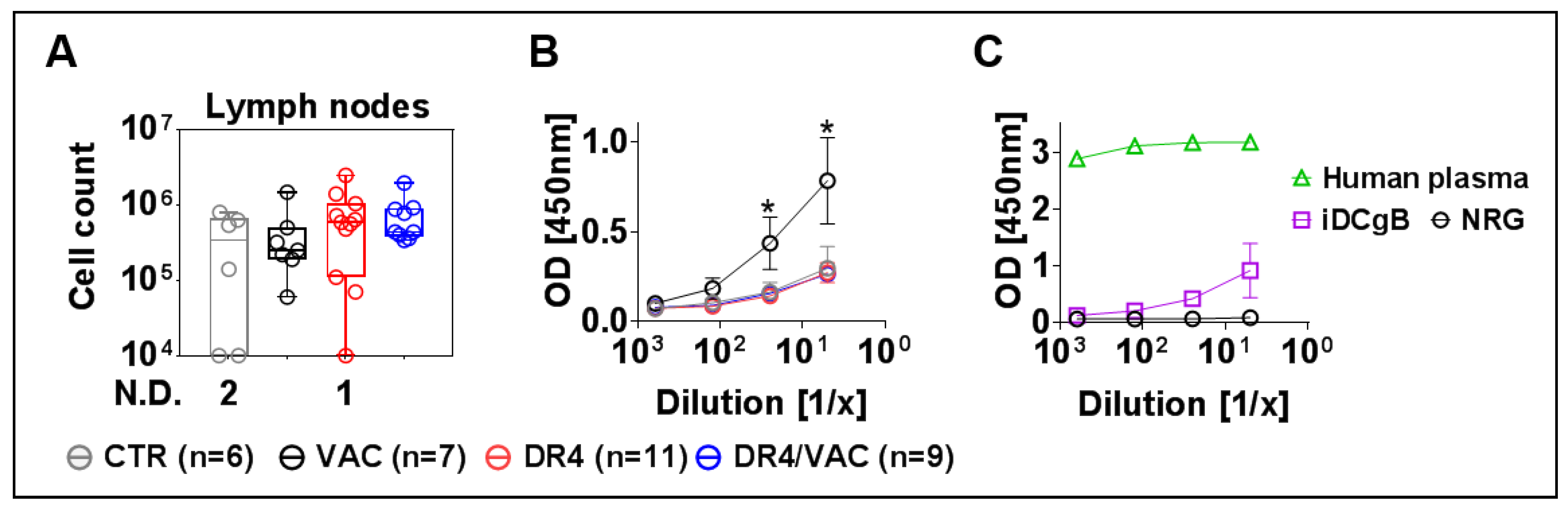
3.11. VAC Promotes IgG Humoral Responses against gB
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCune, J.; Namikawa, R.; Kaneshima, H.; Shultz, L.; Lieberman, M.; Weissman, I. The SCID-hu mouse: Murine model for the analysis of human hematolymphoid differentiation and function. Science 1988, 241, 1632–1639. [Google Scholar] [CrossRef]
- Stripecke, R.; Münz, C.; Schuringa, J.J.; Bissig, K.; Soper, B.; Meeham, T.; Yao, L.; Di Santo, J.P.; Brehm, M.; Rodriguez, E.; et al. Innovations, challenges, and minimal information for standardization of humanized mice. EMBO Mol. Med. 2020, 12, e8662. [Google Scholar] [CrossRef]
- Shultz, L.D.; Brehm, M.; Garcia-Martinez, J.V.; Greiner, D.L. Humanized mice for immune system investigation: Progress, promise and challenges. Nat. Rev. Immunol. 2012, 12, 786–798. [Google Scholar] [CrossRef]
- Wahl, A.; De, C.; Fernandez, M.A.; Lenarcic, E.M.; Xu, Y.; Cockrell, A.S.; Cleary, R.A.; Johnson, C.E.; Schramm, N.J.; Rank, L.M.; et al. Precision mouse models with expanded tropism for human pathogens. Nat. Biotechnol. 2019, 37, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.; Brehm, M.; Bridges, S.; Ferguson, S.; Kumar, P.; Mirochnitchenko, O.; Palucka, K.; Pelanda, R.; Sanders-Beer, B.; Shultz, L.D.; et al. Humanized immune system mouse models: Progress, challenges and opportunities. Nat. Immunol. 2019, 20, 770–774. [Google Scholar] [CrossRef]
- Steinman, R.M. Decisions About Dendritic Cells: Past, Present, and Future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wendland, M.; Willenzon, S.; Kocks, J.; Davalos-Misslitz, A.C.; Hammerschmidt, S.I.; Schumann, K.; Kremmer, E.; Sixt, M.; Hoffmeyer, A.; Pabst, O.; et al. Lymph Node T Cell Homeostasis Relies on Steady State Homing of Dendritic Cells. Immunity 2011, 35, 945–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, R.E.; Richardson, E.K.; Toh, H.C. Revisiting the role of CD4+ T cells in cancer immunotherapy—New insights into old paradigms. Cancer Gene Ther. 2020, 28, 5–17. [Google Scholar] [CrossRef]
- Chen, D.S.; Tang, T.F.; Pulyaeva, H.; Slack, R.; Tu, B.; Wagage, D.; Li, L.; Perlee, L.; Ng, J.; Hartzman, R.J.; et al. Relative hla-drb1*04 allele frequencies in five united states populations found in a hematopoietic stem cell volunteer donor registry and seven new drb1*04 alleles. Hum. Immunol. 2002, 63, 665–672. [Google Scholar] [CrossRef]
- Danner, R.; Chaudhari, S.N.; Rosenberger, J.; Surls, J.; Richie, T.; Brumeanu, T.-D.; Casares, S. Expression of HLA Class II Molecules in Humanized NOD.Rag1KO.IL2RgcKO Mice Is Critical for Development and Function of Human T and B Cells. PLoS ONE 2011, 6, e19826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, M.; Takahashi, T.; Katano, I.; Ito, R.; Ito, M.; Harigae, H.; Ishii, N.; Sugamura, K. Induction of human humoral immune responses in a novel hla-dr-expressing transgenic nod/shi-scid/gammacnull mouse. Int. Immunol. 2012, 24, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Peachman, K.K.; Jobe, O.; Morrison, E.B.; Allam, A.; Jagodzinski, L.; Casares, S.A.; Rao, M. Tracking Human Immunodeficiency Virus-1 Infection in the Humanized DRAG Mouse Model. Front. Immunol. 2017, 8, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, M.; Ballesteros, A.; Qiu, Q.; Sang, L.P.; Shashikumar, S.; Casares, S.; Brumeanu, T.-D. Generation and testing anti-influenza human monoclonal antibodies in a new humanized mouse model (DRAGA: HLA-AHLA-DRRag1 KO. IL-2Rγc KO. NOD). Hum. Vaccines Immunother. 2017, 14, 345–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salguero, G.; Daenthanasanmak, A.; Münz, C.; Raykova, A.; Guzmán, C.A.; Riese, P.; Figueiredo, C.; Länger, F.; Schneider, A.; Macke, L.; et al. Dendritic Cell–Mediated Immune Humanization of Mice: Implications for Allogeneic and Xenogeneic Stem Cell Transplantation. J. Immunol. 2014, 192, 4636–4647. [Google Scholar] [CrossRef] [Green Version]
- Daenthanasanmak, A.; Salguero, G.; Sundarasetty, B.S.; Waskow, C.; Cosgun, K.; Guzman, C.A.; Riese, P.; Gerasch, L.; Schneider, A.; Ingendoh, A.; et al. Engineered dendritic cells from cord blood and adult blood accelerate effector T cell immune reconstitution against HCMV. Mol. Ther. Methods Clin. Dev. 2015, 2, 14060. [Google Scholar] [CrossRef]
- Volk, V.; Reppas, A.I.; Robert, P.A.; Spineli, L.; Sundarasetty, B.S.; Theobald, S.J.; Schneider, A.; Gerasch, L.; Roth, C.D.; Klöss, S.; et al. Multidimensional Analysis Integrating Human T-Cell Signatures in Lymphatic Tissues with Sex of Humanized Mice for Prediction of Responses after Dendritic Cell Immunization. Front. Immunol. 2017, 8, 1709. [Google Scholar] [CrossRef] [Green Version]
- Theobald, S.J.; Kreer, C.; Khailaie, S.; Bonifacius, A.; Eiz-Vesper, B.; Figueiredo, C.; Mach, M.; Backovic, M.; Ballmaier, M.; Koenig, J.; et al. Repertoire characterization and validation of gb-specific human iggs directly cloned from humanized mice vaccinated with dendritic cells and protected against hcmv. PLoS Pathog. 2020, 16, e1008560. [Google Scholar] [CrossRef]
- Stripecke, R. Lentivirus-Induced Dendritic Cells (iDC) for Immune-Regenerative Therapies in Cancer and Stem Cell Transplantation. Biomedicines 2014, 2, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Bialek-Waldmann, J.K.; Domning, S.; Esser, R.; Glienke, W.; Mertens, M.; Aleksandrova, K.; Arseniev, L.; Kumar, S.; Schneider, A.; Koenig, J.; et al. Induced dendritic cells co-expressing GM-CSF/IFN-α/tWT1 priming T and B cells and automated manufacturing to boost GvL. Mol. Ther. Methods Clin. Dev. 2021, 21, 621–641. [Google Scholar] [CrossRef]
- Olbrich, H.; Slabik, C.; Stripecke, R. Reconstructing the immune system with lentiviral vectors. Virus Genes 2017, 53, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Pincha, M.; Sundarasetty, B.S.; Stripecke, R. Lentiviral vectors for immunization: An inflammatory field. Expert Rev. Vaccines 2010, 9, 309–321. [Google Scholar] [CrossRef]
- Pfeiffer, A.; Thalheimer, F.B.; Hartmann, S.; Frank, A.M.; Bender, R.R.; Danisch, S.; Costa, C.; Wels, W.S.; Modlich, U.; Stripecke, R.; et al. In vivo generation of human CD 19- CAR T cells results in B-cell depletion and signs of cytokine release syndrome. EMBO Mol. Med. 2018, 10, e9158. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Koya, R.C.; Anselmi, L.; Sternini, C.; Wang, H.-J.; Comin-Anduix, B.; Prins, R.; Faure-Kumar, E.; Rozengurt, N.; Cui, Y.; et al. Lentiviral Vectors with CMV or MHCII Promoters Administered In Vivo: Immune Reactivity Versus Persistence of Expression. Mol. Ther. 2007, 15, 1390–1399. [Google Scholar] [CrossRef]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-Inactivating Lentivirus Vector for Safe and Efficient In Vivo Gene Delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A Third-Generation Lentivirus Vector with a Conditional Packaging System. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Z.; Elledge, S.J. Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. Nat. Methods 2007, 4, 251–256. [Google Scholar] [CrossRef]
- Godbey, W.; Wu, K.K.; Mikos, A.G. Poly(ethylenimine) and its role in gene delivery. J. Control. Release 1999, 60, 149–160. [Google Scholar] [CrossRef]
- Boddul, S.V.; Sharma, R.K.; Dubnovitsky, A.; Raposo, B.; Gerstner, C.; Shen, Y.; Iyer, V.S.; Kasza, Z.; Kwok, W.W.; Winkler, A.R.; et al. In vitro and ex vivo functional characterization of human HLA-DRB1∗04 restricted T cell receptors. J. Transl. Autoimmun. 2021, 4, 100087. [Google Scholar] [CrossRef]
- Pearson, T.; Shultz, L.D.; Miller, D.; King, M.; Laning, J.; Fodor, W.; Cuthbert, A.; Burzenski, L.; Gott, B.; Lyons, B.; et al. Non-obese diabetic-recombination activating gene-1 (nod-rag1 null) interleukin (il)-2 receptor common gamma chain (il2r gamma null) null mice: A radioresistant model for human lymphohaematopoietic engraftment. Clin. Exp. Immunol. 2008, 154, 270–284. [Google Scholar] [CrossRef]
- Danisch, S.; Slabik, C.; Cornelius, A.; Albanese, M.; Tagawa, T.; Chen, Y.-F.A.; Krönke, N.; Eiz-Vesper, B.; Lienenklaus, S.; Bleich, A.; et al. Spatiotemporally Skewed Activation of Programmed Cell Death Receptor 1–Positive T Cells after Epstein-Barr Virus Infection and Tumor Development in Long-Term Fully Humanized Mice. Am. J. Pathol. 2019, 189, 521–539. [Google Scholar] [CrossRef] [Green Version]
- Sundarasetty, B.; Volk, V.; Theobald, S.J.; Rittinghausen, S.; Schaudien, D.; Neuhaus, V.; Figueiredo, C.; Schneider, A.; Gerasch, L.; Mucci, A.; et al. Human Effector Memory T Helper Cells Engage with Mouse Macrophages and Cause Graft-versus-Host–Like Pathology in Skin of Humanized Mice Used in a Nonclinical Immunization Study. Am. J. Pathol. 2017, 187, 1380–1398. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.H.; Fan, Y.; Wang, X.H.; Mao, Q.; Deng, G.H.; Wang, Y.M. Relationship between hla-dr gene polymorphisms and outcomes of hepatitis b viral infections: A meta-analysis. World J. Gastroenterol. 2012, 18, 3119–3128. [Google Scholar] [CrossRef]
- Luckey, D.; Weaver, E.A.; Osborne, D.G.; Billadeau, D.D.; Taneja, V. Immunity to Influenza is dependent on MHC II polymorphism: Study with 2 HLA transgenic strains. Sci. Rep. 2019, 9, 19061. [Google Scholar] [CrossRef] [Green Version]
- Lanchbury, J.S.; Jaeger, E.E.; Sansom, D.M.; Hall, M.A.; Wordsworth, P.; Stedeford, J.; Bell, J.I.; Panayi, G.S. Strong primary selection for the dw4 subtype of dr4 accounts for the hla-dqw7 association with felty’s syndrome. Hum. Immunol. 1991, 32, 56–64. [Google Scholar] [CrossRef]
- Taneja, V.; Behrens, M.; Basal, E.; Sparks, J.; Griffiths, M.M.; Luthra, H.; David, C.S. Delineating the Role of the HLA-DR4 “Shared Epitope” in Susceptibility versus Resistance to Develop Arthritis. J. Immunol. 2008, 181, 2869–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melchers, F. Checkpoints that control B cell development. J. Clin. Investig. 2015, 125, 2203–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, J.; Theobald, S.J.; Stripecke, R. Modeling Human Cytomegalovirus in Humanized Mice for Vaccine Testing. Vaccines 2020, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theobald, S.J.; Khailaie, S.; Meyer-Hermann, M.; Volk, V.; Olbrich, H.; Danisch, S.; Gerasch, L.; Schneider, A.; Sinzger, C.; Schaudien, D.; et al. Signatures of T and B Cell Development, Functional Responses and PD-1 Upregulation After HCMV Latent Infections and Reactivations in Nod.Rag.Gamma Mice Humanized with Cord Blood CD34+ Cells. Front. Immunol. 2018, 9, 2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marodon, G.; Fisson, S.; Levacher, B.; Fabre, M.; Salomon, B.; Klatzmann, D. Induction of antigen-specific tolerance by intrathymic injection of lentiviral vectors. Blood 2006, 108, 2972–2978. [Google Scholar] [CrossRef] [PubMed]
- Marodon, G.; Klatzmann, D. In situ transduction of stromal cells and thymocytes upon intrathymic injection of lentiviral vectors. BMC Immunol. 2004, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Brehm, M.A.; Kenney, L.L.; Wiles, M.V.; Low, B.E.; Tisch, R.M.; Burzenski, L.; Mueller, C.; Greiner, D.L.; Shultz, L.D. Lack of acute xenogeneic graft- versus-host disease, but retention of t-cell function following engraftment of human peripheral blood mononuclear cells in nsg mice deficient in mhc class i and ii expression. FASEB J. 2019, 33, 3137–3151. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Koenig, J.; Schneider, A.; Wermeling, F.; Boddul, S.; Theobald, S.J.; Vollmer, M.; Kloos, D.; Lachmann, N.; Klawonn, F.; et al. In Vivo Lentiviral Gene Delivery of HLA-DR and Vaccination of Humanized Mice for Improving the Human T and B Cell Immune Reconstitution. Biomedicines 2021, 9, 961. https://doi.org/10.3390/biomedicines9080961
Kumar S, Koenig J, Schneider A, Wermeling F, Boddul S, Theobald SJ, Vollmer M, Kloos D, Lachmann N, Klawonn F, et al. In Vivo Lentiviral Gene Delivery of HLA-DR and Vaccination of Humanized Mice for Improving the Human T and B Cell Immune Reconstitution. Biomedicines. 2021; 9(8):961. https://doi.org/10.3390/biomedicines9080961
Chicago/Turabian StyleKumar, Suresh, Johannes Koenig, Andreas Schneider, Fredrik Wermeling, Sanjaykumar Boddul, Sebastian J. Theobald, Miriam Vollmer, Doreen Kloos, Nico Lachmann, Frank Klawonn, and et al. 2021. "In Vivo Lentiviral Gene Delivery of HLA-DR and Vaccination of Humanized Mice for Improving the Human T and B Cell Immune Reconstitution" Biomedicines 9, no. 8: 961. https://doi.org/10.3390/biomedicines9080961
APA StyleKumar, S., Koenig, J., Schneider, A., Wermeling, F., Boddul, S., Theobald, S. J., Vollmer, M., Kloos, D., Lachmann, N., Klawonn, F., Lienenklaus, S., Talbot, S. R., Bleich, A., Wenzel, N., von Kaisenberg, C., Keck, J., & Stripecke, R. (2021). In Vivo Lentiviral Gene Delivery of HLA-DR and Vaccination of Humanized Mice for Improving the Human T and B Cell Immune Reconstitution. Biomedicines, 9(8), 961. https://doi.org/10.3390/biomedicines9080961