
What Is Known about Theragnostic Strategies in Colorectal Cancer

 ,
,  and
and
Abstract
1. Introduction
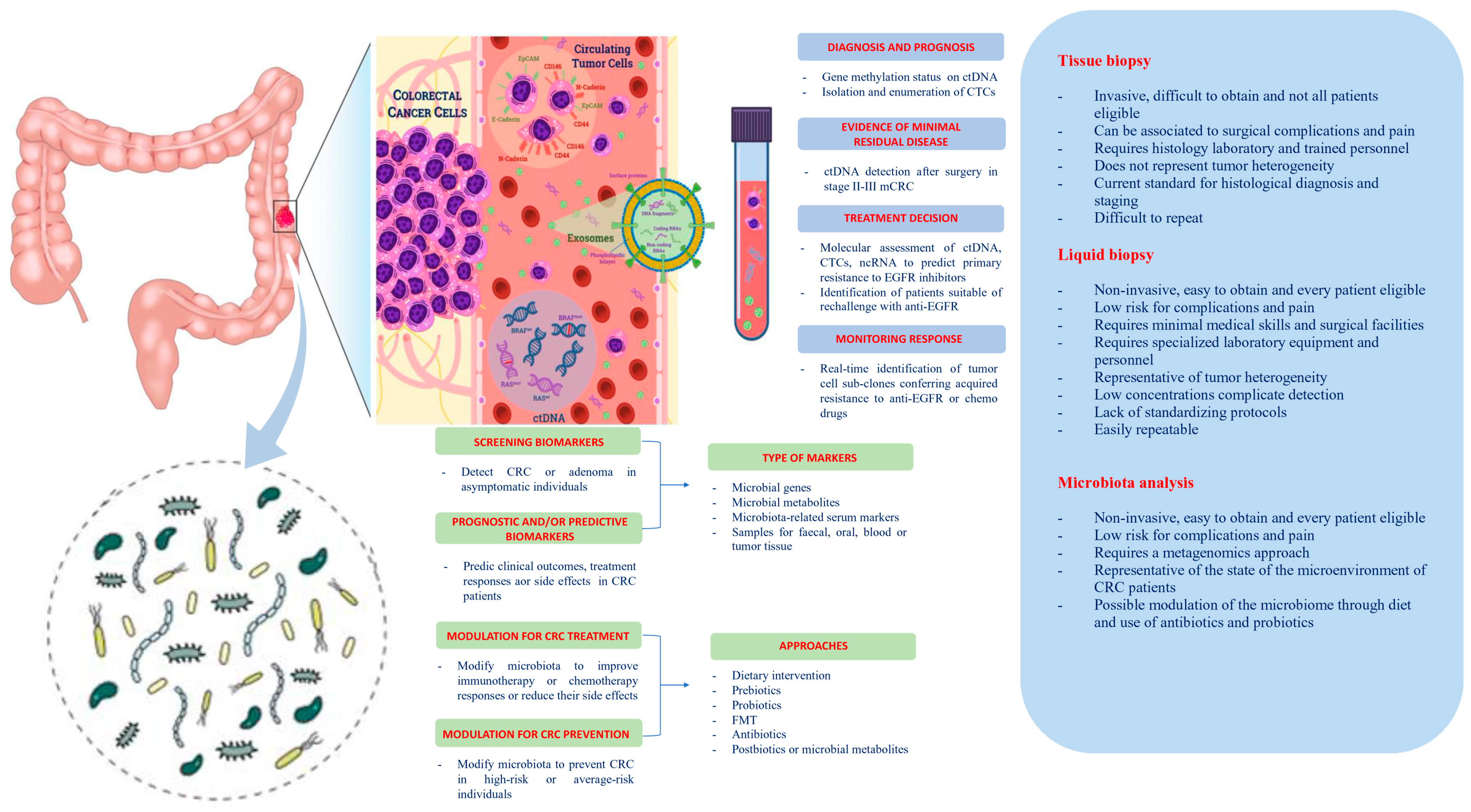
2. Liquid Biopsy
2.1. Circulating Tumor Cells (CTCs)
2.1.1. Screening and Early Diagnosis
2.1.2. Prognostic and Predictive Factor, Staging Tool and Guide for Systemic Treatment, Resistance Evaluation, and MRD Assessment
2.2. Circulating Tumor DNA (ctDNA)
2.2.1. Screening and Early Diagnosis
2.2.2. Prognostic and Predictive Factor, Staging Tool, and Guide for Systemic Treatment, Resistance Evaluation, and MRD Assessment
2.3. MicroRNAs (miRNAs) and Long Non-Coding RNAs (lncRNAs)
2.3.1. Diagnosis and Prognosis
2.3.2. Drug Resistance
3. Microbiota
3.1. Microbiota and Cancer
3.2. Signaling Pathways Activated in Microbiota and Cancer
3.3. Microbiota and Efficacy of Anticancer Agents
3.4. Recent Advances in Metagenomics Technology for Diagnosis and Prognosis
3.5. Organoids Engineering
3.6. Therapeutic Use of Antibiotics, Probiotics, and Fecal Microbiota Transplantation
4. Future Perspectives and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 5-FU | 5-Fluorouracil |
| ACT | Adjuvant Chemotherapy |
| Akt | Protein Kinase B |
| APC | Adenomatous Polyposis Coli |
| APCMin | Multiple Intestinal Neoplasia, a mutant allele of the Murine Adenomatous Polyposis Coli Locus |
| ATM | Ataxia Telangiectasia Mutated Serine/Threonine Kinase |
| AUC | Area Under the Curve |
| BIRC3 | Baculoviral IAP Repeat Containing 3 |
| BMP | Bone Morphogenetic Protein |
| CA19-9 | Carbohydrate Antigen 19-9 |
| cAMP | 3′-5′-Cyclic Adenosine Monophosphate |
| CDI | Clostridium Difficile Infection |
| CEA | Carcino Embryonic Antigen |
| CEA | Carcinoembryonic Antigen |
| cf-DNA | Cell-Free DNA |
| CIMP | CpG Island Methylator Phenotype |
| CPSF4 | Cleavage And Polyadenylation Specific Factor 4 |
| CRC | Colorectal Cancer |
| CSC | Cancer Stem Cells |
| CT | Computed Tomography |
| CT | Chemotherapy |
| CTCs | Circulating Tumor Cells |
| ctDNA | Circulating Tumor DNA |
| ctDNA clear | ctDNA clearance or modification rate of every study arm and correlation with clinical outcome measures (according to the design of each study) |
| ctRNAs | Circulating Tumor RNAs |
| Cxcl2 | C-X-C Motif Chemokine Ligand 2 |
| DCE | Dynamic Contrast-Enhanced |
| ddPCR | Digital Droplet PCR |
| DDR | DNA Damage Response |
| DFS | Disease-free Survival |
| dPCR | Digital PCR |
| DR | Distant Recurrence |
| EGFR | Epidermal Growth Factor Receptor |
| EMT | Epithelial–Mesenchymal Transition |
| Enco/Bini/Cet | Encorafenib/Binimetinib/Cetuximab |
| Endp | Study Endpoints if not Reported in Study Description |
| ETBF | Enterotoxigenic Bacteroides Fragilis |
| EVs | Extracellular Vesicles |
| FMT | Fecal Microbiota Transplantation |
| Fn | Fusobacterium Nucleatum |
| FOBT | Fecal Occult Blood Test |
| FOXO | Forkhead Box O3 |
| GC | Gastric Cancer |
| GI | Gastrointestinal |
| Gli1 | Glioma-Associated Oncogene Homolog 1 |
| GLOBOCAN | Global Cancer Statistics |
| GMMs | Metabolites of the Intestinal Microbiome |
| HIF-1 | Hypoxia-Inducible Factor 1 |
| HR | Hazard Ratio |
| HR | High-Risk According to Histopathological Factors |
| HR | High-Risk |
| IAPs | Inhibitor of Apoptosis Proteins |
| ICIs | Immune Checkpoint Inhibitors |
| IECs | Ileal Intestinal Epithelial Cells |
| IL-17 | Interleukin-17 |
| Int | Interventional |
| IT | Immunotherapy |
| LB | Liquid Biopsy |
| LM | Luminal Microbiota |
| lncRNAs | Long non-coding RNAs |
| LR | Low-Risk According to Histopatological Factors |
| LR | Local Recurrence |
| MAM | Mucosal Associated Microbiota |
| mCRC | Metastatic CRC |
| MDM2 | Mouse Double Minute 2 |
| MEK | Mitogen-Activated Protein Kinase |
| MIR | MicroRNA |
| miRNAs | MicroRNAs |
| MO3UFA | Marine Omega-3 Polyunsaturated Fatty Acid |
| mRCC | Metastatic Renal Cell Carcinoma |
| MRD | Minimal Residual Disease |
| MRI | Magnetic Resonance Imaging |
| mSEPT9 | methylated Septin 9 |
| MSI | Micro-Satellite Instability |
| mTOR | Mammalian Target of Rapamycin |
| MYD88 | Myeloid Differentiation Primary Response 88 |
| N | Planned Enrollment |
| NA | Not Available |
| ncRNAs | non-coding RNAs |
| NEAT1 | Nuclear Enriched Abundant Transcript 1 |
| NFkB | Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells |
| NSCLC | Non-Small Cell Lung Cancer |
| NTBF | Non-Enterotoxigenic Bacteroides Fragilis |
| Obs | Observational |
| OMVs | Outer Membrane Vesicles |
| OS | Overall Survival |
| PACs | Proanthocyanidins |
| PC | Pancreatic Cancer |
| pCR | Pathologic Complete Response |
| PD-1 | Programmed Death-1 |
| PFS | Progression-Free Survival |
| PI3K | Phosphatidylinositol 3-Kinase |
| PLGC | Precancerous lesions of gastric carcinoma |
| PTEN | Phosphatase and Tensin Homolog |
| PUFAs | Omega-3 Polyunsatured Fatty Acids |
| PVAs | Hydroxy-Phenylvaleric Acids |
| PVLs | Hydroxy-Phenyl-γ-Valerolactones |
| QoL | Quality of Life |
| RECIST | Response Evaluation Criteria in Solid Tumors |
| RFS | Relapse-Free Survival |
| ROC | Receiver Operating Characteristic Curve |
| ROS | Reactive Oxygen Species |
| RR | Response Rate |
| SCFA | Short-Chain Fatty Acid |
| SMAD4 | Mothers Against Decapentaplegic Homolog 4 |
| SOC | Standard of Care |
| T | Clinical Trial |
| TDEs | Tumor-Derived Exosomes |
| TFH | Follicular T-helper Cell |
| TGF-β | Transforming Growth Factor Beta |
| TLR4 | Toll-like Receptor 4 |
| TNBC | Triple Negative Breast Cancer |
| TNF-α | Tumor Necrosis Factor- alpha |
| TNM | Classification of Malignant Tumors (Tumor-Nodes-Metastasis) |
| TNMB | Tumor-Nodes-Metastasis-Blood |
| TT | Treatment Toxicity/Treatment-Related Adverse Events |
| TTF | Time to Treatment Failure |
| TTR | Time to Recurrence |
| VEGF | Vascular Endothelial Growth Factor |
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, J.A.; Weinstein, J.N. Biomarkers in cancer staging, prognosis and treatment selection. Nat. Rev. Cancer 2005, 5, 845–856. [Google Scholar] [CrossRef] [PubMed]
- van Zijl, F.; Krupitza, G.; Mikulits, W. Initial steps of metastasis: Cell invasion and endothelial transmigration. Mutat. Res. 2011, 728, 23–34. [Google Scholar] [CrossRef]
- Corcoran, R.B.; Chabner, B.A. Application of Cell-free DNA Analysis to Cancer Treatment. N. Engl. J. Med. 2018, 379, 1754–1765. [Google Scholar] [CrossRef] [PubMed]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Gomez, A.; Espinoza, J.L.; Harkins, D.M.; Leong, P.; Saffery, R.; Bockmann, M.; Torralba, M.; Kuelbs, C.; Kodukula, R.; Inman, J.; et al. Host Genetic Control of the Oral Microbiome in Health and Disease. Cell Host Microbe 2017, 22, 269–278.e263. [Google Scholar] [CrossRef]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host lifestyle affects human microbiota on daily timescales. Genome Biol. 2014, 15, R89. [Google Scholar] [CrossRef]
- Graf, D.; Di Cagno, R.; Fak, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Crowley, E.; Di Nicolantonio, F.; Loupakis, F.; Bardelli, A. Liquid biopsy: Monitoring cancer-genetics in the blood. Nat. Rev. Clin. Oncol. 2013, 10, 472–484. [Google Scholar] [CrossRef]
- Lampis, A.; Hahne, J.C.; Hedayat, S.; Valeri, N. MicroRNAs as mediators of drug resistance mechanisms. Curr. Opin. Pharmacol. 2020, 54, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Osumi, H.; Shinozaki, E.; Yamaguchi, K.; Zembutsu, H. Clinical utility of circulating tumor DNA for colorectal cancer. Cancer Science 2019, 110, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Kilgour, E.; Rothwell, D.G.; Brady, G.; Dive, C. Liquid Biopsy-Based Biomarkers of Treatment Response and Resistance. Cancer Cell 2020, 37, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Hayashi, N.; Iguchi, T.; Ito, S.; Eguchi, H.; Mimori, K. Clinical and biological significance of circulating tumor cells in cancer. Mol. Oncol. 2016, 10, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Romiti, A.; Raffa, S.; Di Rocco, R.; Roberto, M.; Milano, A.; Zullo, A.; Leone, L.; Ranieri, D.; Mazzetta, F.; Medda, E.; et al. Circulating tumor cells count predicts survival in colorectal cancer patients. J. Gastrointest. Liver Dis. JGLD 2014, 23, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Marcuello, M.; Vymetalkova, V.; Neves, R.P.L.; Duran-Sanchon, S.; Vedeld, H.M.; Tham, E.; van Dalum, G.; Flugen, G.; Garcia-Barberan, V.; Fijneman, R.J.; et al. Circulating biomarkers for early detection and clinical management of colorectal cancer. Mol. As. Med. 2019, 69, 107–122. [Google Scholar] [CrossRef]
- Tsai, W.S.; You, J.F.; Hung, H.Y.; Hsieh, P.S.; Hsieh, B.; Lenz, H.J.; Idos, G.; Friedland, S.; Yi-Jiun Pan, J.; Shao, H.J.; et al. Novel Circulating Tumor Cell Assay for Detection of Colorectal Adenomas and Cancer. Clin. Transl. Gastroenterol. 2019, 10, e00088. [Google Scholar] [CrossRef]
- Tan, Y.; Wu, H. The significant prognostic value of circulating tumor cells in colorectal cancer: A systematic review and meta-analysis. Curr. Probl. Cancer 2018, 42, 95–106. [Google Scholar] [CrossRef]
- Tsai, W.S.; Chen, J.S.; Shao, H.J.; Wu, J.C.; Lai, J.M.; Lu, S.H.; Hung, T.F.; Chiu, Y.C.; You, J.F.; Hsieh, P.S.; et al. Circulating Tumor Cell Count Correlates with Colorectal Neoplasm Progression and Is a Prognostic Marker for Distant Metastasis in Non-Metastatic Patients. Sci. Rep. 2016, 6, 24517. [Google Scholar] [CrossRef]
- Cremolini, C.; Schirripa, M.; Antoniotti, C.; Moretto, R.; Salvatore, L.; Masi, G.; Falcone, A.; Loupakis, F. First-line chemotherapy for mCRC-a review and evidence-based algorithm. Nat. Rev. Clin. Oncol. 2015, 12, 607–619. [Google Scholar] [CrossRef]
- Cortellini, A.; Cannita, K.; Parisi, A.; Lanfiuti Baldi, P.; Venditti, O.; D’Orazio, C.; Dal Mas, A.; Calvisi, G.; Giordano, A.V.; Vicentini, V.; et al. Weekly alternate intensive regimen FIrB/FOx in metastatic colorectal cancer patients: An update from clinical practice. OncoTargets Ther. 2019, 12, 2159–2170. [Google Scholar] [CrossRef] [PubMed]
- Aranda, E.; Vieitez, J.M.; Gomez-Espana, A.; Gil Calle, S.; Salud-Salvia, A.; Grana, B.; Garcia-Alfonso, P.; Rivera, F.; Quintero-Aldana, G.A.; Reina-Zoilo, J.J.; et al. FOLFOXIRI plus bevacizumab versus FOLFOX plus bevacizumab for patients with metastatic colorectal cancer and >/=3 circulating tumour cells: The randomised phase III VISNU-1 trial. ESMO Open 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Nordlinger, B.; Cervantes, A.; Group, E.G.W. Advanced colorectal cancer: ESMO Clinical Practice Guidelines for treatment. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2010, 21 (Suppl. 5), v93–v97. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.; Cortellini, A.; Cannita, K.; Venditti, O.; Camarda, F.; Calegari, M.A.; Salvatore, L.; Tortora, G.; Rossini, D.; Germani, M.M.; et al. Evaluation of Second-line Anti-VEGF after First-line Anti-EGFR Based Therapy in RAS Wild-Type Metastatic Colorectal Cancer: The Multicenter “SLAVE” Study. Cancers 2020, 12, 1259. [Google Scholar] [CrossRef]
- Musella, V.; Pietrantonio, F.; Di Buduo, E.; Iacovelli, R.; Martinetti, A.; Sottotetti, E.; Bossi, I.; Maggi, C.; Di Bartolomeo, M.; de Braud, F.; et al. Circulating tumor cells as a longitudinal biomarker in patients with advanced chemorefractory, RAS-BRAF wild-type colorectal cancer receiving cetuximab or panitumumab. Int. J. Cancer 2015, 137, 1467–1474. [Google Scholar] [CrossRef]
- Cristiano, S.; Leal, A.; Phallen, J.; Fiksel, J.; Adleff, V.; Bruhm, D.C.; Jensen, S.O.; Medina, J.E.; Hruban, C.; White, J.R.; et al. Genome-wide cell-free DNA fragmentation in patients with cancer. Nature 2019, 570, 385–389. [Google Scholar] [CrossRef]
- Diehl, F.; Schmidt, K.; Choti, M.A.; Romans, K.; Goodman, S.; Li, M.; Thornton, K.; Agrawal, N.; Sokoll, L.; Szabo, S.A.; et al. Circulating mutant DNA to assess tumor dynamics. Nat. Med. 2008, 14, 985–990. [Google Scholar] [CrossRef]
- Wang, X.; Shi, X.Q.; Zeng, P.W.; Mo, F.M.; Chen, Z.H. Circulating cell free DNA as the diagnostic marker for colorectal cancer: A systematic review and meta-analysis. Oncotarget 2018, 9, 24514–24524. [Google Scholar] [CrossRef]
- Masuda, T.; Wang, X.; Maeda, M.; Canver, M.C.; Sher, F.; Funnell, A.P.; Fisher, C.; Suciu, M.; Martyn, G.E.; Norton, L.J.; et al. Transcription factors LRF and BCL11A independently repress expression of fetal hemoglobin. Science 2016, 351, 285–289. [Google Scholar] [CrossRef]
- Flamini, E.; Mercatali, L.; Nanni, O.; Calistri, D.; Nunziatini, R.; Zoli, W.; Rosetti, P.; Gardini, N.; Lattuneddu, A.; Verdecchia, G.M.; et al. Free DNA and carcinoembryonic antigen serum levels: An important combination for diagnosis of colorectal cancer. Clin. Cancer Res. 2006, 12, 6985–6988. [Google Scholar] [CrossRef]
- Nian, J.; Sun, X.; Ming, S.; Yan, C.; Ma, Y.; Feng, Y.; Yang, L.; Yu, M.; Zhang, G.; Wang, X. Diagnostic Accuracy of Methylated SEPT9 for Blood-based Colorectal Cancer Detection: A Systematic Review and Meta-Analysis. Clin. Transl. Gastroenterol. 2017, 8, e216. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhao, Q.; Wei, W.; Zheng, L.; Yi, S.; Li, G.; Wang, W.; Sheng, H.; Pu, H.; Mo, H.; et al. Circulating tumor DNA methylation profiles enable early diagnosis, prognosis prediction, and screening for colorectal cancer. Sci. Translat. Med. 2020, 12, eaax7533m. [Google Scholar] [CrossRef] [PubMed]
- Spindler, K.G.; Boysen, A.K.; Pallisgard, N.; Johansen, J.S.; Tabernero, J.; Sorensen, M.M.; Jensen, B.V.; Hansen, T.F.; Sefrioui, D.; Andersen, R.F.; et al. Cell-Free DNA in Metastatic Colorectal Cancer: A Systematic Review and Meta-Analysis. Oncologist 2017, 22, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Perdyan, A.; Spychalski, P.; Kacperczyk, J.; Rostkowska, O.; Kobiela, J. Circulating Tumor DNA in KRAS positive colorectal cancer patients as a prognostic factor—A systematic review and meta-analysis. Crit. Rev. Oncol. Hematol. 2020, 154, 103065. [Google Scholar] [CrossRef]
- Douillard, J.Y.; Oliner, K.S.; Siena, S.; Tabernero, J.; Burkes, R.; Barugel, M.; Humblet, Y.; Bodoky, G.; Cunningham, D.; Jassem, J.; et al. Panitumumab-FOLFOX4 treatment and RAS mutations in colorectal cancer. N. Engl. J. Med. 2013, 369, 1023–1034. [Google Scholar] [CrossRef]
- Di Nicolantonio, F.; Martini, M.; Molinari, F.; Sartore-Bianchi, A.; Arena, S.; Saletti, P.; De Dosso, S.; Mazzucchelli, L.; Frattini, M.; Siena, S.; et al. Wild-type BRAF is required for response to panitumumab or cetuximab in metastatic colorectal cancer. J. Clin. Oncol. 2008, 26, 5705–5712. [Google Scholar] [CrossRef]
- Siravegna, G.; Mussolin, B.; Buscarino, M.; Corti, G.; Cassingena, A.; Crisafulli, G.; Ponzetti, A.; Cremolini, C.; Amatu, A.; Lauricella, C.; et al. Clonal evolution and resistance to EGFR blockade in the blood of colorectal cancer patients. Nat. Med. 2015, 21, 827. [Google Scholar] [CrossRef]
- Khan, K.H.; Cunningham, D.; Werner, B.; Vlachogiannis, G.; Spiteri, I.; Heide, T.; Mateos, J.F.; Vatsiou, A.; Lampis, A.; Damavandi, M.D.; et al. Longitudinal Liquid Biopsy and Mathematical Modeling of Clonal Evolution Forecast Time to Treatment Failure in the PROSPECT-C Phase II Colorectal Cancer Clinical Trial. Cancer Dis. 2018, 8, 1270–1285. [Google Scholar] [CrossRef]
- Khan, K.; Rata, M.; Cunningham, D.; Koh, D.M.; Tunariu, N.; Hahne, J.C.; Vlachogiannis, G.; Hedayat, S.; Marchetti, S.; Lampis, A.; et al. Functional imaging and circulating biomarkers of response to regorafenib in treatment-refractory metastatic colorectal cancer patients in a prospective phase II study. Gut 2018, 67, 1484–1492. [Google Scholar] [CrossRef]
- Yang, Y.C.; Wang, D.; Jin, L.; Yao, H.W.; Zhang, J.H.; Wang, J.; Zhao, X.M.; Shen, C.Y.; Chen, W.; Wang, X.L.; et al. Circulating tumor DNA detectable in early- and late-stage colorectal cancer patients. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Yang, M.; Forbes, M.E.; Bitting, R.L.; O’Neill, S.S.; Chou, P.C.; Topaloglu, U.; Miller, L.D.; Hawkins, G.A.; Grant, S.C.; DeYoung, B.R.; et al. Incorporating blood-based liquid biopsy information into cancer staging: Time for a TNMB system? Ann. Oncol. 2018, 29, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Schrag, D.; Rifas-Shiman, S.; Saltz, L.; Bach, P.B.; Begg, C.B. Adjuvant chemotherapy use for Medicare beneficiaries with stage II colon cancer. J. Clin. Oncol. 2002, 20, 3999–4005. [Google Scholar] [CrossRef] [PubMed]
- Tie, J.; Wang, Y.; Tomasetti, C.; Li, L.; Springer, S.; Kinde, I.; Silliman, N.; Tacey, M.; Wong, H.L.; Christie, M.; et al. Circulating tumor DNA analysis detects minimal residual disease and predicts recurrence in patients with stage II colon cancer. Sci. Transl. Med. 2016, 8, 346ra392. [Google Scholar] [CrossRef] [PubMed]
- Reinert, T.; Henriksen, T.V.; Christensen, E.; Sharma, S.; Salari, R.; Sethi, H.; Knudsen, M.; Nordentoft, I.; Wu, H.T.; Tin, A.S.; et al. Analysis of Plasma Cell-Free DNA by Ultradeep Sequencing in Patients With Stages I to III Colorectal Cancer. JAMA Oncol. 2019, 5, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Cohen, J.D.; Kinde, I.; Ptak, J.; Popoli, M.; Schaefer, J.; Silliman, N.; Dobbyn, L.; Tie, J.; et al. Prognostic Potential of Circulating Tumor DNA Measurement in Postoperative Surveillance of Nonmetastatic Colorectal Cancer. JAMA Oncol. 2019, 5, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.J.; Wang, W.; Hann, S.S. Interactions among lncRNAs, miRNAs and mRNA in colorectal cancer. Biochimie 2019, 163, 58–72. [Google Scholar] [CrossRef]
- Qi, F.F.; Yang, Y.; Zhang, H.; Chen, H. Long non-coding RNAs: Key regulators in oxaliplatin resistance of colorectal cancer. Biomed. Pharmacother. 2020, 128, 110329. [Google Scholar] [CrossRef]
- Wei, L.; Wang, X.; Lv, L.; Liu, J.; Xing, H.; Song, Y.; Xie, M.; Lei, T.; Zhang, N.; Yang, M. The emerging role of microRNAs and long noncoding RNAs in drug resistance of hepatocellular carcinoma. Mol. Cancer 2019, 18, 147. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, G.; Ren, W.; Wang, B.; Yang, C.; Li, M. LncRNA NEAT1 Regulates 5-Fu Sensitivity, Apoptosis and Invasion in Colorectal Cancer Through the MiR-150-5p/CPSF4 Axis. OncoTargets Ther. 2020, 13, 6373–6383. [Google Scholar] [CrossRef]
- Xian, Z.; Hu, B.; Wang, T.; Zeng, J.; Cai, J.; Zou, Q.; Zhu, P. lncRNA UCA1 Contributes to 5-Fluorouracil Resistance of Colorectal Cancer Cells Through miR-23b-3p/ZNF281 Axis. OncoTargets Ther. 2020, 13, 7571–7583. [Google Scholar] [CrossRef]
- He, Z.; Dang, J.; Song, A.; Cui, X.; Ma, Z.; Zhang, Y. The involvement of miR-150/beta-catenin axis in colorectal cancer progression. Biomed. Pharmacother. 2020, 121, 109495. [Google Scholar] [CrossRef] [PubMed]
- Sur, D.; Burz, C.; Sabarimurugan, S.; Irimie, A. Diagnostic and Prognostic Significance of MiR-150 in Colorectal Cancer: A Systematic Review and Meta-Analysis. J. Pers. Med. 2020, 10, 99. [Google Scholar] [CrossRef]
- Peng, Q.; Yao, W.; Yu, C.; Zou, L.; Shen, Y.; Zhu, Y.; Cheng, M.; Feng, Z.; Xu, B. Identification of microRNA-181 as a promising biomarker for predicting the poor survival in colorectal cancer. Cancer Med. 2019, 8, 5995–6009. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ke, F.; Chen, T.; Zhou, Q.; Weng, L.; Tan, J.; Shen, W.; Li, L.; Zhou, J.; Xu, C.; et al. MicroRNAs that regulate PTEN as potential biomarkers in colorectal cancer: A systematic review. J. Cancer Res. Clin. Oncol. 2020, 146, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Sabarimurugan, S.; Madhav, M.R.; Kumarasamy, C.; Gupta, A.; Baxi, S.; Krishnan, S.; Jayaraj, R. Prognostic Value of MicroRNAs in Stage II Colorectal Cancer Patients: A Systematic Review and Meta-Analysis. Mol. Diagn. Ther. 2020, 24, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Horimatsu, T.; Okugawa, Y.; Nishida, N.; Honjo, H.; Ida, H.; Kou, T.; Kusaka, T.; Sasaki, Y.; Yagi, M.; et al. Serum miR-21, miR-29a, and miR-125b Are Promising Biomarkers for the Early Detection of Colorectal Neoplasia. Clin. Cancer Res. 2015, 21, 4234–4242. [Google Scholar] [CrossRef]
- Hur, K.; Toiyama, Y.; Okugawa, Y.; Ide, S.; Imaoka, H.; Boland, C.R.; Goel, A. Circulating microRNA-203 predicts prognosis and metastasis in human colorectal cancer. Gut 2017, 66, 654–665. [Google Scholar] [CrossRef]
- Vychytilova-Faltejskova, P.; Radova, L.; Sachlova, M.; Kosarova, Z.; Slaba, K.; Fabian, P.; Grolich, T.; Prochazka, V.; Kala, Z.; Svoboda, M.; et al. Serum-based microRNA signatures in early diagnosis and prognosis prediction of colon cancer. Carcinogenesis 2016, 37, 941–950. [Google Scholar] [CrossRef]
- Oehme, F.; Krahl, S.; Gyorffy, B.; Muessle, B.; Rao, V.; Greif, H.; Ziegler, N.; Lin, K.; Thepkaysone, M.L.; Polster, H.; et al. Low level of exosomal long non-coding RNA HOTTIP is a prognostic biomarker in colorectal cancer. RNA Biol. 2019, 16, 1339–1345. [Google Scholar] [CrossRef]
- Kang, W.; Zheng, Q.; Lei, J.; Chen, C.; Yu, C. Prognostic Value of Long Noncoding RNAs in Patients with Gastrointestinal Cancer: A Systematic Review and Meta-Analysis. Dis. Markers 2018, 2018, 5340894. [Google Scholar] [CrossRef]
- Madurantakam Royam, M.; Kumarasamy, C.; Baxi, S.; Gupta, A.; Ramesh, N.; Kodiveri Muthukaliannan, G.; Jayaraj, R. Current Evidence on miRNAs as Potential Theranostic Markers for Detecting Chemoresistance in Colorectal Cancer: A Systematic Review and Meta-Analysis of Preclinical and Clinical Studies. Mol. Diagn. Ther. 2019, 23, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H. Problems with the concept of gut microbiota dysbiosis. Microb. Biotechnol. 2020, 13, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the human microbiome. Nutr. Rev. 2012, 70 (Suppl. 1), S38–S44. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Ravel, J. The vocabulary of microbiome research: A proposal. Microbiome 2015, 3, 31. [Google Scholar] [CrossRef]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- O’Keefe, S.J. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Tilg, H.; Adolph, T.E.; Gerner, R.R.; Moschen, A.R. The Intestinal Microbiota in Colorectal Cancer. Cancer Cell 2018, 33, 954–964. [Google Scholar] [CrossRef]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef]
- Gagniere, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef]
- Sanchez-Alcoholado, L.; Ramos-Molina, B.; Otero, A.; Laborda-Illanes, A.; Ordonez, R.; Medina, J.A.; Gomez-Millan, J.; Queipo-Ortuno, M.I. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers 2020, 12, 1406. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, A.P.; Redinbo, M.R.; Bultman, S.J. The role of the microbiome in cancer development and therapy. CA A Cancer J. Clin. 2017, 67, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, A.; Nadel, B.; Raoult, D.; Neefjes, J.; Gorvel, J.P. Collateral damage: Insights into bacterial mechanisms that predispose host cells to cancer. Nat. Rev. Microbiol. 2017, 15, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Fahrer, J.; Huelsenbeck, J.; Jaurich, H.; Dorsam, B.; Frisan, T.; Eich, M.; Roos, W.P.; Kaina, B.; Fritz, G. Cytolethal distending toxin (CDT) is a radiomimetic agent and induces persistent levels of DNA double-strand breaks in human fibroblasts. DNA Repair 2014, 18, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Dalmasso, G.; Cougnoux, A.; Delmas, J.; Darfeuille-Michaud, A.; Bonnet, R. The bacterial genotoxin colibactin promotes colon tumor growth by modifying the tumor microenvironment. Gut Microbes 2014, 5, 675–680. [Google Scholar] [CrossRef]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational signature in colorectal cancer caused by genotoxic pks(+) E. coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef]
- Zamani, S.; Taslimi, R.; Sarabi, A.; Jasemi, S.; Sechi, L.A.; Feizabadi, M.M. Enterotoxigenic Bacteroides fragilis: A Possible Etiological Candidate for Bacterially-Induced Colorectal Precancerous and Cancerous Lesions. Front. Cell. Infect. Microbiol. 2019, 9, 449. [Google Scholar] [CrossRef]
- Brennan, C.A.; Garrett, W.S. Fusobacterium nucleatum—Symbiont, opportunist and oncobacterium. Nat. Rev. Microbiol. 2019, 17, 156–166. [Google Scholar] [CrossRef]
- Sun, C.H.; Li, B.B.; Wang, B.; Zhao, J.; Zhang, X.Y.; Li, T.T.; Li, W.B.; Tang, D.; Qiu, M.J.; Wang, X.C.; et al. The role of Fusobacterium nucleatum in colorectal cancer: From carcinogenesis to clinical management. Chronic Dis. Transl. Med. 2019, 5, 178–187. [Google Scholar] [CrossRef]
- Koi, M.; Okita, Y.; Carethers, J.M. Fusobacterium nucleatum Infection in Colorectal Cancer: Linking Inflammation, DNA Mismatch Repair and Genetic and Epigenetic Alterations. J. Anus Rectum Colon 2018, 2, 37–46. [Google Scholar] [CrossRef]
- Zitvogel, L.; Daillere, R.; Roberti, M.P.; Routy, B.; Kroemer, G. Anticancer effects of the microbiome and its products. Nat. Rev. Microbiol. 2017, 15, 465–478. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma-carcinoma sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Lee, Y.J.; Yoo, H.J.; Kim, M.; Chang, Y.; Lee, D.S.; Lee, J.H. Consumption of Dairy Yogurt Containing Lactobacillus paracasei ssp. paracasei, Bifidobacterium animalis ssp. lactis and Heat-Treated Lactobacillus plantarum Improves Immune Function Including Natural Killer Cell Activity. Nutrients 2017, 9, 558. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Yang, Y.; Shen, C.; Wang, Y.; Deng, A.; Qin, Q.; Qiao, L. Metaproteomics characterizes human gut microbiome function in colorectal cancer. NPJ Biofilms Microbiomes 2020, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yu, Y.; Wang, Y.Z.; Wang, J.J.; Guan, R.; Sun, Y.; Shi, F.; Gao, J.; Fu, X.L. Role of SCFAs in gut microbiome and glycolysis for colorectal cancer therapy. J. Cell. Physiol. 2019, 234, 17023–17049. [Google Scholar] [CrossRef]
- Borges-Canha, M.; Portela-Cidade, J.P.; Dinis-Ribeiro, M.; Leite-Moreira, A.F.; Pimentel-Nunes, P. Role of colonic microbiota in colorectal carcinogenesis: A systematic review. Rev. Esp. de Enferm. Dig. 2015, 107, 659–671. [Google Scholar] [CrossRef]
- Grazioso, T.P.; Brandt, M.; Djouder, N. Diet, Microbiota, and Colorectal Cancer. iScience 2019, 21, 168–187. [Google Scholar] [CrossRef]
- Wan, M.L.; Wang, Y.; Zeng, Z.; Deng, B.; Zhu, B.S.; Cao, T.; Li, Y.K.; Xiao, J.; Han, Q.; Wu, Q. Colorectal cancer (CRC) as a multifactorial disease and its causal correlations with multiple signaling pathways. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Duchartre, Y.; Kim, Y.M.; Kahn, M. The Wnt signaling pathway in cancer. Crit. Rev. Oncol. Hematol. 2016, 99, 141–149. [Google Scholar] [CrossRef]
- Zhang, L.; Shay, J.W. Multiple Roles of APC and its Therapeutic Implications in Colorectal Cancer. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Tian, T.; Kalland, K.H.; Ke, X.; Qu, Y. Targeting Wnt/beta-Catenin Signaling for Cancer Immunotherapy. Trends Pharmacol. Sci. 2018, 39, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Koni, M.; Pinnaro, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int. J. Mol. Sci. 2020, 21, 7697. [Google Scholar] [CrossRef] [PubMed]
- Hofman, P.; Vouret-Craviari, V. Microbes-induced EMT at the crossroad of inflammation and cancer. Gut Microbes 2012, 3, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Fearon, E.R.; Hamilton, S.R.; Kern, S.E.; Preisinger, A.C.; Leppert, M.; Nakamura, Y.; White, R.; Smits, A.M.; Bos, J.L. Genetic alterations during colorectal-tumor development. N. England J. Med. 1988, 319, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wang, L.; Yin, R.; Hudlikar, R.; Li, S.; Kuo, H.D.; Peter, R.; Sargsyan, D.; Guo, Y.; Liu, X.; et al. Epigenetics/epigenomics and prevention by curcumin of early stages of inflammatory-driven colon cancer. Mol. Carcinog. 2020, 59, 227–236. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, S.; Li, H.; Yang, F.; Mushtaq, N.; Ullah, S.; Shi, Y.; An, C.; Xu, J. The influence of gut microbiota dysbiosis to the efficacy of 5-Fluorouracil treatment on colorectal cancer. Biomed. Pharmacother. 2018, 108, 184–193. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Weng, W.; Guo, B.; Cai, G.; Ma, Y.; Cai, S. Fusobacterium nucleatum promotes chemoresistance to 5-fluorouracil by upregulation of BIRC3 expression in colorectal cancer. J. Exp. Clin. Cancer Res. CR 2019, 38, 14. [Google Scholar] [CrossRef]
- Andre, T.; Shiu, K.K.; Kim, T.W.; Jensen, B.V.; Jensen, L.H.; Punt, C.; Smith, D.; Garcia-Carbonero, R.; Benavides, M.; Gibbs, P.; et al. Pembrolizumab in Microsatellite-Instability-High Advanced Colorectal Cancer. N. Engl. J. Med. 2020, 383, 2207–2218. [Google Scholar] [CrossRef]
- Lang, M.; Baumgartner, M.; Rozalska, A.; Frick, A.; Riva, A.; Jarek, M.; Berry, D.; Gasche, C. Crypt residing bacteria and proximal colonic carcinogenesis in a mouse model of Lynch syndrome. Int. J. Cancer 2020, 147, 2316–2326. [Google Scholar] [CrossRef]
- Lalani, A.A.; Xie, W.; Braun, D.A.; Kaymakcalan, M.; Bosse, D.; Steinharter, J.A.; Martini, D.J.; Simantov, R.; Lin, X.; Wei, X.X.; et al. Effect of Antibiotic Use on Outcomes with Systemic Therapies in Metastatic Renal Cell Carcinoma. Eur. Urol. Oncol. 2020, 3, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Ngara, T.R.; Zhang, H. Recent Advances in Function-based Metagenomic Screening. Genom. Proteom. Bioinform. 2018, 16, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Coker, O.O.; Nakatsu, G.; Wu, W.K.K.; Zhao, L.; Chen, Z.; Chan, F.K.L.; Kristiansen, K.; Sung, J.J.Y.; Wong, S.H.; et al. Multi-cohort analysis of colorectal cancer metagenome identified altered bacteria across populations and universal bacterial markers. Microbiome 2018, 6, 70. [Google Scholar] [CrossRef] [PubMed]
- Dalal, N.; Jalandra, R.; Sharma, M.; Prakash, H.; Makharia, G.K.; Solanki, P.R.; Singh, R.; Kumar, A. Omics technologies for improved diagnosis and treatment of colorectal cancer: Technical advancement and major perspectives. Biomed. Pharmacother. 2020, 131, 110648. [Google Scholar] [CrossRef]
- Behrouzi, A.; Nafari, A.H.; Siadat, S.D. The significance of microbiome in personalized medicine. Clin. Transl. Med. 2019, 8, 16. [Google Scholar] [CrossRef]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968–976. [Google Scholar] [CrossRef]
- Gao, R.; Kong, C.; Huang, L.; Li, H.; Qu, X.; Liu, Z.; Lan, P.; Wang, J.; Qin, H. Mucosa-associated microbiota signature in colorectal cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2073–2083. [Google Scholar] [CrossRef]
- Zeller, G.; Tap, J.; Voigt, A.Y.; Sunagawa, S.; Kultima, J.R.; Costea, P.I.; Amiot, A.; Bohm, J.; Brunetti, F.; Habermann, N.; et al. Potential of fecal microbiota for early-stage detection of colorectal cancer. Mol. Syst. Biol. 2014, 10, 766. [Google Scholar] [CrossRef]
- Kim, D.J.; Yang, J.; Seo, H.; Lee, W.H.; Ho Lee, D.; Kym, S.; Park, Y.S.; Kim, J.G.; Jang, I.J.; Kim, Y.K.; et al. Colorectal cancer diagnostic model utilizing metagenomic and metabolomic data of stool microbial extracellular vesicles. Sci. Rep. 2020, 10, 2860. [Google Scholar] [CrossRef]
- Ahmadi Badi, S.; Moshiri, A.; Fateh, A.; Rahimi Jamnani, F.; Sarshar, M.; Vaziri, F.; Siadat, S.D. Microbiota-Derived Extracellular Vesicles as New Systemic Regulators. Front. Microbiol. 2017, 8, 1610. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial driver-passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Oke, S.; Martin, A. Insights into the role of the intestinal microbiota in colon cancer. Ther. Adv. Gastroenterol. 2017, 10, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Farin, H.F.; Karthaus, W.R.; Kujala, P.; Rakhshandehroo, M.; Schwank, G.; Vries, R.G.; Kalkhoven, E.; Nieuwenhuis, E.E.; Clevers, H. Paneth cell extrusion and release of antimicrobial products is directly controlled by immune cell-derived IFN-gamma. J. Exp. Med. 2014, 211, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.; Van Es, J.H.; Van den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D.; et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef]
- Rubert, J.; Schweiger, P.J.; Mattivi, F.; Tuohy, K.; Jensen, K.B.; Lunardi, A. Intestinal Organoids: A Tool for Modelling Diet-Microbiome-Host Interactions. Trends Endocrinol. Metab. TEM 2020, 31, 848–858. [Google Scholar] [CrossRef]
- Toden, S.; Ravindranathan, P.; Gu, J.; Cardenas, J.; Yuchang, M.; Goel, A. Oligomeric proanthocyanidins (OPCs) target cancer stem-like cells and suppress tumor organoid formation in colorectal cancer. Sci. Rep. 2018, 8, 3335. [Google Scholar] [CrossRef]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Zhang, L.S.; Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 2016, 8, 46. [Google Scholar] [CrossRef]
- Trost, K.; Ulaszewska, M.M.; Stanstrup, J.; Albanese, D.; De Filippo, C.; Tuohy, K.M.; Natella, F.; Scaccini, C.; Mattivi, F. Host: Microbiome co-metabolic processing of dietary polyphenols—An acute, single blinded, cross-over study with different doses of apple polyphenols in healthy subjects. Food Res. Int. 2018, 112, 108–128. [Google Scholar] [CrossRef]
- Mena, P.; Bresciani, L.; Brindani, N.; Ludwig, I.A.; Pereira-Caro, G.; Angelino, D.; Llorach, R.; Calani, L.; Brighenti, F.; Clifford, M.N.; et al. Phenyl-gamma-valerolactones and phenylvaleric acids, the main colonic metabolites of flavan-3-ols: Synthesis, analysis, bioavailability, and bioactivity. Nat. Prod. Rep. 2019, 36, 714–752. [Google Scholar] [CrossRef]
- Mehta, R.S.; Nishihara, R.; Cao, Y.; Song, M.; Mima, K.; Qian, Z.R.; Nowak, J.A.; Kosumi, K.; Hamada, T.; Masugi, Y.; et al. Association of Dietary Patterns With Risk of Colorectal Cancer Subtypes Classified by Fusobacterium nucleatum in Tumor Tissue. JAMA Oncol. 2017, 3, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Nencioni, A.; Caffa, I.; Cortellino, S.; Longo, V.D. Fasting and cancer: Molecular mechanisms and clinical application. Nat. Rev. Cancer 2018, 18, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, C.; Lu, S.; Nichols, R.G.; Tian, Y.; Li, L.; Patel, D.; Ma, Y.; Brocker, C.N.; Yan, T.; et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell Metabo. 2017, 26, 672–685.e674. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Tonouchi, H.; Sasayama, A.; Ashida, K. A Ketogenic Formula Prevents Tumor Progression and Cancer Cachexia by Attenuating Systemic Inflammation in Colon 26 Tumor-Bearing Mice. Nutrients 2018, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.C.; Seira Oriach, C.; Murphy, K.; Moloney, G.M.; Cryan, J.F.; Dinan, T.G.; Paul Ross, R.; Stanton, C. Omega-3 polyunsaturated fatty acids critically regulate behaviour and gut microbiota development in adolescence and adulthood. Brain Behav. Immun. 2017, 59, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillere, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef]
- Buti, S.; Bersanelli, M.; Perrone, F.; Tiseo, M.; Tucci, M.; Adamo, V.; Stucci, L.S.; Russo, A.; Tanda, E.T.; Spagnolo, F.; et al. Effect of concomitant medications with immune-modulatory properties on the outcomes of patients with advanced cancer treated with immune checkpoint inhibitors: Development and validation of a novel prognostic index. Eur. J. Cancer 2021, 142, 18–28. [Google Scholar] [CrossRef]
- Sanchez, B.; Delgado, S.; Blanco-Miguez, A.; Lourenco, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef]
- Chen, D.; Wu, J.; Jin, D.; Wang, B.; Cao, H. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int. J. Cancer 2019, 145, 2021–2031. [Google Scholar] [CrossRef]
- Wang, Y.; Wiesnoski, D.H.; Helmink, B.A.; Gopalakrishnan, V.; Choi, K.; DuPont, H.L.; Jiang, Z.D.; Abu-Sbeih, H.; Sanchez, C.A.; Chang, C.C.; et al. Fecal microbiota transplantation for refractory immune checkpoint inhibitor-associated colitis. Nat. Med. 2018, 24, 1804–1808. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.W.; Lee, H.C.; Li, L.H.; Chiang Chiau, J.S.; Wang, T.E.; Chuang, W.H.; Chen, M.J.; Wang, H.Y.; Shih, S.C.; Liu, C.Y.; et al. Fecal Microbiota Transplantation Prevents Intestinal Injury, Upregulation of Toll-Like Receptors, and 5-Fluorouracil/Oxaliplatin-Induced Toxicity in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 386. [Google Scholar] [CrossRef] [PubMed]
- Le Bastard, Q.; Ward, T.; Sidiropoulos, D.; Hillmann, B.M.; Chun, C.L.; Sadowsky, M.J.; Knights, D.; Montassier, E. Fecal microbiota transplantation reverses antibiotic and chemotherapy-induced gut dysbiosis in mice. Sci. Rep. 2018, 8, 6219. [Google Scholar] [CrossRef]
- Hefazi, M.; Patnaik, M.M.; Hogan, W.J.; Litzow, M.R.; Pardi, D.S.; Khanna, S. Safety and Efficacy of Fecal Microbiota Transplant for Recurrent Clostridium difficile Infection in Patients With Cancer Treated With Cytotoxic Chemotherapy: A Single-Institution Retrospective Case Series. Mayo Clin. Proc. 2017, 92, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Kopetz, S.; Grothey, A.; Yaeger, R.; Van Cutsem, E.; Desai, J.; Yoshino, T.; Wasan, H.; Ciardiello, F.; Loupakis, F.; Hong, Y.S.; et al. Encorafenib, Binimetinib, and Cetuximab in BRAF V600E-Mutated Colorectal Cancer. N. Engl. J. Med. 2019, 381, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Kolencik, D.; Shishido, S.N.; Pitule, P.; Mason, J.; Hicks, J.; Kuhn, P. Liquid Biopsy in Colorectal Carcinoma: Clinical Applications and Challenges. Cancers 2020, 12, 1376. [Google Scholar] [CrossRef]
- Castro-Giner, F.; Gkountela, S.; Donato, C.; Alborelli, I.; Quagliata, L.; Ng, C.K.Y.; Piscuoglio, S.; Aceto, N. Cancer Diagnosis Using a Liquid Biopsy: Challenges and Expectations. Diagnostics 2018, 8, 31. [Google Scholar] [CrossRef]
- Blaser, M.J. Antibiotic use and its consequences for the normal microbiome. Science 2016, 352, 544–545. [Google Scholar] [CrossRef]
- Nagrath, S.; Sequist, L.V.; Maheswaran, S.; Bell, D.W.; Irimia, D.; Ulkus, L.; Smith, M.R.; Kwak, E.L.; Digumarthy, S.; Muzikansky, A.; et al. Isolation of rare circulating tumour cells in cancer patients by microchip technology. Nature 2007, 450, 1235–1239. [Google Scholar] [CrossRef]
- Mishra, A.; Dubash, T.D.; Edd, J.F.; Jewett, M.K.; Garre, S.G.; Karabacak, N.M.; Rabe, D.C.; Mutlu, B.R.; Walsh, J.R.; Kapur, R.; et al. Ultrahigh-throughput magnetic sorting of large blood volumes for epitope-agnostic isolation of circulating tumor cells. Proc. Natl. Acad. Sci. USA 2020, 117, 16839–16847. [Google Scholar] [CrossRef] [PubMed]
- Kalinich, M.; Bhan, I.; Kwan, T.T.; Miyamoto, D.T.; Javaid, S.; LiCausi, J.A.; Milner, J.D.; Hong, X.; Goyal, L.; Sil, S.; et al. An RNA-based signature enables high specificity detection of circulating tumor cells in hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 2017, 114, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Bardia, A.; Aceto, N.; Bersani, F.; Madden, M.W.; Donaldson, M.C.; Desai, R.; Zhu, H.; Comaills, V.; Zheng, Z.; et al. Cancer therapy. Ex vivo culture of circulating breast tumor cells for individualized testing of drug susceptibility. Science 2014, 345, 216–220. [Google Scholar] [CrossRef]
- Fachin, F.; Spuhler, P.; Martel-Foley, J.M.; Edd, J.F.; Barber, T.A.; Walsh, J.; Karabacak, M.; Pai, V.; Yu, M.; Smith, K.; et al. Monolithic Chip for High-throughput Blood Cell Depletion to Sort Rare Circulating Tumor Cells. Sci. Rep. 2017, 7, 10936. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Brief Study Title | NCT Number/Study Name | Country Study Period | Stage | N | Study Type/Phase/Endp | Target/Assay-Test (If Available) | Study Overview/Schematic Description |
|---|---|---|---|---|---|---|---|
| ctDNA Analysis Informing ACT in Stage III CRC | ACTRN 12617001566325 (DYNAMIC-III) | Australia 2017–2024 | III | 1000 | Int rand/2–3/DFS, OS, ctDNA clear | ctDNA | Surgery followed by ctDNA detection and clinician’s choice ACT (no ACT > fluoropyrimidine > XELOX/FOLFOX) followed by randomization to: (1) SOC arm: SOC ACT (ctDNA blinded) or (2) EXP arm ctDNA+: escalated ACT (FOLFOXIRI allowed) or (3) EXP arm ctDNA-: de-escalated ACT |
| ctDNA Analysis Informing ACT in Stage II CRC | ACTRN 12615000381583 (DYNAMIC II) | Australia 2015–2024 | II | 450 | Int rand/2/DFS, OS, ctDNA clear | ctDNA | Surgery followed by ctDNA detection and randomization to: (1) SOC arm: SOC, (2) EXP arm ctDNA+: ACT (fluoropyrimidine or XELOX/FOLFOX according to clinician’s choice), (3) EXP arm ctDNA-: follow-up |
| Tracking mutations in ctDNA to predict relapse in early CRC | NCT04050345 (TRACC) | UK 2016–2024 | HR II-III | 1000 | Int rand/3/DFS, OS, QoL | ctDNA/RM NGS | Surgery followed by randomization to: (1) SOC arm: SOC ACT, (2) EXP arm: ctDNA guided de-escalated ACT—If ctDNA-: CAPOX × 3 months is reduced to cape x 6 months and CAPE × 6 months is reduced to observation—After 3 months if ctDNA+: switch to CAPOX |
| ctDNA Based Decision for ACT in CRC Stage II Evaluation | NCT04089631 (CIRCULATE) | Germany Austria Switzerland 2019–2026 | II | 4812 | Int rand/3/DFS, OS, TT ctDNA clear | ctDNA/Dresden NGS | After radical surgery, ctDNA+ are randomized to: (1) CAPE × 6 months (or CAPOX × 3–6 months, investigator choice) or (2) follow-up and CtDNA- are randomized to: (1) follow-up inside the study or (2) follow-up outside the study (off-study) |
| Decision for ACT in stage II CRC based on ctDNA | NCT04120701 (CIRCULATE-PRODIGE 70) | France 2019–2026 | II | 1980 | Int rand/3/DFS, TTR, OS, ctDNA clear | ctDNA/ddPCR of 2 methylated probes | After radical surgery, ctDNA+ are randomized to: (1) ACT (FOLFOX) or (2) surveillance inside the trial. CtDNA- are randomized to (1) surveillance inside the trial or (2) surveillance outside the trial |
| ctDNA guided ACT in stage II CRC according the trials within cohorts design | Planning NTR6455 (MEDOCC CrEATE) | Netherland 2018–2022 | LR II | 1320 | Int rand/3/DFS, OS, QoL | ctDNA/gene panel (PG Dx elio platform) | Surgery followed by randomization to: (1) EXP arm: if ctDNA+ FOLFOX/CAPOX × 6 months or follow-up (patient choice), (2) EXP arm: if ctDNA- follow-up, (3) SOC arm: follow up (ctDNA tested later) |
| Intervention Trial Implementing Non-invasive ctDNA Analysis to Optimize the Operative and Postoperative Treatment for CRC Patients | NCT03748680 (IMPROVE-IT) | Denmark 2018–2025 | I-II | 64 | Int rand/2/DFS, LR, TTR, OS, ctDNA clear | ctDNA/NGS + ddPCR | After radical surgery, ctDNA+ patients with no indication to ACT according to DCCG guidelines are randomized to: (1) SOC arm: intensified follow-up or (2) EXP arm: CAPOX or FOLFOX followed by intensified follow-up |
| ctDNA Testing in Predicting Treatment for Patients With Stage IIA CRC After Surgery | NCT04068103 (COBRA) | USA Canada 2019–2024 | IIA | 1408 | Int rand/2–3/ctDNA clear, RFS, OS, TTR | ctDNA | After radical surgery, patients are randomized to: (1) SOC arm: active surveillance (blood stored and tested for ctDNA later), (2) EXP arm ctDNA+: FOLFOX or XELOX × 6 months, (3) EXP arm ctDNA-: active surveillance |
| Early identification and treatment of occult metastatic disease in stage III CRC | NCT03803553 | USA 2020–2023 | III | 500 | Int/3/DFS, OS, ctDNA clear | ctDNA | Surgery followed by SOC ACT followed by ctDNA assessment for MRD: (1) ctDNA+ MSI-h: Nivolumab, (2) ctDNA+ BRAF V600E: Enco/Bini/Cet × 6 months, (3) ctDNA+ others: randomization to FOLFIRI × 6 months or SOC observation, (4) ctDNA-: SOC observation |
| Post-surgical Liquid Biopsy-guided Treatment of Stage III and HR Stage II CRC Patients | NCT04259944 (PEGASUS) | Italy Spain 2020–2023 | HR II- III | 140 | Int/2/DFS, OS, TT, QoL, ctDNA clear | ctDNA/LUNAR1 test | After surgical surgery: (1) ctDNA+: CAPOX × 3 months, (2) ctDNA-: CAPE × 6 months and early switch to CAPOX if ctDNA+ after first cycleAfter ACT: (3) ctDNA+/+: FOLFIRI × 6 months, 2 ctDNA-/+: CAPOX × 3 months, switch to FOLFIRI if still ctDNA-/+, 3ctDNA+/-: CAPE × 3 months, switch to FOLFIRI if ctDNA+/+ after 3 LB, (4) ctDNA-/-: follow-up, switch to CAPOX if ctDNA+ after 2 LB |
| Initial Attack on Latent Metastasis Using TAS-102 for ctDNA Identified CRC After Curative Resection | NCT04457297 (ALTAIR) | Japan 2020–2023 | I-IV resected | 240 | Int rand/3/DFS, OS, TT | ctDNA/Signatera test | After radical surgery and SOC ACT, ctDNA+ patients are randomized to: (1) EXPrm: TAS-102 × 6 months or (2) SOC arm: placebo x 6 months |
| A Phase II Clinical Trial Comparing the Efficacy of RO7198457 Versus Watchful Waiting in Patients With ctDNA positive, Resected HR Stage II and Stage III CRC | NCT04486378 BNT122-01 | USA 2020–2023 | HR II-III | 201 | Int rand/2/DFS, OS, TTR, TTF, TT | ctDNA | After radicalsurgery, ctDNA+ patients receive SOC ACT and are then randomized to: (1) EXP arm: RO7198457 (a personalized cancer vaccine) or (2) SOC arm: watchful waiting |
| ctDNA Analysis to Optimize Treatment for Patients with CRC | NCT03637686 (IMPROVE) | Denmark 2018–2026 | III | 1800 | Obs/DFS | ctDNA | Part I—Surgery: ctDNA detection pre- and postoperative. Part II—Surveillance: ctDNA detection over 5 years follow-up |
| BESPOKE study of ctDNA guided therapy in CRC | NCT04264702 | USA 2020–2024 | II-III | 1000 | Obs/DFS | ctDNA/Signatera test | To examine the impact of SIGNATERA test on ACT decisions and clinical outcomes during a 2-year follow-up |
| Use of ctDNA for Monitoring of Stage III CRC | NCT02842203 (PRO16020374) | USA 2016–2021 | III | 150 | Obs/OS, PFS | ctDNA | ctDNA serial assessment up to 5 years and correlation with CEA and clinical outcomes |
| The implication of ctDNA in the recurrence surveillance of stage II and III CRC | NCT03416478 FFJC2017-01 | China 2018–2020 | II-III | 50 | Obs/DFS, OS | ctDNA | ctDNA serial assessment before and after curative surgery up to 2 years of follow-up |
| A phase II Clinical Trial comparing Efficacy of RO7198457 vs. watchful waiting in ctDNA positive stage II–III resected CRC | NCT04486378 | USA 2020–2027 | II-III | 201 | Int rand/2/DFS, RFS, TTR, TTF, OS, TT, ctDNA clear | ctDNA | To compare the efficacy of RO7198457 vs. watchful waiting after surgery and SOC ACT in ctDNA positive stage II–III CRC |
| ctDNA as a Prognostic Marker for Postoperative Relapse in Early and Intermediate Stage CRC | NCT03312374 | China 2017–2020 | II-III | 350 | Obs/DFS | ctDNA/NGS | ctDNA serial assessment before and after curative surgery and ACT up to 2 years of follow-up |
| The Implication of Plasma ctDNA Methylation Haplotypes in Detecting CRC and Adenomas | NCT03737591 | China 2018–2020 | I-IV adenomas healthy | 500 | Obs | ctDNA/NGS | To evaluate the sensitivity and specificity of ctDNA methylation haplotypes in detecting CRC and adenomas |
| Dynamic monitoring of ctDNA methylation to predict relapse in stage II–III CRC after radical resection | NCT03737539 | China 2018–2022 | II-III | 300 | Obs/DFS | ctDNA/NGS | To correlate and compare postoperative, pre- and post-ACT ctDNA methylation markers with radiological imaging and clinical outcomes |
| Ct-DNA Testing in Guiding Treatment for Patients With Advanced or Metastatic CRC | NCT03844620 | USA 2019–2020 | III (uncurable)-IV | 100 | Int/2/TT, ctDNA clean, QoL, ORR, OS | ctDNA | Monitoring and correlating ctDNA changes and radiological progression or TT incidence during third-line SOC (TAS-102/Regorafenib) (arm A) vs. third-line SOC alone (arm B) |
| Predictive and Prognostic Value of Inflammatory Markers and microRNA in Stage IV CRC | NCT04149613 | USA 2018–2021 | IV | 100 | Obs | miRNAs | To evaluate the expression of selected microRNAs and inflammatory markers in patients with stage IV CRC and assess their correlation with tumor location, dietary patterns, survival rates, response to systemic chemotherapy, and other clinic-pathological parameters |
| Molecular Pathology of CRC: Investigating the Role of Novel Molecular Profiles, microRNAs, and their Targets in CRC Progression | NCT03309722 | UK 2008–2025 | I-IV | 1000 | Obs/OS, DFS, LR, DR | miRNA | Single-center observational cohort study of prospectively recruited patients for biomarker evaluation and identification of novel biomarkers |
| Contents of Circulating Extracellular Vesicles: Biomarkers in CRC Patients | NCT04523389 (ExoColon) | France 2020–2021 | I-IV | 172 | Obs/OS, PFS, LR, DR | Exosomes | To investigate the prognostic and predictive role of exosomes and their contents (miRNAs and others) |
| ColoCare Transdiciplinary Research in CRC Prognosis | NCT02328677 | USA 2007–2030 | I-IV | 5000 | Obs/OS, DFS, QoL, TT | ctDNA, MiRNAs | To investigate the prognostic and predictive role of liquid biopsy in CRC patients in a 5-year follow-up |
| Timing To Minimally Invasive Surgery After Neoadjuvant Chemoradiotherapy For Rectal Cancer: A Multicenter Randomized Controlled Trial—Biomarkers SubStudy | NCT03962088 (TiMiSNAR) | Italy 2019–2023 | II-III | 200 | Obs/pCR, DFS | miRNA/miRNeasy Mini kit by Qiagen | (1) To investigate the association between pre-neoadjuvant and post-neoadjuvant expression levels of miRNA with pCR. (2) To investigate the correlation between changes in expression levels of miRNA following complete surgical resection with DFS and the relation between changes in miRNA during surveillance and tumor relapse |
| microRNAs Tool for Stratifying Stage II CRC: a Perspective Study of ACT | NCT02635087 | China 2015–2025 | II | 630 | Obs/DFS, OS | miRNA | To investigate the predictive and prognostic role of miRNAs in stage II CRC, stratifying patients at “high risk” and at “low risk” of recurrence according to a six miRNAs tool |
| Assessment Of Long Noncoding RNA CCAT1 Using Real-Time Polymerase Chain Reaction In CRC patients | NCT04269746 | Egypt 2020–2021 | Diagnostic | 100 | Obs | lncRNA | To evaluate the clinical utility of detecting long non-coding RNA (CCAT1) expression in diagnosis of CRC patients and its relation to tumor staging |
| Brief Study Title | NCT Number/Study Name | Country Study Period | Stage | N | Study Type/Phase (If Applicable) | Intervention | Study Overview/Schematic Description |
|---|---|---|---|---|---|---|---|
| Gut Microbiome Dynamics in Metastasized or Irresectable CRC | NCT03941080 GIMICC | Netherlands 2020–2022 | IV | 300 | Obs | Fecal and blood sample collection + behavioral questionnaire at baseline and every 3 months | To investigate characteristics and alterations of the gut microbiome and its predictive value for RR and TT during CT for mCRC |
| Gut Microbiome and Oral Fluoropyrimidine Study in Patients With CRC | NCT04054908 GO | USA 2018–2022 | all | 60 | Obs | 1 stool sample at baseline and at least 1 stool sample during treatment + questionnaires regarding bowel habits and dietary habits | To investigate the alterations of the gut microbiome occurring in three cohorts of CRC: Cohort A: patients treated with CAPE as SOC, Cohort B: patients treated with TAS-102 with or without Y-90 radioembolization in T, Cohort C: patients treated with CAPE + pembrolizumab + bevacizumab in T |
| Human Intestinal Microbiome and Surgical Outcomes in Patients Undergoing CRCCancer Surgery | NCT04005118 Microbiota | France | all | 50 | Obs | 2 fresh fecal samples for LM detection (1 pre- and 1 post-operatively) + 1 intraoperatively sample for MAM | To investigate the association between microbiome composition and occurrence of postoperative complications (anastomotic leakage, surgical site infection, prolonged postoperative ileus) |
| Bowel Preparation Impact on the Intestinal Microbiome: Oral Preparation vs. Enema | NCT04013841 BowelPrepMicrobiome | USA 2020–2022 | Left-sided CRC | 60 | Int rand | Stool samples before and after bowel preparation and surgery | To investigate differences in microbiome composition according to oral and enema bowel preparation for left side colon surgery and its correlation with surgical outcomes |
| The Role of Microbiome in Cancer Therapy | NCT02960282 | USA 2016–2021 | IV | 80 | Obs | Fecal specimen collection at baseline, prior to each cycle and at PD or off-treatment | To investigate microbiome composition, its gene and protein expression profile and correlation with RR and TT in two cohorts: Cohort A: patients treated with FOLFOX or FOLFIRI CT backbone as first line regimen, Cohort B: patients treated with pembrolizumab |
| Colorectal Cancer Cohort Study | NCT04185779 COLO-COHORT | UK 2019–2024 | diagnostic | 15000 | Obs | Blood and fecal tests + behavioral questionnaires | To develop a prediction model to stratify patients at risk of having adenomas or CRC (past medical history, family history, blood tests, FIT level, colonscopy, and microbiome stool) |
| Stool and Blood Sample Bank for CRC Patients | NCT04638751 ARGONAUT | USA 2020–2024 | III–IV | 4000 | 2 blood and stool samples each over a 6-month period | To determine whether the microbiome composition can predict PFS and OS in different cohorts of cancer patients (NSCLC, CRC, TNBC, and PC) treated with CT or IT. To identify correlations between microbiome composition and immune markers | |
| Omega-3 Fatty Acid for the Immune Modulation of Colorectal Cancer | NCT03661047 OMICC | USA 2019–2023 | I-III, HR adenomas | 36 | Int rand/2/ | Blood and stool samples + lifestyle questionnaire + nutritional survey between CRC/adenomas detection and surgery | To evaluate the effect of a 30-day administration of AMR101 (VASCEPA, icosapent ethyl) on MO3PUFA composition, gut microbiome, and immune system elements concentration (CD8+ T cells, CD49b, CTLA-4, PD-L1, PD-1, LAG-3, IL10, FOXP3) in both normal and tumor tissue |
| Metagenomic Evaluation of the Gut Microbiome in Patients With Lynch Syndrome and Other Hereditary Colonic Polyposis Syndromes | NCT02371135 | USA 2015–2021 | High hereditary CRC risk | 225 | Obs | Stool sample + Brief Diet and Lifestyle Questionnaire before every colonscopy | To investigate the association of the gut microbiome and dietary factors with risk of adenoma or cancer in Lynch syndrome and other hereditary colonic polyposis syndrome patients |
| Pilot Trial of Resistant Starch in Stage I-III CRC Survivors | NCT03781778 | USA 2018–2020 | I–III | 24 | Int rand/2 | Stool samples + Diet questionnaire at beginning and at 8 weeks | To compare the effect of a 8-week consumption of foods made of resistant (experimental arm) or corn (control arm) starch in addition to usual daily diet in modifying markers of inflammation, insulin resistance, and gut microbiome composition of CRC patients |
| Development and Analysis of a Stool Bank for Cancer Patients | NCT04291755 | USA 2019–2021 | all | 100 | Obs | Five stool, blood, and urine samples each over a 12-month period | To investigate the impact of gut microbiota on the efficacy of immune checkpoint inhibitors in NSCLC and CRC patients |
| Microbiome and Rectal Cancer | NCT04223102 | USA 2020–2027 | II–III | 40 | Int | Serial rectal biopsy specimens in a 5-year follow-up | To investigate the association between microbiome and pathologic response to neoadjuvant therapy in rectal cancer |
| Fecal Microbiota Transplant (FMT) Capsule for Improving the Efficacy of Anti- PD-1 | NCT04130763 | China 2020–2021 | IV | 10 | Int | Induction dose with FMT capsules one week before anti-PD-1 treatment beginning followed by maintenance dose | To determine whether the FMT capsule improves ORR of anti-PD-1 treatment in resistant/refractory gastrointestinal cancer patients |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parisi, A.; Porzio, G.; Pulcini, F.; Cannita, K.; Ficorella, C.; Mattei, V.; Delle Monache, S. What Is Known about Theragnostic Strategies in Colorectal Cancer. Biomedicines 2021, 9, 140. https://doi.org/10.3390/biomedicines9020140
Parisi A, Porzio G, Pulcini F, Cannita K, Ficorella C, Mattei V, Delle Monache S. What Is Known about Theragnostic Strategies in Colorectal Cancer. Biomedicines. 2021; 9(2):140. https://doi.org/10.3390/biomedicines9020140
Chicago/Turabian StyleParisi, Alessandro, Giampiero Porzio, Fanny Pulcini, Katia Cannita, Corrado Ficorella, Vincenzo Mattei, and Simona Delle Monache. 2021. "What Is Known about Theragnostic Strategies in Colorectal Cancer" Biomedicines 9, no. 2: 140. https://doi.org/10.3390/biomedicines9020140
APA StyleParisi, A., Porzio, G., Pulcini, F., Cannita, K., Ficorella, C., Mattei, V., & Delle Monache, S. (2021). What Is Known about Theragnostic Strategies in Colorectal Cancer. Biomedicines, 9(2), 140. https://doi.org/10.3390/biomedicines9020140