
Soluble Factors and Receptors Involved in Skin Innate Immunity—What Do We Know So Far?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
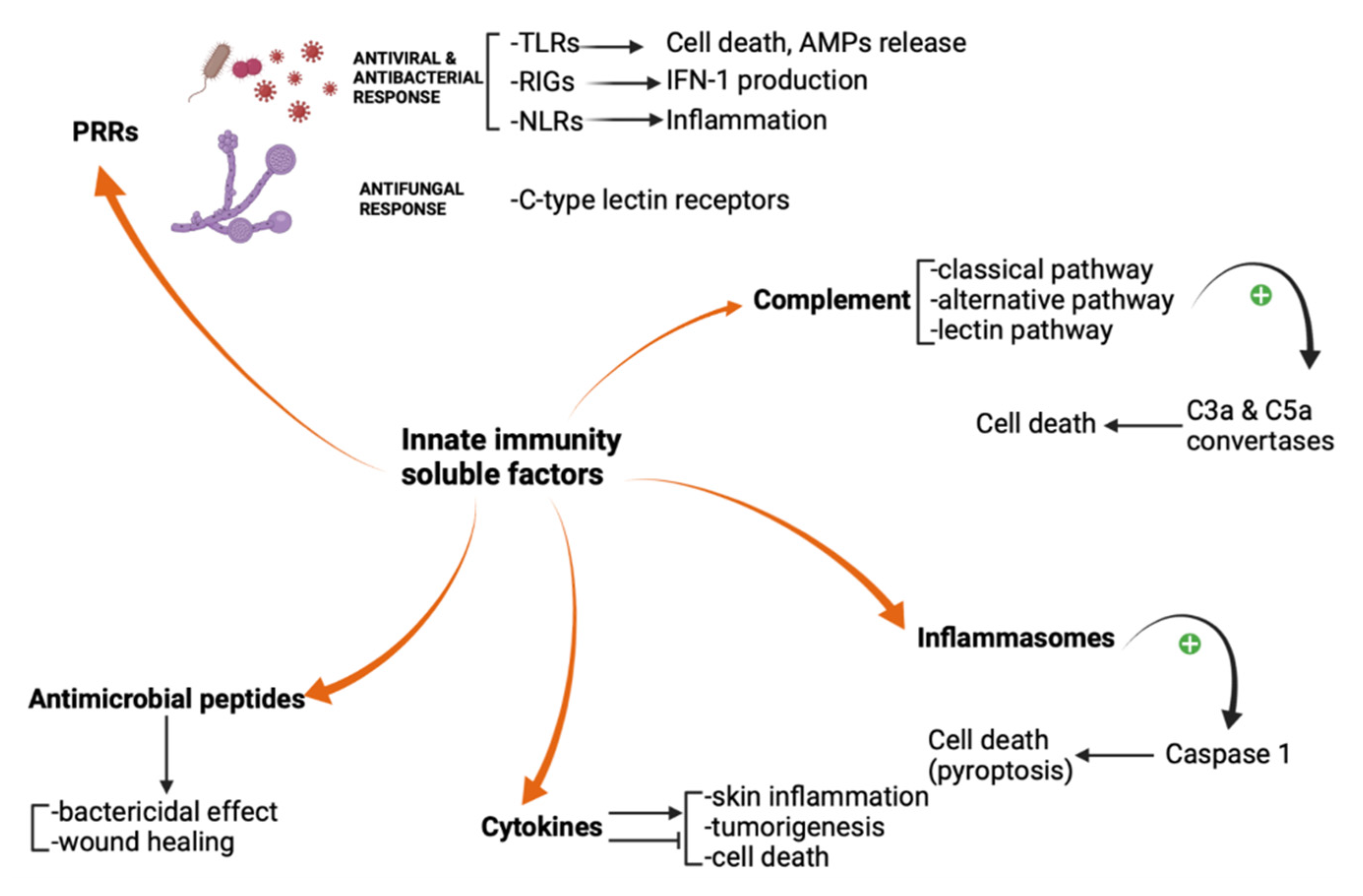
2. Pattern Recognition Receptors
3. Complement
4. Inflammasomes
5. Antimicrobial Peptides
6. Cytokines
7. How Does Imiquimod Target PRRs?
7.1. Imiquimod in Basal Cell Carcinoma (BCC)
7.2. Imiquimod in Actinic Keratosis
7.3. Imiquimod in Condyloma Acuminatum
7.4. Imiquimod Off-Label Use
7.4.1. Bowen Disease
7.4.2. Bowenoid Papulosis
7.4.3. Erythroplasia of Queyrat
7.4.4. Keratoacanthoma (KA)
7.4.5. Squamous Cell Carcinoma (SCC)
7.4.6. In Situ Melanoma
7.4.7. Molluscum Contagiosum
7.4.8. Herpes Simplex Virus (HSV)
7.4.9. Porokeratosis
7.4.10. Morphea
7.4.11. Discoid Lupus Erythematosus (DLE)
8. Targeting Other Soluble Factors
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schwartz, T. Immunology. In Dermatology; Bolognia, J.L., Schaffer, J.V., Cerroni, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 4; pp. 81–83. [Google Scholar]
- Bonefeld, C.M.; Geisler, C. The role of innate lymphoid cells in healthy and inflamed skin. Immunol. Lett. 2016, 179, 25–28. [Google Scholar] [CrossRef]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Hoffert, U. Innate Immunity in the Skin. J. Innate Immun. 2012, 4, 223–224. [Google Scholar] [CrossRef] [PubMed]
- McKernan, D.P. Pattern recognition receptors as potential drug targets in inflammatory disorders. Adv. Protein Chem. Struct. Biol. 2020, 119, 65–109. [Google Scholar] [CrossRef]
- Ermertcan, A.; Ozturk, F.; Gunduz, K. Toll-like receptors and skin. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 997–1006. [Google Scholar] [CrossRef]
- Wenzel, J. Cutaneous lupus erythematosus: New insights into pathogenesis and therapeutic strategies. Nat. Rev. Rheumatol. 2019, 15, 519–532. [Google Scholar] [CrossRef]
- McInturff, J.E.; Modlin, R.L.; Kim, J. The Role of Toll-like Receptors in the Pathogenesis and Treatment of Dermatological Disease. J. Investig. Dermatol. 2005, 125, 1–8. [Google Scholar] [CrossRef]
- Vignesh, P.; Rawat, A.; Sharma, M.; Singh, S. Complement in autoimmune diseases. Clin. Chim. Acta 2017, 465, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Palianus, J.; Meri, S. Complement System in Dermatological Diseases—Fire Under the Skin. Front. Med. 2015, 2, 3. [Google Scholar] [CrossRef]
- Pasch, M.C.; Bos, J.D.; Daha, M.R.; Asghar, S.S. Transforming growth factor-beta isoforms regulate the surface expression of mem-brane cofactor protein (CD46) and CD59 on human keratinocytes [corrected]. Eur. J. Immunol. 1999, 29, 100–108, Erratum in Eur. J. Immunol. 1999, 29, 1751. [Google Scholar] [CrossRef]
- Tang, L.; Zhou, F. Inflammasomes in Common Immune-Related Skin Diseases. Front. Immunol. 2020, 11, 882. [Google Scholar] [CrossRef] [PubMed]
- Beer, H.-D.; Contassot, E.; French, L.E. The Inflammasomes in Autoinflammatory Diseases with Skin Involvement. J. Investig. Dermatol. 2014, 134, 1805–1810. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zeng, M.Y.; Yang, D.; Motro, B.; Núñez, G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 2016, 530, 354–357. [Google Scholar] [CrossRef]
- Contassot, E.; Beer, H.-D.; French, L.E. Interleukin-1, inflammasomes, autoinflammation and the skin. Swiss Med Wkly. 2012, 142, w13590. [Google Scholar] [CrossRef]
- Kuemmerle-Deschner, J.B.; Hachulla, E.; Cartwright, R.; Hawkins, P.N.; Tran, T.A.; Bader-Meunier, B.; Hoyer, J.; Gattorno, M.; Gül, A.; Smith, J.; et al. Two-year results from an open-label, multicentre, phase III study evaluating the safety and efficacy of canakinumab in patients with cryopyrin-associated periodic syndrome across different severity phenotypes. Ann. Rheum. Dis. 2011, 70, 2095–2102. [Google Scholar] [CrossRef]
- Di Micco, A.; Frera, G.; Lugrin, J.; Jamilloux, Y.; Hsu, E.-T.; Tardivel, A.; De Gassart, A.; Zaffalon, L.; Bujisic, B.; Siegert, S.; et al. AIM2 inflammasome is activated by pharmacological disruption of nuclear envelope integrity. Proc. Natl. Acad. Sci. USA 2016, 113, E4671–E4680. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Gallo, R.L.; Kenshi, Y.; Richard, L.G. Antimicrobial peptides in human skin disease. Eur. J. Dermatol. EJD 2007, 18, 11–21. [Google Scholar] [CrossRef]
- Gallo, R.L. Sounding the Alarm: Multiple Functions of Host Defense Peptides. J. Investig. Dermatol. 2008, 128, 5–6. [Google Scholar] [CrossRef]
- Kristian, S.A.; Datta, V.; Weidenmaier, C.; Kansal, R.; Fedtke, I.; Peschel, A.; Gallo, R.L.; Nizet, V. D-Alanylation of Teichoic Acids Promotes Group A Streptococcus Antimicrobial Peptide Resistance, Neutrophil Survival, and Epithelial Cell Invasion. J. Bacteriol. 2005, 187, 6719–6725. [Google Scholar] [CrossRef]
- Schauber, J.; Gallo, R.L. Antimicrobial peptides and the skin immune defense system. J. Allergy Clin. Immunol. 2008, 122, 261–266. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Antimicrobial peptides activity in the skin. Ski. Res. Technol. 2019, 25, 111–117. [Google Scholar] [CrossRef]
- Heilborn, J.D.; Nilsson, M.F.; Sorensen, O.E.; Ståhle-Bäckdahl, M.; Kratz, G.; Weber, G.; Borregaard, N. The Cathelicidin Anti-Microbial Peptide LL-37 is Involved in Re-Epithelialization of Human Skin Wounds and is Lacking in Chronic Ulcer Epithelium. J. Investig. Dermatol. 2003, 120, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Charo, I.F.; Ransohoff, R.M. The Many Roles of Chemokines and Chemokine Receptors in Inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Geginat, J.; Larghi, P.; Paroni, M.; Nizzoli, G.; Penatti, A.; Pagani, M.; Gagliani, N.; Meroni, P.L.; Abrignani, S.; Flavell, R.A. The light and the dark sides of Interleukin-10 in immune-mediated diseases and cancer. Cytokine Growth Factor Rev. 2016, 30, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Gröne, A. Keratinocytes and cytokines. Veter.-Immunol. Immunopathol. 2002, 88, 1–12. [Google Scholar] [CrossRef]
- Sobhan, M.; Hojati, M.; Vafaie, S.-Y.; Ahmadimoghaddam, D.; Mohammadi, Y.; Mehrpooya, M. The Efficacy of Colloidal Oatmeal Cream 1% as Add-on Therapy in the Management of Chronic Irritant Hand Eczema: A Double-Blind Study. Clin. Cosmet. Investig. Dermatol. 2020, 13, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Artaza-Irigaray, C.; Molina-Pineda, A.; Aguilar-Lemarroy, A.; Ortiz-Lazareno, P.; Limón-Toledo, L.P.; Pereira-Suárez, A.L.; Rojo-Contreras, W.; Jave-Suárez, L.F. E6/E7 and E6* From HPV16 and HPV18 Upregulate IL-6 Expression Independently of p53 in Keratinocytes. Front Immunol. 2019, 10, 1676. [Google Scholar] [CrossRef]
- Gasque, P.; Jaffar-Bandjee, M.C. The immunology and inflammatory responses of human melanocytes in infectious diseases. J. Infect. 2015, 71, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Moretti, S.; Pinzi, C.; Spallanzani, A.; Berti, E.; Chiarugi, A.; Mazzoli, S.; Fabiani, M.; Vallecchi, C.; Herlyn, M. Immuno-histochemical evidence of cytokine networks during progression of human melanocytic lesions. Int. J. Cancer 1999, 84, 160–168. [Google Scholar] [CrossRef]
- Zhou, J.; Yuen, N.K.; Zhan, Q.; Velazquez, E.F.; Murphy, G.F.; Giobbie-Hurder, A.; Hodi, F.S. Immunity to the melanoma inhibitor of apoptosis protein (ML-IAP; livin) in patients with malignant melanoma. Cancer Immunol. Immunother. 2012, 61, 655–665. [Google Scholar] [CrossRef][Green Version]
- Hartman, M.L. Non-Apoptotic Cell Death Signaling Pathways in Melanoma. Int. J. Mol. Sci. 2020, 21, 2980. [Google Scholar] [CrossRef]
- Hänel, K.H.; Cornelissen, C.; Lüscher, B.; Baron, J.M. Cytokines and the Skin Barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [PubMed]
- Braunstein, M.J.; Kucharczyk, J.; Adams, S. Targeting Toll-Like Receptors for Cancer Therapy. Target. Oncol. 2018, 13, 583–598. [Google Scholar] [CrossRef]
- Papakostas, D.; Stockfleth, E. Topical treatment of basal cell carcinoma with the immune response modifier imiquimod. Future Oncol. 2015, 11, 2985–2990. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-W.; Wang, S.-T.; Chang, S.-H.; Chuang, K.-C.; Wang, H.-Y.; Kao, J.-K.; Liang, S.-M.; Wu, C.-Y.; Kao, S.-H.; Chen, Y.-J.; et al. Imiquimod Exerts Antitumor Effects by Inducing Immunogenic Cell Death and Is Enhanced by the Glycolytic Inhibitor 2-Deoxyglucose. J. Investig. Dermatol. 2020, 140, 1771–1783.e6. [Google Scholar] [CrossRef]
- Jia, H.-X.; He, Y.-L. Efficacy and safety of imiquimod 5% cream for basal cell carcinoma: A meta-analysis of randomized controlled trial. J. Dermatol. Treat. 2020, 31, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Bath-Hextall, F.; Ozolins, M.; Armstrong, S.J.; Colver, G.B.; Perkins, W.; Miller, P.S.J.; Williams, H.C.; Surgery versus Imiquimod for Nodular Superficial Basal Cell Carcinoma (SINS) Study Group. Surgical excision versus imiquimod 5% cream for nodular and superficial basal-cell carcinoma (SINS): A multicentre, non-inferiority, randomised controlled trial. Lancet Oncol. 2013, 15, 96–105. [Google Scholar] [CrossRef]
- Peris, K.; Fargnoli, M.C.; Garbe, C.; Kaufmann, R.; Bastholt, L.; Seguin, N.B.; Bataille, V.; Marmol, V.D.; Dummer, R.; Harwood, C.A.; et al. Diagnosis and treatment of basal cell carcinoma: European consensus–based interdisciplinary guidelines. Eur. J. Cancer 2019, 118, 10–34. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kawamura, T.; Matsuzawa, T.; Aoki, R.; Shimada, S. Recruitment of plasmacytoid dendritic cells to skin regulates treatment responsiveness of actinic keratosis to imiquimod. J. Dermatol. Sci. 2014, 76, 67–69. [Google Scholar] [CrossRef]
- Jansen, M.H.E.; Kessels, J.P.H.M.; Nelemans, P.J.; Kouloubis, N.; Arits, A.H.; Van Pelt, H.P.; Quaedvlieg, P.J.F.; Essers, B.A.B.; Steijlen, P.M.; Kelleners-Smeets, N.W.; et al. Randomized Trial of Four Treatment Approaches for Actinic Keratosis. N. Engl. J. Med. 2019, 380, 935–946. [Google Scholar] [CrossRef]
- Hadley, G.; Derry, S.; Moore, R.A. Imiquimod for Actinic Keratosis: Systematic Review and Meta-Analysis. J. Investig. Dermatol. 2006, 126, 1251–1255. [Google Scholar] [CrossRef]
- Stockfleth, E. Lmax and imiquimod 3.75%: The new standard in AK management. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.M.; Jung, C.; Lee, W.; Won, C.; Lee, M.; Choi, J.; Chang, S. Topically applied treatments for external genital warts in nonimmunocompromised patients: A systematic review and network meta-analysis. Br. J. Dermatol. 2020, 183, 24–36. [Google Scholar] [CrossRef]
- Leung, A.K.; Barankin, B.; Leong, K.F.; Hon, K.L. Penile warts: An update on their evaluation and management. Drugs Context 2018, 7, 212563. [Google Scholar] [CrossRef]
- Akhavan, S.; Mohammadi, S.R.; Modarres Gillani, M.; Mousavi, A.S.; Shirazi, M. Efficacy of combination therapy of oral zinc sulfate with imiquimod, podophyllin or cryotherapy in the treatment of vulvar warts. J. Obstet. Gynaecol. Res. 2014, 40, 2110–2113. [Google Scholar] [CrossRef]
- David, C.V.; Nguyen, H.; Goldenberg, G. Imiquimod: A review of off-label clinical applications. J. Drugs Dermatol. 2011, 10, 1300–1306. [Google Scholar] [PubMed]
- Rosen, T.; Harting, M.; Gibson, M. Treatment of Bowen’s Disease with Topical 5% Imiquimod Cream: Retrospective Study. Dermatol. Surg. 2007, 33, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie-Wood, A.; Kossard, S.; de Launey, J.; Wilkinson, B.; Owens, M.L. Imiquimod 5% cream in the treatment of Bowen’s disease. J. Am. Acad. Dermatol. 2001, 44, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Matuszewski, M.; Michajłowski, I.; Michajłowski, J.; Sokołowska-Wojdyło, M.; Włodarczyk, A.; Krajka, K. Topical treatment of bowenoid papulosis of the penis with imiquimod. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 978–979. [Google Scholar] [CrossRef]
- Nunes, M.d.G.; Trope, B.M.; Cavalcanti, S.M.; Oliveira, L.d.H.; Ramos-e-Silva, M. Bowenoid papulosis in a patient with AIDS treated with imiquimod: Case report. Acta Dermatovenerol. Croat. 2004, 12, 278–281. [Google Scholar]
- Lim, J.H.-L.; Lim, K.-S.; Chong, W.-S. Dramatic clearance of HIV-associated bowenoid papulosis using combined oral acitretin and topical 5% imiquimod. J. Drugs Dermatol. JDD 2014, 13, 901–902. [Google Scholar] [PubMed]
- Yokoyama, M.; Egawa, G.; Makino, T.; Egawa, K. Erythroplasia of Queyrat treated with imiquimod 5% cream: The necessity of regimen guidelines. Clin. Case Rep. 2019, 7, 723–725. [Google Scholar] [CrossRef]
- Micali, G.; Nasca, M.R.; De Pasquale, R. Erythroplasia of Queyrat treated with imiquimod 5% cream. J. Am. Acad. Dermatol. 2006, 55, 901–903. [Google Scholar] [CrossRef]
- Pancevski, G.; Pepic, S.; Idoska, S.; Tofoski, G.; Nikolovska, S. Topical Imiquimod 5% as a Treatment Option in Solitary Facial Keratoacanthoma. Open Access Maced. J. Med. Sci. 2018, 6, 531–535. [Google Scholar] [CrossRef]
- Barysch, M.J.; Kamarashev, J.; Lockwood, L.L.; Dummer, R. Successful treatment of multiple keratoacanthoma with topical imiquimod and low-dose acitretin. J. Dermatol. 2010, 38, 390–392. [Google Scholar] [CrossRef]
- Tran, D.; Li, S.; Henry, S.; Wood, D.; Chang, A. An 18-year retrospective study on the outcomes of keratoacanthomas with different treatment modalities at a single academic centre. Br. J. Dermatol. 2017, 177, 1749–1751. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kozlow, J.H.; Mittal, B.; Moyer, J.; Olenecki, T.; Rodgers, P.; Alam, M.; Armstrong, A.; Baum, C.; Bordeaux, J.S.; et al. Guidelines of care for the management of cutaneous squamous cell carcinoma. J. Am. Acad. Dermatol. 2018, 78, 560–578. [Google Scholar] [CrossRef] [PubMed]
- Que, S.K.T.; Zwald, F.O.; Schmults, C.D. Cutaneous squamous cell carcinoma: Management of advanced and high-stage tumors. J. Am. Acad. Dermatol. 2018, 78, 249–261. [Google Scholar] [CrossRef]
- Swetter, S.M.; Tsao, H.; Bichakjian, C.K.; Curiel-Lewandrowski, C.; Elder, D.E.; Gershenwald, J.E.; Guild, V.; Grant-Kels, J.M.; Halpern, A.C.; Johnson, T.M.; et al. Guidelines of care for the management of primary cutaneous melanoma. J. Am. Acad. Dermatol. 2019, 80, 208–250. [Google Scholar] [CrossRef] [PubMed]
- Merritt, B.G.; Degesys, C.A.; Brodland, D.G. Extramammary Paget Disease. Dermatol. Clin. 2019, 37, 261–267, Erratum in Dermatol. Clin. 2019, 37, xiii. [Google Scholar] [CrossRef] [PubMed]
- Odyakmaz Demirsoy, E.; Bayramgürler, D.; Çağlayan, Ç.; Bilen, N.; Şikar Aktürk, A.; Kıran, R. Imiquimod 5% Cream Versus Cryotherapy in Classic Kaposi Sarcoma. J. Cutan. Med. Surg. 2019, 23, 488–495. [Google Scholar] [CrossRef]
- Shalabi, D.; Vadalia, N.; Nikbakht, N. Revisiting Imiquimod for Treatment of Folliculotropic Mycosis Fungoides: A Case Report and Review of the Literature. Dermatol. Ther. 2019, 9, 807–814. [Google Scholar] [CrossRef]
- Sisti, A.; Sisti, G.; Oranges, C.M. Topical treatment of melanoma skin metastases with imiquimod: A review. Dermatol. Online J. 2014, 21, 25756475. [Google Scholar] [CrossRef]
- Gualdi, G.; Pascalucci, C.; Panarese, F.; Prignano, F.; Giuliani, F.; Verga, E.; Amerio, P.; Verdolini, R. Molluscum contagiosum in pediatric patients: To treat or not to treat? Could a personalized imiquimod regimen be the answer to the dilemma? J. Dermatol. Treat. 2020, 1–6, epub ahead of print. [Google Scholar] [CrossRef]
- Katz, K.A.; Williams, H.C.; Van Der Wouden, J.C. Imiquimod cream for molluscum contagiosum: Neither safe nor effective. Pediatr. Dermatol. 2018, 35, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.S.; Hasbun, M.T.; Jones, K.M.; Schmidt, B.A.R.; Hussain, S.H. Pemphigus-like eruption as a complication of molluscum contagiosum treatment with imiquimod in a 5-year-old girl. Pediatr. Dermatol. 2020, 37, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, F.; Muhamad, I.I. Acrylamide-based hydrogel drug delivery systems: Release of Acyclovir from MgO nanocomposite hydrogel. J. Taiwan Inst. Chem. Eng. 2017, 72, 182–193. [Google Scholar] [CrossRef]
- Perkins, N.; Nisbet, M.; Thomas, M. Topical imiquimod treatment of aciclovir-resistant herpes simplex disease: Case series and literature review. Sex. Transm. Infect. 2011, 87, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Kan, Y.; Okabayashi, T.; Yokota, S.-I.; Yamamoto, S.; Fujii, N.; Yamashita, T. Imiquimod Suppresses Propagation of Herpes Simplex Virus 1 by Upregulation of Cystatin A via the Adenosine Receptor A1 Pathway. J. Virol. 2012, 86, 10338–10346. [Google Scholar] [CrossRef]
- Jahanban-Esfahlan, R.; Seidi, K.; Majidinia, M.; Karimian, A.; Yousefi, B.; Nabavi, S.M.; Astani, A.; Berindan-Neagoe, I.; Gulei, D.; Fallarino, F.; et al. Toll-like receptors as novel therapeutic targets for herpes simplex virus infection. Rev. Med. Virol. 2019, 29, e2048. [Google Scholar] [CrossRef]
- Valins, W.; Amini, S.; Berman, B. The Expression of Toll-like Receptors in Dermatological Diseases and the Therapeutic Effect of Current and Newer Topical Toll-like Receptor Modulators. J. Clin. Aesthetic Dermatol. 2010, 3, 20–29. [Google Scholar]
- Montes-De-Oca-Sánchez, G.; Tirado-Sánchez, A.; García-Ramírez, V. Porokeratosis of Mibelli of the axillae: Treatment with topical imiquimod. J. Dermatol. Treat. 2006, 17, 319–320. [Google Scholar] [CrossRef]
- Harrison, S.; Sinclair, R. Porokeratosis of Mibelli: Successful treatment with topical 5% imiquimod cream. Australas. J. Dermatol. 2003, 44, 281–283. [Google Scholar] [CrossRef]
- Jensen, J.-M.; Egberts, F.; Proksch, E.; Hauschild, A. Disseminated porokeratosis palmaris and plantaris treated with imiquimod cream to prevent malignancy. Acta Derm. Venereol. 2005, 85, 550–551. [Google Scholar] [CrossRef]
- Ahn, S.-J.; Lee, H.-J.; Chang, S.-E.; Lee, M.-W.; Choi, J.-H.; Moon, K.-C.; Koh, J.-K. Case of linear porokeratosis: Successful treatment with topical 5% imiquimod cream. J. Dermatol. 2007, 34, 146–147. [Google Scholar] [CrossRef] [PubMed]
- Pope, E.; Doria, A.S.; Thériault, M.; Mohanta, A.; Laxer, R.M. Topical Imiquimod 5% Cream for Pediatric Plaque Morphea: A Prospective, Multiple-Baseline, Open-Label Pilot Study. Dermatology 2011, 223, 363–369. [Google Scholar] [CrossRef]
- Dytoc, M.; Wat, H.; Cheung-Lee, M.; Sawyer, D.; Ackerman, T.; Fiorillo, L. Evaluation of the Efficacy and Safety of Topical Imiquimod 5% for Plaque-Type Morphea: A Multicenter, Prospective, Vehicle-Controlled Trial. J. Cutan. Med. Surg. 2015, 19, 132–139. [Google Scholar] [CrossRef]
- Gül, U.; Gönül, M.; Cakmak, S.K.; Kiliç, A.; Demiriz, M. A case of generalized discoid lupus erythematosus: Successful treatment with imiquimod cream 5%. Adv. Ther. 2006, 23, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Turan, E.; Sinem Bagci, I.; Turgut Erdemir, A.; Salih Gurel, M. Successful treatment of generalized discoid lupus erythematosus with imiquimod cream 5%: A case report and review of the literature. Acta Dermatovenerol. Croat. 2014, 22, 150–159. [Google Scholar] [PubMed]
- Jiménez-Gallo, D.; Brandão, B.J.F.; Arjona-Aguilera, C.; Báez-Perea, J.M.; Linares-Barrios, M. Imiquimod-induced cutaneous lupus erythematosus with antinuclear antibodies showing a homogenous pattern. Clin. Exp. Dermatol. 2017, 42, 795–797. [Google Scholar] [CrossRef]
- Giang, J.; Seelen, M.A.J.; Van Doorn, M.B.A.; Rissmann, R.; Prens, E.P.; Damman, J. Complement Activation in Inflammatory Skin Diseases. Front. Immunol. 2018, 9, 639. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, M.S.; Ballanti, E.; Triggianese, P.; Perricone, R. Vasculitides and the Complement System: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2015, 49, 333–346. [Google Scholar] [CrossRef]
- Ghias, M.H.; Hyde, M.J.; Tomalin, L.E.; Morgan, B.P.; Alavi, A.; Lowes, M.A.; Piguet, V. Role of the Complement Pathway in Inflammatory Skin Diseases: A Focus on Hidradenitis Suppurativa. J. Investig. Dermatol. 2020, 140, 531–536.e1. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03001622 (accessed on 12 August 2021).
- De Benedetti, F.; Gattorno, M.; Anton, J.; Ben-Chetrit, E.; Frenkel, J.; Hoffman, H.M.; Koné-Paut, I.; Lachmann, H.; Ozen, S.; Simon, A.; et al. Canakinumab for the Treatment of Autoinflammatory Recurrent Fever Syndromes. N. Engl. J. Med. 2018, 378, 1908–1919. [Google Scholar] [CrossRef]
- Betrains, A.; Staels, F.; Vanderschueren, S. Efficacy and safety of canakinumab treatment in Schnitzler syndrome: A systematic literature review. Semin. Arthritis Rheum. 2020, 50, 636–642. [Google Scholar] [CrossRef]
- Kolios, A.; Maul, J.-T.; Meier, B.; Kerl, K.; Traidl-Hoffmann, C.; Hertl, M.; Zillikens, D.; Röcken, M.; Ring, J.; Facchiano, A.; et al. Canakinumab in adults with steroid-refractory pyoderma gangrenosum. Br. J. Dermatol. 2015, 173, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.Z.; Ro, T.; Jolly, P.; Sayed, C.J. Non-response to Interleukin-1 Antagonist Canakinumab in Two Patients with Refractory Pyoderma Gangrenosum and Hidradenitis Suppurativa. J. Clin. Aesthet. Dermatol. 2017, 10, 36–38. [Google Scholar]
- Leslie, K.S.; Tripathi, S.V.; Nguyen, T.V.; Pauli, M.; Rosenblum, M.D. An open-label study of anakinra for the treatment of moderate to severe hidradenitis suppurativa. J. Am. Acad. Dermatol. 2014, 70, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Seegräber, M.; Srour, J.; Walter, A.; Knop, M.; Wollenberg, A. Dupilumab for treatment of atopic dermatitis. Expert Rev. Clin. Pharmacol. 2018, 11, 467–474. [Google Scholar] [CrossRef]
- Papp, K.A.; Blauvelt, A.; Bukhalo, M.; Gooderham, M.; Krueger, J.G.; Lacour, J.-P.; Menter, A.; Philipp, S.; Sofen, H.; Tyring, S.; et al. Risankizumab versus Ustekinumab for Moderate-to-Severe Plaque Psoriasis. N. Engl. J. Med. 2017, 376, 1551–1560. [Google Scholar] [CrossRef]
- Deodhar, A.; Gottlieb, A.B.; Boehncke, W.-H.; Dong, B.; Wang, Y.; Zhuang, Y.; Barchuk, W.; Xu, X.L.; Hsia, E.C.; CNTO1959PSA2001 Study Group. Efficacy and safety of guselkumab in patients with active psoriatic arthritis: A randomised, double-blind, placebo-controlled, phase 2 study. Lancet 2018, 391, 2213–2224. [Google Scholar] [CrossRef]
- Bai, F.; Li, G.G.; Liu, Q.; Niu, X.; Li, R.; Ma, H. Short-Term Efficacy and Safety of IL-17, IL-12/23, and IL-23 Inhibitors Brodalumab, Secukinumab, Ixekizumab, Ustekinumab, Guselkumab, Tildrakizumab, and Risankizumab for the Treatment of Moderate to Severe Plaque Psoriasis: A Systematic Review and Network Meta-Analysis of Randomized Controlled Trials. J. Immunol. Res. 2019, 2019, 2546161. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Read, C. Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. JAMA 2020, 323, 1945–1960. [Google Scholar] [CrossRef]
- Mahil, S.K.; Ezejimofor, M.C.; Exton, L.S.; Manounah, L.; Burden, A.D.; Coates, L.C.; De Brito, M.; McGuire, A.; Murphy, R.; Owen, C.M.; et al. Comparing the efficacy and tolerability of biologic therapies in psoriasis: An updated network meta-analysis. Br. J. Dermatol. 2020, 183, 638–649. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scurtu, L.G.; Simionescu, O. Soluble Factors and Receptors Involved in Skin Innate Immunity—What Do We Know So Far? Biomedicines 2021, 9, 1795. https://doi.org/10.3390/biomedicines9121795
Scurtu LG, Simionescu O. Soluble Factors and Receptors Involved in Skin Innate Immunity—What Do We Know So Far? Biomedicines. 2021; 9(12):1795. https://doi.org/10.3390/biomedicines9121795
Chicago/Turabian StyleScurtu, Lucian G., and Olga Simionescu. 2021. "Soluble Factors and Receptors Involved in Skin Innate Immunity—What Do We Know So Far?" Biomedicines 9, no. 12: 1795. https://doi.org/10.3390/biomedicines9121795
APA StyleScurtu, L. G., & Simionescu, O. (2021). Soluble Factors and Receptors Involved in Skin Innate Immunity—What Do We Know So Far? Biomedicines, 9(12), 1795. https://doi.org/10.3390/biomedicines9121795