Identification of Enzymes Oxidizing the Tyrosine Kinase Inhibitor Cabozantinib: Cabozantinib Is Predominantly Oxidized by CYP3A4 and Its Oxidation Is Stimulated by cyt b5 Activity


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Oxidation of Cabozantinib by Hepatic microsomes and Human Recombinant Enzymes
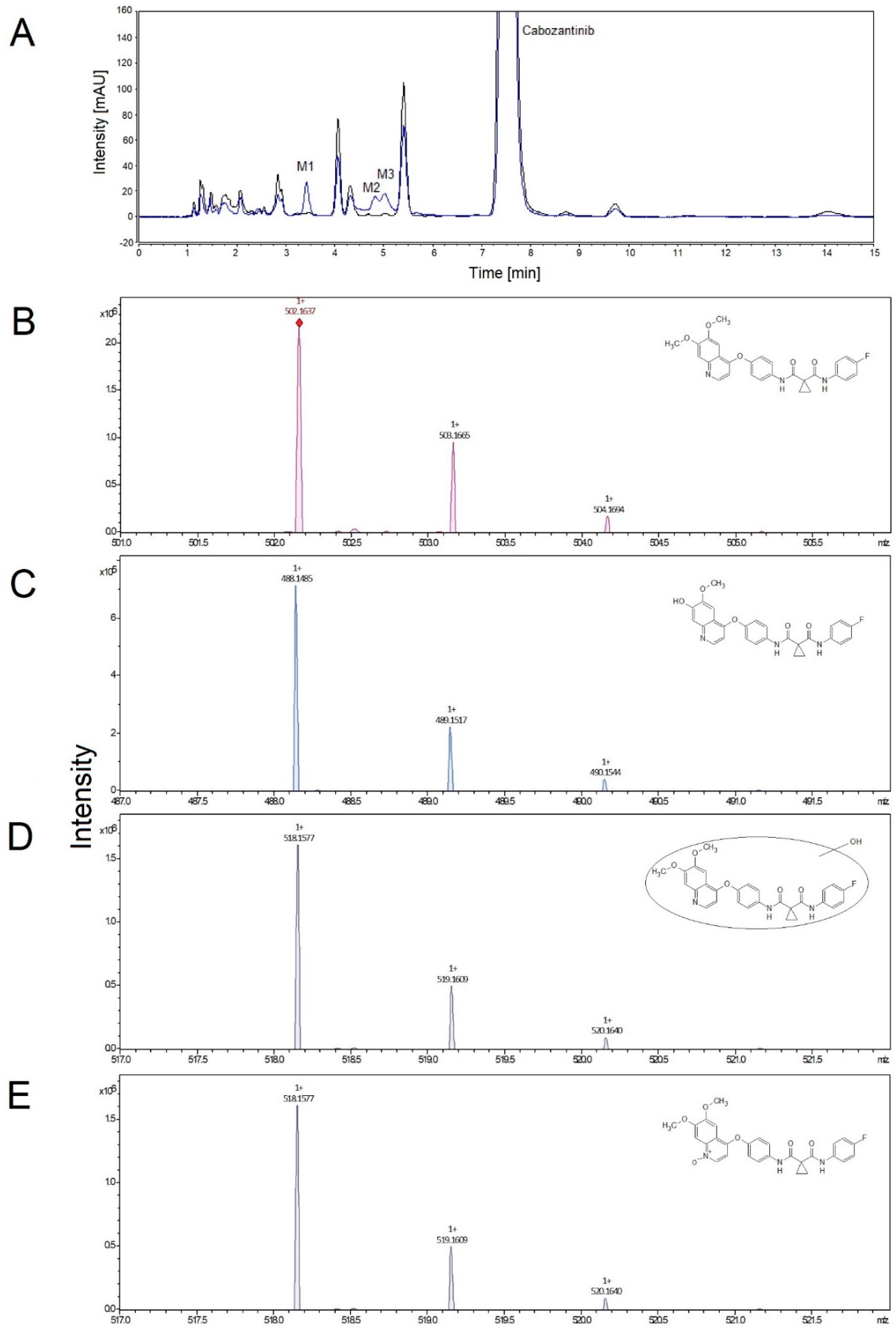
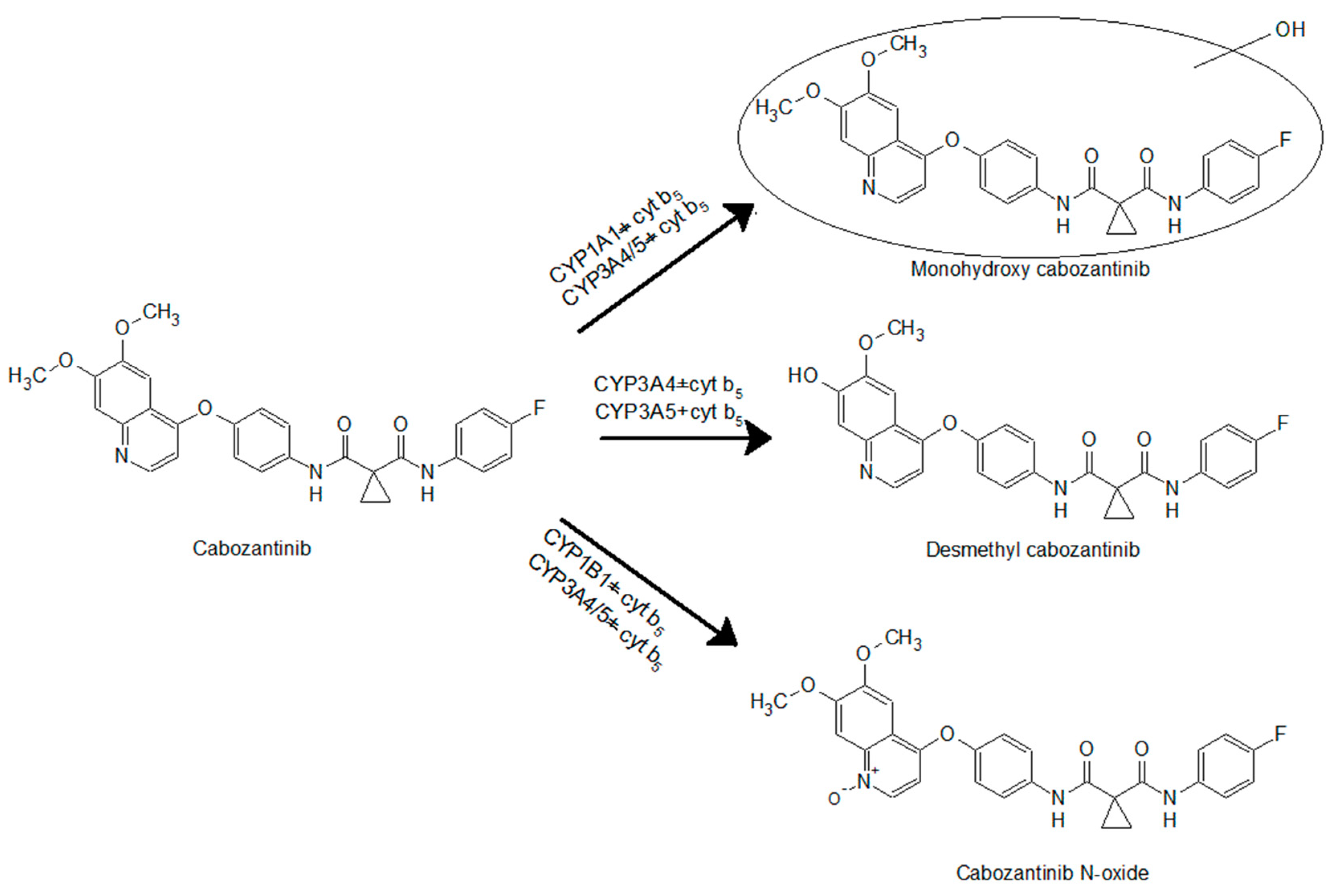
2.3. Identification of Cabozantinib Metabolites by Mass Spectrometry
2.4. Inhibition Study
2.5. Contributions of CYP Enzymes to Formation of Cabozantinib Metabolites in Human Livers
2.6. Statistical Analysis
3. Results
3.1. Oxidation of Cabozantinib by Human Hepatic Microsomes and Correlation of CYPs Activities with Cabozantinib Oxidation
3.2. The Effect of CYP Enzyme Inhibitors on Cabozantinib Oxidation in Human Liver Microsomes
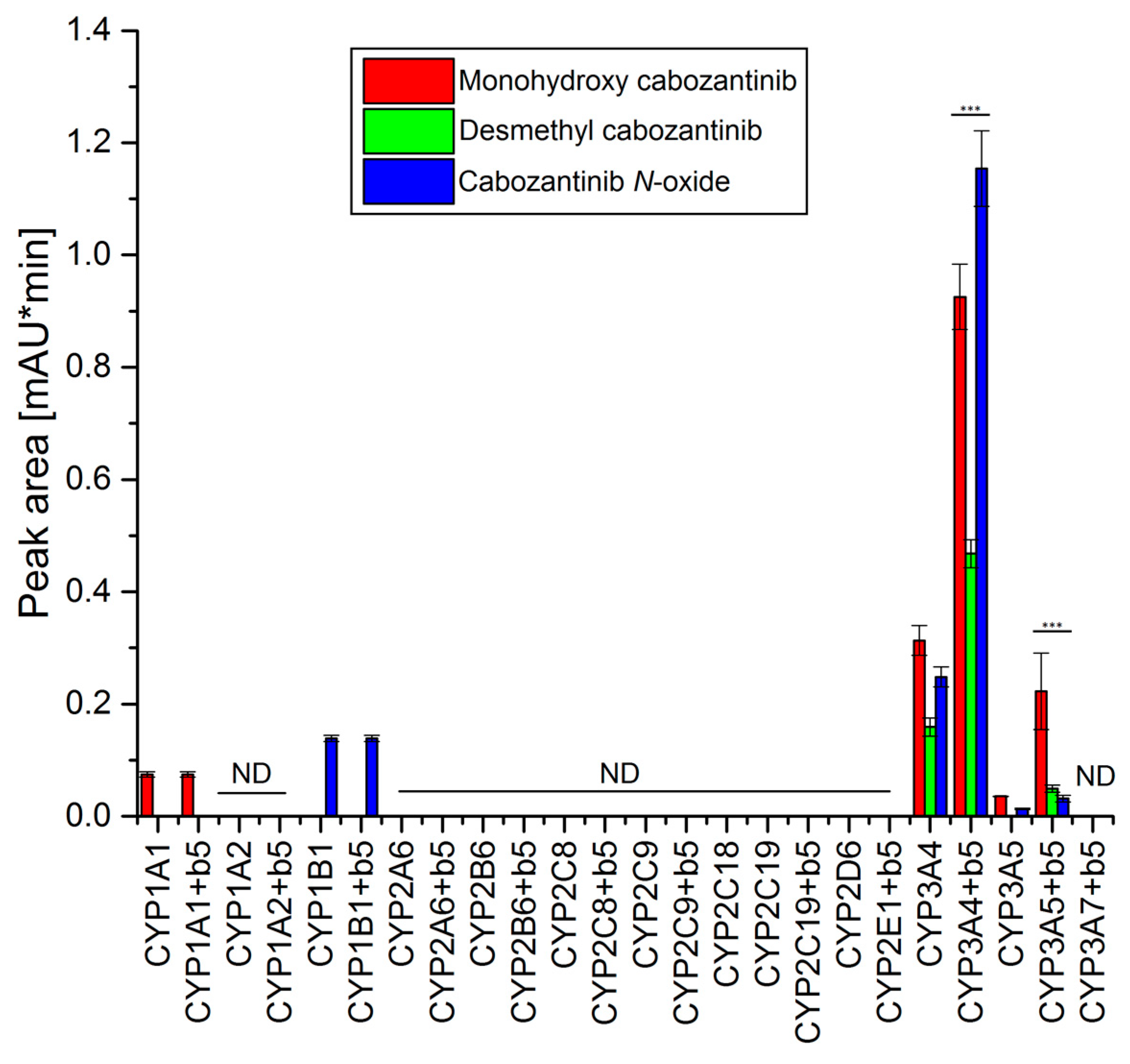
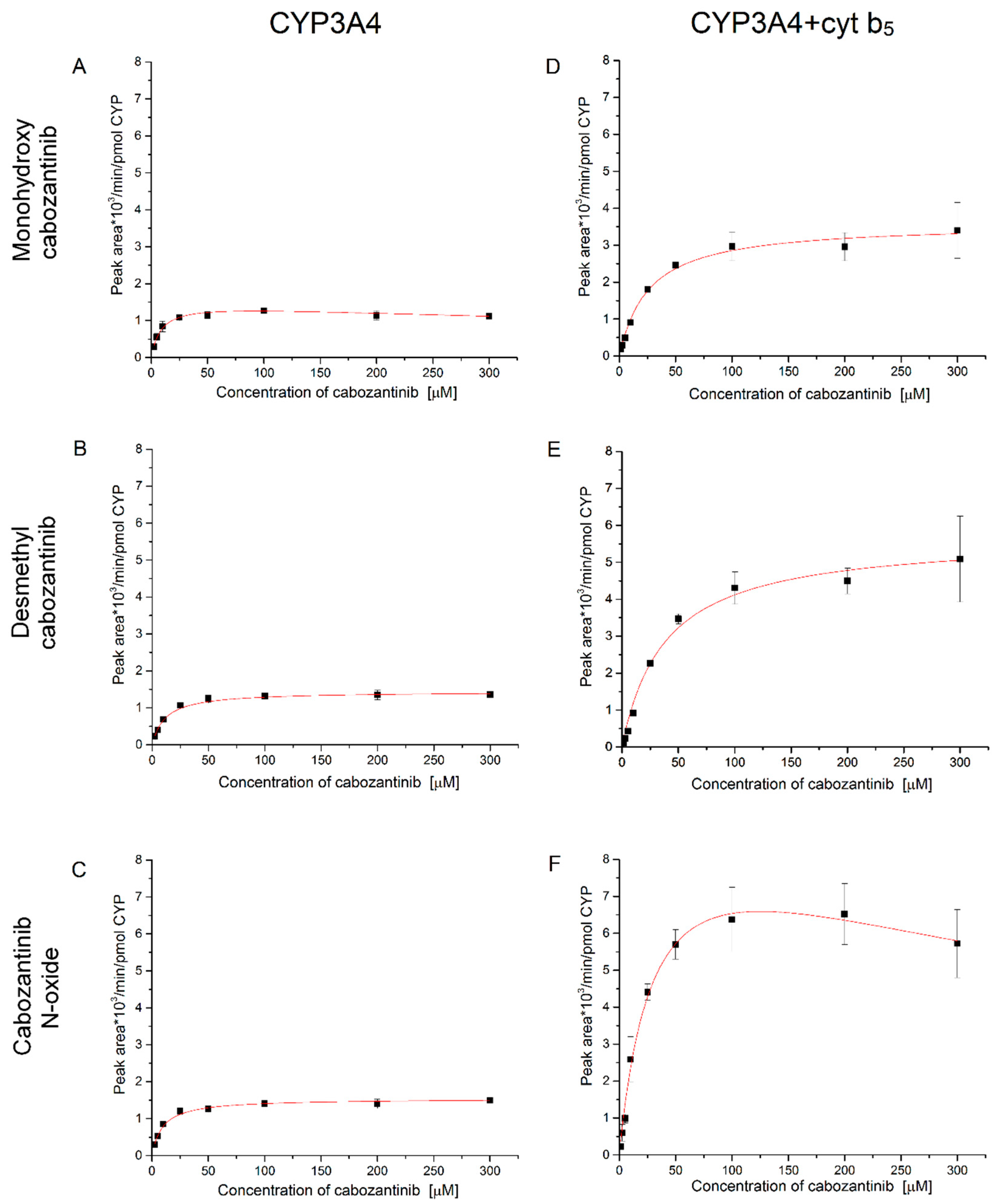
3.3. Oxidation of Cabozantinib by Recombinant Human Cytochromes P450
3.4. Oxidation of Cabozantinib by Recombinant Human Flavin-Containing Mono-oxygenases and Aldehyde Oxidase
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| α-NF | α-Naphthoflavone |
| AO | Aldehyde oxidase |
| CYP | Cytochrome P450 |
| Cyt b5 | Cytochrome b5 |
| DDTC | Diethyldithiocarbamate |
| FMO | Flavin-containing mono-oxygenase |
| HPLC | High-performance liquid chromatography |
| M1-3 | Cabozantinib metabolites |
| MET | Hepatocyte growth factor receptor |
| POR | NADPH:CYP oxidoreductase |
| RET | Rearranged during transfection receptor |
| TKI | Tyrosine kinase inhibitor |
| VEGFR | Vascular endothelial growth factor receptor |
References
- Wilkinson, G.R. Drug metabolism and variability among patients in drug response. N. Engl. J. Med. 2005, 352, 2211–2221. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K. Cytochrome P450 and anticancer drugs. Curr. Drug Metab. 2006, 7, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Kurzrock, R.; Sherman, S.I.; Ball, D.W.; Forastiere, A.A.; Cohen, R.B.; Mehra, R.; Pfister, D.G.; Cohen, E.E.W.; Janisch, L.; Nauling, F.; et al. Activity of XL184 (Cabozantinib), an oral tyrosine kinase inhibitor, in patients with medullary thyroid cancer. J. Clin. Oncol. 2011, 29, 2660–2666. [Google Scholar] [CrossRef] [PubMed]
- Tolaney, S.M.; Nechushtan, H.; Ron, I.; Schöffski, P.; Awada, A.; Yasenchak, C.A.; Laird, A.D.; O’Keeffe, B.; Shapiro, G.I.; Winer, E.R. Cabozantinib for metastatic breast carcinoma: Results of a phase II placebo-controlled randomized discontinuation study. Breast Cancer Res. Treat. 2016, 160, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Tolaney, S.M.; Ziehr, D.R.; Guo, H.; Ng, M.R.; Barry, W.T.; Higgins, M.J.; Isakoff, S.J.; Brock, J.E.; Ivanova, E.V.; Paweletz, C.P.; et al. Phase II and biomarker study of cabozantinib in metastatic triple-negative breast cancer patients. Oncologist 2017, 22, 25–32. [Google Scholar] [CrossRef]
- Kelley, R.K.; Verslype, C.; Cohn, A.L.; Yang, T.S.; Su, W.C.; Burris, H.; Braiteh, F.; Vogelzang, N.; Spira, A.; Foster, P.; et al. Cabozantinib in hepatocellular carcinoma: Results of a phase 2 placebo-controlled randomized discontinuation study. Ann. Oncol. 2017, 28, 528–534. [Google Scholar] [CrossRef]
- Abou-Alfa, G.K.; Meyer, T.; Cheng, A.L.; El-Khoueiry, A.B.; Rimassa, L.; Ryoo, B.Y.; Cicin, I.; Merle, P.; Chen, Y.H.; Park, J.W.; et al. Cabozantinib in patients with advanced and progressing hepatocellular carcinoma. N. Engl. J. Med. 2018, 379, 54–63. [Google Scholar] [CrossRef]
- Drilon, A.; Wang, L.; Hasanovic, A.; Suehara, Y.; Lipson, D.; Stephens, P.; Roos, J.; Miller, V.; Ginsberg, M.; Zakowski, M.F.; et al. Response to Cabozantinib in patients with RET fusion-positive lung adenocarcinomas. Cancer Discov. 2013, 3, 630–635. [Google Scholar] [CrossRef]
- Nokihara, H.; Nishio, M.; Yamamoto, N.; Fujiwara, Y.; Horinouchi, H.; Kanda, S.; Horiike, A.; Ohyanagi, F.; Yanagitani, N.; Nguyen, L.; et al. Phase 1 Study of Cabozantinib in Japanese Patients with Expansion Cohorts in Non–Small-Cell Lung Cancer. Clin. Lung Cancer 2019, 20, e317–e328. [Google Scholar] [CrossRef]
- Smith, D.C.; Smith, M.R.; Sweeney, C.; Elfiky, A.A.; Logothetis, C.; Corn, P.G.; Vogelzang, N.J.; Small, E.J.; Harzstark, A.L.; Gordon, S.; et al. Cabozantinib in patients with advanced prostate cancer: Results of a phase II randomized discontinuation trial. J. Clin. Oncol. 2013, 31, 412–419. [Google Scholar] [CrossRef]
- Dai, J.; Zhang, H.; Karatsinides, A.; Keller, J.M.; Kozloff, K.M.; Aftab, D.T.; Schimmoller, F.; Keller, E.T. Cabozantinib inhibits prostate cancer growth and prevents tumor-induced bone lesions. Clin. Cancer Res. 2014, 20, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, T.K.; Escudier, B.; Powles, T.; Mainwaring, P.N.; Rini, B.I.; Donskov, F.; Hammers, H.; Hutson, T.E.; Lee, J.L.; Peltola, K.; et al. Cabozantinib versus everolimus in advanced renal-cell carcinoma. N. Engl. J. Med. 2015, 373, 1814–1823. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Escudier, B.; Powles, T.; Tannir, N.M.; Mainwaring, P.N.; Rini, B.I.; Hammers, H.J.; Donskov, F.; Roth, B.J.; Peltola, K.; et al. Cabozantinib versus everolimus in advanced renal cell carcinoma (METEOR): Final results from a randomised, open-label, phase 3 trial. Lancet Oncol. 2016, 17, 917–927. [Google Scholar] [CrossRef]
- Hage, C.; Rausch, V.; Giese, N.; Giese, T.; Schönsiegel, F.; Labsch, S.; Nwaeburu, C.; Mattern, J.; Gladkich, J.; Herr, I. The novel c-Met inhibitor cabozantinib overcomes gemcitabine resistance and stem cell signaling in pancreatic cancer. Cell Death Dis. 2013, 4, e627. [Google Scholar] [CrossRef] [PubMed]
- Yakes, F.M.; Chen, J.; Tan, J.; Yamaguchi, K.; Shi, Y.; Yu, P.; Qian, F.; Chu, F.; Bentzien, F.; Cancilla, B.; et al. Cabozantinib (XL184), a novel MET and VEGFR2 inhibitor, simultaneously suppresses metastasis, angiogenesis, and tumor growth. Mol. Cancer Ther. 2011, 10, 2298–2308. [Google Scholar] [CrossRef]
- Bentzien, F.; Zuzow, M.; Heald, N.; Gibson, A.; Shi, Y.; Goon, L.; Yu, P.; Engst, S.; Zhang, W.; Huang, D.; et al. In vitro and in vivo activity of cabozantinib (XL184), an inhibitor of RET, MET, and VEGFR2, in a model of medullary thyroid cancer. Thyroid 2013, 23, 1569–1577. [Google Scholar] [CrossRef]
- Xiang, Q.; Chen, W.; Ren, M.; Wang, J.; Zhang, H.; Deng, D.Y.B.; Zhang, L.; Shang, C.; Chen, Y. Cabozantinib suppresses tumor growth and metastasis in hepatocellular carcinoma by a dual blockade of VEGFR2 and MET. Clin. Cancer Res. 2014, 20, 2959–2970. [Google Scholar] [CrossRef]
- Lacy, S.; Hsu, B.; Miles, D.; Aftab, D.; Wang, R.; Nguyen, L. Metabolism and disposition of cabozantinib in healthy male volunteers and pharmacologic characterization of its major metabolites. Drug Metab. Dispos. 2015, 43, 1190–1207. [Google Scholar] [CrossRef]
- Gerendash, B.S.; Creel, P.A. Practical management of adverse events associated with cabozantinib treatment in patients with renal-cell carcinoma. OncoTargets Ther. 2017, 10, 5053. [Google Scholar] [CrossRef]
- FDA Center for Drug Evaluation and Research. Cabometyx Full Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2016/208692s000lbl.pdf (accessed on 20 September 2020).
- Stiborová, M.; Indra, R.; Mizerovská, M.; Frei, E.; Schmeiser, H.H.; Kopka, K.; Philips, D.H.; Arlt, V.M. NADH:Cytochrome b5 reductase and cytochrome b5 can act as sole electron donors to human cytochrome P450 1A1-mediated oxidation and DNA adduct formation by benzo[a]pyrene. Chem. Res. Toxicol. 2016, 29, 1325–1334. [Google Scholar] [CrossRef]
- Kotrbova, V.; Mrazova, B.; Moserova, M.; Martinek, V.; Hodek, P.; Hudecek, J.; Frei, E.; Stiborova, M. Cytochrome b5 shifts oxidation of the anticancer drug ellipticine by cytochromes P450 1A1 and 1A2 from its detoxication to activation, thereby modulating its pharmacological efficacy. Biochem. Pharmacol. 2011, 82, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Šulc, M.; Indra, R.; Moserová, M.; Schmeiser, H.H.; Frei, E.; Arlt, V.M.; Stiborová, M. The impact of individual cytochrome P450 enzymes on oxidative metabolism of benzo[a]pyrene in human livers. Environ. Mol. Mutagen. 2016, 57, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Stiborová, M.; Borek-Dohalska, L.; Aimova, D.; Kotrbova, V.; Kukackova, K.; Janouchova, K.; Rupertova, M.; Ryslava, H.; Hudecek, J.; Frei, E. Oxidation pattern of the anticancer drug ellipticine by hepatic microsomes—Similarity between human and rat systems. Gen. Physiol. Biophys. 2006, 25, 245–261. [Google Scholar] [PubMed]
- Rendic, S.; DiCarlo, F.J. Human cytochrome P450 enzymes: A status report summarizing their reactions, substrates, inducers, and inhibitors. Drug Metab. Rev. 1997, 29, 413–480. [Google Scholar] [CrossRef] [PubMed]
- Stiborová, M.; Martínek, V.; Rýdlová, H.; Hodek, P.; Frei, E. Sudan I is a potential carcinogen for humans: Evidence for its metabolic activation and detoxication by human recombinant cytochrome P450 1A1 and liver microsomes. Cancer Res. 2002, 62, 5678–5684. [Google Scholar] [PubMed]
- Stiborová, M.; Martínek, V.; Rýdlová, H.; Koblas, T.; Hodek, P. Expression of cytochrome P450 1A1 and its contribution to oxidation of a potential human carcinogen 1-phenylazo-2-naphthol (Sudan I) in human livers. Cancer Lett. 2005, 220, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Nakano, M.; Omak, Y.; Ueng, Y.F.; Guengerich, F.P.; Shimada, T. Roles of cytochrome b5 in the oxidation of testosterone and nifedipine by recombinant cytochrome P450 3A4 and by human liver microsomes. Arch. Biochem. Biophys. 1996, 325, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.D. The roles of cytochrome b5 in cytochrome P450 reactions. J. Biochem. Mol. Toxicol. 2002, 16, 311–316. [Google Scholar] [CrossRef]
- Stiborová, M.; Indra, R.; Frei, E.; Kopečková, K.; Schmeiser, H.H.; Eckschlager, T.; Adam, V.; Heger, Z.; Arlt, V.M.; Martínek, V. Cytochrome b5 plays a dual role in the reaction cycle of cytochrome P450 3A4 during oxidation of the anticancer drug ellipticine. Monatsh. Chem. 2017, 148, 1983–1991. [Google Scholar] [CrossRef]
- Indra, R.; Pompach, P.; Martínek, V.; Takácsová, P.; Vavrová, K.; Heger, Z.; Adam, V.; Eckschlager, T.; Kopečková, K.; Arlt, V.M.; et al. Identification of Human Enzymes Oxidizing the Anti-Thyroid-Cancer Drug Vandetanib and Explanation of the High Efficiency of Cytochrome P450 3A4 in its Oxidation. Int. J. Mol. Sci. 2019, 20, 3392. [Google Scholar] [CrossRef]
- Schenkman, J.B.; Jansson, I. The many roles of cytochrome b5. Pharmacol. Ther. 2003, 97, 139–152. [Google Scholar] [CrossRef]
- Achira, M.; Ito, K.; Suzuki, H.; Sugiyama, Y. Comparative studies to determine the selective inhibitors for P-glycoprotein and cytochrome P 4503A4. AAPS Pharmsci. 1999, 1, 14–19. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Greenblatt, D.J.; Zhao, Y.; Venkatakrishnan, K.; Duan, S.X.; Harmatz, J.S.; Parent, S.J.; Court, M.H.; von Moltke, L.L. Mechanism of cytochrome P450-3A inhibition by ketoconazole. J. Pharm. Pharmacol. 2011, 63, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Walsky, R.L.; Obach, R.S.; Hyland, R.; Kang, P.; Zhou, S.; West, M.; Geoghegan, K.F.; Helal, C.J.; Walker, G.S.; Goosen, T.C.; et al. Selective mechanism-based inactivation of CYP3A4 by CYP3cide (PF-04981517) and its utility as an in vitro tool for delineating the relative roles of CYP3A4 versus CYP3A5 in the metabolism of drugs. Drug Metab. Dispos. 2012, 40, 1686–1697. [Google Scholar] [CrossRef]
- Nguyen, L.; Holland, J.; Miles, D.; Engel, C.; Benrimoh, N.; O’Reilly, T.; Lacy, S. Pharmacokinetic (PK) drug interaction studies of cabozantinib: Effect of CYP3A inducer rifampin and inhibitor ketoconazole on cabozantinib plasma PK and effect of cabozantinib on CYP2C8 probe substrate rosiglitazone plasma PK. J. Clin. Pharmacol. 2015, 55, 1012–1023. [Google Scholar] [CrossRef]
- Lin, Q.M.; Li, Y.H.; Lu, X.R.; Wang, R.; Pang, N.H.; Xu, R.A.; Cai, J.P.; Hu, G.X. Characterization of Genetic Variation in CYP3A4 on the Metabolism of Cabozantinib in Vitro. Chem. Res. Toxicol. 2019, 32, 1583–1590. [Google Scholar] [CrossRef]
- Li, J.; Zhao, M.; He, P.; Hidalgo, M.; Baker, S.D. Differential metabolism of gefitinib and erlotinib by human cytochrome P450 enzymes. Clin. Cancer Res. 2007, 13, 3731–3737. [Google Scholar] [CrossRef]
- Van Erp, N.P.; Gelderblom, H.; Karlsson, M.O.; Li, J.; Zhao, M.; Ouwerkerk, J.; Nortier, J.W.; Guchelaar, H.J.; Baker, S.D.; Sparreboom, A. Influence of CYP3A4 inhibition on the steady-state pharmacokinetics of imatinib. Clin. Cancer Res. 2007, 13, 7394–7400. [Google Scholar] [CrossRef]
- Kim, A.; Balis, F.M.; Widemann, B.C. Sorafenib and sunitinib. Oncologist 2009, 14, 800. [Google Scholar] [CrossRef]
- Jackson, K.D.; Durandis, R.; Vergne, M.J. Role of cytochrome P450 enzymes in the metabolic activation of tyrosine kinase inhibitors. Int. J. Mol. Sci. 2018, 19, 2367. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Voice, M.W.; Zhang, Y.; Wolf, C.R.; Burchell, B.; Friedberg, T. Effects of Human Cytochrome b5on CYP3A4 Activity and Stability in Vivo. Arch. Biochem. Biophys. 1999, 366, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Stiborová, M.; Sejbal, J.; Bořek-Dohalská, L.; Aimová, D.; Poljaková, J.; Forsterová, K.; Rupertová, M.; Wiesner, J.; Hudeček, J.; Wiessler, M.; et al. The anticancer drug ellipticine forms covalent DNA adducts, mediated by human cytochromes P450, through metabolism to 13-hydroxyellipticine and ellipticine N2-oxide. Cancer Res. 2004, 64, 8374–8380. [Google Scholar] [CrossRef] [PubMed]
- Jushchyshyn, M.I.; Hutzler, J.M.; Schrag, M.L.; Wienkers, L.C. Catalytic turnover of pyrene by CYP3A4: Evidence that cytochrome b5 directly induces positive cooperativity. Arch. Biochem. Biophys. 2005, 438, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C.J.; McLaughlin, L.A.; Scheer, N.; Stanley, L.A.; Wolf, C.R. Cytochrome b5 is a major determinant of human cytochrome P450 CYP2D6 and CYP3A4 activity in vivo. Mol. Pharmacol. 2015, 87, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Ingelman-Sundberg, M.; Sim, S.C.; Gomez, A.; Rodriguez-Antona, C. Influence of cytochrome P450 polymorphisms on drug therapies: Pharmacogenetic, pharmacoepigenetic and clinical aspects. Pharmacol. Therap. 2007, 116, 496–526. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.E.; Yi, M.; Kim, W.Y.; Cho, S.A.; Lee, S.S.; Lee, S.J.; Shin, J.G. Influences of cytochrome b5 expression and its genetic variant on the activity of CYP2C9, CYP2C19 and CYP3A4. Drug Metab. Pharmacokinet. 2019, 34, 201–208. [Google Scholar] [CrossRef]
- Zhang, H.; Gao, N.; Liu, T.; Fang, Y.; Qi, B.; Wen, Q.; Zhou, J.; Jia, L.; Qiao, H. Effect of cytochrome b5 content on the activity of polymorphic CYP1A2, 2B6, and 2E1 in human liver microsomes. PLoS ONE 2015, 10, e0128547. [Google Scholar] [CrossRef]
- Takahashi, K.; Oda, Y.; Toyoda, Y.; Fukami, T.; Yokoi, T.; Nakajima, M. Regulation of cytochrome b5 expression by miR-223 in human liver: Effects on cytochrome P450 activities. Pharm. Res. 2014, 31, 780–794. [Google Scholar] [CrossRef]
- Sacco, J.C.; Trepanier, L.A. Cytochrome b5 and NADH cytochrome b5 reductase: Genotype-phenotype correlations for hydroxylamine reduction. Pharmacogenet. Genom. 2010, 20, 26. [Google Scholar] [CrossRef]
- Ruiz, J.N.; Belum, V.R.; Creel, P.; Cohn, A.; Ewer, M.; Lacouture, M.E. Current practices in the management of adverse events associated with targeted therapies for advanced renal cell carcinoma: A national survey of oncologists. Clin. Genitourin. Cancer 2014, 12, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, F.; Solas, C.; Giocanti, M.; Lacarelle, B.; Deville, J.L.; Gravis, G.; Ciccolini, J. A simple and rapid liquid chromatography-mass spectrometry method to assay cabozantinib in plasma: Application to therapeutic drug monitoring in patients with renal cell carcinoma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2020, 1138, 121968. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.; Jumbe, N.L.; Lacy, S.; Nguyen, L. Population pharmacokinetic model of cabozantinib in patients with medullary thyroid carcinoma and its application to an exposure-response analysis. Clin. Pharmacokinet. 2016, 55, 93–105. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total CYPs a | POR b | Cyt b5 c | CYP1A2 d | CYP2A6 d | CYP2B6 d | CYP2C8 d | CYP2C9 d | CYP2C19 d | CYP2D6 d | CYP2E1 d | CYP3A4 d | CYP4A11 d | FMO d | M1 e | M2 e | M3 e | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HG03 | 290 | 450 | 380 | 170 | 2000 | 51 | 200 | 1700 | 44 | 110 | 1800 | 6100 | 1600 | 0.287 | 0.335 | 1.672 | |
| HG103 | 340 | 210 | 790 | 310 | 440 | 7.2 | 39 | 2300 | 23 | 65 | 1100 | 2200 | 1600 | 1400 | 0.109 | 0.144 | 0.644 |
| HG24 | 260 | 260 | 550 | 1700 | 1500 | 35 | 190 | 3000 | 41 | - | 2300 | 4000 | 1800 | 1500 | 0.132 | 0.149 | 0.752 |
| HG32 | 170 | 330 | 580 | 730 | 520 | 0.68 | 20 | 450 | 4.8 | 46 | 1200 | 2000 | 680 | 920 | 0.208 | 0.185 | 1.338 |
| HG42 | 670 | 510 | 500 | 700 | 2200 | 150 | 480 | 1600 | 7.4 | 95 | 1600 | 15,000 | 1400 | 2000 | 0.301 | 0.329 | 1.609 |
| HG43 | 270 | 210 | 640 | 580 | 770 | 14 | 25 | 1800 | 440 | 4 | 780 | 4600 | 1800 | 920 | 0.755 | 0.625 | 3.491 |
| HG74 | 220 | 200 | 600 | 520 | 360 | 13 | 130 | 2100 | 55 | 120 | 1400 | 2700 | 1300 | 1200 | 0.148 | 0.256 | 1.106 |
| HG93 | 430 | 320 | 450 | 691 | 350 | 43 | 270 | 2200 | 75 | 49 | 2800 | 2800 | 1800 | 3500 | 0.292 | 0.371 | 1.935 |
| HK23 | 380 | 380 | 700 | 960 | 1100 | 24 | 160 | 2100 | 110 | 140 | 2100 | 6800 | 780 | 2500 | 0.094 | 0.130 | 0.607 |
| HK27 | 300 | 450 | 730 | 1320 | 1320 | 31 | 180 | 480 | 460 | 130 | 3000 | 4910 | 1110 | 2230 | 0.273 | 0.467 | 2.228 |
| HK31 | 580 | 540 | 770 | 1220 | 2160 | 8.1 | 130 | 1690 | 172 | 3.4 | 1660 | 8210 | 2010 | 3020 | 0.218 | 0.377 | 1.537 |
| HK34 | 500 | 460 | 890 | 1000 | 1500 | 39 | 220 | 1900 | 45 | 100 | 6000 | 5200 | 1100 | 2700 | 0.209 | 0.305 | 1.359 |
| Total CYPs | CYP1A2 | CYP2A6 | CYP2B6 | CYP2C8 | CYP2C9 | CYP2C19 | CYP2D6 | CYP2E1 | CYP3A4 | CYP4A11 | FMO | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M1 | 0.705 * | 0.104 | 0.607 * | 0.886 *** | 0.785 ** | 0.142 | −0.119 | 0.028 | −0.065 | 0.918 *** | 0.163 | 0.137 |
| M2 | 0.794 ** | 0.280 | 0.841 *** | 0.695 * | 0.654 * | 0.086 | 0.033 | −0.015 | 0.140 | 0.935 *** | 0.193 | 0.315 |
| M3 | 0.811 ** | 0.216 | 0.728 ** | 0.776 ** | 0.737 ** | 0.130 | −0.021 | −0.043 | 0.030 | 0.947 *** | 0.223 | 0.341 |
| Enzyme | Metabolite | Kinetic Characteristics a | ||
|---|---|---|---|---|
| VMax (peak area/min/nmol CYP) | K0.5 (μM) | Ki (μM) | ||
| CYP3A4 | Monohydroxy cabozantinib | 1.50 ± 0.05 | 8.98 ± 1.04 | 947.87 ± 202.96 |
| Desmethyl cabozantinib | 1.44 ± 0.03 | 11.69 ± 1.00 | NA | |
| Cabozantinib N-oxide | 1.54 ± 0.04 | 9.56 ± 0.69 | NA | |
| CYP3A4 + cyt b5 | Monohydroxy cabozantinib | 3.60 ± 0.11 | 26.00 ± 2.92 | NA |
| Desmethyl cabozantinib | 5.70 ± 0.24 | 38.53 ± 5.35 | NA | |
| Cabozantinib N-oxide | 9.96 ± 0.92 | 32.03 ± 5.68 | 490.22 ± 139.7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Indra, R.; Vavrová, K.; Pompach, P.; Heger, Z.; Hodek, P. Identification of Enzymes Oxidizing the Tyrosine Kinase Inhibitor Cabozantinib: Cabozantinib Is Predominantly Oxidized by CYP3A4 and Its Oxidation Is Stimulated by cyt b5 Activity. Biomedicines 2020, 8, 547. https://doi.org/10.3390/biomedicines8120547
Indra R, Vavrová K, Pompach P, Heger Z, Hodek P. Identification of Enzymes Oxidizing the Tyrosine Kinase Inhibitor Cabozantinib: Cabozantinib Is Predominantly Oxidized by CYP3A4 and Its Oxidation Is Stimulated by cyt b5 Activity. Biomedicines. 2020; 8(12):547. https://doi.org/10.3390/biomedicines8120547
Chicago/Turabian StyleIndra, Radek, Katarína Vavrová, Petr Pompach, Zbyněk Heger, and Petr Hodek. 2020. "Identification of Enzymes Oxidizing the Tyrosine Kinase Inhibitor Cabozantinib: Cabozantinib Is Predominantly Oxidized by CYP3A4 and Its Oxidation Is Stimulated by cyt b5 Activity" Biomedicines 8, no. 12: 547. https://doi.org/10.3390/biomedicines8120547
APA StyleIndra, R., Vavrová, K., Pompach, P., Heger, Z., & Hodek, P. (2020). Identification of Enzymes Oxidizing the Tyrosine Kinase Inhibitor Cabozantinib: Cabozantinib Is Predominantly Oxidized by CYP3A4 and Its Oxidation Is Stimulated by cyt b5 Activity. Biomedicines, 8(12), 547. https://doi.org/10.3390/biomedicines8120547