Relaxin-2 May Suppress Endometriosis by Reducing Fibrosis, Scar Formation, and Inflammation


 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Reagents and Materials
2.2. Immunohistochemistry
2.3. Isolation and Culture of Human Endometriotic Stromal Cells (ESCs)
2.4. Reverse Transcription (RT), Conventional Polymerase Chain Reaction (PCR), and Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
2.5. Measurement of Cyclic AMP (cAMP) and IL-8
2.6. Western Blotting
2.7. Collagen Gel Contraction Assay
2.8. Endometrial Transplantation Mouse Model
2.9. Statistical Analysis
3. Results
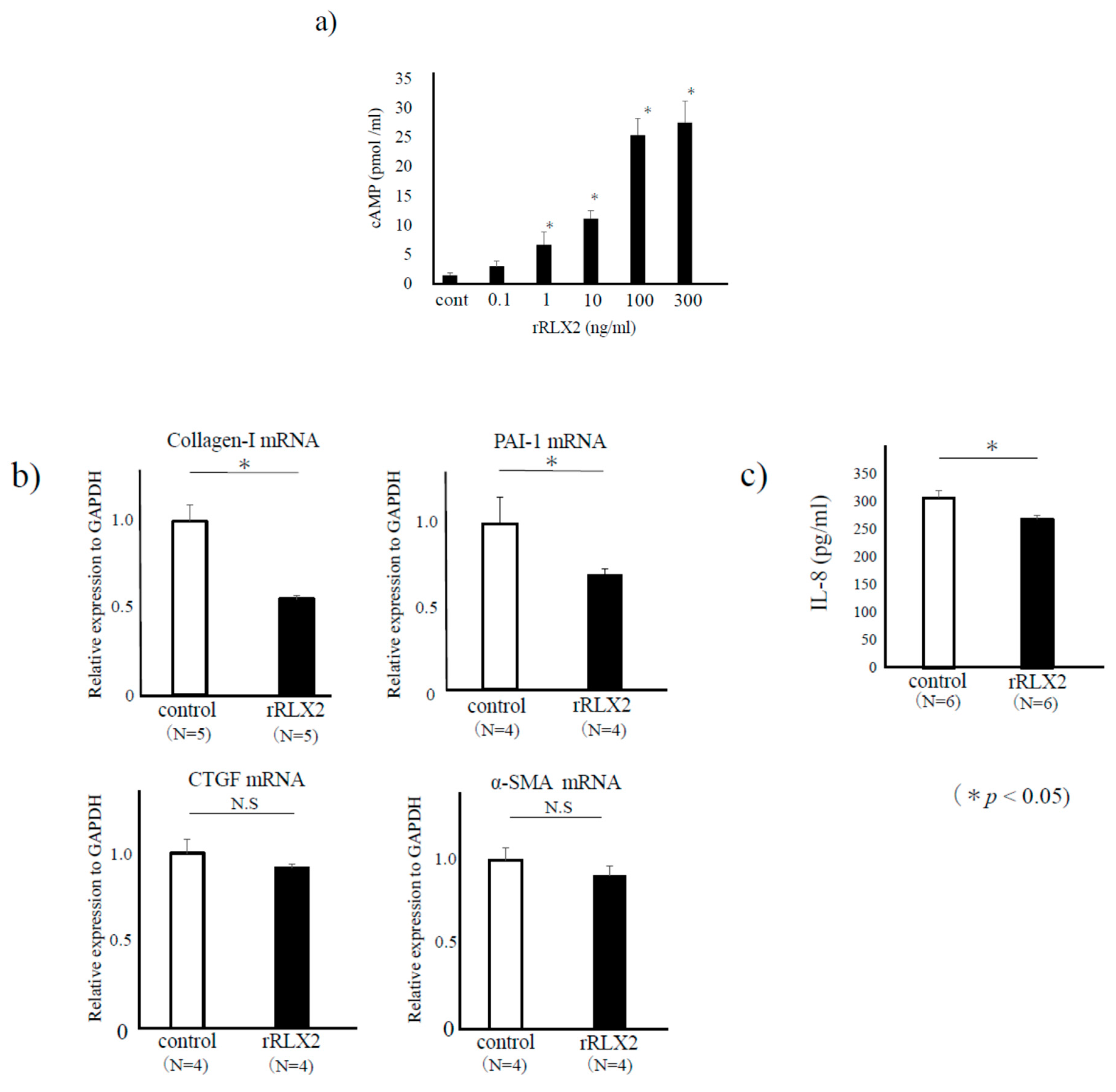
3.1. Expression of LGR-7 in Endometriotic Lesions
3.2. The Effect of RLX-2 for Endometriotic Stromal Cells (ESCs)
3.3. The Effect of RLX-2 for Collagen Gel Contractility of ESCs
3.4. Endometriosis Mouse Model (Endometrial Implantation Mouse Model)
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Laganà, A.S.; Garzon, S.; Götte, M.; Viganò, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The pathogenesis of endometriosis: Molecular and cell biology insights. Int. J. Mol. Sci. 2019, 20, 5615. [Google Scholar] [CrossRef] [PubMed]
- Zondervan, K.T.; Becker, C.M.; Koga, K.; Missmer, S.A.; Taylor, R.N.; Viganò, P. Endometriosis. Nat. Rev. Dis. Prim. 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Nnoaham, K.E.; Hummelshoj, L.; Webster, P.; d’Hooghe, T.; de Cicco Nardone, F.; de Cicco Nardone, C.; Jenkinson, C.; Kennedy, S.H.; Zondervan, K.T. World Endometriosis Research Foundation Global Study of Women’s Health consortium Impact of endometriosis on quality of life and work productivity: A multicenter study across ten countries. Fertil. Steril. 2011, 96. [Google Scholar] [CrossRef] [PubMed]
- Vigano, P.; Candiani, M.; Monno, A.; Giacomini, E.; Vercellini, P.; Somigliana, E. Time to redefine endometriosis including its pro-fibrotic nature. Hum. Reprod. 2018, 33, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β mediated SMAD signaling for the prevention of fibrosis. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Laganà, A.S.; Salmeri, F.M.; Ban Frangež, H.; Ghezzi, F.; Vrtačnik-Bokal, E.; Granese, R. Evaluation of M1 and M2 macrophages in ovarian endometriomas from women affected by endometriosis at different stages of the disease. Gynecol. Endocrinol. 2020, 36, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Viganò, P.; Ottolina, J.; Bartiromo, L.; Bonavina, G.; Schimberni, M.; Villanacci, R.; Candiani, M. Cellular Components Contributing to Fibrosis in Endometriosis: A Literature Review. J. Minim. Invasive Gynecol. 2020, 27, 287–295. [Google Scholar] [CrossRef]
- Kitajima, M.; Defrère, S.; Dolmans, M.-M.; Colette, S.; Squifflet, J.; Van Langendonckt, A.; Donnez, J. Endometriomas as a possible cause of reduced ovarian reserve in women with endometriosis. Fertil. Steril. 2011, 96, 685–691. [Google Scholar] [CrossRef]
- Kasapoglu, I.; Ata, B.; Uyaniklar, O.; Seyhan, A.; Orhan, A.; Oguz, S.Y.; Uncu, G. Endometrioma-related reduction in ovarian reserve (ERROR): A prospective longitudinal study. Fertil. Steril. 2018, 110, 122–127. [Google Scholar] [CrossRef]
- Kanai, A.J.; Konieczko, E.M.; Bennett, R.G.; Samuel, C.S.; Royce, S.G. Relaxin and fibrosis: Emerging targets, challenges, and future directions. Mol. Cell. Endocrinol. 2019, 487, 66–74. [Google Scholar] [CrossRef]
- Becker, I.; Woodley, S.J.; Stringer, M.D. The adult human pubic symphysis: A systematic review. J. Anat. 2010, 217, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, L.T.; Weiss, G. Relaxin in human pregnancy. In Annals of the New York Academy of Sciences; Blackwell Publishing Inc.: Malden, MA, USA, 2009; Volume 1160, pp. 130–135. [Google Scholar] [CrossRef]
- Morelli, S.S.; Petraglia, F.; Weiss, G.; Luisi, S.; Florio, P.; Wojtczuk, A.; Goldsmith, L.T. Endometrial expression of relaxin and relaxin receptor in endometriosis. Fertil. Steril. 2010, 94, 2885–2887. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Arai, K.; Takei, T.; Shinozaki, R.; Noguchi, M.; Fujisawa, S.; Katayama, H.; Moroder, L.; Ando, S.; Okumura, M.; Inaba, K.; et al. Characterization and optimization of two-chain folding pathways of insulin via native chain assembly. Commun. Chem. 2018, 1, 1–11. [Google Scholar] [CrossRef]
- Yoshino, O.; Osuga, Y.; Hirota, Y.; Koga, K.; Hirata, T.; Harada, M.; Morimoto, C.; Yano, T.; Nishii, O.; Tsutsumi, O.; et al. Possible pathophysiological roles of mitogen-activated protein kinases (MAPKs) in endometriosis. Am. J. Reprod. Immunol. 2004, 52, 306–311. [Google Scholar] [CrossRef]
- Yamada-Nomoto, K.; Yoshino, O.; Akiyama, I.; Iwase, A.; Ono, Y.; Nakamura, T.; Harada, M.; Nakashima, A.; Shima, T.; Ushijima, A.; et al. PAI-1 in granulosa cells is suppressed directly by statin and indirectly by suppressing TGF-β and TNF-α in mononuclear cells by insulin-sensitizing drugs. Am. J. Reprod. Immunol. 2017, 78. [Google Scholar] [CrossRef]
- Han, J.; Richter, B.; Li, Z.; Kravchenko, V.V.; Ulevitch, R.J. Molecular cloning of human p38 MAP kinase. BBA Mol. Cell Res. 1995, 1265, 224–227. [Google Scholar] [CrossRef][Green Version]
- Yuge, A.; Nasu, K.; Matsumoto, H.; Nishida, M.; Narahara, H. Collagen gel contractility is enhanced in human endometriotic stromal cells: A possible mechanism underlying the pathogenesis of endometriosis- associated fibrosis. Hum. Reprod. 2007, 22, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Hattori, K.; Ito, Y.; Honda, M.; Sekiguchi, K.; Hosono, K.; Shibuya, M.; Unno, N.; Majima, M. Lymphangiogenesis induced by vascular endothelial growth factor receptor 1 signaling contributes to the progression of endometriosis in mice. J. Pharmacol. Sci. 2020. [Google Scholar] [CrossRef]
- Becker, C.M.; Wright, R.D.; Satchi-Fainaro, R.; Funakoshi, T.; Folkman, J.; Kung, A.L.; D’Amato, R.J. A novel noninvasive model of endometriosis for monitoring the efficacy of antiangiogenic therapy. Am. J. Pathol. 2006, 168, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, K.; Ito, Y.; Hattori, K.; Inoue, T.; Hosono, K.; Honda, M.; Numao, A.; Amano, H.; Shibuya, M.; Unno, N.; et al. VEGF Receptor 1-Expressing Macrophages Recruited from Bone Marrow Enhances Angiogenesis in Endometrial Tissues. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.; Jiang, H.; Liu, X.; Guo, S.W. Enriched Environment Decelerates the Development of Endometriosis in Mouse. Reprod. Sci. 2020, 27, 1423–1435. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Pouly, J.L.; Canis, M. Dose-dependent pro- or anti-fibrotic responses of endometriotic stromal cells to interleukin-1β and tumor necrosis factor α. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Canis, M.; Darcha, C.; Dechelotte, P.; Pouly, J.L.; Bruhat, M.A. Fibrogenesis in peritoneal endometriosis. A semi-quantitative analysis of type-I collagen. Gynecol. Obstet. Investig. 1999, 47, 197–199. [Google Scholar] [CrossRef]
- Bruse, C.; Bergqvist, A.; Carlström, K.; Fianu-Jonasson, A.; Lecander, I.; Åstedt, B. Fibrinolytic factors in endometriotic tissue, endometrium, peritoneal fluid, and plasma from women with endometriosis and in endometrium and peritoneal fluid from healthy women. Fertil. Steril. 1998, 70, 821–826. [Google Scholar] [CrossRef]
- Gilabert-Estelles, J.; Castello, R.; Gilabert, J.; Ramon, L.A.; Espana, F.; Romeu, A.; Estelles, A. Plasminogen activators and plasminogen activator inhibitors in endometriosis. Front. Biosci. 2005, 10, 1162–1176. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Darcha, C. Involvement of the Wnt/β-Catenin Signaling Pathway in the Cellular and Molecular Mechanisms of Fibrosis in Endometriosis. PLoS ONE 2013, 8, e76808. [Google Scholar] [CrossRef] [PubMed]
- Arici, A. Local cytokines in endometrial tissue: The role of interleukin-8 in the pathogenesis of endometriosis. In Annals of the New York Academy of Sciences; Academy of Sciences: New York, NY, USA, 2002; Volume 955, pp. 101–109. [Google Scholar] [CrossRef]
- Koga, K.; Takemura, Y.; Osuga, Y.; Yoshino, O.; Hirota, Y.; Hirata, T.; Morimoto, C.; Harada, M.; Yano, T.; Taketani, Y. Recurrence of ovarian endometrioma after laparoscopic excision. Hum. Reprod. 2006, 21. [Google Scholar] [CrossRef]
- Chae, U.; Min, J.Y.; Kim, S.H.; Ihm, H.J.; Oh, Y.S.; Park, S.Y.; Chae, H.D.; Kim, C.H.; Kang, B.M. Decreased progesterone receptor B/A ratio in endometrial cells by tumor necrosis factor-alpha and peritoneal fluid from patients with endometriosis. Yonsei Med. J. 2016, 57, 1468–1474. [Google Scholar] [CrossRef]
- Wu, S.-P.; Li, R.; DeMayo, F.J. Progesterone Receptor Regulation of Uterine Adaptation for Pregnancy. Trends Endocrinol. Metab. 2018, 29, 481–491. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Dahoud, W.; Skomorovska-Prokvolit, Y.; Yi, L.; Liu, J.H.; Falcone, T.; Hurd, W.W.; Mesiano, S. Abundance and Localization of Progesterone Receptor Isoforms in Endometrium in Women with and Without Endometriosis and in Peritoneal and Ovarian Endometriotic Implants. Reprod. Sci. 2015, 22, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Attia, G.R.; Zeitoun, K.; Edwards, D.; Johns, A.; Carr, B.R.; Bulun, S.E. Progesterone receptor isoform A but not B is expressed in endometriosis. J. Clin. Endocrinol. Metab. 2000, 85, 2897–2902. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Cheng, Y.H.; Pavone, M.E.; Xue, Q.; Attar, E.; Trukhacheva, E.; Tokunaga, H.; Utsunomiya, H.; Yin, P.; Luo, X.; et al. Estrogen receptor-β, Estrogen receptor-α, and progesterone resistance in endometriosis. Semin. Reprod. Med. 2010, 28, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Bathgate, R.A.; Ivell, R.; Sanborn, B.M.; Sherwood, O.D.; Summers, R.J. International union of pharmacology LVII: Recommendations for the nomenclature of receptors for relaxin family peptides. Pharmacol. Rev. 2006, 58, 7–31. [Google Scholar] [CrossRef]
- Sheau, Y.H.; Nakabayashi, K.; Nishi, S.; Kumagai, J.; Kudo, M.; Sherwood, O.D.; Hsueh, A.J.W. Activation of orphan receptors by the hormone relaxin. Science 2002, 295, 671–674. [Google Scholar] [CrossRef]
- Stovall, D.W.; Anners, J.A.; Halme, J. Immunohistochemical detection of type, I, III, and IV collagen in endometriosis implants. Fertil. Steril. 1992, 57, 984–989. [Google Scholar] [CrossRef]
- Mulayim, N.; Savlu, A.; Guzeloglu-Kayisli, O.; Kayisli, U.A.; Arici, A. Regulation of endometrial stromal cell matrix metalloproteinase activity and invasiveness by interleukin-8. Fertil. Steril. 2004, 81 (Suppl. 1), 904–911. [Google Scholar] [CrossRef]
- Selam, B.; Kayisli, U.A.; Garcia-Velasco, J.A.; Eda Akbas, G.; Arici, A. Regulation of Fas ligand expression by IL-8 in human endometrium. J. Clin. Endocrinol. Metab. 2002, 87, 3921–3927. [Google Scholar] [CrossRef][Green Version]
- Jonas, O.; Duschl, C. Force propagation and force generation in cells. Cytoskeleton 2010, 67, 555–563. [Google Scholar] [CrossRef]
- Yoshino, O.; Osuga, Y.; Koga, K.; Hirota, Y.; Hirata, T.; Ruimeng, X.; Na, L.; Yano, T.; Tsutsumi, O.; Taketani, Y. FR 167653, a p38 mitogen-activated protein kinase inhibitor, suppresses the development of endometriosis in a murine model. J. Reprod. Immunol. 2006, 72. [Google Scholar] [CrossRef]
- Zhou, W.D.; Yang, H.M.; Wang, Q.; Su, D.Y.; Liu, F.A.; Zhao, M.; Chen, Q.H.; Chen, Q.X. SB203580, a p38 mitogen-activated protein kinase inhibitor, suppresses the development of endometriosis by down-regulating proinflammatory cytokines and proteolytic factors in a mouse model. Hum. Reprod. 2010, 25, 3110–3116. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, A.; Haluska, P.; Tolcher, A.W.; Erlichman, C.; Papadopoulos, K.P.; Lensing, J.L.; Beeram, M.; Molina, J.R.; Rasco, D.W.; Arcos, R.R.; et al. A first-in-human phase i study of the oral p38 MAPK inhibitor, ralimetinib (LY2228820 Dimesylate), in patients with advanced cancer. Clin. Cancer Res. 2016, 22, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, O.; Osuga, Y.; Hirota, Y.; Koga, K.; Hirata, T.; Yano, T.; Ayabe, T.; Tsutsumi, O.; Taketani, Y. Endometrial stromal cells undergoing decidualization down-regulate their properties to produce proinflammatory cytokines in response to interleukin-1β via reduced p38 mitogen-activated protein kinase phosphorylation. J. Clin. Endocrinol. Metab. 2003, 88. [Google Scholar] [CrossRef] [PubMed]
- Metra, M.; Teerlink, J.R.; Cotter, G.; Davison, B.A.; Felker, G.M.; Filippatos, G.; Greenberg, B.H.; Pang, P.S.; Ponikowski, P.; Voors, A.A.; et al. Effects of serelaxin in patients with acute heart failure. N. Engl. J. Med. 2019, 381, 716–726. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshino, O.; Ono, Y.; Honda, M.; Hattori, K.; Sato, E.; Hiraoka, T.; Ito, M.; Kobayashi, M.; Arai, K.; Katayama, H.; et al. Relaxin-2 May Suppress Endometriosis by Reducing Fibrosis, Scar Formation, and Inflammation. Biomedicines 2020, 8, 467. https://doi.org/10.3390/biomedicines8110467
Yoshino O, Ono Y, Honda M, Hattori K, Sato E, Hiraoka T, Ito M, Kobayashi M, Arai K, Katayama H, et al. Relaxin-2 May Suppress Endometriosis by Reducing Fibrosis, Scar Formation, and Inflammation. Biomedicines. 2020; 8(11):467. https://doi.org/10.3390/biomedicines8110467
Chicago/Turabian StyleYoshino, Osamu, Yosuke Ono, Masako Honda, Kyoko Hattori, Erina Sato, Takehiro Hiraoka, Masami Ito, Mutsumi Kobayashi, Kenta Arai, Hidekazu Katayama, and et al. 2020. "Relaxin-2 May Suppress Endometriosis by Reducing Fibrosis, Scar Formation, and Inflammation" Biomedicines 8, no. 11: 467. https://doi.org/10.3390/biomedicines8110467
APA StyleYoshino, O., Ono, Y., Honda, M., Hattori, K., Sato, E., Hiraoka, T., Ito, M., Kobayashi, M., Arai, K., Katayama, H., Tsuchida, H., Yamada-Nomoto, K., Iwahata, S., Fukushi, Y., Wada, S., Iwase, H., Koga, K., Osuga, Y., Iwaoka, M., & Unno, N. (2020). Relaxin-2 May Suppress Endometriosis by Reducing Fibrosis, Scar Formation, and Inflammation. Biomedicines, 8(11), 467. https://doi.org/10.3390/biomedicines8110467