Current Status of In Vitro Models and Assays for Susceptibility Testing for Wound Biofilm Infections




Abstract

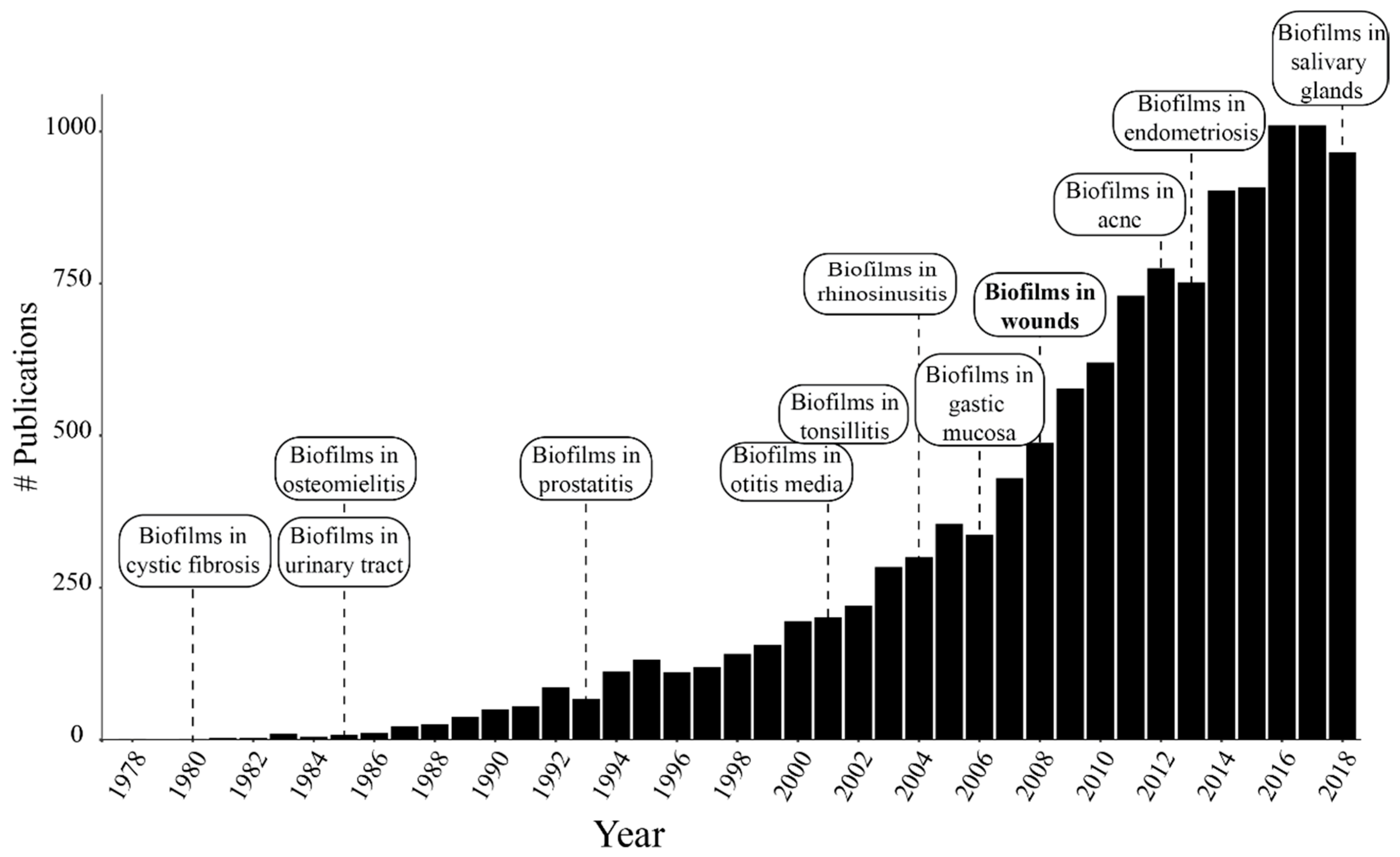
1. Introduction
1.1. Biofilms in Chronic Wounds and Other Chronic Infections
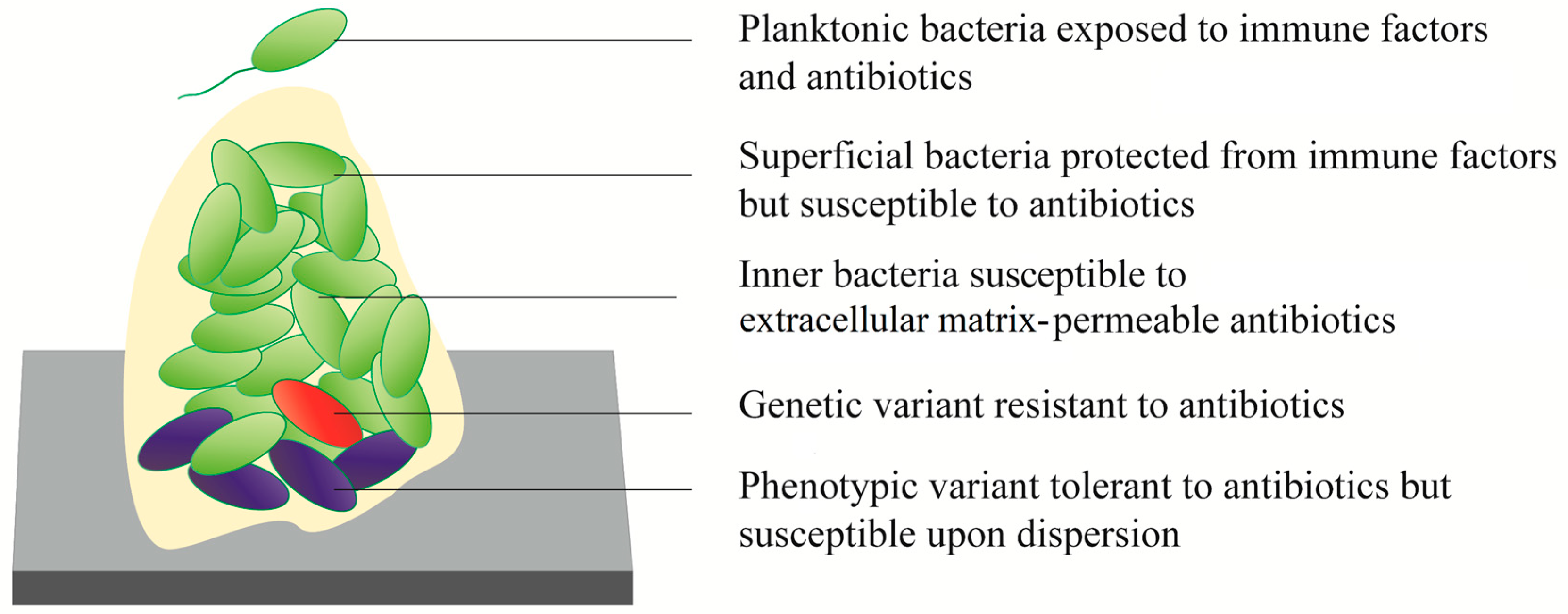
1.2. Mechanism of Biofilm Resistance
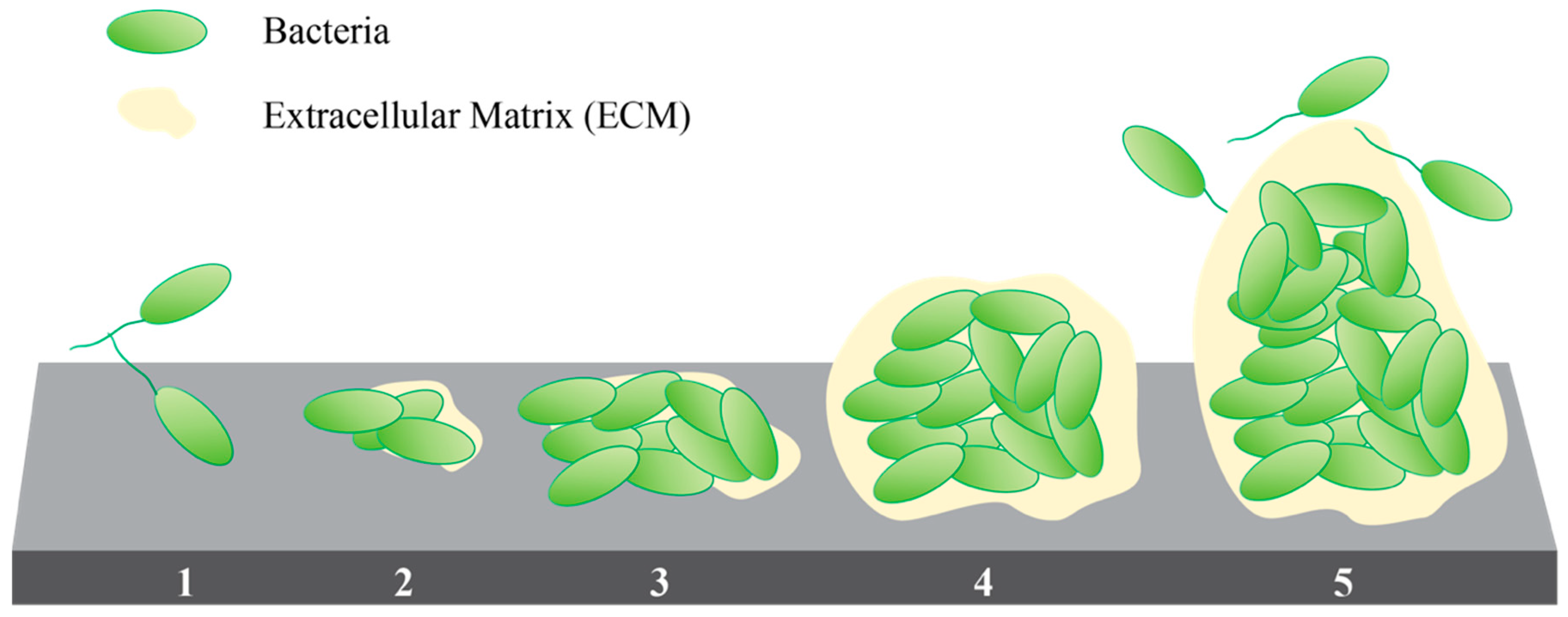
1.2.1. Extracellular Matrix (ECM)
1.2.2. Phenotypic Tolerance
1.2.3. Genetic Resistance
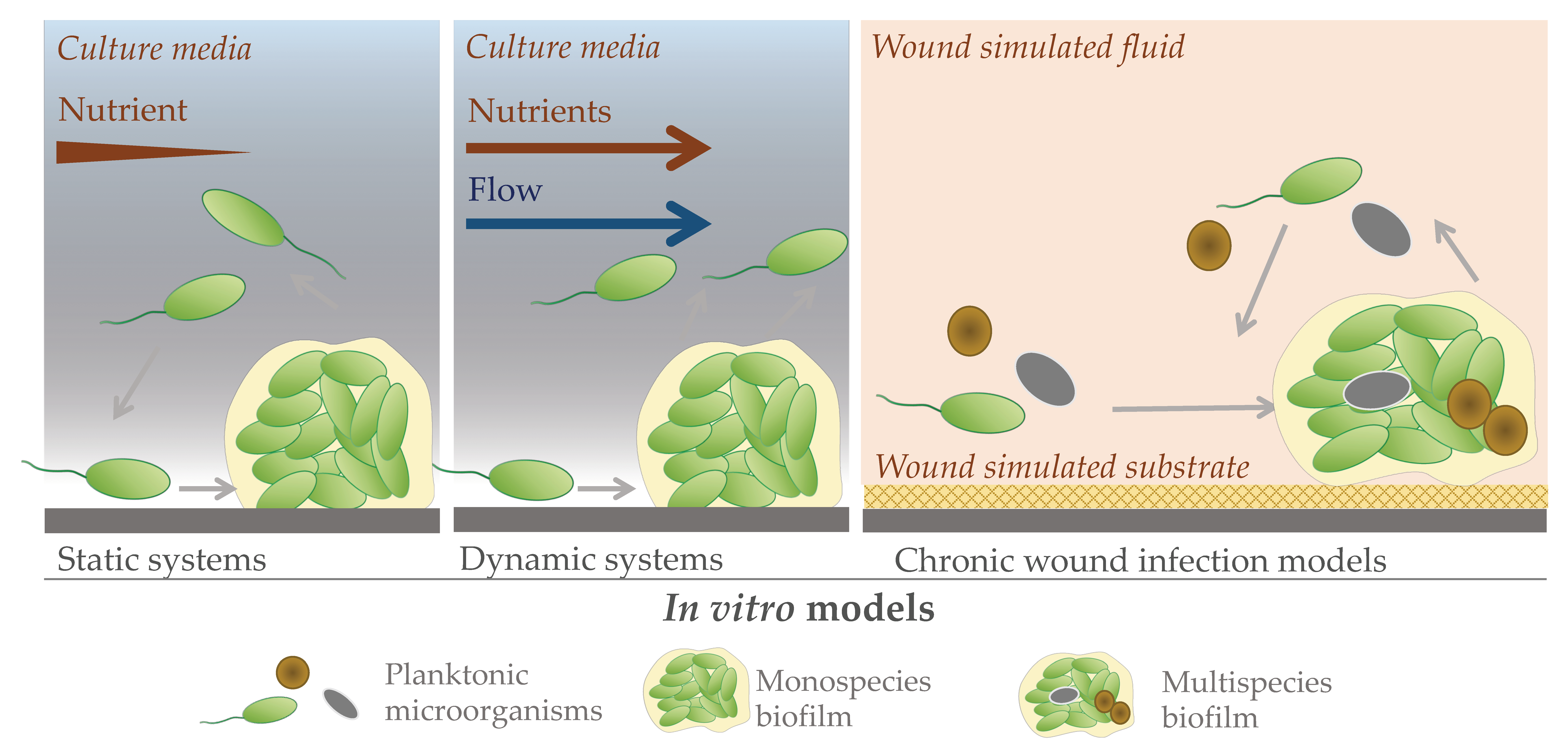
2. In Vitro Models to Study Chronic Wound Biofilms
2.1. Static Models
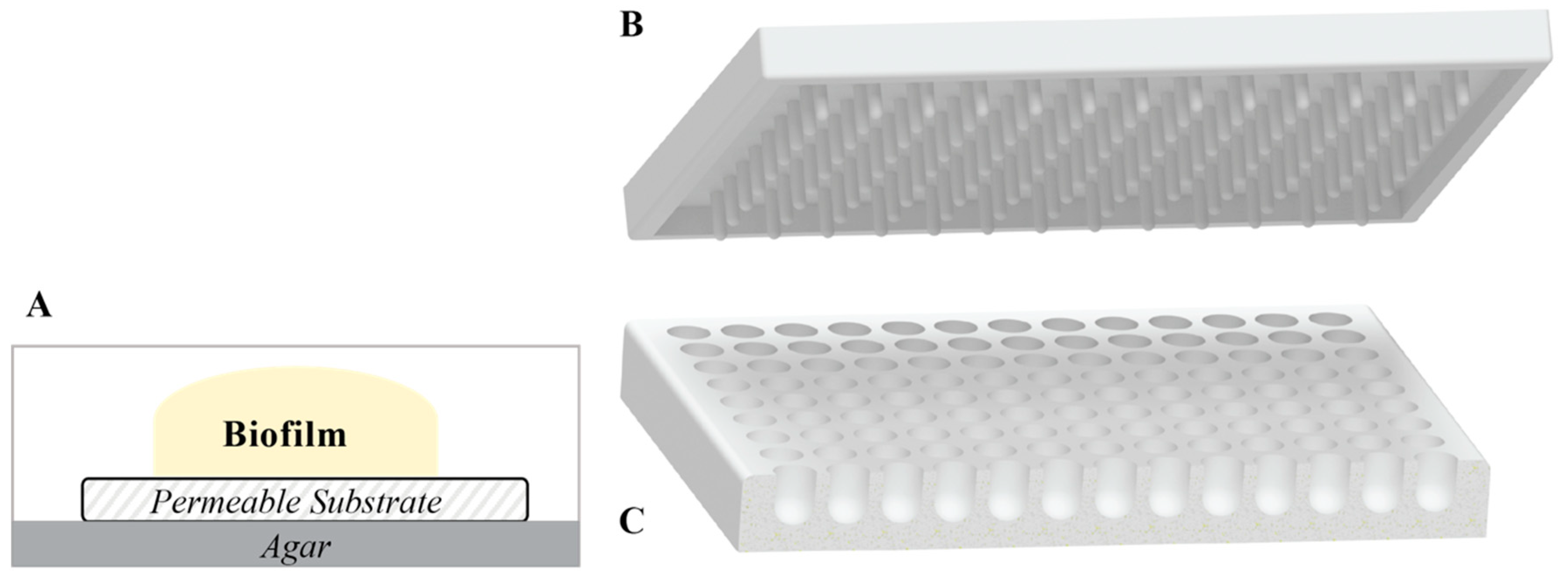
2.1.1. Agar Plates
2.1.2. Colony Biofilms
2.1.3. Microtiter Plates
2.1.4. Calgary Biofilm Device
2.2. Dynamic Models
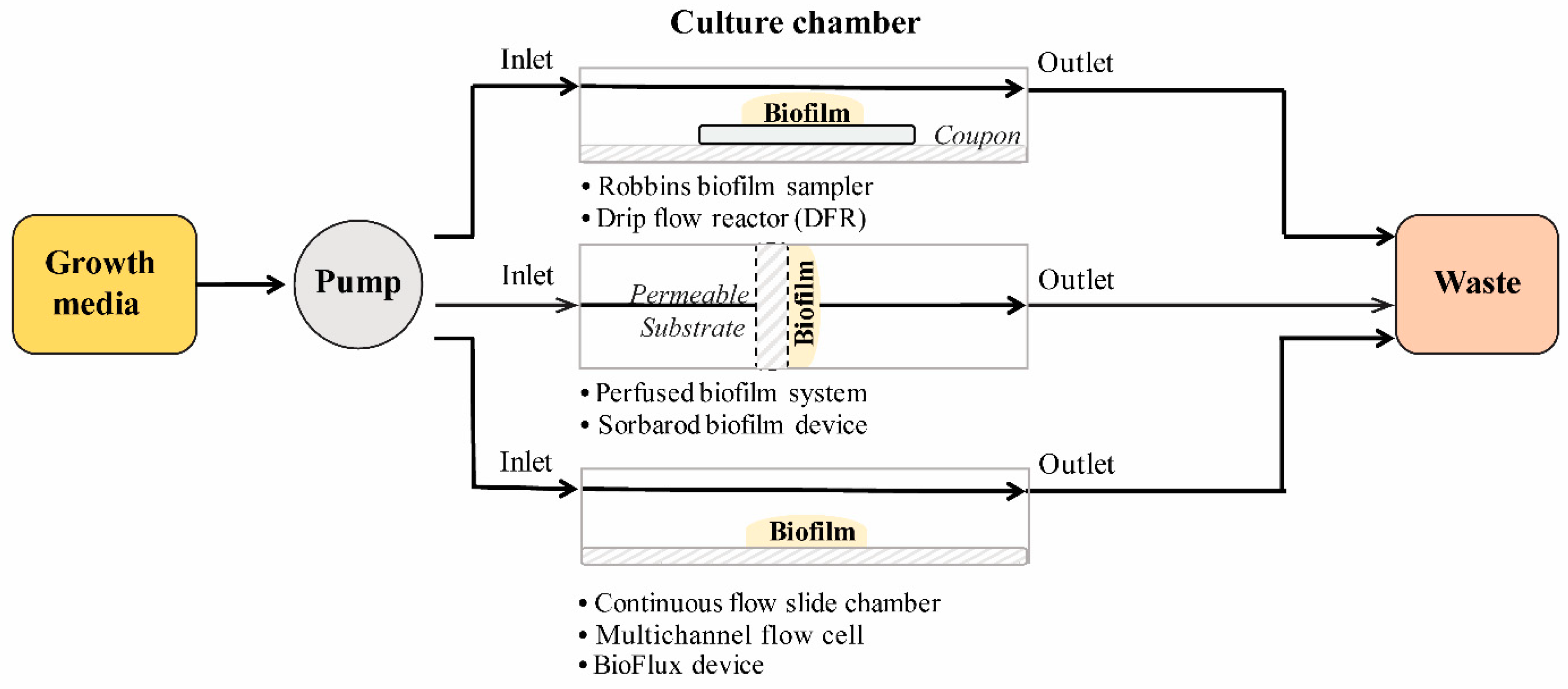
2.2.1. Flow Cell Systems
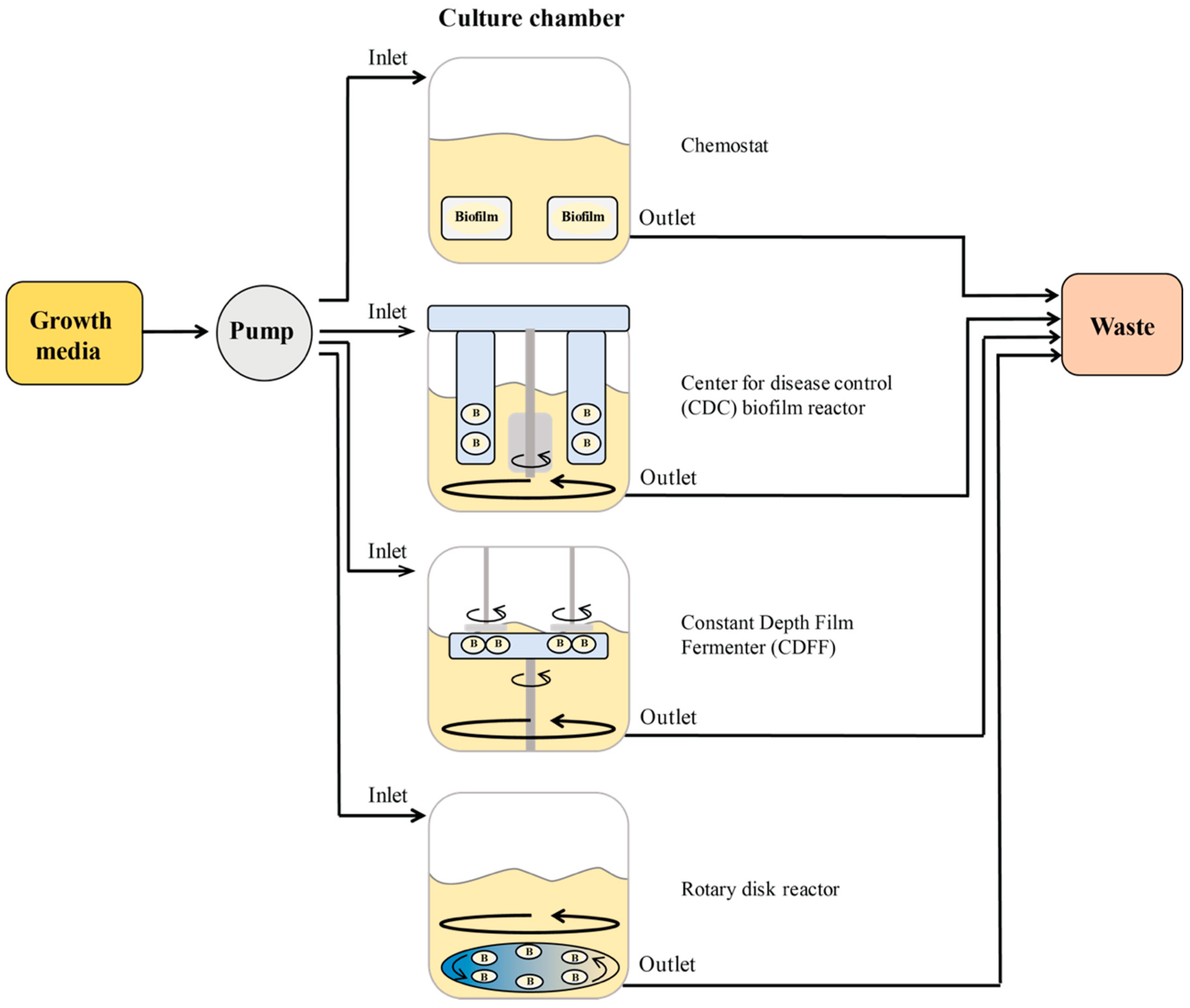
2.2.2. Biofilm Reactors
2.3. Advances in In Vitro Biofilm Models
In Vitro P. aeruginosa Biofilm Wound Infection Models
2.4. Summary of In Vitro Model Systems
3. Evaluation of In Vitro Models
3.1. Viability Determination by Colony Counting and Cell Staining
3.1.1. Colony Forming Units
3.1.2. SYTO 9/PI (Live/Dead BacLight Dye)
3.1.3. Acridine Orange
3.2. Metabolic Activity
3.2.1. Tetrazolium Salts (INT, TTC, CTC, MTT, & XTT)
3.2.2. Resazurin (Alamar Blue, PrestoBlue, & CellTiter-Blue)
3.2.3. Bioluminescence
3.2.4. Fluorescein Diacetate (FDA)
3.3. Biofilm Biomass and Molecular Stains
3.3.1. Total Biomass: Crystal Violet and Congo Red
3.3.2. Cell Components: Safranin and Calcofluor White
3.3.3. Proteins: SYPRO Ruby and FITC
3.3.4. Polysaccharides: Concanavalin A, Periodic Acid–Schiff, and van Gieson
3.3.5. Lipids: DiD
3.4. Turbidity
3.5. Biofilm Structure
3.5.1. Fluorescence Microscopy
3.5.2. Raman Microscopy
3.5.3. Electron Microscopy
3.6. Mechanical Stability
3.7. Zone of Inhibition
3.8. Summary of In Vitro Assays
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Geesey, G.; Mutch, R.; Costerton, J.T.; Green, R. Sessile bacteria: An important component of the microbial population in small mountain streams. Limnol. Oceanogr. 1978, 23, 1214–1223. [Google Scholar] [CrossRef]
- Costerton, J.W.; Geesey, G.; Cheng, K.-J. How bacteria stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Geesey, G.; Richardson, W.; Yeomans, H.; Irvin, R.; Costerton, J. Microscopic examination of natural sessile bacterial populations from an alpine stream. Can. J. Microbiol. 1977, 23, 1733–1736. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.; Bradshaw, D. Dental plaque as a biofilm. J. Ind. Microbiol. 1995, 15, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Gest, H. The discovery of microorganisms by Robert Hooke and Antoni Van Leeuwenhoek, fellows of the Royal Society. Notes Rec. R. Soc. 2004, 58, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.; Chan, R.; Lam, K.; Costerton, J. Production of mucoid microcolonies by Pseudomonas aeruginosa within infected lungs in cystic fibrosis. Infect. Immun. 1980, 28, 546–556. [Google Scholar]
- Nickel, J.C.; Emtage, J.; Costerton, J.W. Ultrastructural microbial ecology of infection-induced urinary stones. J. Urol. 1985, 133, 622–627. [Google Scholar] [CrossRef]
- Gristina, A.G.; Webb, L.X.; Hobgood, C.D.; Oga, M. Adherent bacterial colonization in the pathogenesis of osteomyelitis. Science 1985, 228, 990–994. [Google Scholar] [CrossRef]
- Nickel, J.C.; Costerton, J.W. Bacterial localization in antibiotic-refractory chronic bacterial prostatitis. Prostate 1993, 23, 107–114. [Google Scholar] [CrossRef]
- Post, J.C. Candidate’s Thesis: Direct evidence of bacterial biofilms in otitis media. Laryngoscope 2001, 111, 2083–2094. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Hu, F.Z.; Gieseke, A.; Nistico, L.; Nguyen, D.; Hayes, J.; Forbes, M.; Greenberg, D.P.; Dice, B.; Burrows, A. Direct detection of bacterial biofilms on the middle-ear mucosa of children with chronic otitis media. JAMA 2006, 296, 202–211. [Google Scholar] [CrossRef]
- Chen, C. Periodontitis as a biofilm infection. J. Calif. Dent. Assoc. 2001, 29, 362–369. [Google Scholar]
- Chole, R.A.; Faddis, B.T. Anatomical evidence of microbial biofilms in tonsillar tissues: A possible mechanism to explain chronicity. Arch. Otolaryngol.-Head Neck Surg. 2003, 129, 634–636. [Google Scholar] [CrossRef]
- Cryer, J.; Schipor, I.; Perloff, J.R.; Palmer, J.N. Evidence of bacterial biofilms in human chronic sinusitis. ORL 2004, 66, 155–158. [Google Scholar] [CrossRef]
- Capon, M.A.; Tran, T.R.; Sugawa, C.; Coticchia, J.M. Identification of Helicobacter pylori biofilms in human gastric mucosa. J. Gastrointest. Surg. 2006, 10, 712–717. [Google Scholar]
- Kirketerp-Møller, K.; Jensen, P.Ø.; Fazli, M.; Madsen, K.G.; Pedersen, J.; Moser, C.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Distribution, organization, and ecology of bacteria in chronic wounds. J. Clin. Microbiol. 2008, 46, 2717–2722. [Google Scholar] [CrossRef]
- Jahns, A.C.; Lundskog, B.; Ganceviciene, R.; Palmer, R.H.; Golovleva, I.; Zouboulis, C.C.; McDowell, A.; Patrick, S.; Alexeyev, O.A. An increased incidence of Propionibacterium acnes biofilms in acne vulgaris: A case-control study. Br. J. Dermatol. 2012, 167, 50–58. [Google Scholar] [CrossRef]
- Swidsinski, A.; Verstraelen, H.; Loening-Baucke, V.; Swidsinski, S.; Mendling, W.; Halwani, Z. Presence of a polymicrobial endometrial biofilm in patients with bacterial vaginosis. PLoS ONE 2013, 8, e53997. [Google Scholar] [CrossRef]
- Schrøder, S.A.; Eickhardt, S.; Bjarnsholt, T.; Nørgaard, T.; Homøe, P. Morphological evidence of biofilm in chronic obstructive sialadenitis. J. Laryngol. Otol. 2018, 132, 611–614. [Google Scholar] [CrossRef]
- Zhao, G.; Usui, M.L.; Lippman, S.I.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Biofilms and inflammation in chronic wounds. Adv. Wound Care 2013, 2, 389–399. [Google Scholar] [CrossRef]
- Cochrane, D.M.; Brown, M.; Anwar, H.; Weller, P.; Lam, K.; Costerton, J. Antibody response to Pseudomonas aeruginosa surface protein antigens in a rat model of chronic lung infection. J. Med. Microbiol. 1988, 27, 255–261. [Google Scholar] [CrossRef]
- Jahoor, A.; Patel, R.; Bryan, A.; Do, C.; Krier, J.; Watters, C.; Wahli, W.; Li, G.; Williams, S.C.; Rumbaugh, K.P. Peroxisome Proliferator-Activated Receptors Mediate Host Cell Proinflammatory Responses to Pseudomonas aeruginosa Autoinducer. J. Bacteriol. 2008, 190, 4408–4415. [Google Scholar] [CrossRef] [PubMed]
- Leid, J.G.; Willson, C.J.; Shirtliff, M.E.; Hassett, D.J.; Parsek, M.R.; Jeffers, A.K. The Exopolysaccharide Alginate Protects Pseudomonas aeruginosa Biofilm Bacteria from IFN-γ-Mediated Macrophage Killing. J. Immunol. 2005, 175, 7512–7518. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Rhoads, D.D.; Dowd, S.E. Biofilms and chronic wound inflammation. J. Wound Care 2008, 17, 333–341. [Google Scholar] [CrossRef]
- Stadelmann, W.K.; Digenis, A.G.; Tobin, G.R. Physiology and healing dynamics of chronic cutaneous wounds. Am. J. Surg. 1998, 176, 26S–38S. [Google Scholar] [CrossRef]
- Sen, C.K.; Gordillo, G.M.; Roy, S.; Kirsner, R.; Lambert, L.; Hunt, T.K.; Gottrup, F.; Gurtner, G.C.; Longaker, M.T. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair Regen. 2009, 17, 763–771. [Google Scholar] [CrossRef]
- Gould, L.; Abadir, P.; Brem, H.; Carter, M.; Conner-Kerr, T.; Davidson, J.; DiPietro, L.; Falanga, V.; Fife, C.; Gardner, S. Chronic wound repair and healing in older adults: Current status and future research. Wound Repair Regen. 2015, 23, 1–13. [Google Scholar] [CrossRef]
- Cutting, K.F. Wound exudate: Composition and functions. Br. J. Community Nurs. 2003, 8, 4–9. [Google Scholar] [CrossRef]
- Reiber, G.E.; Boyko, E.J.; Smith, D.G. Lower extremity foot ulcers and amputations in diabetes. Diabetes Am. 1995, 2, 409–427. [Google Scholar]
- Gjødsbøl, K.; Christensen, J.J.; Karlsmark, T.; Jørgensen, B.; Klein, B.M.; Krogfelt, K.A. Multiple bacterial species reside in chronic wounds: A longitudinal study. Int. Wound J. 2006, 3, 225–231. [Google Scholar] [CrossRef]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Hidron, A.; Edwards, J.; Patel, J.; Horan, T.; Sievert, D.; Pollock, D.; Fridkin, S. Antimicrobial-resistant pathogens associated with healthcare-associated infections: Annual summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006–2007. Infect. Control. Hosp. Epidemiol. 2008, 29, 996–1011. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Priority pathogens list for R&D of new antibiotics (27 February 2017). Available online: https://www.who.int/en/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 29 April 2019).
- Khan, H.A.; Ahmad, A.; Mehboob, R. Nosocomial infections and their control strategies. Asian Pac. J. Trop. Biomed. 2015, 5, 509–514. [Google Scholar] [CrossRef]
- Roche, E.D.; Renick, P.J.; Tetens, S.P.; Ramsay, S.J.; Daniels, E.Q.; Carson, D.L. Increasing the presence of biofilm and healing delay in a porcine model of MRSA-infected wounds. Wound Repair Regen. 2012, 20, 537–543. [Google Scholar] [CrossRef]
- Metcalf, D.G.; Bowler, P.G. Biofilm delays wound healing: A review of the evidence. Burn. Trauma 2013, 1, 5. [Google Scholar] [CrossRef]
- Serralta, V.W.; Harrison-Balestra, C.; Cazzaniga, A.L.; Davis, S.C.; Mertz, P.M. Lifestyles of bacteria in wounds: Presence of biofilms? Wounds 2001, 13, 29–34. [Google Scholar]
- Percival, S.; Bowler, P. Understanding the effects of bacterial communities and biofilms on wound healing. World Wide Wounds 2004, 1, 1–5. [Google Scholar]
- Bjarnsholt, T.; Kirketerp-Møller, K.; Jensen, P.Ø.; Madsen, K.G.; Phipps, R.; Krogfelt, K.; Høiby, N.; Givskov, M. Why chronic wounds will not heal: A novel hypothesis. Wound Repair Regen. 2008, 16, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.; Séguin, D.L.; Asselin, A.-E.; Déziel, E.; Cantin, A.M.; Frost, E.H.; Michaud, S.; Malouin, F. Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide. BMC Microbiol. 2010, 10, 33. [Google Scholar] [CrossRef]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an in vitro wound model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef]
- Pastar, I.; Nusbaum, A.G.; Gil, J.; Patel, S.B.; Chen, J.; Valdes, J.; Stojadinovic, O.; Plano, L.R.; Tomic-Canic, M.; Davis, S.C. Interactions of methicillin resistant Staphylococcus aureus USA300 and Pseudomonas aeruginosa in polymicrobial wound infection. PLoS ONE 2013, 8, e56846. [Google Scholar] [CrossRef]
- Fazli, M.; Bjarnsholt, T.; Kirketerp-Møller, K.; Jørgensen, B.; Andersen, A.S.; Krogfelt, K.A.; Givskov, M.; Tolker-Nielsen, T. Nonrandom distribution of Pseudomonas aeruginosa and Staphylococcus aureus in chronic wounds. J. Clin. Microbiol. 2009, 47, 4084–4089. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti Infect. 2015, 13, 605–613. [Google Scholar] [CrossRef]
- Zhao, G.; Hochwalt, P.C.; Usui, M.L.; Underwood, R.A.; Singh, P.K.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Delayed wound healing in diabetic (db/db) mice with Pseudomonas aeruginosa biofilm challenge: A model for the study of chronic wounds. Wound Repair Regen. 2010, 18, 467–477. [Google Scholar] [CrossRef]
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I.M. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 1: Normal and chronic wounds: Biology, causes, and approaches to care. Adv. Skin Wound Care 2012, 25, 304–314. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Rhoads, D.D. A study of biofilm-based wound management in subjects with critical limb ischaemia. J. Wound Care 2008, 17, 145–148, 150–152, 154–155. [Google Scholar] [CrossRef]
- Nixon, G.M.; Armstrong, D.S.; Carzino, R.; Carlin, J.B.; Olinsky, A.; Robertson, C.F.; Grimwood, K. Clinical outcome after early Pseudomonas aeruginosa infection in cystic fibrosis. J. Pediatr. 2001, 138, 699–704. [Google Scholar] [CrossRef]
- Rubin, B.K. Mucus structure and properties in cystic fibrosis. Paediatr. Respir. Rev. 2007, 8, 4–7. [Google Scholar] [CrossRef]
- Costerton, J.W. Cystic fibrosis pathogenesis and the role of biofilms in persistent infection. Trends Microbiol. 2001, 9, 50–52. [Google Scholar] [CrossRef]
- Murphy, T.F.; Brauer, A.L.; Eschberger, K.; Lobbins, P.; Grove, L.; Cai, X.; Sethi, S. Pseudomonas aeruginosa in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2008, 177, 853–860. [Google Scholar] [CrossRef]
- De Soyza, A.; Winstanley, C. Pseudomonas aeruginosa and Bronchiectasis. In Bronchiectasis; Springer: Berlin/Heidelberg, Germany, 2018; pp. 157–180. [Google Scholar]
- Psaltis, A.J.; Ha, K.R.; Beule, A.G.; Tan, L.W.; Wormald, P.J. Confocal scanning laser microscopy evidence of biofilms in patients with chronic rhinosinusitis. Laryngoscope 2007, 117, 1302–1306. [Google Scholar] [CrossRef]
- Mittal, R.; Aggarwal, S.; Sharma, S.; Chhibber, S.; Harjai, K. Urinary tract infections caused by Pseudomonas aeruginosa: A minireview. J. Infect. Public Health 2009, 2, 101–111. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Fact Sheet: Antimicrobial Resistance. Available online: http://www.who.int/mediacentre/factsheets/fs194/en/ (accessed on 11 October 2017).
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef]
- De Kievit, T.R.; Gillis, R.; Marx, S.; Brown, C.; Iglewski, B.H. Quorum-Sensing Genes in Pseudomonas aeruginosa Biofilms: Their Role and Expression Patterns. Appl. Environ. Microbiol. 2001, 67, 1865–1873. [Google Scholar] [CrossRef]
- Shrout, J.D.; Chopp, D.L.; Just, C.L.; Hentzer, M.; Givskov, M.; Parsek, M.R. The impact of quorum sensing and swarming motility on Pseudomonas aeruginosa biofilm formation is nutritionally conditional. Mol. Microbiol. 2006, 62, 1264–1277. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef]
- Kim, H.-S.; Lee, S.-H.; Byun, Y.; Park, H.-D. 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Sci. Rep. 2015, 5, 8656. Available online: https://www.nature.com/articles/srep08656#supplementary-information (accessed on 29 April 2019). [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605. [Google Scholar] [CrossRef]
- Riedel, K.; Hentzer, M.; Geisenberger, O.; Huber, B.; Steidle, A.; Wu, H.; Høiby, N.; Givskov, M.; Molin, S.; Eberl, L. N-acylhomoserine-lactone-mediated communication between Pseudomonas aeruginosa and Burkholderia cepacia in mixed biofilms. Microbiology 2001, 147, 3249–3262. [Google Scholar] [CrossRef]
- Driffield, K.; Miller, K.; Bostock, J.M.; O’Neill, A.J.; Chopra, I. Increased mutability of Pseudomonas aeruginosa in biofilms. J. Antimicrob. Chemother. 2008, 61, 1053–1056. [Google Scholar] [CrossRef]
- Arvidson, S. Extracellular enzymes. In Gram-Positive Pathogens, 2nd ed.; Fischetti, V., Novick, R., Ferretti, J., Portnoy, D., Rood, J., Eds.; ASM Press: Washington, DC, USA, 2006; pp. 478–485. [Google Scholar] [CrossRef]
- Chiang, W.C.; Nilsson, M.; Jensen, P.O.; Hoiby, N.; Nielsen, T.E.; Givskov, M.; Tolker-Nielsen, T. Extracellular DNA shields against aminoglycosides in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2013, 57, 2352–2361. [Google Scholar] [CrossRef]
- Khan, W.; Bernier, S.P.; Kuchma, S.L.; Hammond, J.H.; Hasan, F.; O’Toole, G.A. Aminoglycoside resistance of Pseudomonas aeruginosa biofilms modulated by extracellular polysaccharide. Int. Microbiol. 2010, 13, 207–212. [Google Scholar] [PubMed]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Rachid, S.; Ohlsen, K.; Witte, W.; Hacker, J.; Ziebuhr, W. Effect of Subinhibitory Antibiotic Concentrations on Polysaccharide Intercellular Adhesin Expression in Biofilm-Forming Staphylococcus epidermidis. Antimicrob. Agents Chemother. 2000, 44, 3357–3363. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Ghafoor, A.; Hay, I.D.; Rehm, B.H. Role of exopolysaccharides in Pseudomonas aeruginosa biofilm formation and architecture. Appl. Environ. Microbiol. 2011, 77, 5238–5246. [Google Scholar] [CrossRef]
- Lee, B.; Schjerling, C.K.; Kirkby, N.; Hoffmann, N.; Borup, R.; Molin, S.; Høiby, N.; Ciofu, O. Mucoid Pseudomonas aeruginosa isolates maintain the biofilm formation capacity and the gene expression profiles during the chronic lung infection of CF patients. Apmis 2011, 119, 263–274. [Google Scholar] [CrossRef]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.; Parsek, M.R. The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa. Plos Pathog. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Jennings, L.K.; Storek, K.M.; Ledvina, H.E.; Coulon, C.; Marmont, L.S.; Sadovskaya, I.; Secor, P.R.; Tseng, B.S.; Scian, M.; Filloux, A. Pel is a cationic exopolysaccharide that cross-links extracellular DNA in the Pseudomonas aeruginosa biofilm matrix. Proc. Natl. Acad. Sci. USA 2015, 112, 11353–11358. [Google Scholar] [CrossRef]
- Muto, Y.; Goto, S. Transformation by extracellular DNA produced by Pseudomonas aeruginosa. Microbiol. Immunol. 1986, 30, 621–628. [Google Scholar] [CrossRef]
- Rani, S.A.; Pitts, B.; Beyenal, H.; Veluchamy, R.A.; Lewandowski, Z.; Davison, W.M.; Buckingham-Meyer, K.; Stewart, P.S. Spatial patterns of DNA replication, protein synthesis, and oxygen concentration within bacterial biofilms reveal diverse physiological states. J. Bacteriol. 2007, 189, 4223–4233. [Google Scholar] [CrossRef] [PubMed]
- Borriello, G.; Werner, E.; Roe, F.; Kim, A.M.; Ehrlich, G.D.; Stewart, P.S. Oxygen limitation contributes to antibiotic tolerance of Pseudomonas aeruginosa in biofilms. Antimicrob. Agents Chemother. 2004, 48, 2659–2664. [Google Scholar] [CrossRef]
- Werner, E.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Heydorn, A.; Molin, S.; Pitts, B.; Stewart, P.S. Stratified growth in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2004, 70, 6188–6196. [Google Scholar] [CrossRef]
- Wessel, A.K.; Arshad, T.A.; Fitzpatrick, M.; Connell, J.L.; Bonnecaze, R.T.; Shear, J.B.; Whiteley, M. Oxygen Limitation within a Bacterial Aggregate. Mbio 2014, 5, e00992. [Google Scholar] [CrossRef]
- Percival, S.L.; Hill, K.E.; Malic, S.; Thomas, D.W.; Williams, D.W. Antimicrobial tolerance and the significance of persister cells in recalcitrant chronic wound biofilms. Wound Repair Regen. 2011, 19, 1–9. [Google Scholar] [CrossRef]
- Conlon, B.P.; Rowe, S.E.; Lewis, K. Persister cells in biofilm associated infections. Adv. Exp. Med. Biol 2015, 831, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Walters, M.C.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418. [Google Scholar] [CrossRef]
- Livermore, D.M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Ross, S.S.; Fiegel, J. Nutrient dispersion enhances conventional antibiotic activity against Pseudomonas aeruginosa biofilms. Int. J. Antimicrob. Agents 2012, 40, 177–181. [Google Scholar] [CrossRef]
- Lamppa, J.W.; Griswold, K.E. Alginate lyase exhibits catalysis-independent biofilm dispersion and antibiotic synergy. Antimicrob. Agents Chemother. 2013, 57, 137–145. [Google Scholar] [CrossRef]
- Roizman, D.; Vidaillac, C.; Givskov, M.; Yang, L. In Vitro Evaluation of Biofilm Dispersal as a Therapeutic Strategy To Restore Antimicrobial Efficacy. Antimicrob. Agents Chemother. 2017, 61, e01088-17. [Google Scholar] [CrossRef]
- McBain, A.J. In Vitro Biofilm Models. In Advances in Applied Microbiology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; Volume 69, pp. 99–132. [Google Scholar]
- Kovach, K.; Davis-Fields, M.; Irie, Y.; Jain, K.; Doorwar, S.; Vuong, K.; Dhamani, N.; Mohanty, K.; Touhami, A.; Gordon, V.D. Evolutionary adaptations of biofilms infecting cystic fibrosis lungs promote mechanical toughness by adjusting polysaccharide production. Biofilms Microbiomes 2017, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Billings, N.; Birjiniuk, A.; Samad, T.S.; Doyle, P.S.; Ribbeck, K. Material properties of biofilms-a review of methods for understanding permeability and mechanics. Rep. Prog. Phys. 2015, 78, 036601. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.-F.; Pitts, B.; Pellock, B.; Walker, G.C.; Stewart, P.S.; O’Toole, G.A. A genetic basis for Pseudomonas aeruginosa biofilm antibiotic resistance. Nature 2003, 426, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Anderl, J.N.; Franklin, M.J.; Stewart, P.S. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob. Agents Chemother. 2000, 44, 1818–1824. [Google Scholar] [CrossRef]
- James, G.A.; Zhao, A.G.; Usui, M.; Underwood, R.A.; Hguyen, H.; Beyenal, H.; Pulchini, E.D.; Hunt, A.A.; Bernstein, H.C.; Fleckman, P.; et al. Microsensor and transcriptomic signatures of oxygen depletion in biofilms associated with chronic wounds. Wound Repair Regen. 2015, 24, 373–383. [Google Scholar] [CrossRef]
- Filloux, A.; Ramos, J.-L. Pseudomonas Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Pitts, B.; Hamilton, M.A.; Zelver, N.; Stewart, P.S. A microtiter-plate screening method for biofilm disinfection and removal. J. Microbiol. Methods 2003, 54, 269–276. [Google Scholar] [CrossRef]
- Field, T.; White, A.; Elborn, J.; Tunney, M. Effect of oxygen limitation on the in vitro antimicrobial susceptibility of clinical isolates of Pseudomonas aeruginosa grown planktonically and as biofilms. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 677–687. [Google Scholar] [CrossRef]
- Lopes, S.P.; Azevedo, N.F.; Pereira, M.O. Emergent bacteria in cystic fibrosis: In vitro biofilm formation and resilience under variable oxygen conditions. Biomed. Res. Int. 2014, 2014, 678301. [Google Scholar] [CrossRef]
- Sun, Y.; Dowd, S.E.; Smith, E.; Rhoads, D.D.; Wolcott, R.D. In vitro multispecies Lubbock chronic wound biofilm model. Wound Repair Regen. 2008, 16, 805–813. [Google Scholar] [CrossRef]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar]
- Tre-Hardy, M.; Traore, H.; El Manssouri, N.; Vanderbist, F.; Vaneechoutte, M.; Devleeschouwer, M.J. Evaluation of long-term co-administration of tobramycin and clarithromycin in a mature biofilm model of cystic fibrosis clinical isolates of Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2009, 34, 370–374. [Google Scholar] [CrossRef][Green Version]
- Nickel, J.C.; Ruseska, I.; Wright, J.B.; Costerton, J.W. Tobramycin resistance of Pseudomonas aeruginosa cells growing as a biofilm on urinary catheter material. Antimicrob. Agents Chemother. 1985, 27, 619–624. [Google Scholar] [CrossRef]
- Pereira, M.O.; Kuehn, M.; Wuertz, S.; Neu, T.; Melo, L.F. Effect of flow regime on the architecture of a Pseudomonas fluorescens biofilm. Biotechnol. Bioeng. 2002, 78, 164–171. [Google Scholar] [CrossRef] [PubMed]
- McCoy, W.F.; Costerton, J.W. Fouling Biofilm Development in a Tubular Flow System. Dev. Ind. Microbiol. 1982, 23, 551–558. [Google Scholar]
- Xu, K.D.; Stewart, P.S.; Xia, F.; Huang, C.-T.; McFeters, G.A. Spatial physiological heterogeneity inPseudomonas aeruginosa biofilm is determined by oxygen availability. Appl. Environ. Microbiol. 1998, 64, 4035–4039. [Google Scholar]
- Gilbert, P.; Allison, D.; Evans, D.; Handley, P.; Brown, M. Growth rate control of adherent bacterial populations. Appl. Environ. Microbiol. 1989, 55, 1308–1311. [Google Scholar]
- Ghani, M.; Soothill, J.S. Ceftazidime, gentamicin, and rifampicin, in combination, kill biofilms of mucoid Pseudomonas aeruginosa. Can. J. Microbiol 1997, 43, 999–1004. [Google Scholar] [CrossRef]
- McBain, A.; Sissons, C.; Ledder, R.; Sreenivasan, P.; De Vizio, W.; Gilbert, P. Development and characterization of a simple perfused oral microcosm. J. Appl. Microbiol. 2005, 98, 624–634. [Google Scholar] [CrossRef]
- Hodgson, A.; Nelson, S.; Brown, M.; Gilbert, P. A simple in vitro model for growth control of bacterial biofilms. J. Appl. Microbiol. 1995, 79, 87–93. [Google Scholar]
- Caldwell, D.E.; Lawrence, J.R. Growth kinetics ofPseudomonas fluorescens microcolonies within the hydrodynamic boundary layers of surface microenvironments. Microb. Ecol. 1986, 12, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Benoit, M.R.; Conant, C.G.; Ionescu-Zanetti, C.; Schwartz, M.; Matin, A. New device for high-throughput viability screening of flow biofilms. Appl. Environ. Microbiol. 2010, 76, 4136–4142. [Google Scholar] [CrossRef]
- Ombaka, E.A.; Cozens, R.M.; Brown, M.R. Influence of nutrient limitation of growth on stability and production of virulence factors of mucoid and nonmucoid strains of Pseudomonas aeruginosa. Rev. Infect. Dis. 1983, 5, S880–S888. [Google Scholar] [CrossRef]
- McNeer, N.A.; Anandalingam, K.; Fields, R.J.; Caputo, C.; Kopic, S.; Gupta, A.; Quijano, E.; Polikoff, L.; Kong, Y.; Bahal, R.; et al. Nanoparticles that deliver triplex-forming peptide nucleic acid molecules correct F508del CFTR in airway epithelium. Nat. Commun. 2015, 6, 6952. [Google Scholar] [CrossRef]
- Peters, A.C.; Wimpenny, J.W. A constant-depth laboratory model film fermentor. Biotechnol. Bioeng. 1988, 32, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.E.; Malic, S.; McKee, R.; Rennison, T.; Harding, K.G.; Williams, D.W.; Thomas, D.W. An in vitro model of chronic wound biofilms to test wound dressings and assess antimicrobial susceptibilities. J. Antimicrob. Chemother. 2010, 65, 1195–1206. [Google Scholar] [CrossRef]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects Pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef]
- Woods, D.E.; Schaffer, M.S.; Rabin, H.R.; Campbell, G.; Sokol, P.A. Phenotypic comparison of Pseudomonas aeruginosa strains isolated from a variety of clinical sites. J. Clin. Microbiol. 1986, 24, 260–264. [Google Scholar]
- Trivedi, U.; Madsen, J.S.; Rumbaugh, K.P.; Wolcott, R.D.; Burmolle, M.; Sorensen, S.J. A post-planktonic era of in vitro infectious models: Issues and changes addressed by a clinically relevant wound like media. Crit. Rev. Microbiol. 2017, 43, 453–465. [Google Scholar] [CrossRef]
- Sun, Y.; Smith, E.; Wolcott, R.; Dowd, S.E. Propagation of anaerobic bacteria within an aerobic multi-species chronic wound biofilm model. J. Wound Care 2009, 18, 426–431. [Google Scholar] [CrossRef]
- Kwan, B.W.; Chowdhury, N.; Wood, T.K. Combatting bacterial infections by killing persister cells with mitomycin C. Environ. Microbiol. 2015, 17, 4406–4414. [Google Scholar] [CrossRef] [PubMed]
- Craven, M.; Kasper, S.; Canfield, M.; Diaz-Morales, R.; Hrabie, J.; Cady, N.; Strickland, A. Nitric oxide-releasing polyacrylonitrile disperses biofilms formed by wound-relevant pathogenic bacteria. J. Appl. Microbiol. 2016, 120, 1085–1099. [Google Scholar] [CrossRef]
- Nair, S.; Desai, S.; Poonacha, N.; Vipra, A.; Sharma, U. Antibiofilm Activity and Synergistic Inhibition of Staphylococcus aureus Biofilms by Bactericidal Protein P128 in Combination with Antibiotics. Antimicrob. Agents Chemother. 2016, 60, 7280–7289. [Google Scholar] [PubMed]
- Dowd, S.E.; Sun, Y.; Smith, E.; Kennedy, J.P.; Jones, C.E.; Wolcott, R. Effects of biofilm treatments on the multi-species Lubbock chronic wound biofilm model. J. Wound Care 2009, 18, 508, 510–512. [Google Scholar] [CrossRef]
- Kucera, J.; Sojka, M.; Pavlik, V.; Szuszkiewicz, K.; Velebny, V.; Klein, P. Multispecies biofilm in an artificial wound bed—A novel model for in vitro assessment of solid antimicrobial dressings. J. Microbiol. Methods 2014, 103, 18–24. [Google Scholar] [CrossRef]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in vivo polymicrobial biofilm wound infection model to study interspecies interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Rumbaugh, K.P.; James, G.; Schultz, G.; Phillips, P.; Yang, Q.; Watters, C.; Stewart, P.S.; Dowd, S.E. Biofilm maturity studies indicate sharp debridement opens a time-dependent therapeutic window. J. Wound Care 2010, 19, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Larose, C.; Della Porta, A.C.; Schultz, G.S.; Gibson, D.J. A surfactant-based wound dressing can reduce bacterial biofilms in a porcine skin explant model. Int. Wound J. 2017, 14, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Bowler, P.G.; Dolman, J. Antimicrobial activity of silver-containing dressings on wound microorganisms using an in vitro biofilm model. Int. Wound J. 2007, 4, 186–191. [Google Scholar] [CrossRef]
- Werthén, M.; Henriksson, L.; Jensen, P.Ø.; Sternberg, C.; Givskov, M.; Bjarnsholt, T. An in vitro model of bacterial infections in wounds and other soft tissues. Apmis 2010, 118, 156–164. [Google Scholar] [CrossRef]
- Brackman, G.; Cos, P.; Maes, L.; Nelis, H.J.; Coenye, T. Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob. Agents Chemother. 2011, 55, 2655–2661. [Google Scholar] [CrossRef]
- Bowler, P.G.; Parsons, D. Combatting wound biofilm and recalcitrance with a novel anti-biofilm Hydrofiber® wound dressing. Wound Med. 2016, 14, 6–11. [Google Scholar] [CrossRef]
- Martineau, L.; Dosch, H.M. In vitro bactericidal efficacy of a new sun- and heat burn gel. Burns 2006, 32, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.; Boegli, L.; Kirker, K.R.; Agostinho, A.M.; Durch, A.M.; deLancey Pulcini, E.; Stewart, P.S.; James, G.A. Development and application of a polymicrobial, in vitro, wound biofilm model. J. Appl. Microbiol. 2012, 112, 998–1006. [Google Scholar] [CrossRef]
- Townsend, E.M.; Sherry, L.; Rajendran, R.; Hansom, D.; Butcher, J.; Mackay, W.G.; Williams, C.; Ramage, G. Development and characterisation of a novel three-dimensional inter-kingdom wound biofilm model. Biofouling 2016, 32, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Grassi, L.; Batoni, G.; Ostyn, L.; Rigole, P.; Van den Bossche, S.; Rinaldi, A.C.; Maisetta, G.; Esin, S.; Coenye, T.; Crabbé, A. The Antimicrobial Peptide lin-SB056-1 and Its Dendrimeric Derivative Prevent Pseudomonas aeruginosa Biofilm Formation in Physiologically Relevant Models of Chronic Infections. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Lebeaux, D.; Ghigo, J.M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef]
- Naghili, H.; Tajik, H.; Mardani, K.; Razavi Rouhani, S.M.; Ehsani, A.; Zare, P. Validation of drop plate technique for bacterial enumeration by parametric and nonparametric tests. Vet. Res. Forum 2013, 4, 179–183. [Google Scholar] [PubMed]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Goeres, D.M.; Beck, N.A.; Buckingham-Meyer, K.; Hilyard, J.D.; Loetterle, L.R.; Lorenz, L.A.; Walker, D.K.; Stewart, P.S. A method for growing a biofilm under low shear at the air–liquid interface using the drip flow biofilm reactor. Nat. Protoc. 2009, 4, 783–788. [Google Scholar] [CrossRef]
- Bevilacqua, M.P.; Huang, D.J.; Wall, B.D.; Lane, S.J.; Edwards, C.K., 3rd; Hanson, J.A.; Benitez, D.; Solomkin, J.S.; Deming, T.J. Amino Acid Block Copolymers with Broad Antimicrobial Activity and Barrier Properties. Macromol. Biosci. 2017, 17. [Google Scholar] [CrossRef]
- Microbiological Tests 〈61〉 Microbiological Examination of Nonsterile Products: Microbial Enumeration Tests. In United States Pharmacopeia and National Formulary (USP 40-NF 35); United States Pharmacopeial Convention: Rockville, MD, USA, 2017.
- Percival, S.; Cutting, K. Microbiology of Wounds; Percival, S., Cutting, K., Eds.; CRC Press: New York, NY, USA, 2010. [Google Scholar]
- Stiefel, P.; Rosenberg, U.; Schneider, J.; Mauerhofer, S.; Maniura-Weber, K.; Ren, Q. Is biofilm removal properly assessed? Comparison of different quantification methods in a 96-well plate system. Appl. Microbiol. Biotechnol. 2016, 100, 4135–4145. [Google Scholar] [CrossRef]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef]
- Hentzer, M.; Wu, H.; Andersen, J.B.; Riedel, K.; Rasmussen, T.B.; Bagge, N.; Kumar, N.; Schembri, M.A.; Song, Z.; Kristoffersen, P. Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. Embo J. 2003, 22, 3803–3815. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Kirketerp-Møller, K.; Kristiansen, S.; Phipps, R.; Nielsen, A.K.; Jensen, P.Ø.; Høiby, N.; Givskov, M. Silver against Pseudomonas aeruginosa biofilms. Apmis 2007, 115, 921–928. [Google Scholar] [CrossRef]
- Molecular Probes Inc. Product information sheet MP07007. LIVE/DEAD® BacLight™ Bacterial Viability Kits; Eugene, OR, USA, 2004. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/mp07007.pdf (accessed on 29 April 2019).
- Percival, S.L.; Slone, W.; Linton, S.; Okel, T.; Corum, L.; Thomas, J.G. Use of flow cytometry to compare the antimicrobial efficacy of silver-containing wound dressings against planktonic Staphylococcus aureus and Pseudomonas aeruginosa. Wound Repair Regen. 2011, 19, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Ammons, M.C.; Ward, L.S.; Dowd, S.; James, G.A. Combined treatment of Pseudomonas aeruginosa biofilm with lactoferrin and xylitol inhibits the ability of bacteria to respond to damage resulting from lactoferrin iron chelation. Int. J. Antimicrob. Agents 2011, 37, 316–323. [Google Scholar] [CrossRef][Green Version]
- Kim, M.H.; Yamayoshi, I.; Mathew, S.; Lin, H.; Nayfach, J.; Simon, S.I. Magnetic nanoparticle targeted hyperthermia of cutaneous Staphylococcus aureus infection. Ann. Biomed. Eng. 2013, 41, 598–609. [Google Scholar] [CrossRef]
- Ghaseminezhad, S.M.; Shojaosadati, S.A.; Meyer, R.L. Ag/Fe3O4 nanocomposites penetrate and eradicate S. aureus biofilm in an in vitro chronic wound model. Colloids Surf. B Biointerfaces 2018, 163, 192–200. [Google Scholar] [CrossRef]
- Percival, S.L.; Bowler, P.; Woods, E.J. Assessing the effect of an antimicrobial wound dressing on biofilms. Wound Repair Regen. 2008, 16, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Boulton, C.A. Encyclopedia of Brewing; John Wiley & Sons Inc.: London, UK, 2013. [Google Scholar]
- Gold, O.G.; Jordan, H.V.; Houte, J.V. Identification of Streptococcus mutans colonies by mannitol-dependent tetrazolium reduction. Archs Oral Biol. 1974, 19, 271–272. [Google Scholar] [CrossRef]
- Sabaeifard, P.; Abdi-Ali, A.; Soudi, M.R.; Dinarvand, R. Optimization of tetrazolium salt assay for Pseudomonas aeruginosa biofilm using microtiter plate method. J. Microbiol. Methods 2014, 105, 134–140. [Google Scholar] [CrossRef]
- Stevens, M.G.; Olsen, S.C. Comparative analysis of using MTT and XTT in colorimetric assays for quantitating bovine neutrophil bactericidal activity. J. Immunol. Methods 1993, 157, 225–231. [Google Scholar] [CrossRef]
- Huang, C.-T.; Yu, F.P.; McFeters, G.A.; Stewart, P.S. Nonuniform spatial patterns of respiratory activity within biofilms during disinfection. Appl. Environ. Microbiol. 1995, 61, 2252–2256. [Google Scholar] [PubMed]
- Zheng, Z.; Stewart, P.S. Growth limitation of Staphylococcus epidermidis in biofilms contributes to rifampin tolerance. Biofilms 2004, 1, 31–35. [Google Scholar] [CrossRef]
- Tunney, M.M.; Ramage, G.; Field, T.R.; Moriarty, T.F.; Storey, D.G. Rapid colorimetric assay for antimicrobial susceptibility testing of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2004, 48, 1879–1881. [Google Scholar] [CrossRef] [PubMed]
- Sigma-Aldrich. In Vitro Toxicity Assay Kit, XTT Based; Sigma-Aldrich: St. Louis, MO, USA, 2005. [Google Scholar]
- Ramage, G. Comparing apples and oranges: Considerations for quantifying candidal biofilms with XTT [2–bis(2-methoxy-4-nitro-5-sulfo-phenyl)-2H-tetrazolium-5-carboxanilide] and the need for standardized testing. J. Med. Microbiol. 2016, 65, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.; Karrasch, B.; Hoppe, H.; Jeskulke, K.; Mehrens, M. Toxic effects on bacterial metabolism of the redox dye 5-cyano-2, 3-ditolyl tetrazolium chloride. Appl. Environ. Microbiol. 1996, 62, 4587–4593. [Google Scholar] [PubMed]
- Zimmermann, R.; Iturriaga, R.; Becker-Birck, J. Simultaneous determination of the total number of aquatic bacteria and the number thereof involved in respiration. Appl. Environ. Microbiol. 1978, 36, 926–935. [Google Scholar] [PubMed]
- Alonso, B.; Cruces, R.; Perez, A.; Sanchez-Carrillo, C.; Guembe, M. Comparison of the XTT and resazurin assays for quantification of the metabolic activity of Staphylococcus aureus biofilm. J. Microbiol. Methods 2017, 139, 135–137. [Google Scholar] [CrossRef]
- Junka, A.F.; Zywicka, A.; Szymczyk, P.; Dziadas, M.; Bartoszewicz, M.; Fijalkowski, K.A.D.A.M. test (Antibiofilm Dressing’s Activity Measurement)—Simple method for evaluating anti-biofilm activity of drug-saturated dressings against wound pathogens. J. Microbiol. Methods 2017, 143, 6–12. [Google Scholar] [CrossRef]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef]
- Van den Driessche, F.; Rigole, P.; Brackman, G.; Coenye, T. Optimization of resazurin-based viability staining for quantification of microbial biofilms. J. Microbiol. Methods 2014, 98, 31–34. [Google Scholar] [CrossRef]
- Invitrogen. Product information sheet MAN0003232. PrestoBlue Cell Viability Reagent Protocol; Carlsbad, CA, USA, 2010. Available online: http://tools.thermofisher.com/content/sfs/manuals/PrestoBlue_Reagent_PIS_15Oct10.pdf (accessed on 29 April 2019).
- Invitrogen. Application Note: Processing Absorbance Data Obtained Using PrestoBlue Viability Reagen; Carlsbad, CA, USA, 2011. Available online: http://www.ulab360.com/files/prod/manuals/201509/13/2142001.pdf (accessed on 29 April 2019).
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.L.; Bonaventura, G.D.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Bahamondez-Canas, T.F.; Zhang, H.; Tewes, F.; Leal, J.; Smyth, H.D.C. PEGylation of Tobramycin Improves Mucus Penetration and Antimicrobial Activity against Pseudomonas aeruginosa Biofilms in Vitro. Mol. Pharm. 2018. [Google Scholar] [CrossRef]
- Thorn, R.M.; Nelson, S.M.; Greenman, J. Use of a bioluminescent Pseudomonas aeruginosa strain within an in vitro microbiological system, as a model of wound infection, to assess the antimicrobial efficacy of wound dressings by monitoring light production. Antimicrob. Agents Chemother 2007, 51, 3217–3224. [Google Scholar] [CrossRef] [PubMed]
- Thorn, R.M.; Greenman, J. A novel in vitro flat-bed perfusion biofilm model for determining the potential antimicrobial efficacy of topical wound treatments. J. Appl. Microbiol 2009, 107, 2070–2079. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Wanandy, S.; Brouwer, N.; Liu, Q.; Mahon, A.; Cork, S.; Karuso, P.; Vemulpad, S.; Jamie, J. Optimisation of the fluorescein diacetate antibacterial assay. J. Microbiol. Methods 2005, 60, 21–30. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011. [Google Scholar] [CrossRef]
- Tote, K.; Horemans, T.; Vanden Berghe, D.; Maes, L.; Cos, P. Inhibitory effect of biocides on the viable masses and matrices of Staphylococcus aureus and Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2010, 76, 3135–3142. [Google Scholar] [CrossRef]
- Harrison-Balestra, C.; Cazzaniga, A.L.; Davis, S.C.; Mertz, P.M. A Wound-Isolated Pseudomonas aeruginosa Grows a Biofilm In Vitro Within 10 Hours and Is Visualized by Light Microscopy. Derm. Surg. 2003, 29, 631–635. [Google Scholar]
- Ommen, P.; Zobek, N.; Meyer, R.L. Quantification of biofilm biomass by staining: Non-toxic safranin can replace the popular crystal violet. J. Microbiol. Methods 2017, 141, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Baird, F.J.; Wadsworth, M.P.; Hill, J.E. Evaluation and optimization of multiple fluorophore analysis of a Pseudomonas aeruginosa biofilm. J. Microbiol. Methods 2012, 90, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.Y.; Lee, D.J.; Tay, J.H.; Show, K.Y. Staining of extracellular polymeric substances and cells in bioaggregates. Appl. Microbiol. Biotechnol. 2007, 75, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Strathmann, M.; Wingender, J.; Flemming, H.-C. Application of fluorescently labelled lectins for the visualization and biochemical characterization of polysaccharides in biofilms of Pseudomonas aeruginosa. J. Microbiol. Methods 2002, 50, 237–248. [Google Scholar] [CrossRef]
- Bandara, H.M.; Nguyen, D.; Mogarala, S.; Osinski, M.; Smyth, H.D. Magnetic fields suppress Pseudomonas aeruginosa biofilms and enhance ciprofloxacin activity. Biofouling 2015, 31, 443–457. [Google Scholar] [CrossRef]
- Ross, S.S.; Gharse, S.; Sanchez, L.; Fiegel, J. Dry powder aerosols to co-deliver antibiotics and nutrient dispersion compounds for enhanced bacterial biofilm eradication. Int. J. Pharm. 2017, 531, 14–23. [Google Scholar] [CrossRef]
- Lewandowski, Z.; Beyenal, H. Imaging and characterizing biofilm components. In Fundamentals of Biofilm Research, 2nd ed.; CRC Press: New York, NY, USA, 2014; pp. 67–141. [Google Scholar]
- Wagner, M.; Ivleva, N.P.; Haisch, C.; Niessner, R.; Horn, H. Combined use of confocal laser scanning microscopy (CLSM) and Raman microscopy (RM): Investigations on EPS-Matrix. Water Res. 2009, 43, 63–76. [Google Scholar] [CrossRef]
- Kusić, D.; Kampe, B.; Ramoji, A.; Neugebauer, U.; Rösch, P.; Popp, J. Raman spectroscopic differentiation of planktonic bacteria and biofilms. Anal. Bioanal. Chem. 2015, 407, 6803–6813. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.B.; Nam, S.W.; Choi, S.; Lee, G.J.; Park, H.K. Evaluation of antibiotic effects on Pseudomonas aeruginosa biofilm using Raman spectroscopy and multivariate analysis. Biomed. Opt. Express 2014, 5, 3238–3251. [Google Scholar] [CrossRef]
- Alhede, M.; Qvortrup, K.; Liebrechts, R.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Combination of microscopic techniques reveals a comprehensive visual impression of biofilm structure and composition. FEMS Immunol. Med. Microbiol. 2012, 65, 335–342. [Google Scholar] [CrossRef] [PubMed]
- El Abed, S.; Ibnsouda, S.K.; Latrache, H.; Hamadi, F. Scanning electron microscopy (SEM) and environmental SEM: Suitable tools for study of adhesion stage and biofilm formation. In Scanning Electron Microscopy; Kazmiruk, V., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 717–730. [Google Scholar] [CrossRef]
- Gordon, V.D.; Davis-Fields, M.; Kovach, K.; Rodesney, C.A. Biofilms and mechanics: A review of experimental techniques and findings. J. Phys. D Appl. Phys. 2017, 50. [Google Scholar] [CrossRef]
- Bhat, S.; Dong, J.; Biplab, P.; Dahms, T. Viscoelasticity in Biological Systems: A Special Focus on Microbes. In Viscoelasticity-From Theory to Biological Applications; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Rogers, S.S.; Walle, C.V.D.; Waigh, T.A. Microrheology of Bacterial Biofilms In Vitro: Staphylococcus aureus and Pseudomonas aeruginosa. Langmuir 2008, 24, 13549–13555. [Google Scholar] [CrossRef] [PubMed]
- Rupp, C.J.; Fux, C.A.; Stoodley, P. Viscoelasticity of Staphylococcus aureus biofilms in response to fluid shear allows resistance to detachment and facilitates rolling migration. Appl. Environ. Microbiol. 2005, 71, 2175–2178. [Google Scholar] [CrossRef]
- Gloag, E.S.; German, G.K.; Stoodley, P.; Wozniak, D.J. Viscoelastic properties of Pseudomonas aeruginosa variant biofilms. Sci. Rep. 2018, 8, 9691. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, S.; Stoodley, P. Mechanical properties of Biofilms. In The Perfect Slime: Microbial Extracellular Polymeric Substances (EPS); Flemming, H.C., Neu, D.T.R., Wingender, D.J., Eds.; IWA Publishing: London, UK, 2016. [Google Scholar]
- Heatley, N.G. A method for the assay of penicillin. Biochem. J. 1944, 38, 61–65. [Google Scholar] [CrossRef]
- Bradshaw, C.E. An in vitro comparison of the antimicrobial activity of honey, iodine and silver wound dressings. Biosci. Horiz. 2011, 4, 61–70. [Google Scholar] [CrossRef]
- Prabhu, Y.T.; Rao, K.V.; Kumari, B.S.; Kumar, V.S.S.; Pavani, T. Synthesis of Fe3O4 nanoparticles and its antibacterial application. Int. Nano Lett. 2015, 5, 85–92. [Google Scholar] [CrossRef]
- Pettit, R.K.; Weber, C.A.; Pettit, G.R. Application of a high throughput Alamar blue biofilm susceptibility assay to Staphylococcus aureus biofilms. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Hill, K.E.; Williams, D.W.; Hooper, S.J.; Thomas, D.W.; Costerton, J.W. A review of the scientific evidence for biofilms in wounds. Wound Repair Regen. 2012, 20, 647–657. [Google Scholar] [CrossRef]
- Schlafer, S.; Meyer, R.L. Confocal microscopy imaging of the biofilm matrix. J. Microbiol. Methods 2017, 138, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Rodesney, C.A.; Roman, B.; Dhamani, N.; Cooley, B.J.; Touhami, A.; Gordon, V.D. Mechanosensing of shear by Pseudomonas aeruginosa leads to increased levels of the cyclic-di-GMP signal initiating biofilm development. Proc. Natl. Acad. Sci. USA 2017, 114, 5906–5911. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Strains | Support | Surface | Media | Incubation | Application | References |
|---|---|---|---|---|---|---|---|
| Chronic Wound Infection Models | |||||||
| Germ carrier model | PA ATCC 27317 | Petri dish | Polyurethane sponge | TSB | 4, 7, and 24 h | Biofilm susceptibility in in vitro to semisolid formulations | Martineau, 2006 [139] |
| Poloxamer biofilm | PA NCIMB 8626 | Petri dish | Poloxamer hydrogel | MHB | 24 h | Biofilm formation and susceptibility testing of different commercial silver-containing dressings. | Percival, 2007 [135] |
| Lubbock chronic wound pathogenic biofilm (LCWPB) | PAO1, E. faecalis, and S. aureus | Microtiter plate | Polystyrene 96-well plate | WLM media: 45% Bolton broth, 50% bovine plasma, and 5% freeze-thaw laked horse red blood cells | Anaerobic for 24 h | Definition of media composition for multispecies biofilm formation and susceptibility testing | Sun, 2008 [106] |
| Porcine explant wound biofilm model | PAO1 and S. aureus | TSA plate | Partial thickness wound beds in fresh porcine skin explants | TSB | Up to 5 days | Biofilm susceptibility testing | Wolcott, 2010 [133] |
| Collagen wound biofilm | PAO1 and S. aureus | Culture slides | Collagen matrices | 50% fetal calf serum and 50% physiological NaCl in 0.1% Peptone | 48 h | Biofilm formation within collagen matrices and susceptibility testing | Werthen, 2010 [136] |
| Polymicrobial colony-DFR wound biofilm | PA, MRSA, and C. perfringens wound isolates | DFR | Polycarbonate membrane | 100% BHI with 5% adult bovine serum | 3 days | Biofilm formation and susceptibility testing of dressings | Woods, 2012 [140] |
| Cellulose interkingdom biofilm wound model | PA14, S. aureus, and C. albicans SC5314 | N/D | Cellulose matrices on top of hydrogel | PBS | 24 h | Biofilm formation within cellulose matrices and susceptibility testing | Townsend, 2016 [141] |
| Artificial wound model | PAO1 and clinical isolates | Microtiter plate | Hyaluronic acid and collagen scaffold | Bolton broth with 50% bovine plasma and 5% freeze-thaw laked horse blood | 16 h | Biofilm formation and susceptibility testing of antimicrobial peptides | Grassi, 2019 [142] |
| Category of Evaluation | Principle/Target | Method Overview | Detection Method | Example Detection Settings | Example Model Systems | References |
|---|---|---|---|---|---|---|
| Viability | ||||||
| Colony counting | Viable cells are able to form colonies when plated on appropriate agar substrates | Dispersions of cells are spread or drop-plated. Colonies formed counted after appropriate growth period | Visual | Agar plate | [136,180,208] | |
| SYTO 9 | All Cells | Nucleic acids stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, FM | Ex: 485 nm Em: 528 nm | Microtiter plate | [150] |
| SYTO 9/PI | All cells (SYTO9) Dead/membrane permeable cells (PI) | Nucleic acids stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, FM | Ex: 485 nm Em: 528 or 645 nm | Microtiter plate, flow cell, collagen model | [150,157,158,159] |
| Acridine orange | All cells (nucleic acids) | Nucleic acids stained and dispersed by vortexing in appropriate media | FS | Ex: 485 nm Em: 528 nm | Microtiter plate | [150] |
| Ethidium bromide | DNA | DNA stained and visualized. Appears orange when excited | FS, FM | Ex: 210 or 285 nm Em: 605 nm | Constant depth film fermenter (CDFF) and glass microscopy slide | [122] |
| Ziehl carbol fuchsin | Bacterial cells | Stains bacterial cells red/purple | LM | CDFF and glass microscopy slide | [122] | |
| DAPI | DNA | DNA stained and visualized. | FM, LM | Ex: 350 nm Em: 470 nm | Microtiter plate and glass slide | [136] |
| Metabolic Activity | ||||||
| Tetrazolium Salts (INT, TTC, CTC, XTT, and MTT) | Reduction of Tetrazolium to formazan | Dissolved dye from stained biofilms recovered and quantified | AS | INT: 470 nm TTC: 405, 450, 490, 540 nm XTT: 450-492 nm (486nm) | Microtiter plate, modified agar plate | [150,161,162,172] |
| Resazurin (Alamar Blue, PrestoBlue, CellTiter-Blue) | Reduction of Resazurin to resorufin | Reagent incubated with media and biofilms | FS, AS | Ex: 560 nm Em: 590 nm Abs: 570 and 600 nm | Microtiter plate | [173,174,208] |
| Bioluminescent Assay (BacTiter) | Catalysis of ATP and luciferin by luciferase | D-luciferin is used in these assays as it undergoes conversion by luciferase to oxyluciferin a light generating compound when in the presence of ATP | L*S | Microtiter plate | [150,179,180,181,209] | |
| Fluorescein diacetate (FDA) | Cleavage of acetate by intracellular esterases | FDA converted to yellow fluorescent fluorescein | FS | Ex: 494 nm Em: 518 nm | Microtiter plate | [182] |
| Biomass | ||||||
| Crystal violet | Stains negatively-charged molecules and polymers. All biomass (live, dead, and matrix) | Stained biofilms dissolved in appropriate solvent | AS | 550 - 600 nm | Microtiter plate | [102,185] |
| Congo red | Polysaccharides and cell membranes | Stained biofilms dissolved in appropriate solvent | AS | 500nm | Microtiter plate CDFF and glass microscopy slide | [122,150] |
| Safranin | Nuclei and mucin red | Stained biofilms dissolved in appropriate solvent | AS | ~535 nm | Microtiter plate | [150,187] |
| Calcofluor white | beta-polysaccharides in matrix | Biofilms stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, AS | Ex: 360, 365, or 400 nm Em: 460, 435, 410 nm | Microtiter plate CDFF and glass microscopy slide | [122,150,188,189] |
| SYPRO Ruby | proteins | Biofilms stained and dispersed by vortexing in appropriate media | FS | Ex: 450 or 460 nm Em: 610 or 645 nm | Microtiter plate | [150,188] |
| FITC | proteins and amino-sugars | Biofilms stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, FM | Ex: ~488 nm Em: 500-550 nm | Microtiter plate | [150,188,189] |
| Concanavalin A (Con A) conjugates | alpha-Mannopyranosyl and alpha-glucopyranosyl sugars | Biofilms stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, FM | Ex: 543 nm Em: 550-600 nm | [189] | |
| FITC-Con A | polysaccharides | Biofilms stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, FM | Ex: 485 nm Em: 528 nm | Microtiter plate | [150] |
| Con A - Tetramethylrhodamine | Alpha polysaccharides | Biofilms stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FS, FM | Ex: 555 nm Em: 580 nm | Biofilm reactor | [188] |
| Periodic acid-Schiff (PAS) | Stains polysaccharides | LM | Microtiter plate and glass slide | [136] | ||
| van Gieson | Stains collagen Fibers and Bacterial DNA | LM | Microtiter plate and glass slide | [136] | ||
| Vybrant DiD | Lipids and Membranes | Biofilms stained and visualized (M) or dispersed by vortexing in appropriate media (S) | FM | Ex: 644 nm Em: 665 nm | Biofilm reactor | [188] |
| Turbidity | ||||||
| Turbidity threshold method | Quantification of dispersed cells | Measure absorbance of bacterial suspension and bacteria-free media and compare to a known dilution series | AS | 600 nm | Microtiter plate | [150,209] |
| MacFarland standards | Quantification of dispersed cells | Measure absorbance of bacterial suspension and McFarland Standards (mixtures of H2SO4+BaCl2 or latex particles) | AS | 625 nm | Microtiter plate | |
| Structure | ||||||
| Scanning electron microscopy (SEM) | Visualization of morphology and distribution of microorganisms and extracellular matrix (ECM) | Biofilms typically fixed and negatively stained (SEM) | SEM/Cryo-SEM/ESEM | Varies by instrument | Flat-bed perfusion system, collagen model | [158,181,209,210] |
| Confocal scanning laser microscopy (CLSM) | Isolation of 3D microbial community | Use applicable stains and dyes listed above to visualize various aspects of the biofilm | FM | Varies by stain/dye | Glass microscopy slide, flow cell | [209,211] |
| Fluorescent in-situ hybridization (FISH/PNA-FISH) | Visualize patterns of microbial colonization | Fluorescently labeled oligonucleotide probes hybridize to ribosomal RNA in cells that have been fixed and permeabilized | FM | Varies by stain/dye | Glass microscopy slide | [136,177,209] |
| Raman microscopy | Mapping of microorganisms and ECM Raman spectra | RM | Varies by instrument/target | Raman-neutral slide | [209] | |
| Mechanics | ||||||
| Atomic force microscopy (AFM) | Mapping of local and global adhesive and cohesive forces | Measure force-displacement curves | Varies by instrument/target | [97,212] | ||
| Micro-rheology | Measure behavior of isolated bacteria under different physical conditions | FM, LM | Varies by stain/dye | Flow cell | [202] | |
| Bulk rheology | Biofilms have viscoelastic properties | Measure viscoelastic properties of ECM matrix with microorganisms | Rheometer | Varies by instrument | Agar plate, colony system | [97] |
| Other | ||||||
| Agar disk/well-diffusion | Zone-of-inhibition of therapies measured | Agar plates inoculated with bacteria are exposed to a therapy within a defined area. Following growth period, area of new growth measured | Visual | Agar plate | [178] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahamondez-Canas, T.F.; Heersema, L.A.; Smyth, H.D.C. Current Status of In Vitro Models and Assays for Susceptibility Testing for Wound Biofilm Infections. Biomedicines 2019, 7, 34. https://doi.org/10.3390/biomedicines7020034
Bahamondez-Canas TF, Heersema LA, Smyth HDC. Current Status of In Vitro Models and Assays for Susceptibility Testing for Wound Biofilm Infections. Biomedicines. 2019; 7(2):34. https://doi.org/10.3390/biomedicines7020034
Chicago/Turabian StyleBahamondez-Canas, Tania F., Lara A. Heersema, and Hugh D. C. Smyth. 2019. "Current Status of In Vitro Models and Assays for Susceptibility Testing for Wound Biofilm Infections" Biomedicines 7, no. 2: 34. https://doi.org/10.3390/biomedicines7020034
APA StyleBahamondez-Canas, T. F., Heersema, L. A., & Smyth, H. D. C. (2019). Current Status of In Vitro Models and Assays for Susceptibility Testing for Wound Biofilm Infections. Biomedicines, 7(2), 34. https://doi.org/10.3390/biomedicines7020034