Short-Term Soy Protein Isolate Feeding Prevents Liver Steatosis and Reduces Serum ALT and AST Levels in Obese Female Zucker Rats

Abstract
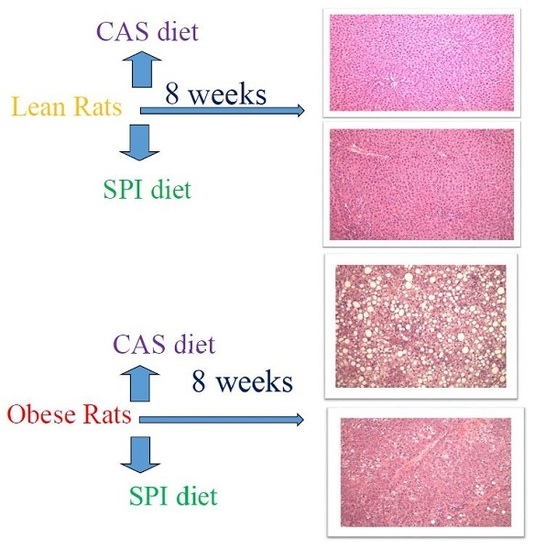
:
1. Introduction
2. Materials and Methods
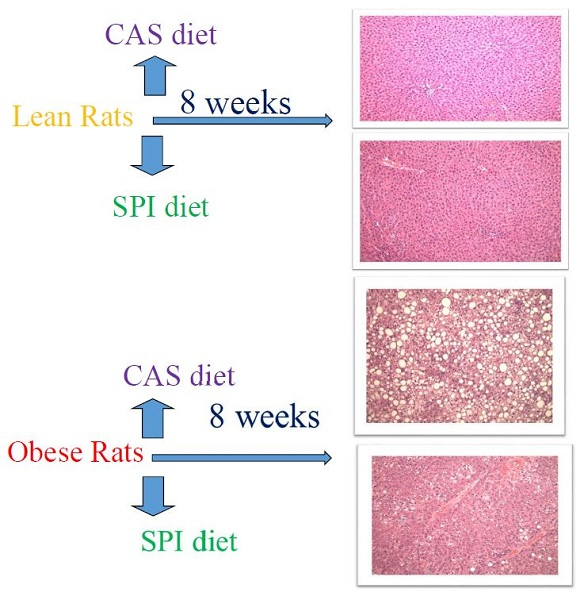
2.1. Experimental Design
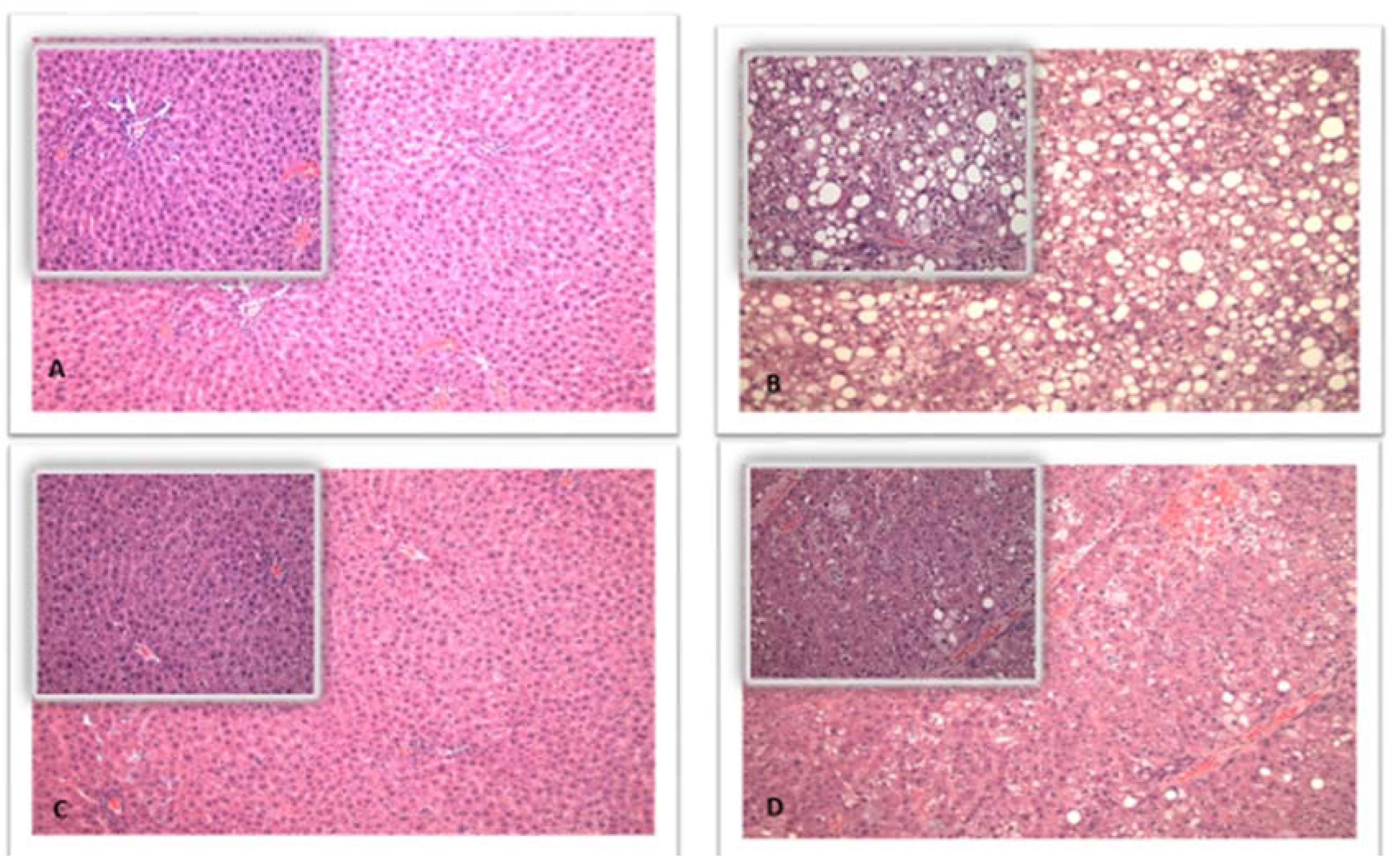
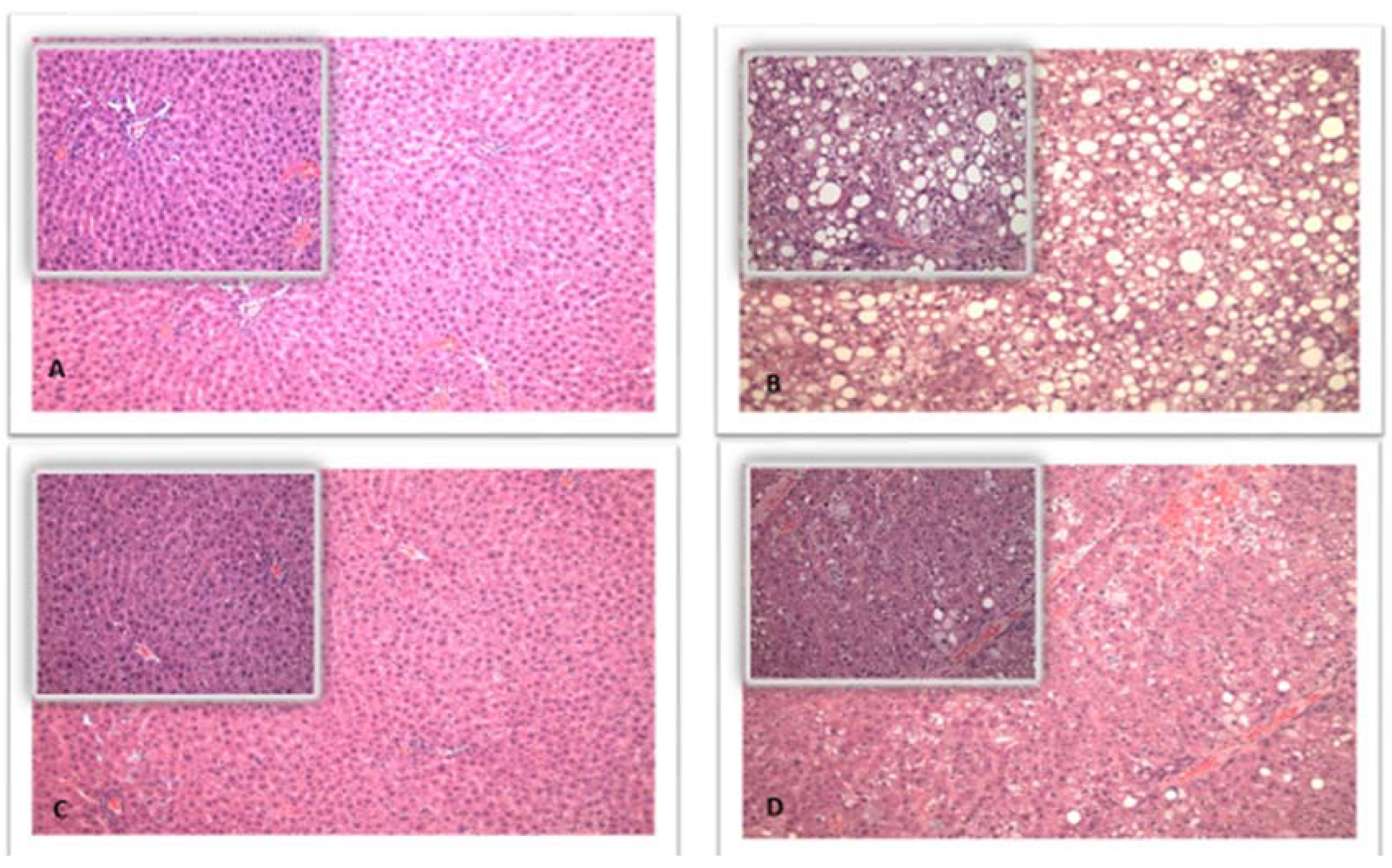
2.2. Liver Histology and Weights
2.3. Serum Measurements
2.4. Statistical Analysis
3. Results
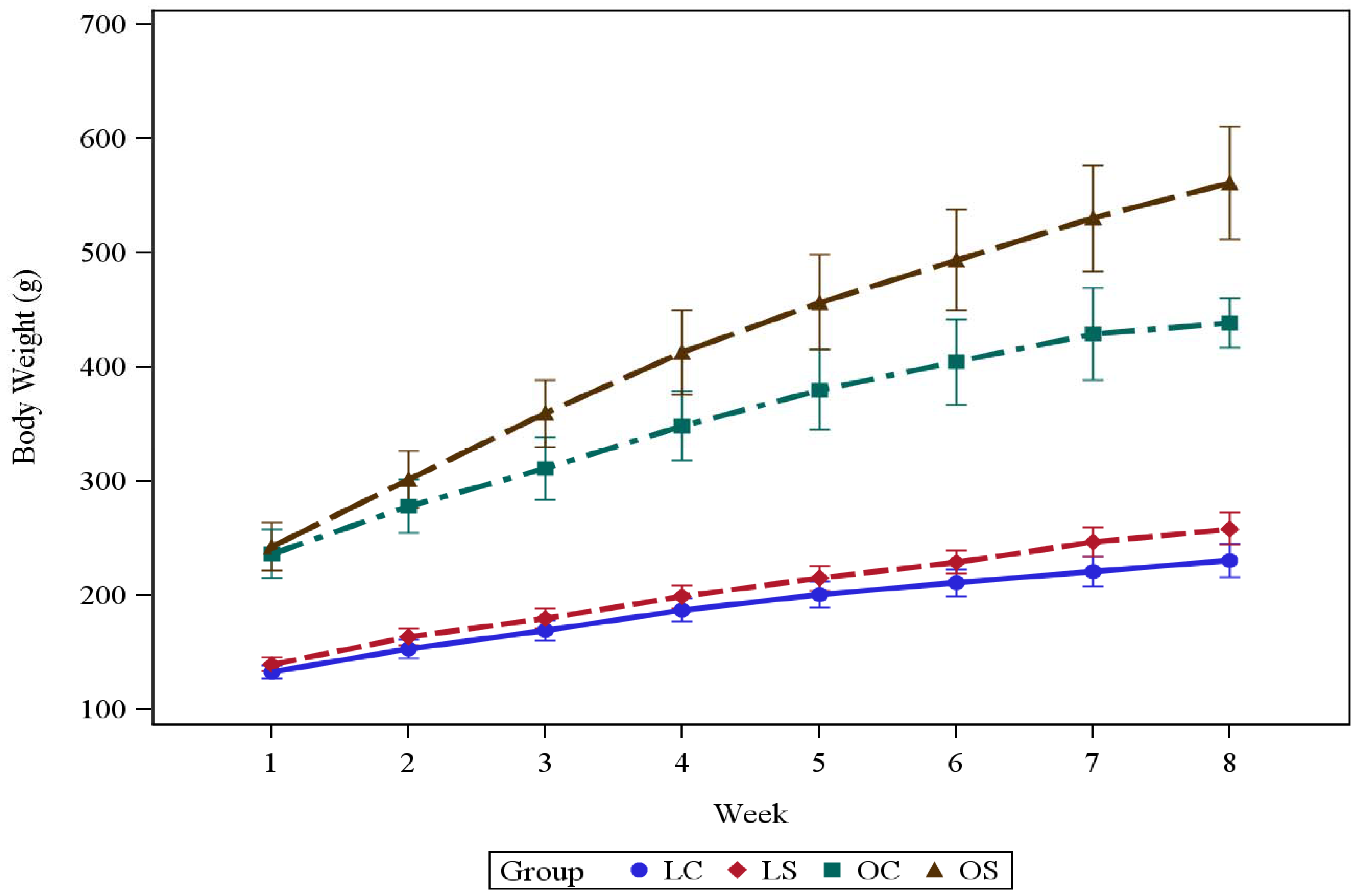
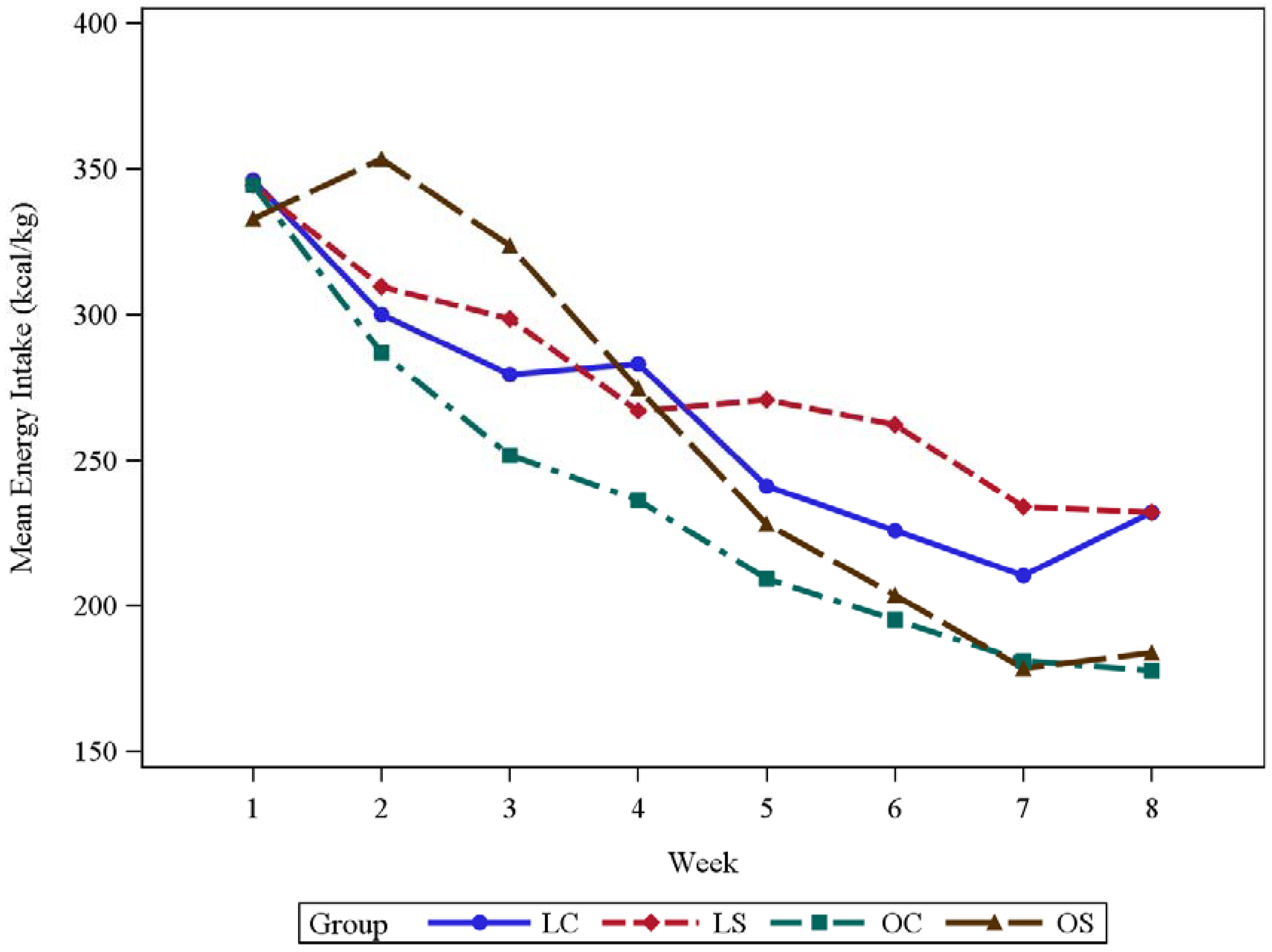
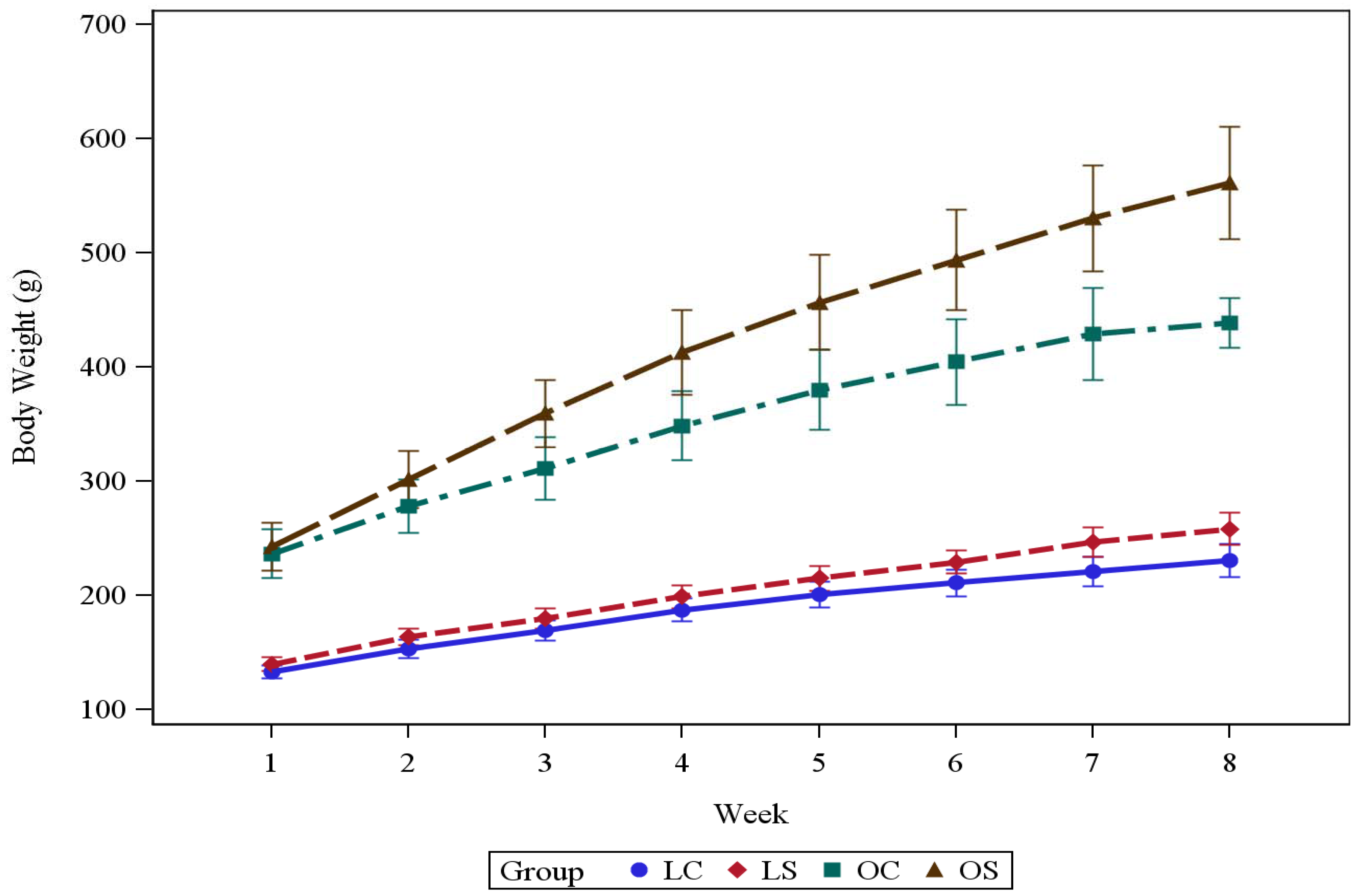
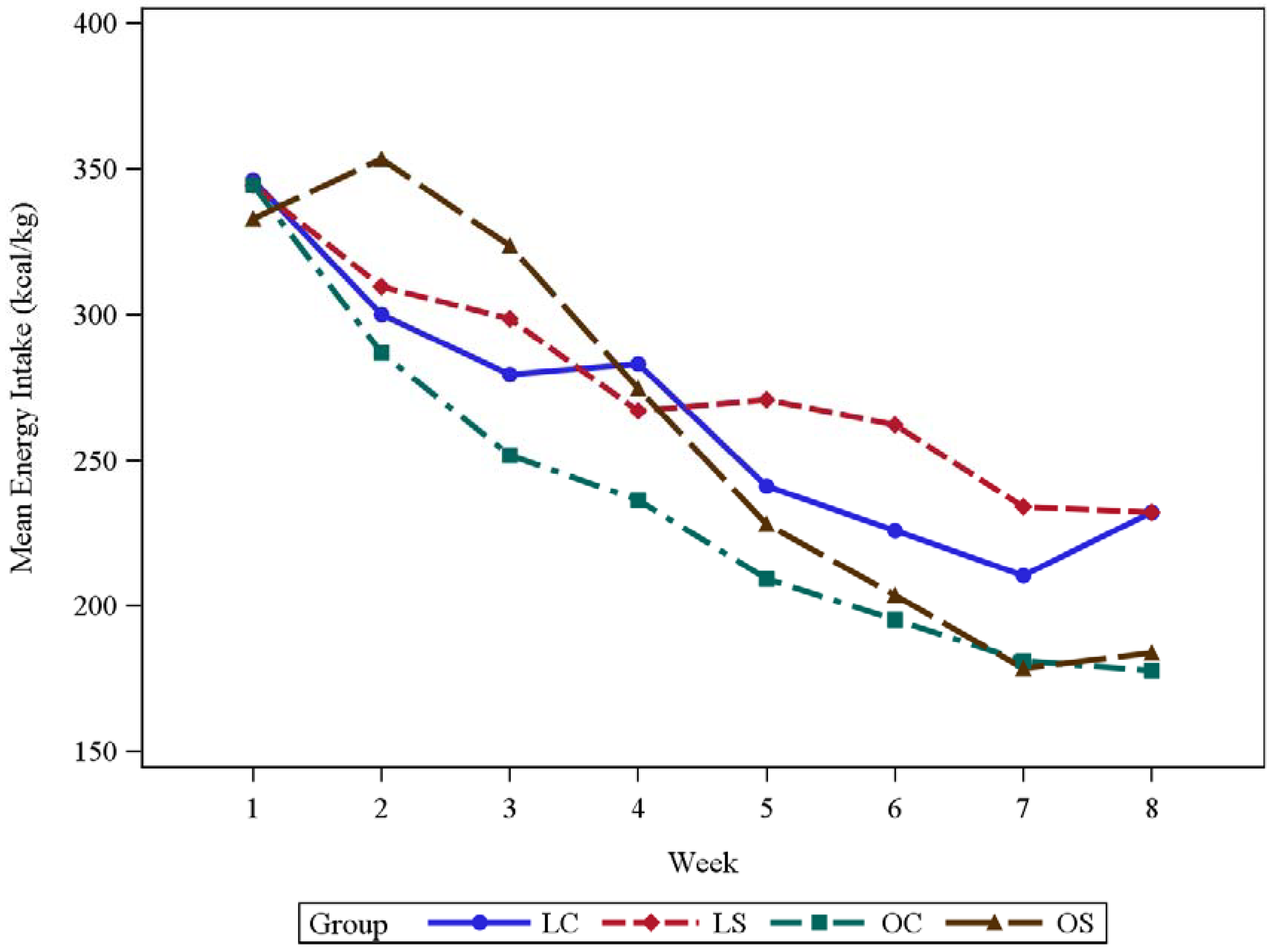
3.1. Body Weight and Energy Intake
3.2. Serum Measurement for Leptin, AST, and ALT
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ogden, C.L.; Carroll, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of Childhood and Adult Obesity in the United States, 2011–2012. JAMA 2014, 311, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention. Overweight and Obesity. Available online: http://www.cdc.gov/obesity/index.html (accessed on 6 February 2018.).
- World Health Organization. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en (accessed on 6 February 2018).
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Mokdad, A.H.; Ford, E.S.; Bowman, B.A.; Dietz, W.H.; Vinicor, F.; Bales, V.S.; Marks, J.S. Prevalence of obesity, diabetes, and obesity-related health risk factors, 2001. JAMA 2003, 289, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.; Szczepaniak, L.; Dobbins, R.; Nuremberg, P.; Horton, J.; Cohen, J.; Grundy, S.; Hobbs, H. Prevalence of hepatic steatosis in an urban population in the United States: Impact of ethnicity. Hepatology 2004, 40, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.; Younossi, Z. Epidemiology and natural history of NAFLD and NASH. Clin. Liver Dis. 2007, 11, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Johnstone, B.; Cook-Newell, M. Meta-analysis of the effects of soy protein intake on serum lipids. NEJM 1995, 333, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Ascencio, C.; Torres, N.; Isoard-Acosta, F.; Gómez-Pérez, F.; Hernández-Pando, R.; Tovar, A. Soy protein affects serum insulin and hepatic SREBP-1 mRNA and reduces fatty liver in rats. J. Nutr. 2004, 134, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Tovar, A.; Murguía, F.; Cruz, C.; Hernández-Pando, R.; Aguilar-Salinas, C.; Pedraza-Chaverri, J.; Correa-Rotter, R.; Torres, N. A soy protein diet alters hepatic lipid metabolism gene expression and reduces serum lipids and renal fibrogenic cytokines in rats with chronic nephrotic syndrome. J. Nutr. 2002, 132, 2562–2569. [Google Scholar] [CrossRef] [PubMed]
- Zhan, S.; Ho, S. Meta-analysis of the effects of soy protein containing isoflavones on the lipid profile. Am. J. Clin. Nutr. 2005, 81, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Gudbrandsen, O.A.; Wergedahl, H.; Mørk, S.; Liaset, B.; Espe, M.; Berge, R.K. Dietary soya protein concentrate enriched with isoflavones reduced fatty liver, increased hepatic fatty acid oxidation and decreased the hepatic mRNA level of VLDL receptor in obese Zucker rats. Br. J. Nutr. 2007, 96, 249–257. [Google Scholar] [CrossRef]
- Gudbrandsen, O.A.; Wergedahl, H.; Berge, R.K. A casein diet added isoflavone-enriched soy protein favorably affects biomarkers of steatohepatitis in obese Zucker rats. Nutrition 2009, 25, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Wergedahl, H.; Liaset, B.; Gudbrandsen, O.; Lied, E.; Espe, M.; Muna, Z.; Mørk, S.; Berge, R. Fish protein hydrolysate reduces plasma total cholesterol, increases the proportion of HDL cholesterol, and lowers acyl-CoA:cholesterol acyltransferase activity in liver of Zucker rats. J. Nutr. 2004, 134, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Mezei, O.; Banz, W.; Steger, R.; Peluso, M.; Winters, T.; Shay, N. Soy Isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr. 2003, 133, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Tovar, A.; Torre-Villalvazo, I.; Ochoa, M.; Elías, A.; Ortíz, V.; Aguilar-Salinas, C.; Torres, N. Soy protein reduces hepatic lipotoxicity in hyperinsulinemic obese Zucker fa/fa rats. J. Lipid Res. 2005, 46, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Jobgena, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2005, 17, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Hakkak, R.; Al-Dwairi, A.; Fuchs, G.; Korourian, S.; Simmen, F. Dietary soy protein induces hepatic lipogenic enzyme gene expression while suppressing hepatosteatosis in obese female Zucker rats bearing DMBA-initiated mammary tumors. Gen. Nutr. 2012, 7, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Hakkak, R.; Zeng, H.; Dhakal, I.B.; Korourian, S. Short- and long-term soy diet versus casein protects liver steatosis independent of the arginine content. J. Med. Foods 2015, 18, 1274–1280. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Korourian, S.; Zeng, H.; Phelps, J.; Hakkak, R. A Diet Containing a High—Versus Low-Daidzein Level Does Not Protect Against Liver Steatosis in the Obese Female Zucker Rat model. Food Funct. 2017, 8, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Korourian, S.; Hakkak, R.; Ronis, M.J.; Badger, T.M. Diet and risk of ethanol-induced hepatotoxicity: Carbohydrate-fat relationships in rats. Toxicol. Sci. 1999, 47, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Chua, S.C.; Chung, W.K.; Wu-Peng, X.S.; Zhang, Y.; Liu, S.M.; Tartaglia, L.; Leibel, R.L. Phenotypes of mouse diabetes and rat fatty liver due to mutations in the OB (leptin) receptor. Science 1996, 271, 994–996. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, L.A.; Dembski, M.; Weng, X.; Deng, N.; Culpepper, J.; Devos, R.; Richards, G.J.; Campfield, L.A.; Clark, FT.; Deeds, J.; et al. Identification and expression cloning of a leptin receptor, OB-R. Cell 1995, 83, 1263–1271. [Google Scholar] [CrossRef]
- Zucker, L.M. Fat mobilization in vitro and in vivo in the genetically obese Zucker rat “fatty”. J. Lipid Res. 1972, 13, 234–243. [Google Scholar]
- Bray, G.A.; York, D.A.; Fisler, J.S. Experimental obesity: A homeostatic failure due to defective nutrient stimulation of the sympathetic nervous system. Vitam. Horm. 1989, 45, 1–125. [Google Scholar] [CrossRef] [PubMed]
- Buque, X.; Martinez, M.J.; Cano, A.; Miquilena-Colina, M.E.; Garcia-Monzon, C.; Aspichueta, P.; Ochoa, B. A subset of dysregulated metabolic and survival genes is associated with severity of hepatic steatosis in obese Zucker rats. J. Lipid Res. 2010, 51, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Whitten, P.L.; Patisaul, H.B. Cross-species and interassay comparisons of phytoestrogen action. Environ. Health Perspect. 2001, 109, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Mazur, W. Phyto-oestrogens and Western diseases. Ann. Med. 1997, 29, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Bouker, K.B.; Hilakivi-Clarke, L. Genistein: Does it prevent or promote breast cancer? Environ. Health Perspect. 2000, 108, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.; Cassidy, A. Dietary isoflavones: Biological effects and relevance to human health. J. Nutr. 1999, 129, 758S–767S. [Google Scholar] [CrossRef] [PubMed]
- Cain, J.; Banz, W.J.; Butteiger, D.; Davis, J.E. Soy protein isolate modified metabolic phenotype and hepatic wnt signaling in obese Zucker rats. Horm. Metab. Res. 2011, 43, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Peluso, M.R.; Winters, T.A.; Shanahan, M.F.; Banz, W.J. A cooperative interaction between soy protein and its isoflavone-enriched fraction lowers hepatic lipids in male obese zucker rats and reduced blood platelet sensitivity in male sprague-dawley rats. J. Nutr. 2000, 130, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.J.; Yaegashi, S.; Ahsan, R.; Lightfoot, D.A.; Banz, W.J. Differently abundant mRNAs in rat liver in response to diets containing soy protein isolate. Physiol. Genom. 2002, 11, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Lezmi, S.; Wang, H.; Davis, J.; Banz, W.; Chen, H. Fat accumulation in the liver of obese rats is alleviated by soy protein isolate through β-catenin signaling. Obesity 2014, 22, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Van Nielen, M.; Feskens, E.J.M.; Rietman, A.; Siebelink, E.; Mensink, M. Partly replacing meat protein with soy protein alters insulin resistance and blood lipids in post-menopausal women with abdominal obesity. J. Nutr. 2014, 144, 1423–1426. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Marini, H.; Bitto, A.; Altavilla, D.; Polito, F.; Adamo, E.B.; D’Anna, R.; Arcoraci, V.; Burnett, B.P.; Minutoli, L.; et al. Genistein in the metabolic syndrome: Results of a randomized clinical trial. J. Clin. Endocrinol. Metab. 2013, 98, 3366–3374. [Google Scholar] [CrossRef] [PubMed]
- Hakkak, R.; Shaaf, S.; Jo, C.H.; Macleaod, S.; Korourian, S. Effects of high-isoflavone soy diet vs. casein protein diet and obesity on DMBA-induced mammary tumor development. Oncol. Lett. 2011, 2, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Odbayar, T.O.; Ide, T. A comparative analysis of genistein and daidzein in affecting lipid metabolism in rat liver. J. Clin. Biochem. Nutr. 2009, 44, 223–230. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC | LS | OC | OS | LC vs. LS | LC vs. OC | LS vs. OS | OC vs. OS | |
|---|---|---|---|---|---|---|---|---|
| Final BW (g) | 231 ± 14.6 | 259 ± 14.4 | 439 ± 21.8 | 561 ± 49.2 | 1.0000 | <0.0001 | <0.0001 | <0.0001 |
| Liver weight (g) | 7.6 ± 0.9 | 8.6 ± 0.8 | 33.3 ± 4.4 | 20.2 ± 4.1 | 0.0893 | <0.0001 | <0.0001 | <0.0001 |
| Liver weight (% BW) | 3.3 ± 0.4 | 3.3 ± 0.3 | 7.6 ± 0.7 | 3.6 ± 0.8 | 1.0000 | <0.0001 | 1.0000 | <0.0001 |
| Steatosis score | 0.1 ± 0.4 | 0.0 ± 0.0 | 3.5 ± 0.5 | 1.2 ± 0.4 | 1.0000 | <0.0001 | <0.0001 | <0.0001 |
| Leptin | 14.1 ± 5.3 | 20.0 ± 8.9 | 183.0 ± 22.3 | 206.6 ± 25.7 | 0.4524 | <0.0001 | <0.0001 | 0.2303 |
| AST | 108.1 ± 15.4 | 114.2 ± 17.4 | 222.5 ± 32.1 | 103.4 ± 12.1 | 1.0000 | <0.0001 | 0.5834 | <0.0001 |
| ALT | 38.4 ± 3.7 | 49.4 ± 8.9 | 71.9 ± 12.2 | 56.2 ± 10.2 | 0.0109 | <0.0001 | 0.5823 | 0.0357 |
| LC | LS | OC | OS | LC vs. LS | LC vs. OC | LS vs. OS | OC vs. OS | |
|---|---|---|---|---|---|---|---|---|
| Week 1 | 346 ± 27.4 | 344 ± 23.1 | 344 ± 34.4 | 333 ± 31.7 | 1.0000 | 1.0000 | 1.0000 | 1.0000 |
| Week 2 | 300 ± 16.8 | 310 ± 18.3 | 287 ± 30.8 | 353 ± 22.9 | 1.0000 | 1.0000 | 0.0108 | <0.0001 |
| Week 3 | 279 ± 30.5 | 298 ± 30.4 | 252 ± 19.1 | 324 ± 15.6 | 1.0000 | 1.0000 | 1.0000 | <0.0001 |
| Week 4 | 283 ± 25.7 | 267 ± 37.7 | 236 ± 20.0 | 275 ± 20.3 | 1.0000 | 0.0611 | 1.0000 | 0.2459 |
| Week 5 | 241 ± 31.5 | 271 ± 19.6 | 209 ± 34.7 | 228 ± 21.4 | 1.0000 | 0.9376 | 0.0828 | 1.0000 |
| Week 6 | 226 ± 21.4 | 262 ± 34.1 | 195 ± 22.4 | 204 ± 18.6 | 0.2051 | 0.7391 | 0.0010 | 1.0000 |
| Week 7 | 210 ± 28.4 | 234 ± 19.7 | 181 ± 20.3 | 179 ± 23.7 | 1.0000 | 0.6161 | 0.0007 | 1.0000 |
| Week 8 | 232 ± 25.8 | 232 ± 30.5 | 178 ± 18.2 | 184 ± 19.5 | 1.0000 | 0.0034 | 0.0071 | 1.0000 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hakkak, R.; Gauss, C.H.; Bell, A.; Korourian, S. Short-Term Soy Protein Isolate Feeding Prevents Liver Steatosis and Reduces Serum ALT and AST Levels in Obese Female Zucker Rats. Biomedicines 2018, 6, 55. https://doi.org/10.3390/biomedicines6020055
Hakkak R, Gauss CH, Bell A, Korourian S. Short-Term Soy Protein Isolate Feeding Prevents Liver Steatosis and Reduces Serum ALT and AST Levels in Obese Female Zucker Rats. Biomedicines. 2018; 6(2):55. https://doi.org/10.3390/biomedicines6020055
Chicago/Turabian StyleHakkak, Reza, C. Heath Gauss, Andrea Bell, and Soheila Korourian. 2018. "Short-Term Soy Protein Isolate Feeding Prevents Liver Steatosis and Reduces Serum ALT and AST Levels in Obese Female Zucker Rats" Biomedicines 6, no. 2: 55. https://doi.org/10.3390/biomedicines6020055
APA StyleHakkak, R., Gauss, C. H., Bell, A., & Korourian, S. (2018). Short-Term Soy Protein Isolate Feeding Prevents Liver Steatosis and Reduces Serum ALT and AST Levels in Obese Female Zucker Rats. Biomedicines, 6(2), 55. https://doi.org/10.3390/biomedicines6020055