
Asiatic Acid Alleviates Renal Damage by Upregulating STBD1-Mediated Glycophagy in Diabetic Kidney Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Models
2.2. Glucose Sensitivity and Insulin Tolerance Testing
2.3. Blood Glucose and Urinary Albumin-to-Creatinine Ratio (UACR)
2.4. Reverse Transcription-Quantitative PCR (RT-qPCR) and Western Blot Analysis
2.5. Glycogen Detection
2.6. Electron Microscopy
2.7. Immunohistochemistry
2.8. Hematoxylin-Eosin (HE) Staining and Masson Staining
2.9. Molecular Docking Analysis
2.10. Network Pharmacology
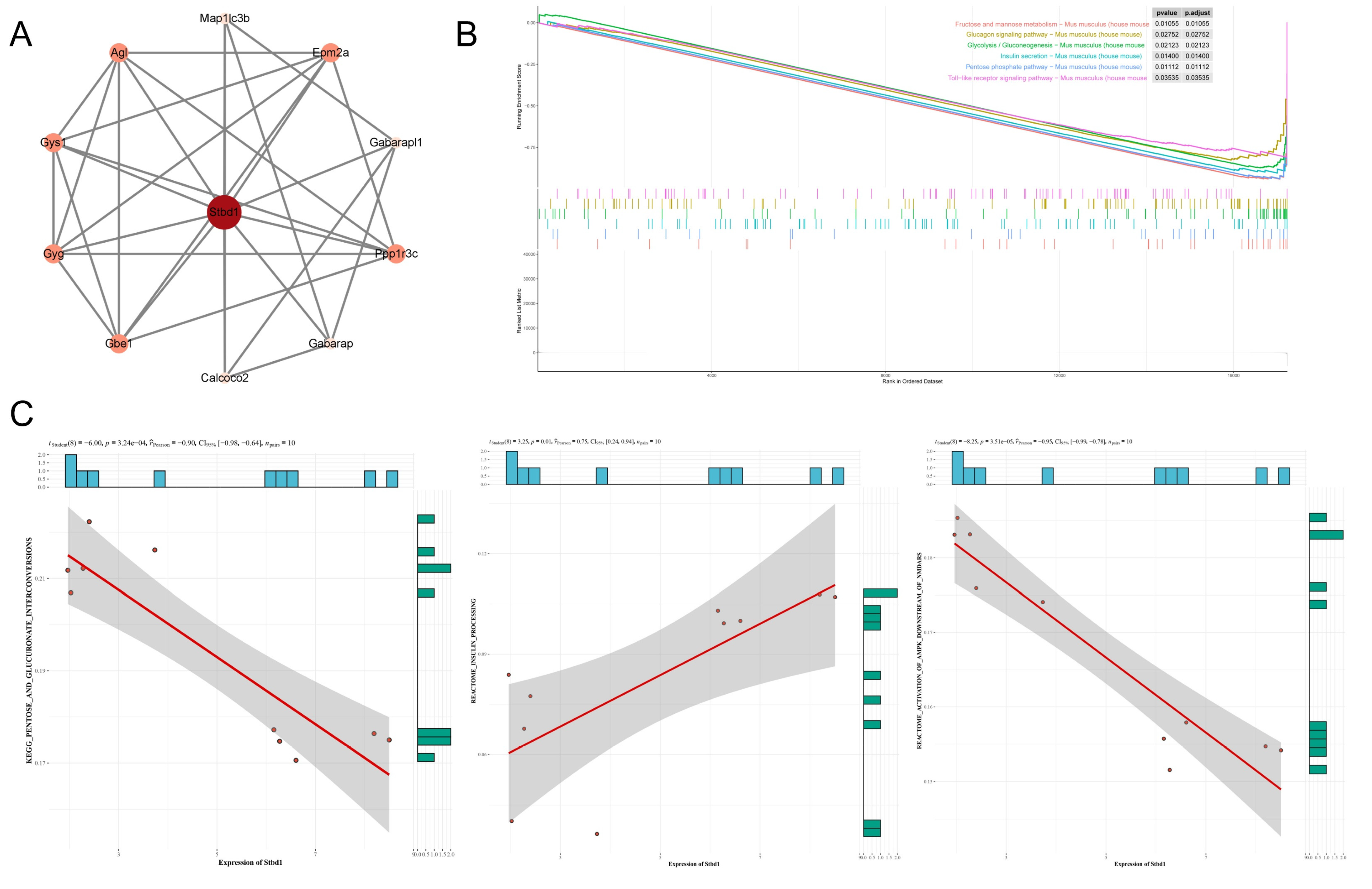
2.11. Gene Set Enrichment Analysis
2.12. Statistical Analyses
3. Results
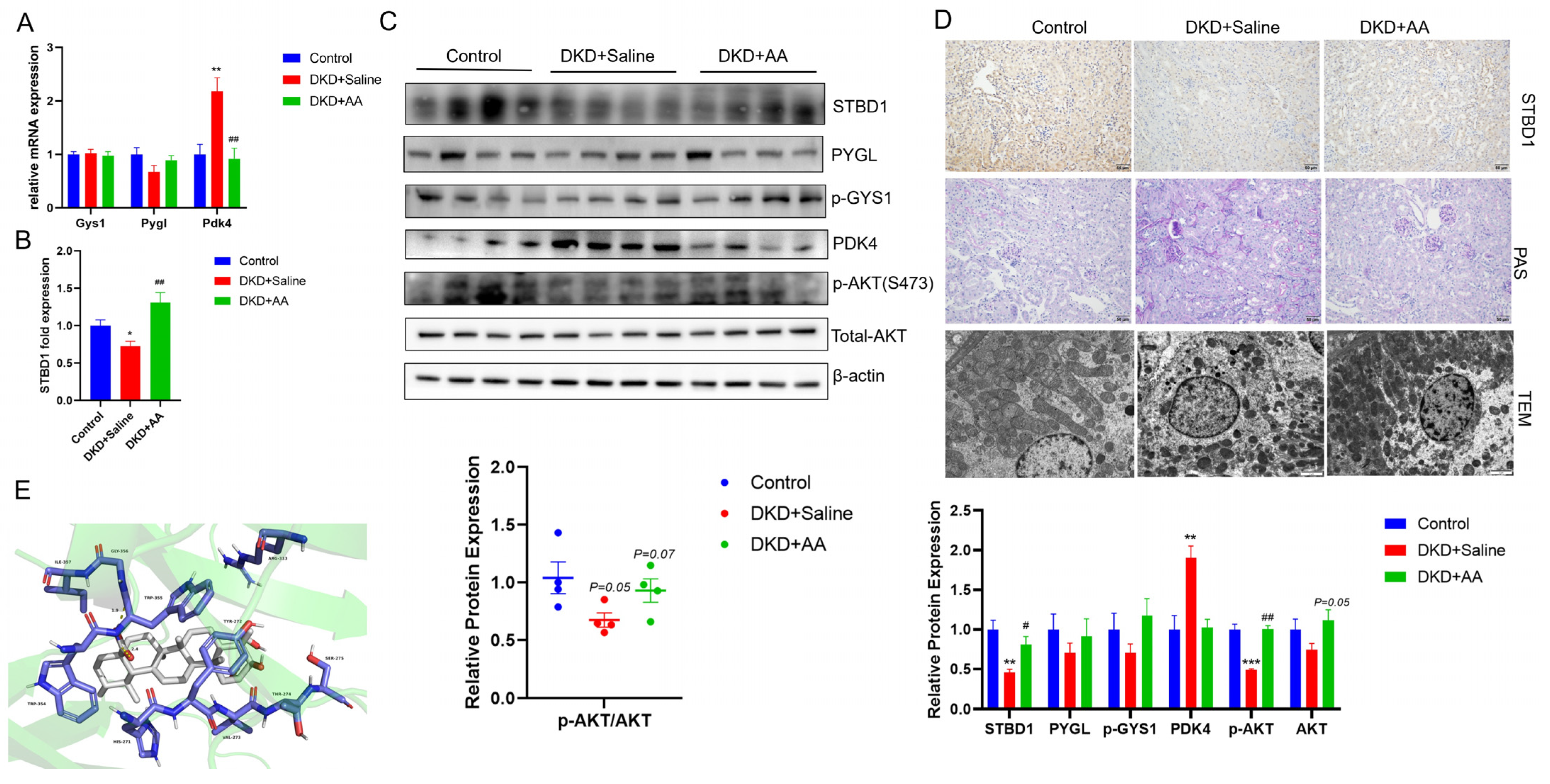
3.1. AA Improves Blood Glucose and UACR in DKD
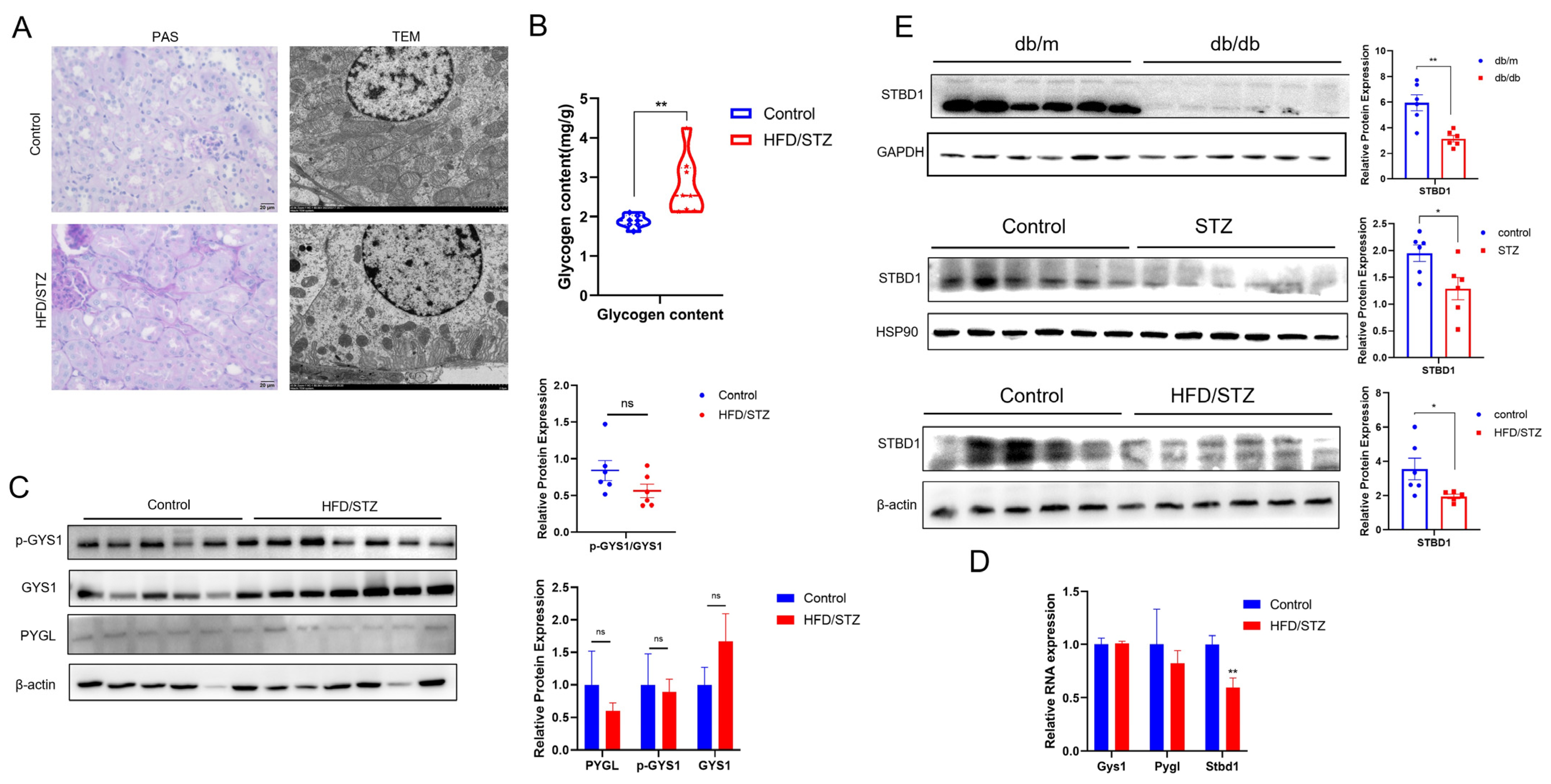
3.2. AA Modulates Kidney Glycogen Metabolism in DKD
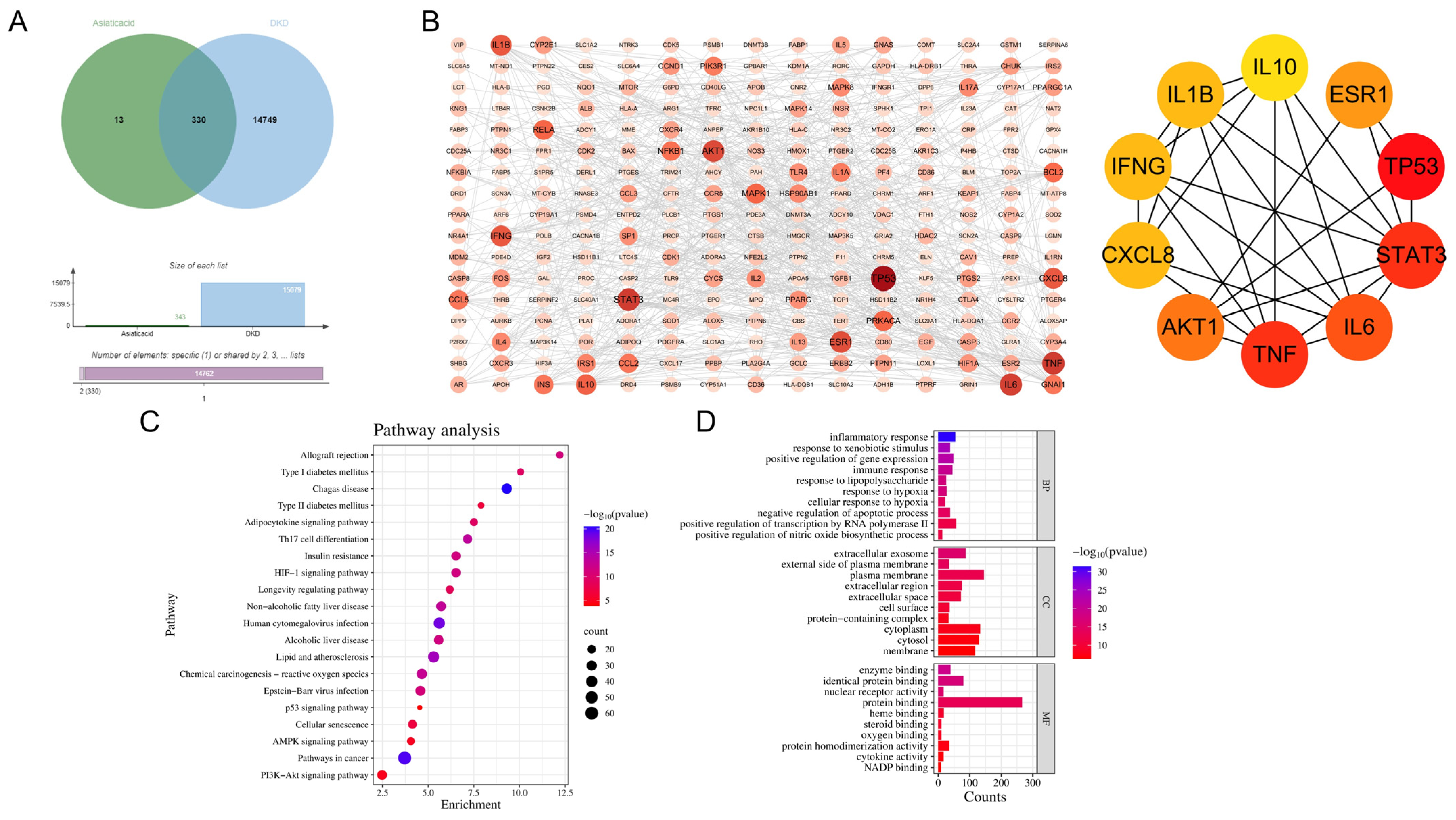
3.3. Network Pharmacology of AA and DKD
3.4. Glycogen Deposition in DKD
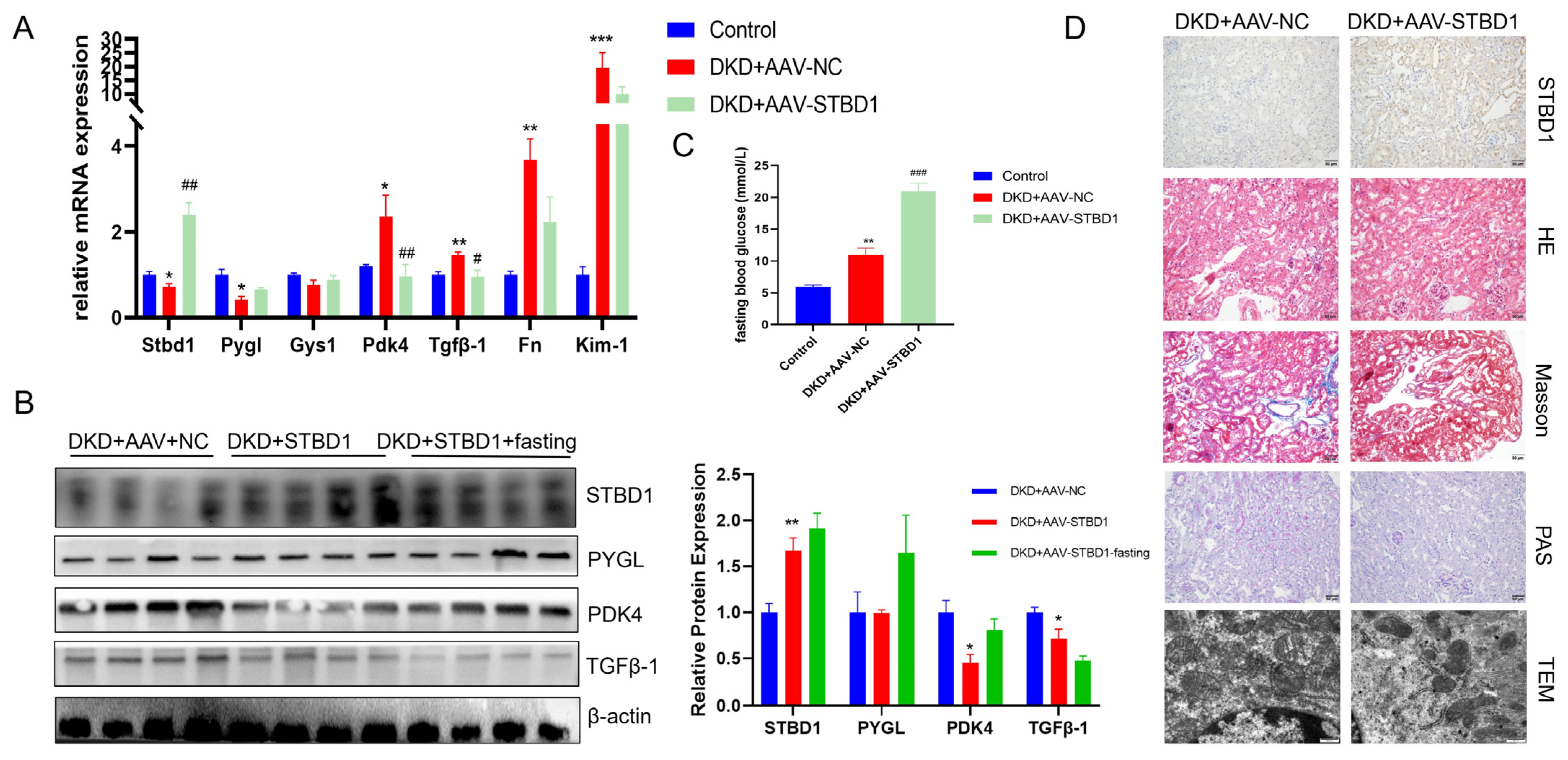
3.5. Modulatory Effects of STBD1 Overexpression in DKD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Asiatic acid |
| AKT | Akt (protein kinase B) |
| DKD | Diabetic kidney disease |
| ESRD | End-stage kidney disease |
| FN | Fibronectin |
| GYS1 | Glycogen synthase 1 |
| HFD | High-fat diet |
| IPGTT | Intraperitoneal glucose tolerance test |
| IPITT | Intraperitoneal insulin tolerance test |
| KIM-1 | Kidney injury molecule-1 |
| p-GYS1 | Phosphorylated glycogen synthase 1 |
| PDK4 | Pyruvate dehydrogenase kinase, isozyme 4 |
| PYGL | Glycogen phosphorylase liver |
| SGLT2i | Sodium-glucose cotransporter 2 inhibitors |
| STBD1 | Starch-binding domain-containing protein 1 |
| STZ | Streptozotocin |
| TGFβ-1 | Transforming growth factor beta-1 |
| UACR | Urinary albumin-to-creatinine ratio |
References
- Alicic, R.; Nicholas, S.B. Diabetic Kidney Disease Back in Focus: Management Field Guide for Health Care Professionals in the 21st Century. Mayo Clin. Proc. 2022, 97, 1904–1919. [Google Scholar] [CrossRef]
- Rayego-Mateos, S.; Rodrigues-Diez, R.R.; Fernandez-Fernandez, B.; Mora-Fernández, C.; Marchant, V.; Donate-Correa, J.; Navarro-González, J.F.; Ortiz, A.; Ruiz-Ortega, M. Targeting Inflammation to Treat Diabetic Kidney Disease: The Road to 2030. Kidney Int. 2023, 103, 282–296. [Google Scholar] [CrossRef]
- Cai, T.; Ke, Q.; Fang, Y.; Wen, P.; Chen, H.; Yuan, Q.; Luo, J.; Zhang, Y.; Sun, Q.; Lv, Y.; et al. Sodium-Glucose Cotransporter 2 Inhibition Suppresses HIF-1α-Mediated Metabolic Switch from Lipid Oxidation to Glycolysis in Kidney Tubule Cells of Diabetic Mice. Cell Death Dis. 2020, 11, 390. [Google Scholar] [CrossRef]
- Natesan, V.; Kim, S.-J. Natural Compounds in Kidney Disease: Therapeutic Potential and Drug Development. Biomol. Ther. 2025, 33, 39–53. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Mandal, S.; Ghorai, M.; Jha, N.K.; Kumar, M.; Radha, null; Ghosh, A.; Proćków, J.; Pérez de la Lastra, J.M.; Dey, A. Therapeutic Properties and Pharmacological Activities of Asiaticoside and Madecassoside: A Review. J. Cell Mol. Med. 2023, 27, 593–608. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, X.; Chen, J.; Song, S.; Fang, S.; Wang, Z.; Xu, S.; Xu, Y.; Liu, J.; Jiang, C.; et al. Asiatic Acid Attenuates Tubular Injury in Diabetic Kidney Disease by Regulating Mitochondrial Dynamics via the Nrf-2 Pathway. Phytomedicine 2023, 109, 154552. [Google Scholar] [CrossRef]
- Huang, C.-F.; Hung, T.-W.; Yang, S.-F.; Tsai, Y.-L.; Yang, J.-T.; Lin, C.-L.; Hsieh, Y.-H. Asiatic Acid from Centella Asiatica Exert Anti-Invasive Ability in Human Renal Cancer Cells by Modulation of ERK/p38MAPK-Mediated MMP15 Expression. Phytomedicine 2022, 100, 154036. [Google Scholar] [CrossRef]
- Wiciński, M.; Fajkiel-Madajczyk, A.; Kurant, Z.; Gajewska, S.; Kurant, D.; Kurant, M.; Sousak, M. Can Asiatic Acid from Centella Asiatica Be a Potential Remedy in Cancer Therapy?-A Review. Cancers 2024, 16, 1317. [Google Scholar] [CrossRef]
- Bylka, W.; Znajdek-Awiżeń, P.; Studzińska-Sroka, E.; Dańczak-Pazdrowska, A.; Brzezińska, M. Centella Asiatica in Dermatology: An Overview. Phytother. Res. 2014, 28, 1117–1124. [Google Scholar] [CrossRef]
- Qiu, F.; Yuan, Y.; Luo, W.; Gong, Y.-S.; Zhang, Z.-M.; Liu, Z.-M.; Gao, L. Asiatic Acid Alleviates Ischemic Myocardial Injury in Mice by Modulating Mitophagy- and Glycophagy-Based Energy Metabolism. Acta Pharmacol. Sin. 2022, 43, 1395–1407. [Google Scholar] [CrossRef]
- Ritchie, S.; Waugh, D. The Pathology of Armanni-Ebstein Diabetic Nephropathy. Am. J. Pathol. 1957, 33, 1035–1057. [Google Scholar] [PubMed]
- Sullivan, M.A.; Forbes, J.M. Glucose and Glycogen in the Diabetic Kidney: Heroes or Villains? EBioMedicine 2019, 47, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Chen, D.; Wang, Q.; Guo, F.; Wei, F.; Zhang, W.; Gan, T.; Luo, Y.; Fan, X.; Du, P.; et al. Canagliflozin Regulates Metabolic Reprogramming in Diabetic Kidney Disease by Inducing Fasting-like and Aestivation-like Metabolic Patterns. Diabetologia 2024, 67, 738–754. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Q.; Wang, L.; Tan, X.; Deng, B.; Li, E.; Gilbert, R.G.; Sullivan, M.A. Some Molecular Structural Features of Glycogen in the Kidneys of Diabetic Rats. Carbohydr. Polym. 2020, 229, 115526. [Google Scholar] [CrossRef]
- Holck, P.; Rasch, R. Structure and Segmental Localization of Glycogen in the Diabetic Rat Kidney. Diabetes 1993, 42, 891–900. [Google Scholar] [CrossRef]
- Jiang, S.; Wells, C.D.; Roach, P.J. Starch-Binding Domain-Containing Protein 1 (Stbd1) and Glycogen Metabolism: Identification of the Atg8 Family Interacting Motif (AIM) in Stbd1 Required for Interaction with GABARAPL1. Biochem. Biophys. Res. Commun. 2011, 413, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Prats, C.; Graham, T.E.; Shearer, J. The Dynamic Life of the Glycogen Granule. J. Biol. Chem. 2018, 293, 7089–7098. [Google Scholar] [CrossRef] [PubMed]
- Koutsifeli, P.; Varma, U.; Daniels, L.J.; Annandale, M.; Li, X.; Neale, J.P.H.; Hayes, S.; Weeks, K.L.; James, S.; Delbridge, L.M.D.; et al. Glycogen-Autophagy: Molecular Machinery and Cellular Mechanisms of Glycophagy. J. Biol. Chem. 2022, 298, 102093. [Google Scholar] [CrossRef]
- Mancini, M.C.; Noland, R.C.; Collier, J.J.; Burke, S.J.; Stadler, K.; Heden, T.D. Lysosomal Glucose Sensing and Glycophagy in Metabolism. Trends Endocrinol. Metab. 2023, 34, 764–777. [Google Scholar] [CrossRef]
- Zhao, H.; Tang, M.; Liu, M.; Chen, L. Glycophagy: An Emerging Target in Pathology. Clin. Chim. Acta 2018, 484, 298–303. [Google Scholar] [CrossRef]
- Tang, Q.; Liu, M.; Zhao, H.; Chen, L. Glycogen-Binding Protein STBD1: Molecule and Role in Pathophysiology. J. Cell Physiol. 2023, 238, 2010–2025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, N.; Liu, M.; Li, X.; Zhou, L.; Huang, W.; Xu, Z.; Liu, J.; Musi, N.; DeFronzo, R.A.; et al. Disruption of Growth Factor Receptor-Binding Protein 10 in the Pancreas Enhances β-Cell Proliferation and Protects Mice from Streptozotocin-Induced β-Cell Apoptosis. Diabetes 2012, 61, 3189–3198. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.-Y.; Zhang, H.-B.; Fan, C.-X.; Zeng, Y.-M.; Zou, S.-Z.; Wu, C.-Y.; Wang, L.; Fang, S.; Li, P.; Xue, Y.-M.; et al. Renoprotective Effects of Brown Adipose Tissue Activation in Diabetic Mice. J. Diabetes 2019, 11, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Chang, D.-Y.; Zhao, M.-H.; Chen, M. Dapagliflozin Alleviates Diabetic Kidney Disease via Hypoxia Inducible Factor 1α/Heme Oxygenase 1-Mediated Ferroptosis. Antioxid. Redox Signal. 2024, 40, 492–509. [Google Scholar] [CrossRef]
- Reidy, K.; Kang, H.M.; Hostetter, T.; Susztak, K. Molecular Mechanisms of Diabetic Kidney Disease. J. Clin. Investig. 2014, 124, 2333–2340. [Google Scholar] [CrossRef]
- Wang, M.; Pang, Y.; Guo, Y.; Tian, L.; Liu, Y.; Shen, C.; Liu, M.; Meng, Y.; Cai, Z.; Wang, Y.; et al. Metabolic Reprogramming: A Novel Therapeutic Target in Diabetic Kidney Disease. Front. Pharmacol. 2022, 13, 970601. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, D.; Zhang, Z.; Zhang, Y.; Zhang, Y.; Kang, X.; Jiang, L.; Tong, X.; Lian, F. Effects of Antioxidants on Diabetic Kidney Diseases: Mechanistic Interpretations and Clinical Assessment. Chin. Med. 2023, 18, 3. [Google Scholar] [CrossRef]
- Masola, B.; Oguntibeju, O.O.; Oyenihi, A.B. Centella Asiatica Ameliorates Diabetes-Induced Stress in Rat Tissues via Influences on Antioxidants and Inflammatory Cytokines. Biomed. Pharmacother. 2018, 101, 447–457. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Goyal, S.N.; Suchal, K.; Sharma, C.; Patil, C.R.; Ojha, S.K. Pharmacological Properties, Molecular Mechanisms, and Pharmaceutical Development of Asiatic Acid: A Pentacyclic Triterpenoid of Therapeutic Promise. Front. Pharmacol. 2018, 9, 892. [Google Scholar] [CrossRef]
- He, J.; Chen, H.; Zhou, W.; Chen, M.; Yao, Y.; Zhang, Z.; Tan, N. Kidney Targeted Delivery of Asiatic Acid Using a FITC Labeled Renal Tubular-Targeting Peptide Modified PLGA-PEG System. Int. J. Pharm. 2020, 584, 119455. [Google Scholar] [CrossRef] [PubMed]
- Haase, V.H. Got Glycogen? An Energy Resource in HIF-Mediated Prevention of Ischemic Kidney Injury. Kidney Int. 2020, 97, 645–647. [Google Scholar] [CrossRef]
- Kang, J.; Dai, X.-S.; Yu, T.-B.; Wen, B.; Yang, Z.-W. Glycogen Accumulation in Renal Tubules, a Key Morphological Change in the Diabetic Rat Kidney. Acta Diabetol. 2005, 42, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Mellor, K.M.; Varma, U.; Stapleton, D.I.; Delbridge, L.M.D. Cardiomyocyte Glycophagy Is Regulated by Insulin and Exposure to High Extracellular Glucose. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1240-1245. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, M.E.; Mellor, K.M.; Curl, C.L.; Stapleton, D.; Delbridge, L.M.D. Myocardial Glycophagy—A Specific Glycogen Handling Response to Metabolic Stress Is Accentuated in the Female Heart. J. Mol. Cell Cardiol. 2013, 65, 67–75. [Google Scholar] [CrossRef]
- Mellor, K.M.; Varma, U.; Koutsifeli, P.; Daniels, L.J.; Benson, V.L.; Annandale, M.; Li, X.; Nursalim, Y.; Janssens, J.V.; Weeks, K.L.; et al. Myocardial Glycophagy Flux Dysregulation and Glycogen Accumulation Characterize Diabetic Cardiomyopathy. J. Mol. Cell Cardiol. 2024, 189, 83–89. [Google Scholar] [CrossRef]
- Sun, T.; Yi, H.; Yang, C.; Kishnani, P.S.; Sun, B. Starch Binding Domain-Containing Protein 1 Plays a Dominant Role in Glycogen Transport to Lysosomes in Liver. J. Biol. Chem. 2016, 291, 16479–16484. [Google Scholar] [CrossRef]
- Heden, T.D.; Chow, L.S.; Hughey, C.C.; Mashek, D.G. Regulation and Role of Glycophagy in Skeletal Muscle Energy Metabolism. Autophagy 2022, 18, 1078–1089. [Google Scholar] [CrossRef]
- Kyriakoudi, S.; Theodoulou, A.; Potamiti, L.; Schumacher, F.; Zachariou, M.; Papacharalambous, R.; Kleuser, B.; Panayiotidis, M.I.; Drousiotou, A.; Petrou, P.P. Stbd1-Deficient Mice Display Insulin Resistance Associated with Enhanced Hepatic ER-Mitochondria Contact. Biochimie 2022, 200, 172–183. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Wu, P.; Feng, Q.; Lin, X.; Wang, Y.; Wu, M.; Cai, F.; Zhang, J.; Yang, C.; Li, X.; et al. Asiatic Acid Alleviates Renal Damage by Upregulating STBD1-Mediated Glycophagy in Diabetic Kidney Disease. Biomedicines 2025, 13, 1544. https://doi.org/10.3390/biomedicines13071544
Guo L, Wu P, Feng Q, Lin X, Wang Y, Wu M, Cai F, Zhang J, Yang C, Li X, et al. Asiatic Acid Alleviates Renal Damage by Upregulating STBD1-Mediated Glycophagy in Diabetic Kidney Disease. Biomedicines. 2025; 13(7):1544. https://doi.org/10.3390/biomedicines13071544
Chicago/Turabian StyleGuo, Lei, Peili Wu, Qijian Feng, Xiaochun Lin, Yuan Wang, Minghai Wu, Feifei Cai, Jin Zhang, Chuyi Yang, Xuelin Li, and et al. 2025. "Asiatic Acid Alleviates Renal Damage by Upregulating STBD1-Mediated Glycophagy in Diabetic Kidney Disease" Biomedicines 13, no. 7: 1544. https://doi.org/10.3390/biomedicines13071544
APA StyleGuo, L., Wu, P., Feng, Q., Lin, X., Wang, Y., Wu, M., Cai, F., Zhang, J., Yang, C., Li, X., Wen, C., Lin, Y., Liu, N., Hu, Y., Wang, H., Fan, X., & Guan, M. (2025). Asiatic Acid Alleviates Renal Damage by Upregulating STBD1-Mediated Glycophagy in Diabetic Kidney Disease. Biomedicines, 13(7), 1544. https://doi.org/10.3390/biomedicines13071544