

Computational Neuroscience’s Influence on Autism Neuro-Transmission Research: Mapping Serotonin, Dopamine, GABA, and Glutamate





Abstract

1. Introduction
2. Theoretical Knowledge
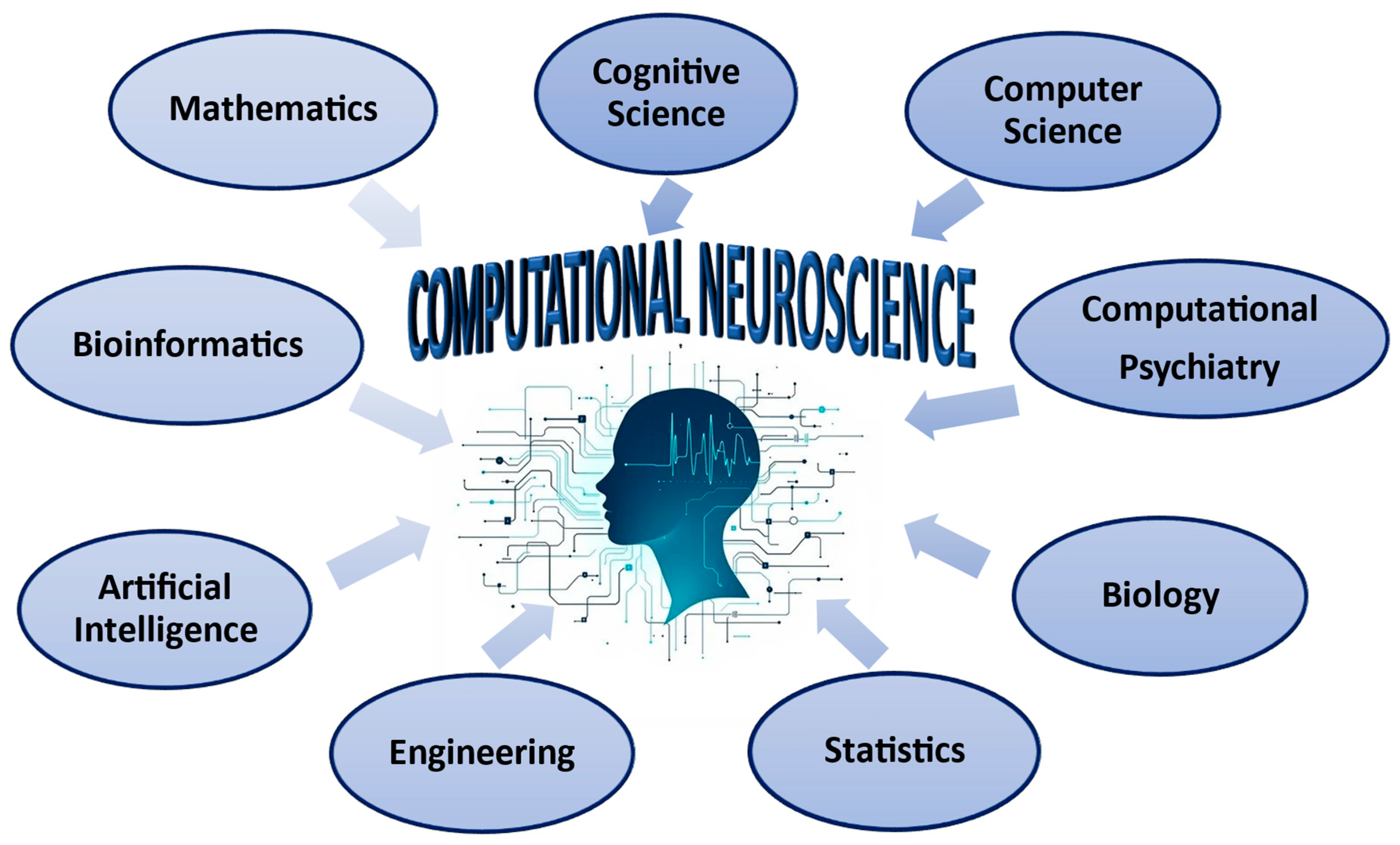
2.1. Computational Neuroscience
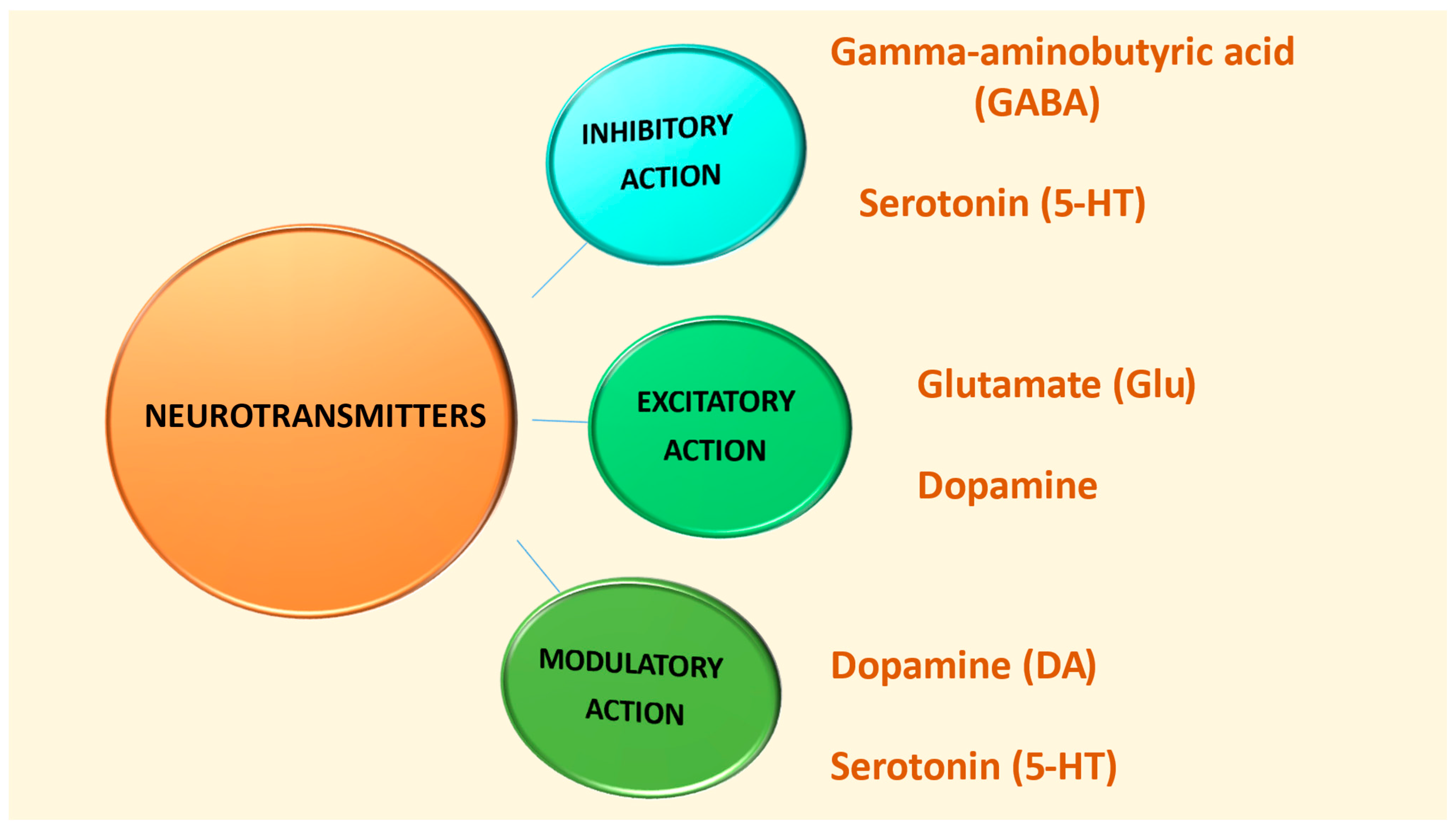
2.2. Neurotransmitters
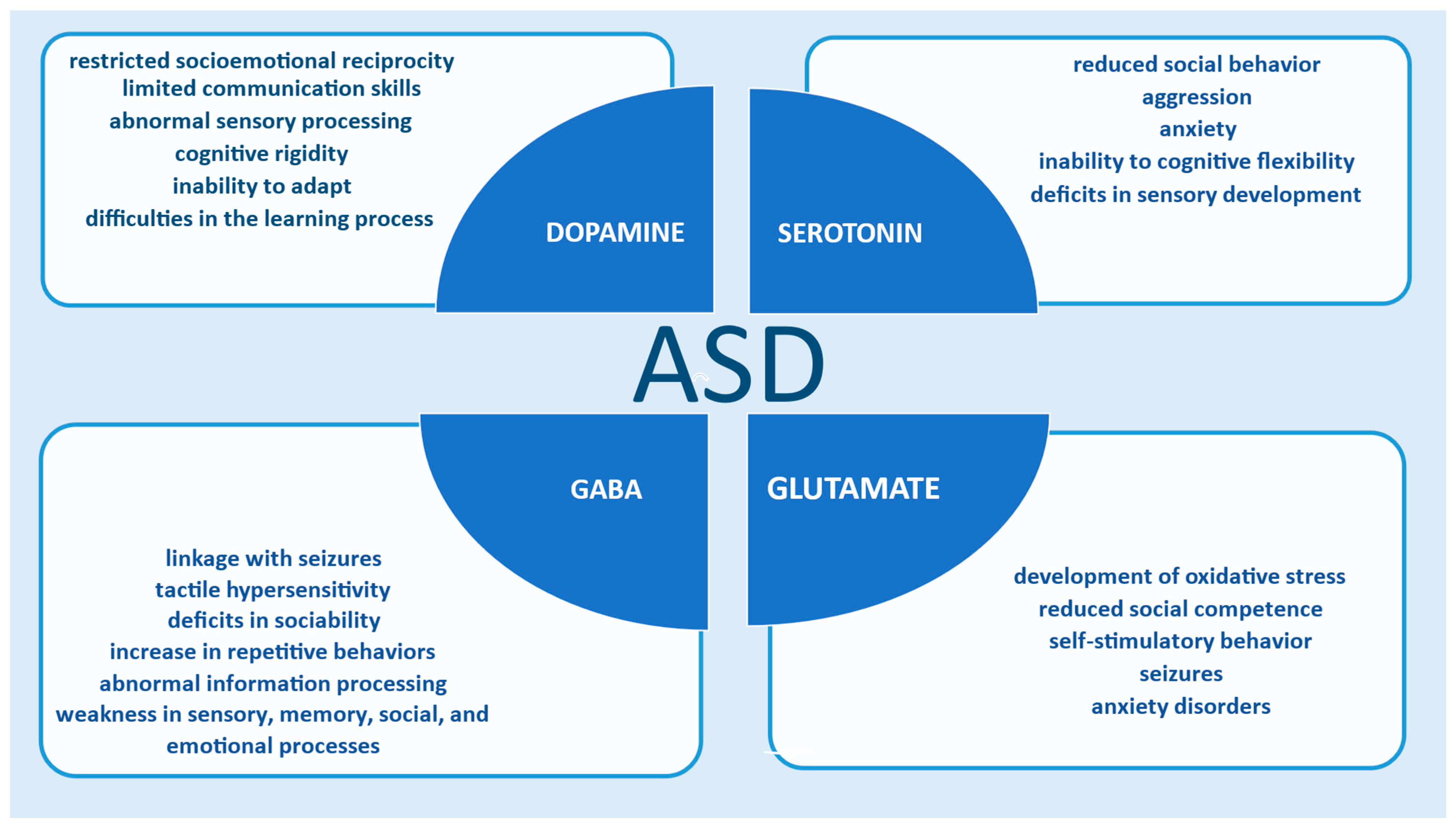
2.3. Autism
3. The Computational Neuroscience Approach to Understanding Neurotransmitter Function in ASD
4. Serotonergic System in ASD and the Use of Computational Methods
5. Dopaminergic System in ASD and the Use of Computational Methods
6. Glutamatergic and GABAergic Systems in ASD and the Use of Computational Methods
7. Computational Approaches’ Contribution to the Identification and Management of ASD
8. Discussion
Limitations and Suggestions for Future Research
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kriegeskorte, N.; Douglas, P.K. Cognitive computational neuroscience. Nat. Neurosci. 2018, 21, 1148–1160. [Google Scholar] [CrossRef] [PubMed]
- Sejnowski, T.; Koch, C.; Churchland, P. Computational Neuroscience. Science 1988, 241, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F. A Quantitative Description of Membrane Current and Its Application to Conduction and Excitation in Nerve. Physiol. J. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed]
- Rall, W. Branching Dendritic Trees and Motoneuron Membrane Resistivity. Exp. Neurol. 1959, 1, 491–527. [Google Scholar] [CrossRef]
- Mäki-Marttunen, T.; Kaufmann, T.; Elvsåshagen, T.; Devor, A.; Djurovic, S.; Westlye, L.T.; Linne, M.-L.; Rietschel, M.; Schubert, D.; Borgwardt, S.; et al. Biophysical Psychiatry—How Computational Neuroscience Can Help to Understand the Complex Mechanisms of Mental Disorders. Front. Psychiatry 2019, 10, 534. [Google Scholar] [CrossRef]
- Sancristóbal, B.; Tastekin, I.; Dierssen, M. Computational models: How do they help to understand neurologic diseases? In Molecular-Genetic and Statistical Techniques for Behavioral and Neural Research; Academic Press: Cambridge, MA, USA; Elsevier: San Diego, CA, USA, 2018; pp. 105–131. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders DSM-5; American Psychiatric Association: Washington, DC, USA, 2013; Volume 5. [Google Scholar] [CrossRef]
- Montanari, M.; Martella, G.; Bonsi, P.; Meringolo, M. Autism Spectrum Disorder: Focus on Glutamatergic Neurotransmission. Int. J. Mol. Sci. 2022, 23, 3861. [Google Scholar] [CrossRef]
- Pergantis, P.; Bamicha, V.; Doulou, A.; Christou, A.I.; Bardis, N.; Skianis, C.; Drigas, A. Assistive and Emerging Technologies to Detect and Reduce Neurophysiological Stress and Anxiety in Children and Adolescents with Autism and Sensory Processing Disorders: A Systematic Review. Technologies 2025, 13, 144. [Google Scholar] [CrossRef]
- Bamicha, V.; Pergantis, P.; Drigas, A. The Effect of Gut Microbiome, Neurotransmitters, and Digital Insights in Autism. Appl. Microbiol. 2024, 4, 1677–1701. [Google Scholar] [CrossRef]
- Bamicha, V.; Salapata, Y. LLLT Applications May Enhance ASD Aspects Related to Disturbances in the Gut Microbiome, Mitochondrial Activity, and Neural Network Function. Braz. J. Sci. 2023, 3, 140–158. [Google Scholar] [CrossRef]
- Salgado-Cacho, J.M.; Moreno-Jiménez, M.d.P.; de Diego-Otero, Y. Detection of Early Warning Signs in Autism Spectrum Disorders: A Systematic Review. Children 2021, 8, 164. [Google Scholar] [CrossRef]
- Randolph-Gips, M.; Srinivasan, P. Modeling autism: A system biology approach. J. Clin. Bioinform. 2012, 2, 17. [Google Scholar] [CrossRef]
- Friston, K. Computational psychiatry: From synapses to sentience. Mol. Psychiatry 2022, 28, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, D.; Boiani, G.M.; Bigiani, A.; Mapelli, J. Modeling Neurotransmission: Computational Tools to Investigate Neurological Disorders. Int. J. Mol. Sci. 2021, 22, 4565. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Wolff, J.J.; Steinbach, M.S.; Doyle, C.B.; Kumar, V.; Elison, J.T. Neurodevelopmental Heterogeneity and Computational Approaches for Understanding Autism. Transl. Psychiatry 2019, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, K.; He, X.; Zhou, J.; Jin, C.; Shen, L.; Gao, Y.; Tian, M.; Zhang, H. Structural, Functional, and Molecular Imaging of Autism Spectrum Disorder. Neurosci. Bull. 2021, 37, 1051–1071. [Google Scholar] [CrossRef]
- Cetin, F.H.; Tunca, H.; Güney, E.; Iseri, E. Neurotransmitter Systems in Autism Spectrum Disorder; Tech: Houston, TX, USA, 2015; pp. 15–30. [Google Scholar] [CrossRef]
- Huys, Q.J.M.; Maia, T.V.; Frank, M.J. Computational Psychiatry as a Bridge from Neuroscience to Clinical Applications. Nat. Neurosci. 2016, 19, 404–413. [Google Scholar] [CrossRef]
- Collins, J.A.; Fauser, B.C. Balancing the strengths of systematic and narrative reviews. Hum. Reprod. Update 2005, 11, 103–104. [Google Scholar] [CrossRef]
- Snyder, H. Literature Review as a Research Methodology: An Overview and Guidelines. J. Bus. Res. 2019, 104, 333–339. [Google Scholar] [CrossRef]
- Greenhalgh, T.; Thorne, S.; Malterud, K. Time to Challenge the Spurious Hierarchy of Systematic over Narrative Reviews? Eur. J. Clin. Investig. 2018, 48, e12931. [Google Scholar] [CrossRef]
- Bamicha, V.; Drigas, A. Strengthening AI via ToM and MC Dimensions. Sci. Electron. Arch. 2024, 17. [Google Scholar] [CrossRef]
- Yahata, N.; Kasai, K.; Kawato, M. Computational Neuroscience Approach to Biomarkers and Treatments for Mental Disorders. Psychiatry Clin. Neurosci. 2017, 71, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Valton, V.; Romaniuk, L.; Steele, J.D.; Lawrie, S.; Seriès, P. Comprehensive Review: Computational Modelling of Schizophrenia. Neurosci. Biobehav. Rev. 2017, 83, 631–646. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Jirsa, V.K. The Quest for Multiscale Brain Modeling. Trends Neurosci. 2022, 45, 777–790. [Google Scholar] [CrossRef]
- Kietzmann, T.C.; McClure, P.; Kriegeskorte, N. Deep Neural Networks in Computational Neuroscience. In Oxford Research Encyclopedia of Neuroscience; Oxford University Press: Oxford, UK, 2019. [Google Scholar] [CrossRef]
- Surianarayanan, C.; Lawrence, J.J.; Chelliah, P.R.; Prakash, E.; Hewage, C. Convergence of Artificial Intelligence and Neuroscience towards the Diagnosis of Neurological Disorders—A Scoping Review. Sensors 2023, 23, 3062. [Google Scholar] [CrossRef]
- Hauser, T.U.; Skvortsova, V.; De Choudhury, M.; Koutsouleris, N. The Promise of a Model-Based Psychiatry: Building Computational Models of Mental Ill Health. Lancet Digit. Health 2022, 4, e816–e828. [Google Scholar] [CrossRef]
- Barnfield, A. Role of Neurotransmitters. In Encyclopedia of Personality and Individual Differences; Zeigler-Hill, V., Shackelford, T., Eds.; Springer: Cham, Switzerland, 2019; pp. 1–7. [Google Scholar] [CrossRef]
- Kapalka, G.M. Substances Involved in Neurotransmission. In Practical Resources for the Mental Health Professional, Nutritional and Herbal Therapies for Children and Adolescents; Kapalka, G.M., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: San Diego, CA, USA, 2010; pp. 71–99. [Google Scholar] [CrossRef]
- Rieder, R.; Wisniewski, P.J.; Alderman, B.L.; Campbell, S.C. Microbes and Mental Health: A Review. Brain Behav. Immun. 2017, 66, 9–17. [Google Scholar] [CrossRef]
- Hussain, S.; Iltaf, M.; Nazir, E.; Hamid, A.; Shamim, S.; Khan, N.; Bibi, H.; Rasheed, S.J.; Khan, F.D. A Comprehensive Review on Neurotransmitters, Its Biological Importance and Methods of Analysis. J. Mol. Genet. 2023, 4, 11–41. [Google Scholar] [CrossRef]
- Dhailappan, A.; Samiappan, S. Impact of Diet on Neurotransmitters. In Role of Nutrients in Neurological Disorders; Rajagopal, S., Ramachandran, S., Sundararaman, G., Gadde Venkata, S., Eds.; Springer: Singapore, 2022; pp. 363–383. [Google Scholar] [CrossRef]
- Kanova, M.; Kohout, P. Serotonin—Its Synthesis and Roles in the Healthy and the Critically Ill. Int. J. Mol. Sci. 2021, 22, 4837. [Google Scholar] [CrossRef] [PubMed]
- Best, J.; Nijhout, H.F.; Reed, M. Serotonin Synthesis, Release and Reuptake in Terminals: A Mathematical Model. Theor. Biol. Med. Model. 2010, 7, 34. [Google Scholar] [CrossRef]
- Hohmann, C.F.; Walker, E.M.; Boylan, C.B.; Blue, M.E. Neonatal Serotonin Depletion Alters Behavioral Responses to Spatial Change and Novelty. Brain Res. 2007, 1139, 163–177. [Google Scholar] [CrossRef]
- Rodnyy, A.Y.; Kondaurova, E.M.; Tsybko, A.S.; Popova, N.K.; Kudlay, D.A.; Naumenko, V.S. The Brain Serotonin System in Autism. Rev. Neurosci. 2023, 35, 1–20. [Google Scholar] [CrossRef]
- Muller, C.L.; Anacker, A.M.J.; Veenstra-VanderWeele, J. The Serotonin System in Autism Spectrum Disorder: From Biomarker to Animal Models. Neuroscience 2016, 321, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Ciranna, L. Serotonin as a Modulator of Glutamate- and GABA-Mediated Neurotransmission: Implications in Physiological Functions and in Pathology. Curr. Neuropharmacol. 2006, 4, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Ayano, G. Dopamine: Receptors, Functions, Synthesis, Pathways, Locations and Mental Disorders: Review of Literatures. J. Nerv. Ment. Dis. 2016, 2, 2. [Google Scholar] [CrossRef]
- Pizzarelli, R.; Cherubini, E. Alterations of GABAergic Signaling in Autism Spectrum Disorders. Neural Plast. 2011, 2011, 297153. [Google Scholar] [CrossRef]
- Zhao, H.; Mao, X.; Zhu, C.; Zou, X.; Peng, F.; Yang, W.; Li, B.; Li, G.; Ge, T.; Cui, R. GABAergic System Dysfunction in Autism Spectrum Disorders. Front. Cell Dev. Biol. 2022, 9, 781327. [Google Scholar] [CrossRef]
- Rojas, D.C. The Role of Glutamate and Its Receptors in Autism and the Use of Glutamate Receptor Antagonists in Treatment. J. Neural Transm. 2014, 121, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The Neurochemistry of Autism. Brain Sci. 2020, 10, 163. [Google Scholar] [CrossRef]
- Shimmura, C.; Suzuki, K.; Iwata, Y.; Tsuchiya, K.J.; Ohno, K.; Matsuzaki, H.; Iwata, K.; Kameno, Y.; Takahashi, T.; Wakuda, T.; et al. Enzymes in the Glutamate-Glutamine Cycle in the Anterior Cingulate Cortex in Postmortem Brain of Subjects with Autism. Mol. Autism 2013, 4, 6. [Google Scholar] [CrossRef]
- Fernández, M.; Mollinedo-Gajate, I.; Peñagarikano, O. Neural Circuits for Social Cognition: Implications for Autism. Neuroscience 2018, 370, 148–162. [Google Scholar] [CrossRef]
- Al-Otaish, H.; Al-Ayadhi, L.; Bjørklund, G.; Chirumbolo, S.; Urbina, M.A.; El-Ansary, A. Relationship between Absolute and Relative Ratios of Glutamate, Glutamine and GABA and Severity of Autism Spectrum Disorder. Metab. Brain Dis. 2018, 33, 843–854. [Google Scholar] [CrossRef] [PubMed]
- El-Ansary, A.; Al-Ayadhi, L. GABAergic/Glutamatergic Imbalance Relative to Excessive Neuroinflammation in Autism Spectrum Disorders. J. Neuroinflamm. 2014, 11, 189. [Google Scholar] [CrossRef] [PubMed]
- Kosillo, P.; Bateup, H.S. Dopaminergic Dysregulation in Syndromic Autism Spectrum Disorders: Insights from Genetic Mouse Models. Front. Neural Circuits 2021, 15, 700968. [Google Scholar] [CrossRef] [PubMed]
- Abg Abd Wahab, D.Y.; Gau, C.H.; Zakaria, R.; Muthu Karuppan, M.K.; A-rahbi, B.S.; Abdullah, Z.; Alrafiah, A.; Abdullah, J.M.; Muthuraju, S. Review on Cross Talk between Neurotransmitters and Neuroinflammation in Striatum and Cerebellum in the Mediation of Motor Behaviour. Biomed Res. Int. 2019, 2019, 1767203. [Google Scholar] [CrossRef]
- Tartaglione, A.M.; Camoni, L.; Calamandrei, G.; Chiarotti, F.; Venerosi, A. The Contribution of Environmental Pollutants to the Risk of Autism and Other Neurodevelopmental Disorders: A Systematic Review of Case-Control Studies. Neurosci. Biobehav. Rev. 2024, 164, 105815. [Google Scholar] [CrossRef]
- Pagalan, L.; Bickford, C.; Weikum, W.; Lanphear, B.; Brauer, M.; Lanphear, N.; Hanley, G.E.; Oberlander, T.F.; Winters, M. Association of Prenatal Exposure to Air Pollution with Autism Spectrum Disorder. JAMA Pediatr. 2019, 173, 86. [Google Scholar] [CrossRef]
- Tau, G.Z.; Peterson, B.S. Normal Development of Brain Circuits. Neuropsychopharmacology 2009, 35, 147–168. [Google Scholar] [CrossRef]
- Yin, W.; Chen, M.-H.; Hung, S.-C.; Baluyot, K.R.; Li, T.; Lin, W. Brain Functional Development Separates into Three Distinct Time Periods in the First Two Years of Life. NeuroImage 2019, 189, 715–726. [Google Scholar] [CrossRef]
- Chaidi, I.; Pergantis, P.; Drigas, A.; Karagiannidis, C. Gaming Platforms for People with ASD. J. Intell. 2024, 12, 122. [Google Scholar] [CrossRef]
- Boylan, C.; Blue, M.; Hohmann, C. Modeling Early Cortical Serotonergic Deficits in Autism. Behav. Brain Res. 2007, 176, 94–108. [Google Scholar] [CrossRef]
- Chattopadhyaya, B.; Cristo, G.D. GABAergic Circuit Dysfunctions in Neurodevelopmental Disorders. Front. Psychiatry 2012, 3, 21012. [Google Scholar] [CrossRef] [PubMed]
- Pergantis, P.; Drigas, A. Sensory Integration Therapy as Enabler for Developing Emotional Intelligence in Children with Autism Spectrum Disorder and the ICT’s Role. Braz. J. Sci. 2023, 2, 53–65. [Google Scholar] [CrossRef]
- Pergantis, P.; Drigas, A. Assistive Technology for Autism Spectrum Disorder Children That Experiences Stress and Anxiety. Braz. J. Sci. 2023, 2, 77–93. [Google Scholar] [CrossRef]
- Chatterjee, R.; Paluh, J.L.; Chowdhury, S.; Mondal, S.; Raha, A.; Mukherjee, A. SyNC, a Computationally Extensive and Realistic Neural Net to Identify Relative Impacts of Synaptopathy Mechanisms on Glutamatergic Neurons and Their Networks in Autism and Complex Neurological Disorders. Front. Cell. Neurosci. 2021, 15, 674030. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, M.; Zhou, L.; Wang, X.; Jia, J. Cognitive Psychology-Based Artificial Intelligence Review. Front. Neurosci. 2022, 16, 1024316. [Google Scholar] [CrossRef]
- Macpherson, T.; Churchland, A.; Sejnowski, T.; DiCarlo, J.; Kamitani, Y.; Takahashi, H.; Hikida, T. Natural and Artificial Intelligence: A Brief Introduction to the Interplay between AI and Neuroscience Research. Neural Netw. 2021, 144, 603–613. [Google Scholar] [CrossRef]
- Lake, B.M.; Ullman, T.D.; Tenenbaum, J.B.; Gershman, S.J. Building Machines That Learn and Think like People. Behav. Brain Sci. 2016, 40, e253. [Google Scholar] [CrossRef]
- Duch, W.; Nowak, W.; Meller, J.; Osiński, G.; Dobosz, K.; Mikołajewski, D.; Wójcik, G.M. Computational approach to understanding autism spectrum disorders. Comput. Sci. 2012, 13, 47. [Google Scholar] [CrossRef]
- Wang, S.S.-H.; Kloth, A.D.; Badura, A. The Cerebellum, Sensitive Periods, and Autism. Neuron 2014, 83, 518–532. [Google Scholar] [CrossRef]
- Krishnan, A.; Zhang, R.; Yao, V.; Theesfeld, C.L.; Wong, A.K.; Tadych, A.; Volfovsky, N.; Packer, A.; Lash, A.; Troyanskaya, O.G. Genome-Wide Prediction and Functional Characterization of the Genetic Basis of Autism Spectrum Disorder. Nat. Neurosci. 2016, 19, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Satterstrom, F.K.; Kosmicki, J.A.; Wang, J.; Breen, M.S.; De Rubeis, S.; An, J.-Y.; Peng, M.; Collins, R.; Grove, J.; Klei, L.; et al. Large-Scale Exome Sequencing Study Implicates Both Developmental and Functional Changes in the Neurobiology of Autism. Cell 2020, 180, 568–584.e23. [Google Scholar] [CrossRef]
- Muhle, R.A.; Reed, H.E.; Stratigos, K.A.; Veenstra-VanderWeele, J. The Emerging Clinical Neuroscience of Autism Spectrum Disorder. JAMA Psychiatry 2018, 75, 514. [Google Scholar] [CrossRef]
- Piccinini, G.; Shagrir, O. Foundations of Computational. Curr. Opin. Neurobiol. 2014, 25, 25–30. [Google Scholar] [CrossRef]
- De Schutter, E. Why Are Computational Neuroscience and Systems Biology so Separate? PLoS Comput. Biol. 2008, 4, e1000078. [Google Scholar] [CrossRef]
- Vladusich, T. Towards a Computational Neuroscience of Autism-Psychosis Spectrum Disorders. Behav. Brain Sci. 2008, 31, 282–283. [Google Scholar] [CrossRef]
- Olson, A.; Zhang, F.; Cao, H.; Ancha Baranova; Slavin, M. In Silico Gene Set and Pathway Enrichment Analyses Highlight Involvement of Ion Transport in Cholinergic Pathways in Autism: Rationale for Nutritional Intervention. Front. Neurosci. 2021, 15, 648410. [Google Scholar] [CrossRef]
- Lord, L.-D.; Stevner, A.B.; Deco, G.; Kringelbach, M.L. Understanding Principles of Integration and Segregation Using Whole-Brain Computational Connectomics: Implications for Neuropsychiatric Disorders. Philos. Trans. A Math. Phys. Eng. Sci. 2017, 375, 20160283. [Google Scholar] [CrossRef]
- Robertson, C.E.; Baron-Cohen, S. Sensory Perception in Autism. Nat. Rev. Neurosci. 2017, 18, 671–684. [Google Scholar] [CrossRef]
- Thomas, M.S.C.; Johnson, M.H. The Computational Modeling of Sensitive Periods. Dev. Psychobiol. 2006, 48, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Freche, D.; Lee, C.-Y.; Rouach, N.; Holcman, D. Synaptic Transmission in Neurological Disorders Dissected by a Quantitative Approach. Commun. Integr. Biol. 2012, 5, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wang, J.; Zhang, Z.; Jin, X.; Xu, Y.; Si, Y.; Liang, Y.; Ge, Y.; Zhan, H.; Peng, L.; et al. Shank3 Deficiency Elicits Autistic-like Behaviors by Activating P38α in Hypothalamic AgRP Neurons. Mol. Autism 2024, 15, 14. [Google Scholar] [CrossRef]
- Uchino, S.; Waga, C. SHANK3 as an Autism Spectrum Disorder-Associated Gene. Brain Dev. 2013, 35, 106–110. [Google Scholar] [CrossRef]
- Masoli, S.; Rizza, M.F.; Tognolina, M.; Prestori, F.; D’Angelo, E. Computational Models of Neurotransmission at Cerebellar Synapses Unveil the Impact on Network Computation. Front. Comput. Neurosci. 2022, 16, 1006989. [Google Scholar] [CrossRef]
- Gilbert, T.L. The Allen Brain Atlas as a Resource for Teaching Undergraduate Neuroscience. J. Undergrad. Neurosci. Educ. 2018, 16, A261. Available online: https://pmc.ncbi.nlm.nih.gov/articles/PMC6153011 (accessed on 7 March 2025).
- Martin, H.; Choi, J.E.; Rodrigues, A.R.; Eshel, N. Review: Dopamine, Serotonin, and the Translational Neuroscience of Aggression in Autism Spectrum Disorder. J. Am. Acad. Child Adolesc. Psychiatry 2024, 3, 29–41. [Google Scholar] [CrossRef]
- Lee, A.; Choo, H.; Jeon, B. Serotonin Receptors as Therapeutic Targets for Autism Spectrum Disorder Treatment. Int. J. Mol. Sci. 2022, 23, 6515. [Google Scholar] [CrossRef]
- Kareva, I. Serotonin Signaling in the Enteric Nervous System and Connection to Autism Spectrum Disorder: A Translational Mathematical Model. Appl. Sci. 2023, 13, 2970. [Google Scholar] [CrossRef]
- Azmitia, E.C.; Singh, J.S.; Whitaker-Azmitia, P.M. Increased Serotonin Axons (Immunoreactive to 5-HT Transporter) in Postmortem Brains from Young Autism Donors. Neuropharmacology 2011, 60, 1347–1354. [Google Scholar] [CrossRef]
- Janušonis, S.; Haiman, J.H.; Metzler, R.; Vojta, T. Predicting the Distribution of Serotonergic Axons: A Supercomputing Simulation of Reflected Fractional Brownian Motion in a 3D-Mouse Brain Model. Front. Comput. Neurosci. 2023, 17, 1189853. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, S.; Li, Q.; Li, B. Fringe Projection Profilometry by Conducting Deep Learning from Its Digital Twin. Opt. Express 2020, 28, 36568. [Google Scholar] [CrossRef] [PubMed]
- Mishro, P.K.; Agrawal, S.; Panda, R.; Abraham, A. Novel Fuzzy Clustering-Based Bias Field Correction Technique for Brain Magnetic Resonance Images. IET Image Process. 2020, 14, 1929–1936. [Google Scholar] [CrossRef]
- Hu, M.; Zhong, Y.; Xie, S.; Lv, H.; Lv, Z. Fuzzy System Based Medical Image Processing for Brain Disease Prediction. Front. Neurosci. 2021, 15, 714318. [Google Scholar] [CrossRef]
- Tan, Z.; Wei, H.; Song, X.; Mai, W.; Yan, J.; Ye, W.; Ling, X.; Hou, L.; Zhang, S.; Yan, S.; et al. Positron Emission Tomography in the Neuroimaging of Autism Spectrum Disorder: A Review. Front. Neurosci. 2022, 16, 806876. [Google Scholar] [CrossRef]
- Shinde, A.V.; Patil, D.D. A Multi-Classifier-Based Recommender System for Early Autism Spectrum Disorder Detection Using Machine Learning. Healthc. Anal. 2023, 4, 100211. [Google Scholar] [CrossRef]
- Pereira, F.; Mitchell, T.; Botvinick, M. Machine Learning Classifiers and FMRI: A Tutorial Overview. NeuroImage 2009, 45, S199–S209. [Google Scholar] [CrossRef]
- Itahashi, T.; Yamashita, A.; Takahara, Y.; Noriaki Yahata; Aoki, Y.Y.; Fujino, J.; Yoshihara, Y.; Nakamura, M.; Aoki, R.; Okimura, T.; et al. Generalizable and Transportable Resting-State Neural Signatures Characterized by Functional Networks, Neurotransmitters, and Clinical Symptoms in Autism. Mol. Psychiatry 2024, 30, 1466–1478. [Google Scholar] [CrossRef] [PubMed]
- Pavăl, D. A dopamine hypothesis of autism spectrum disorder. Dev. Neurosci. 2017, 39, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Asl, M.M.; Vahabie, A.-H.; Valizadeh, A. Dopaminergic Modulation of Synaptic Plasticity, Its Role in Neuropsychiatric Disorders, and Its Computational Modeling. Basic Clin. Neurosci. 2018, 10, 1–12. [Google Scholar] [CrossRef]
- Béroule, D.G. Offline Encoding Impaired by Epigenetic Regulations of Monoamines in the Guided Propagation Model of Autism. BMC Neurosci. 2018, 19, 80. [Google Scholar] [CrossRef]
- Hass, J.; Durstewitz, D. Models of Dopaminergic Modulation. Scholarpedia 2011, 6, 4215. [Google Scholar] [CrossRef]
- Montague, P.R.; Hyman, S.E.; Cohen, J.D. Computational Roles for Dopamine in Behavioural Control. Nature 2004, 431, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.X.; Frank, M.J. Neurocomputational Models of Basal Ganglia Function in Learning, Memory and Choice. Behav. Brain Res. 2009, 199, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Kriete, T.; Noelle, D.C. Dopamine and the Development of Executive Dysfunction in Autism Spectrum Disorders. PLoS ONE 2015, 10, e0121605. [Google Scholar] [CrossRef] [PubMed]
- Cartier, E.; Hamilton, P.J.; Belovich, A.N.; Shekar, A.; Campbell, N.G.; Saunders, C.; Andreassen, T.F.; Gether, U.; Veenstra-Vanderweele, J.; Sutcliffe, J.S.; et al. Rare Autism-Associated Variants Implicate Syntaxin 1 (STX1 R26Q) Phosphorylation and the Dopamine Transporter (HDAT R51W) in Dopamine Neurotransmission and Behaviors. EBioMedicine 2015, 2, 135–146. [Google Scholar] [CrossRef]
- Binda, F.; Dipace, C.; Bowton, E.; Robertson, S.D.; Lute, B.J.; Fog, J.U.; Zhang, M.; Sen, N.; Colbran, R.J.; Gnegy, M.E.; et al. Syntaxin 1A Interaction with the Dopamine Transporter Promotes Amphetamine-Induced Dopamine Efflux. Mol. Pharmacol. 2008, 74, 1101–1108. [Google Scholar] [CrossRef]
- Zald, D.H. Dopamine Transmission in the Human Striatum during Monetary Reward Tasks. J. Neurosci. 2004, 24, 4105–4112. [Google Scholar] [CrossRef]
- Makkonen, I.; Riikonen, R.; Kokki, H.; Airaksinen, M.M.; Kuikka, J.T. Serotonin and Dopamine Transporter Binding in Children with Autism Determined by SPECT. Dev. Med. Child Neurol. 2008, 50, 593–597. [Google Scholar] [CrossRef]
- Haker, H.; Schneebeli, M.; Stephan, K.E. Can Bayesian Theories of Autism Spectrum Disorder Help Improve Clinical Practice? Front. Psychiatry 2016, 7, 107. [Google Scholar] [CrossRef]
- Nakamura, K.; Sekine, Y.; Ouchi, Y.; Tsujii, M.; Yoshikawa, E.; Futatsubashi, M.; Tsuchiya, K.J.; Sugihara, G.; Iwata, Y.; Suzuki, K.; et al. Brain Serotonin and Dopamine Transporter Bindings in Adults with High-Functioning Autism. Arch. Gen. Psychiatry 2010, 67, 59. [Google Scholar] [CrossRef]
- Foss-Feig, J.H.; Adkinson, B.D.; Ji, J.L.; Yang, G.; Srihari, V.H.; McPartland, J.C.; Krystal, J.H.; Murray, J.D.; Anticevic, A. Searching for Cross-Diagnostic Convergence: Neural Mechanisms Governing Excitation and Inhibition Balance in Schizophrenia and Autism Spectrum Disorders. Biol. Psychiatry 2017, 81, 848–861. [Google Scholar] [CrossRef]
- Rosenberg, A.; Patterson, J.S.; Angelaki, D.E. A Computational Perspective on Autism. Proc. Natl. Acad. Sci. USA 2015, 112, 9158–9165. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Cline, H.T. What Is Excitation/Inhibition and How Is It Regulated? A Case of the Elephant and the Wisemen. J. Exp. Neurosci. 2019, 13, 117906951985937. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, J.L.R.; Merzenich, M.M. Model of Autism: Increased Ratio of Excitation/Inhibition in Key Neural Systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Abu, A.; Al-Ayadhi, L.Y.; Al-Dbass, A.M.; El-Ansary, A.K. Mechanism of Nitrogen Metabolism-Related Parameters and Enzyme Activities in the Pathophysiology of Autism. J. Neurodev. Disord. 2012, 4, 4. [Google Scholar] [CrossRef]
- Harada, M.; Taki, M.M.; Nose, A.; Kubo, H.; Mori, K.; Nishitani, H.; Matsuda, T. Non-Invasive Evaluation of the GABAergic/Glutamatergic System in Autistic Patients Observed by MEGA-Editing Proton MR Spectroscopy Using a Clinical 3 Tesla Instrument. J. Autism Dev. Disord. 2011, 41, 447–454. [Google Scholar] [CrossRef]
- Vattikuti, S.; Chow, C.C. A Computational Model for Cerebral Cortical Dysfunction in Autism Spectrum Disorders. Biol. Psychiatry 2010, 67, 672–678. [Google Scholar] [CrossRef]
- Deco, G.; Kringelbach, M.L.; Aurina Arnatkeviciute; Oldham, S.; Sabaroedin, K.; Rogasch, N.C.; Aquino, K.M.; Fornito, A. Dynamical Consequences of Regional Heterogeneity in the Brain’s Transcriptional Landscape. Sci. Adv. 2021, 7, eabf4752. [Google Scholar] [CrossRef]
- Park, B.; Hong, S.-J.; Valk, S.L.; Paquola, C.; Benkarim, O.; Bethlehem, R.A.I.; Di Martino, A.; Milham, M.P.; Gozzi, A.; Yeo, B.T.T.; et al. Differences in Subcortico-Cortical Interactions Identified from Connectome and Microcircuit Models in Autism. Nat. Commun. 2021, 12, 2225. [Google Scholar] [CrossRef]
- Schirner, M.; McIntosh, A.R.; Jirsa, V.; Deco, G.; Ritter, P. Inferring Multi-Scale Neural Mechanisms with Brain Network Modelling. eLife 2018, 7, e28927. [Google Scholar] [CrossRef]
- Breakspear, M. Dynamic Models of Large-Scale Brain Activity. Nat. Neurosci. 2017, 20, 340–352. [Google Scholar] [CrossRef]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate Receptor Ion Channels: Structure, Regulation, and Function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed]
- Nisar, S.; Bhat, A.A.; Masoodi, T.; Hashem, S.; Akhtar, S.; Ali, T.A.; Amjad, S.; Chawla, S.; Bagga, P.; Frenneaux, M.P.; et al. Genetics of Glutamate and Its Receptors in Autism Spectrum Disorder. Mol. Psychiatry 2022, 27, 2380–2392. [Google Scholar] [CrossRef] [PubMed]
- Gandal, M.J.; Anderson, R.L.; Billingslea, E.N.; Carlson, G.C.; Roberts, T.P.L.; Siegel, S.J. Mice with Reduced NMDA Receptor Expression: More Consistent with Autism than Schizophrenia? Genes Brain Behav. 2012, 11, 740–750. [Google Scholar] [CrossRef]
- Fatemi, S.H. The Hyperglutamatergic Hypothesis of Autism. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 911. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.E.; Jeon, O.H.; Zimmerman, A.W.; Blue, M.E.; Pevsner, J. Postmortem Brain Abnormalities of the Glutamate Neurotransmitter System in Autism. Neurology 2001, 57, 1618–1628. [Google Scholar] [CrossRef]
- Hanada, T. Ionotropic Glutamate Receptors in Epilepsy: A Review Focusing on AMPA and NMDA Receptors. Biomolecules 2020, 10, 464. [Google Scholar] [CrossRef]
- Tuchman, R. What Is the Relationship between Autism Spectrum Disorders and Epilepsy? Semin. Pediatr. Neurol. 2017, 24, 292–300. [Google Scholar] [CrossRef]
- Ferrante, M.; Blackwell, K.T.; Migliore, M.; Ascoli, G.A. Computational Models of Neuronal Biophysics and the Characterization of Potential Neuropharmacological Targets. Curr. Med. Chem. 2008, 15, 2456–2471. [Google Scholar] [CrossRef]
- Goulas, A.; Changeux, J.-P.; Wagstyl, K.; Amunts, K.; Palomero-Gallagher, N.; Hilgetag, C.C. The Natural Axis of Transmitter Receptor Distribution in the Human Cerebral Cortex. Proc. Natl. Acad. Sci. USA 2021, 118, e2020574118. [Google Scholar] [CrossRef]
- Mohammad, F.K.; Palukuri, M.V.; Shivakumar, S.; Rengaswamy, R.; Sahoo, S. A Computational Framework for Studying Gut-Brain Axis in Autism Spectrum Disorder. Front. Physiol. 2022, 13, 760753. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol. Clin. N. Am. 2017, 46, 77–89. [Google Scholar] [CrossRef]
- Horn, J.; Mayer, D.E.; Chen, S.; Mayer, E.A. Role of Diet and Its Effects on the Gut Microbiome in the Pathophysiology of Mental Disorders. Transl. Psychiatry 2022, 12, 164. [Google Scholar] [CrossRef]
- Shanmugam, H.; Ganguly, S.; Priya, B. Plant Food Bioactives and Its Effects on Gut Microbiota Profile Modulation for Better Brain Health and Functioning in Autism Spectrum Disorder Individuals: A Review. Food Front. 2022, 3, 124–141. [Google Scholar] [CrossRef]
- Eicher, T.P.; Mohajeri, M.H. Overlapping Mechanisms of Action of Brain-Active Bacteria and Bacterial Metabolites in the Pathogenesis of Common Brain Diseases. Nutrients 2022, 14, 2661. [Google Scholar] [CrossRef]
- Hollestein, V.; Poelmans, G.; Forde, N.J.; Beckmann, C.F.; Ecker, C.; Mann, C.; Schäfer, T.; Moessnang, C.; Baumeister, S.; Banaschewski, T.; et al. Excitatory/Inhibitory Imbalance in Autism: The Role of Glutamate and GABA Gene-Sets in Symptoms and Cortical Brain Structure. Transl. Psychiatry 2023, 13, 18. [Google Scholar] [CrossRef]
- Leithead, A.B.; Tasker, J.G.; Harony-Nicolas, H. The Interplay between Glutamatergic Circuits and Oxytocin Neurons in the Hypothalamus and Its Relevance to Neurodevelopmental Disorders. J. Neuroendocrinol. 2021, 33, e13061. [Google Scholar] [CrossRef] [PubMed]
- Freche, D.; Pannasch, U.; Rouach, N.; Holcman, D. Synapse Geometry and Receptor Dynamics Modulate Synaptic Strength. PLoS ONE 2011, 6, e25122. [Google Scholar] [CrossRef] [PubMed]
- Campagnola, L.; Seeman, S.C.; Chartrand, T.; Kim, L.; Hoggarth, A.; Gamlin, C.; Ito, S.; Trinh, J.; Davoudian, P.A.; Radaelli, C.; et al. Local Connectivity and Synaptic Dynamics in Mouse and Human Neocortex. Science 2022, 375, eabj5861. [Google Scholar] [CrossRef]
- Bristy, S.A.; Islam, A.M.H.; Andalib, K.M.S.; Khan, U.; Awal, M.A.; Rahman, M.H. Determination of Molecular Signatures and Pathways Common to Brain Tissues of Autism Spectrum Disorder: Insights from Comprehensive Bioinformatics Approach. Inform. Med. Unlocked 2022, 29, 100871. [Google Scholar] [CrossRef]
- Syriopoulou-Delli, C.K. Advances in Autism Spectrum Disorder (ASD) Diagnostics: From Theoretical Frameworks to AI-Driven Innovations. Electronics 2025, 14, 951. [Google Scholar] [CrossRef]
- Qin, L.; Wang, H.; Ning, W.; Cui, M.; Wang, Q. New Advances in the Diagnosis and Treatment of Autism Spectrum Disorders. Eur. J. Med. Res. 2024, 29, 322. [Google Scholar] [CrossRef]
- Thabtah, F. Machine Learning in Autistic Spectrum Disorder Behavioral Research: A Review and Ways Forward. Inform. Health Soc. Care 2018, 44, 278–297. [Google Scholar] [CrossRef]
- Moridian, P.; Ghassemi, N.; Jafari, M.; Salloum-Asfar, S.; Sadeghi, D.; Khodatars, M.; Shoeibi, A.; Khosravi, A.; Ling, S.H.; Subasi, A.; et al. Automatic Autism Spectrum Disorder Detection Using Artificial Intelligence Methods with MRI Neuroimaging: A Review. Front. Mol. Neurosci. 2022, 15, 999605. [Google Scholar] [CrossRef]
- Sherkatghanad, Z.; Akhondzadeh, M.; Salari, S.; Zomorodi-Moghadam, M.; Abdar, M.; Acharya, U.R.; Khosrowabadi, R.; Salari, V. Automated Detection of Autism Spectrum Disorder Using a Convolutional Neural Network. Front. Neurosci. 2020, 13, 1325. [Google Scholar] [CrossRef]
- Gao, L.; Wang, Z.; Long, Y.; Zhang, X.; Su, H.; Yu, Y.; Hong, J. Autism Spectrum Disorders Detection Based on Multi-Task Transformer Neural Network. BMC Neurosci. 2024, 25, 27. [Google Scholar] [CrossRef]
- Rajagopalan, S.S.; Zhang, Y.; Yahia, A.; Kristiina Tammimies. Machine Learning Prediction of Autism Spectrum Disorder from a Minimal Set of Medical and Background Information. JAMA Netw. Open 2024, 7, e2429229. [Google Scholar] [CrossRef]
- Song, C.; Jiang, Z.-Q.; Hu, L.-F.; Li, W.-H.; Liu, X.-L.; Wang, Y.-Y.; Jin, W.-Y.; Zhu, Z.-W. A Machine Learning-Based Diagnostic Model for Children with Autism Spectrum Disorders Complicated with Intellectual Disability. Front. Psychiatry 2022, 13, 993077. [Google Scholar] [CrossRef]
- Manglunia, A.; Puranik, A. FDG PET/CT Findings in a Clinically Diagnosed Case of Childhood Autism. Indian J. Nucl. Med. 2016, 31, 138. [Google Scholar] [CrossRef]
- Keeratitanont, K.; Theerakulpisut, D.; Auvichayapat, N.; Suphakunpinyo, C.; Patjanasoontorn, N.; Tiamkao, S.; Tepmongkol, S.; Khiewvan, B.; Raruenrom, Y.; Srisuruk, P.; et al. Brain Laterality Evaluated by F-18 Fluorodeoxyglucose Positron Emission Computed Tomography in Autism Spectrum Disorders. Front. Mol. Neurosci. 2022, 15, 901016. [Google Scholar] [CrossRef] [PubMed]
- Hellmuth, S.G.; Lars Egholm Pedersen; Caroline Borregaard Miltoft; Olav Bjørn Petersen; Kjærgaard, S.; Charlotte Kvist Ekelund; Tabor, A. Increased Nuchal Translucency Thickness and Risk of Neurodevelopmental Disorders. Ultrasound Obstet. Gynecol. 2017, 49, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Kasera, B.; Shinar, S.; Edke, P.; Pruthi, V.; Goldenberg, A.; Erdman, L.; Van Mieghem, T. Deep-Learning Computer Vision Can Identify Increased Nuchal Translucency in the First Trimester of Pregnancy. Prenat. Diagn. 2024, 44, 535–543. [Google Scholar] [CrossRef]
- Nisar, S.; Haris, M. Neuroimaging Genetics Approaches to Identify New Biomarkers for the Early Diagnosis of Autism Spectrum Disorder. Mol. Psychiatry 2023, 28, 4995–5008. [Google Scholar] [CrossRef]
- Dimitrakopoulos, G.N.; Di Miceli, M. Editorial: Bioinformatics for Modern Neuroscience. Front. Comput. Neurosci. 2024, 18, 1385658. [Google Scholar] [CrossRef]
- Thomas, M.S.C.; Fedor, A.; Davis, R.; Yang, J.; Alireza, H.; Charman, T.; Masterson, J.; Best, W. Computational Modeling of Interventions for Developmental Disorders. Psychol. Rev. 2019, 126, 693–726. [Google Scholar] [CrossRef]
- Liu, W.; Pluta, A.; Charpentier, C.J.; Rosenblau, G. A Computational Cognitive Neuroscience Approach for Characterizing Individual Differences in Autism: Introduction to Special Issue. Pers. Neurosci. 2025, 8, e2. [Google Scholar] [CrossRef]
- Bamicha, V.; Drigas, A. Human-Social Robot Interaction in the Light of ToM and Metacognitive Functions. Sci. Electron. Arch. 2024, 17, 1–19. [Google Scholar] [CrossRef]
- Ganggayah, M.D.; Zhao, D.; Liew, J.Y.; Nurul Aqilah; Thayapari Paramasivam; Lee, Y.Y.; Abu, I.; Shaharuddin, S. Accelerating Autism Spectrum Disorder Care: A Rapid Review of Data Science Applications in Diagnosis and Intervention. Asian J. Psychiatr. 2025, 108, 104498. [Google Scholar] [CrossRef]
- Alexopoulou, A.; Pergantis, P.; Koutsojannis, C.; Triantafillou, V.; Drigas, A. Non-Invasive BCI-VR Applied Protocols as Intervention Paradigms on School-Aged Subjects with ASD: A Systematic Review. Sensors 2025, 25, 1342. [Google Scholar] [CrossRef]
- Gupta, N.S.; Kumar, P. Perspective of Artificial Intelligence in Healthcare Data Management: A Journey towards Precision Medicine. Comput. Biol. Med. 2023, 162, 107051. [Google Scholar] [CrossRef]
- Maia, T.V. Introduction to the Series on Computational Psychiatry. Clin. Psychol. Sci. 2015, 3, 374–377. [Google Scholar] [CrossRef]
- Loosen, A.M.; Kato, A.; Gu, X. Revisiting the Role of Computational Neuroimaging in the Era of Integrative Neuroscience. Neuropsychopharmacology 2024, 50, 103–113. [Google Scholar] [CrossRef]
- Lawn, T.; Howard, M.A.; Turkheimer, F.; Misic, B.; Deco, G.; Martins, D.; Dipasquale, O. From Neurotransmitters to Networks: Transcending Organisational Hierarchies with Molecular-Informed Functional Imaging. Neurosci. Biobehav. Rev. 2023, 150, 105193. [Google Scholar] [CrossRef]
- Teleanu, R.I. Neurotransmitters—Key Factors in Neurological and Neurodegenerative Disorders of the Central Nervous System. Int. J. Mol. Sci. 2022, 23, 5954. [Google Scholar] [CrossRef]
- Banerjee, S.; McCracken, S.; Hossain, M.F.; Slaughter, G. Electrochemical detection of neurotransmitters. Biosensors 2020, 10, 101. [Google Scholar] [CrossRef]
- Bakhtiari, R.; Mohammadi Sephavand, N.; Nili Ahmadabadi, M.; Nadjar Araabi, B.; Esteky, H. Computational Model of Excitatory/Inhibitory Ratio Imbalance Role in Attention Deficit Disorders. J. Comput. Neurosci. 2012, 33, 389–404. [Google Scholar] [CrossRef]
- Wegiel, J.; Chadman, K.; London, E.; Wisniewski, T.; Wegiel, J. Contribution of the Serotonergic System to Developmental Brain Abnormalities in Autism Spectrum Disorder. J. Autism 2024, 17, 1300–1321. [Google Scholar] [CrossRef]
- Olia, J.B.H.; Raman, A.; Hsu, C.-Y.; Alkhayyat, A.; Nourazarian, A. A Comprehensive Review of Neurotransmitter Modulation via Artificial Intelligence: A New Frontier in Personalized Neurobiochemistry. Comput. Biol. Med. 2025, 189, 109984. [Google Scholar] [CrossRef]
- Iglesias, S.; Tomiello, S.; Schneebeli, M.; Stephan, K.E. Models of Neuromodulation for Computational Psychiatry. Wiley Interdiscip. Rev. Cogn. 2016, 8, e1420. [Google Scholar] [CrossRef]
- Movassaghi, C.S.; Perrotta, K.A.; Yang, H.; Iyer, R.; Cheng, X.; Dagher, M.; Fillol, M.A.; Andrews, A.M. Simultaneous Serotonin and Dopamine Monitoring across Timescales by Rapid Pulse Voltammetry with Partial Least Squares Regression. Anal. Bioanal. Chem. 2021, 413, 6747–6767. [Google Scholar] [CrossRef]
- Dayan, P.; Durstewitz, D. Computational modelling: After the Middle Ages of computational neuroscience. Curr. Opin. Behav. Sci. 2016, 11, 5. [Google Scholar]
- Pergantis, P.; Bamicha, V.; Skianis, C.; Drigas, A. AI Chatbots and Cognitive Control: Enhancing Executive Functions Through Chatbot Interactions: A Systematic Review. Brain Sci. 2025, 15, 47. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neurotransmitter | Type | Functions | Associated Conditions | Pathways/Mechanisms |
|---|---|---|---|---|
| Serotonin (5-HT) [35,36,37,38,39,40] | Inhibitory, Modulatory | Regulates feeding, sleep, aggression, emotion, body weight, circadian rhythm, reward processing, and plasticity | OCD, anxiety, alcoholism, emotional disorders, ASD, memory, and learning deficits | Produced in dorsal and median raphe nuclei; modulates glutamate and GABA transmission; acts via multiple receptors; impacts cortical synaptic plasticity and morphogenesis |
| Dopamine (DA) [17,41] | Modulatory, Excitatory | Influences behavior, reward processing, emotional regulation, and cognition | ASD, Parkinson’s, emotional dysregulation, aggression, and prolactin dysregulation | Four pathways: Nigrostriatal, Mesolimbic, Mesocortical, and Tuberoinfundibular; overactivity leads to impulsivity and underactivity leads to cognitive deficits |
| Glutamate (Glu) [31,41,46,47,48,49] | Excitatory | Enhances memory and promotes synaptic excitation across brain | ASD, seizures, hypersensitivity, oxidative stress, and sensory/memory/emotional dysfunctions | Synthesized from glutamine; converted in astrocytes; interacts with GABA system; involved in excitatory neurotransmission across nearly all synapses |
| GABA [42,43,44,45,46,47,48,49] | Inhibitory | Regulates cell proliferation, migration, maturation, and death | ASD, information processing challenges, social skill development deficits, seizures, anxiety, and repetitive behaviors | Main inhibitory neurotransmitter; interacts with glutamate; part of GABAergic system; critical to neurodevelopment and neurotransmission balance |
| Neurotransmitter | Dysfunction in ASD | Computational Methods |
|---|---|---|
| Serotonin (5-HT) [36,38,82,83,84,85,86,89,90] | Altered serotogenic signaling; hyperseretonemia in 25–33% of ASD patients; linked to aggression, hyperactivity, sleep, and sensory issues | Mathematical models of serotonergic homeostasis; gut-brain axis modeling; PET data provide information about serotonin function; fMRI classifiers model growth and spatial distribution of axon density |
| Dopamine (DA) [50,94,95,96,97,98,99,100,101,102,103,104,105,106,107] | Abnormal DA signaling affects reward processing, motivation, repetitive behaviors, and executive functions | Reinforcement learning (TDRL); cross-task models (XT); biophysical models; genetic simulations of DA transport (DAT); fMRI and PET with computational models |
| Glutamate (Glu) [108,117,118,119,120,121,122,123,124,134] | Excitatory/inhibitory imbalance; hyperactive NMDA/AMPA receptors; associated with seizures and oxidative stress | Biophysical models of AMPA receptor activity; synaptic learning parameter models; neural field models (NFMs); structural synapse simulations |
| GABA [75,108,109,110,111,112,113,114,115,116,126,127,128,129,130,131,132] | Reduced GABAergic activity; decreased inhibition leads to E/I imbalance and sensory/perceptual abnormalities | Neural circuit simulations; binocular competition modeling; hybrid EEG models; gut microbiome interaction modeling |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamicha, V.; Pergantis, P.; Skianis, C.; Drigas, A. Computational Neuroscience’s Influence on Autism Neuro-Transmission Research: Mapping Serotonin, Dopamine, GABA, and Glutamate. Biomedicines 2025, 13, 1420. https://doi.org/10.3390/biomedicines13061420
Bamicha V, Pergantis P, Skianis C, Drigas A. Computational Neuroscience’s Influence on Autism Neuro-Transmission Research: Mapping Serotonin, Dopamine, GABA, and Glutamate. Biomedicines. 2025; 13(6):1420. https://doi.org/10.3390/biomedicines13061420
Chicago/Turabian StyleBamicha, Victoria, Pantelis Pergantis, Charalabos Skianis, and Athanasios Drigas. 2025. "Computational Neuroscience’s Influence on Autism Neuro-Transmission Research: Mapping Serotonin, Dopamine, GABA, and Glutamate" Biomedicines 13, no. 6: 1420. https://doi.org/10.3390/biomedicines13061420
APA StyleBamicha, V., Pergantis, P., Skianis, C., & Drigas, A. (2025). Computational Neuroscience’s Influence on Autism Neuro-Transmission Research: Mapping Serotonin, Dopamine, GABA, and Glutamate. Biomedicines, 13(6), 1420. https://doi.org/10.3390/biomedicines13061420