
Effect of Recombinant Human Growth Hormone (rhGH) Use on Genetic Methylation Patterns and Their Relationship with Body Composition in Small-for-Gestational-Age (SGA) Newborns

 , ,
, ,  , , , ,
, , , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Collection of Clinical and Sociodemographic Data
2.3. Blood Biochemical Evaluation
2.4. DNA Sample Extraction
2.5. Quantification and Evaluation of DNA Quality
2.6. Epigenetic Sequencing of Human Methylation Profiling
2.7. Bioinformatic Prediction of Protein Interactions
2.8. Structural Modeling and Functional Bioinformatics Analysis
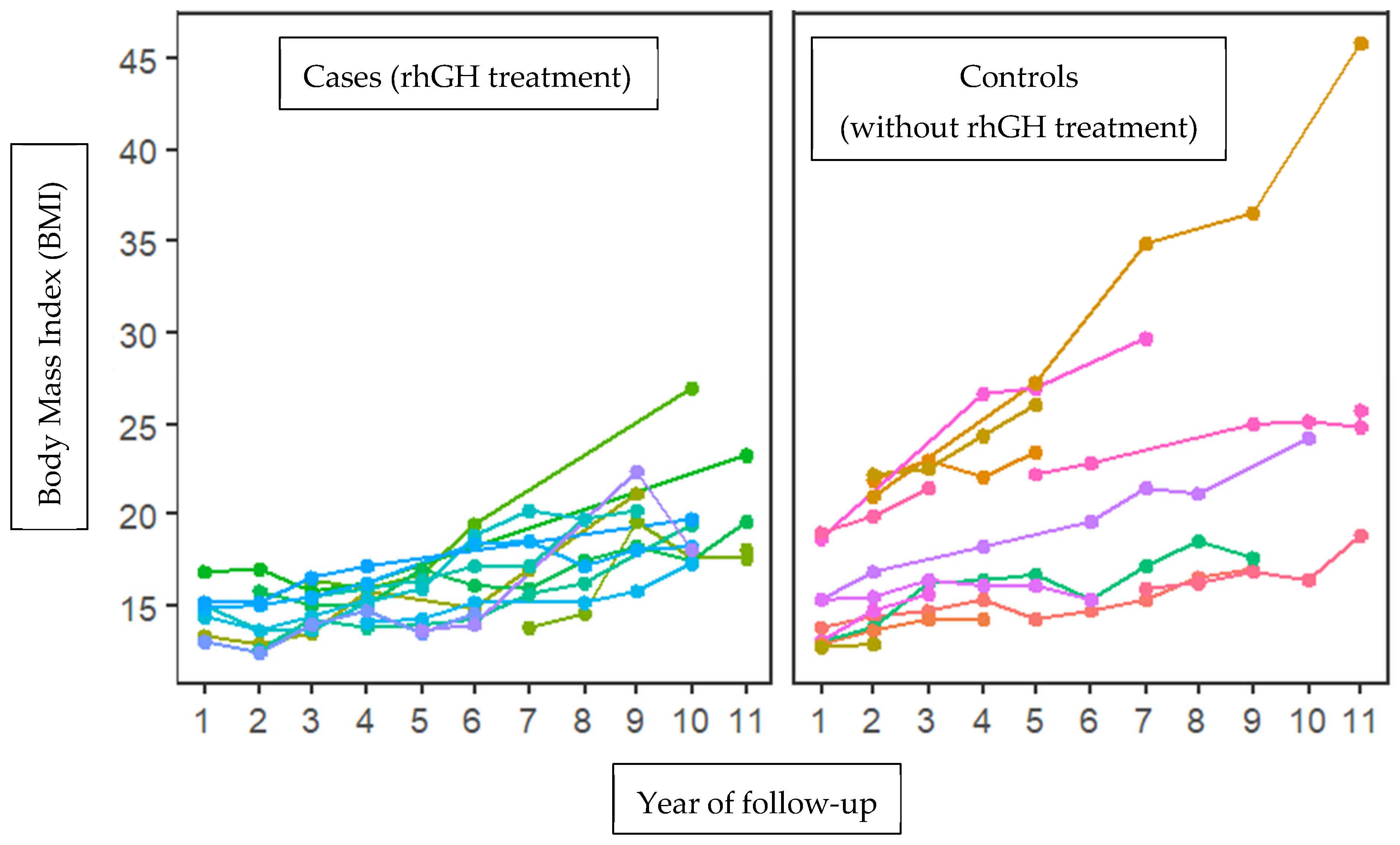
3. Results
3.1. Study Population
3.2. Collection of Clinical and Sociodemographic Data
3.3. Blood Biochemical Evaluation
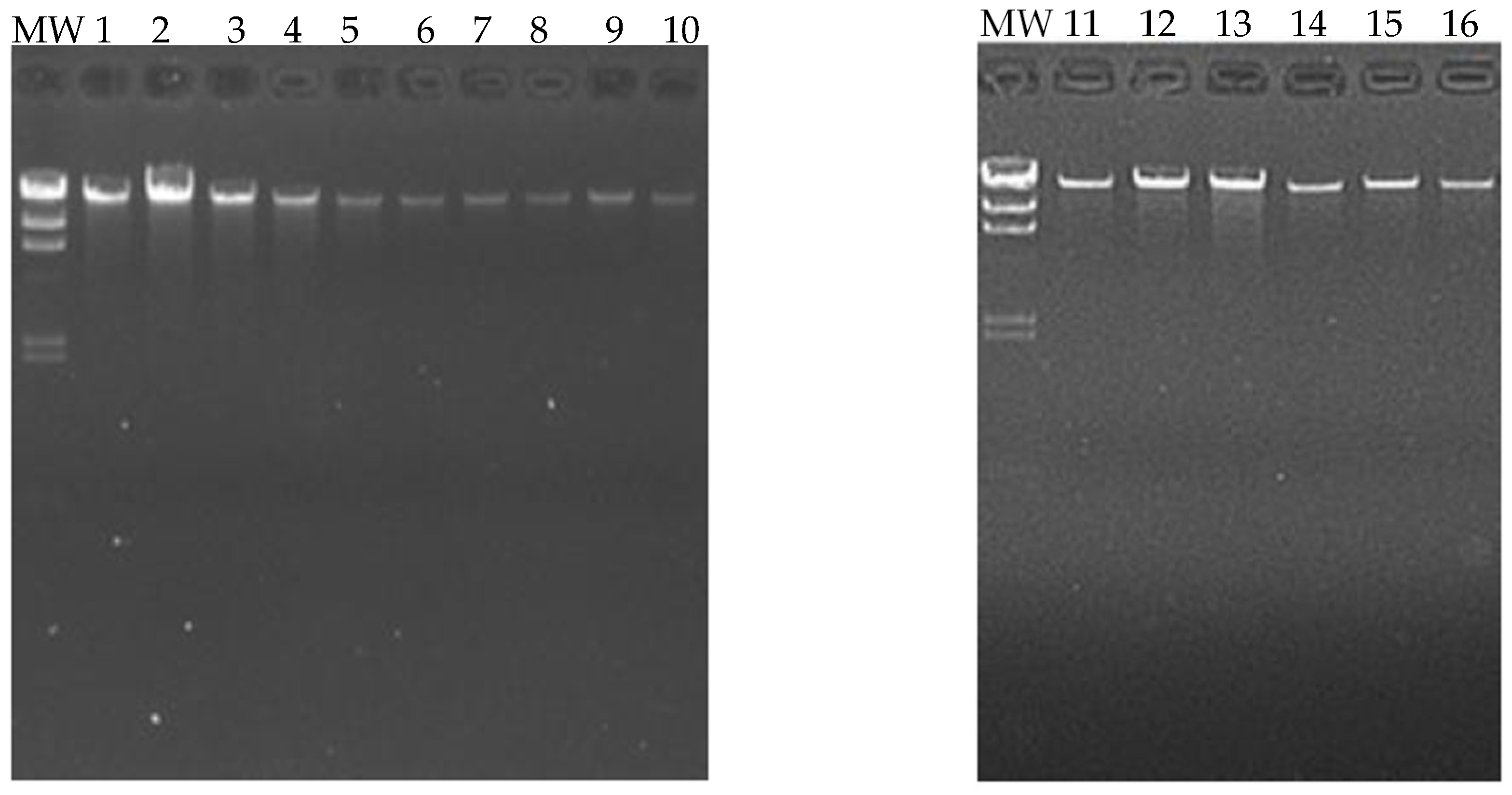
3.4. DNA Sample Extraction
3.5. Quantification and Evaluation of DNA Quality
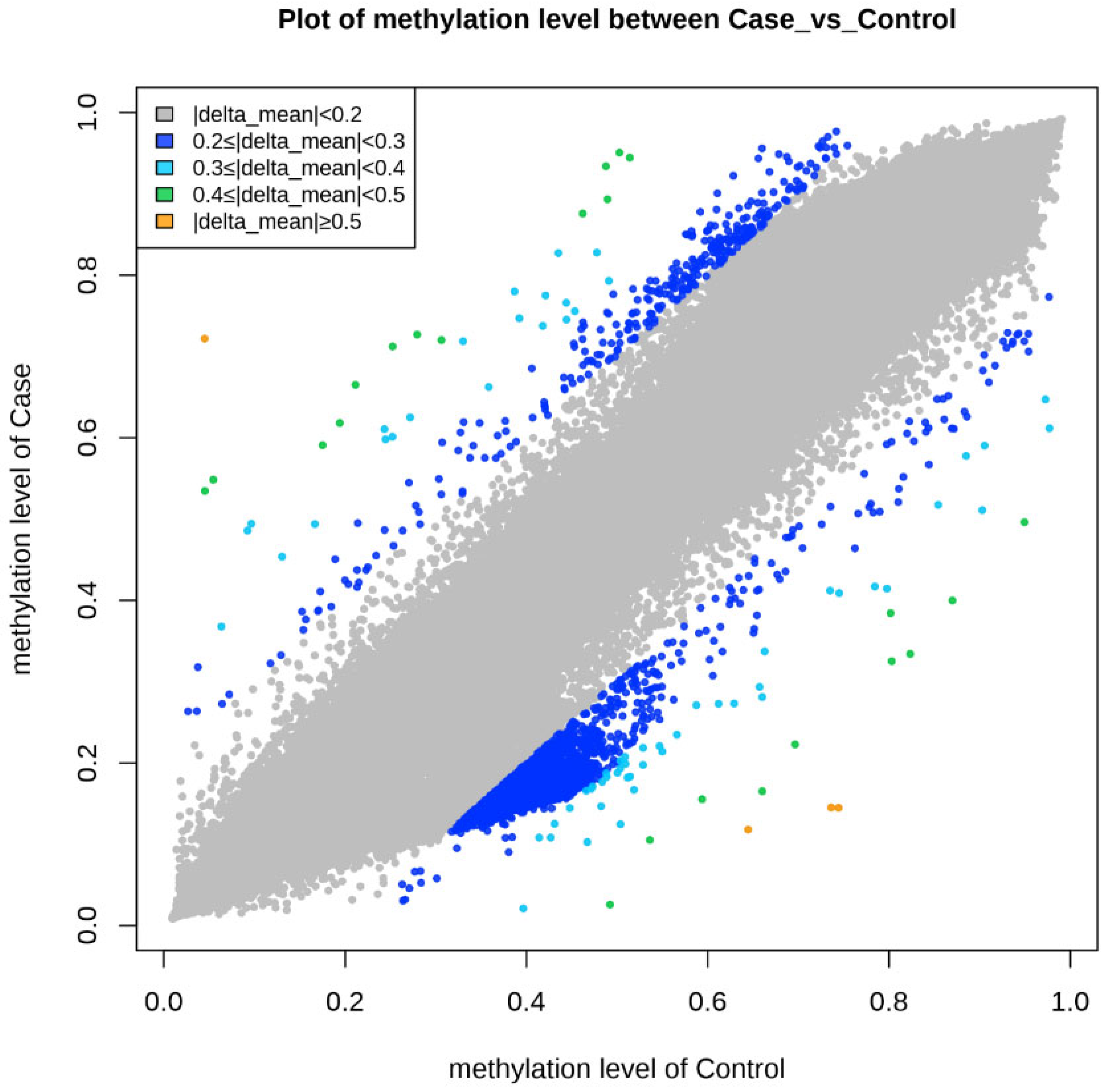
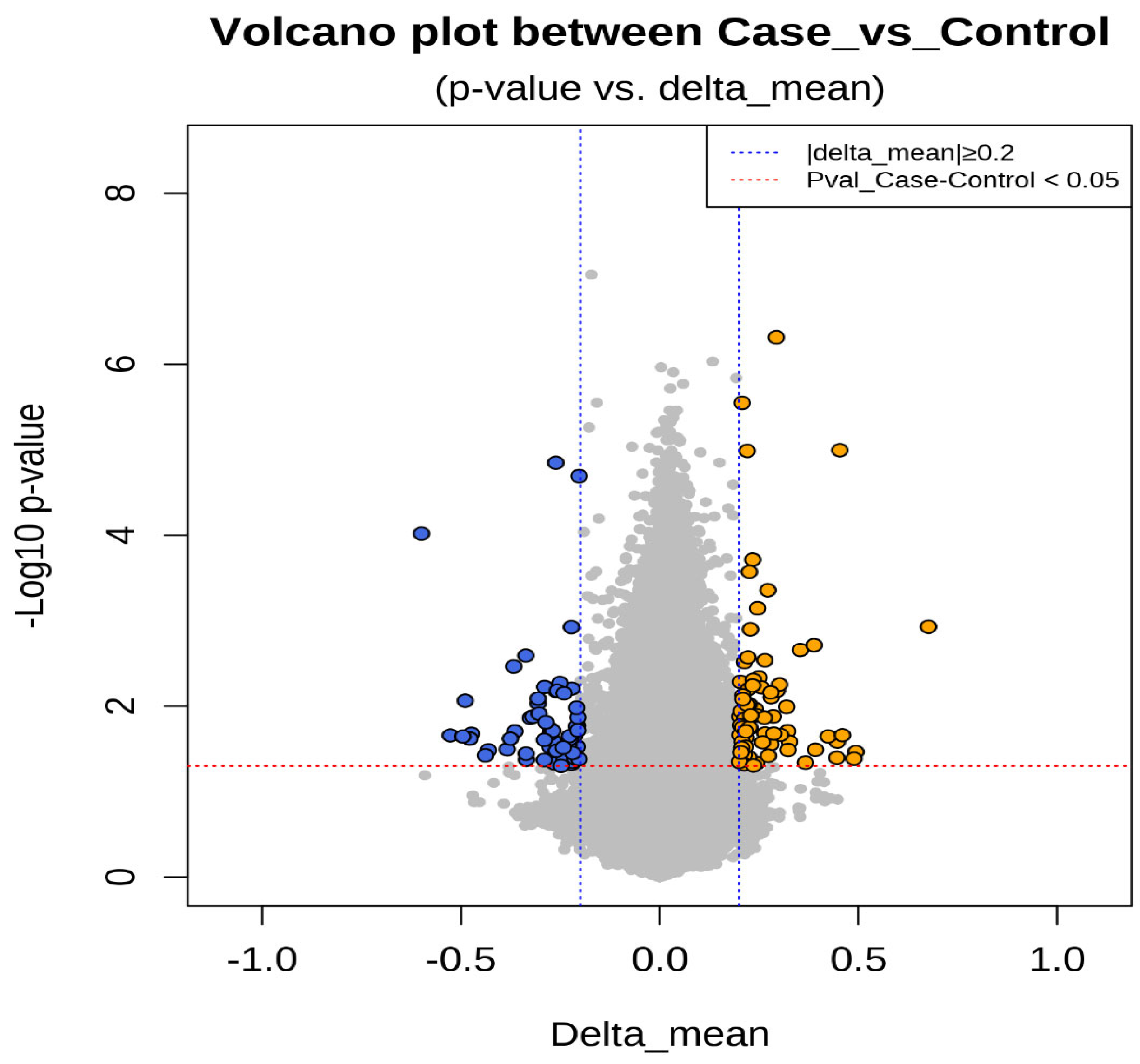
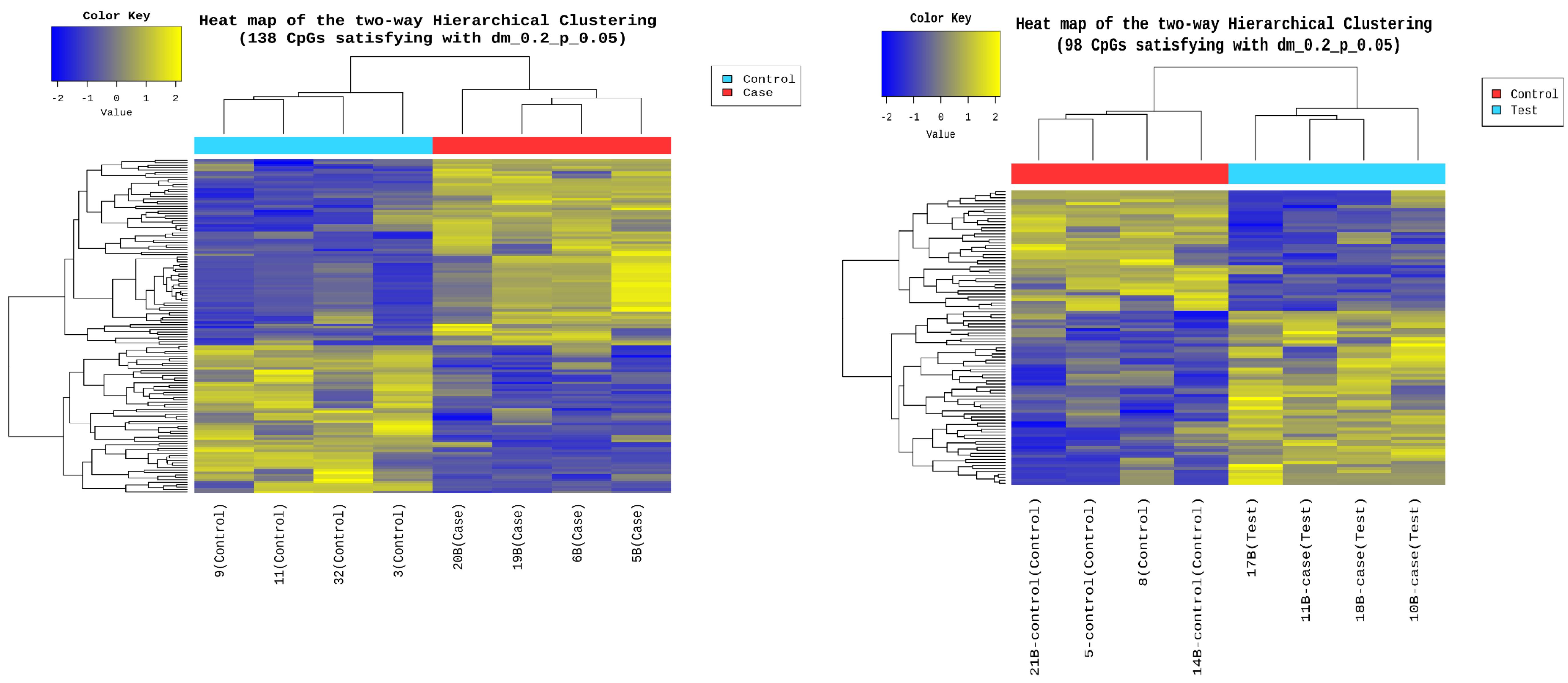
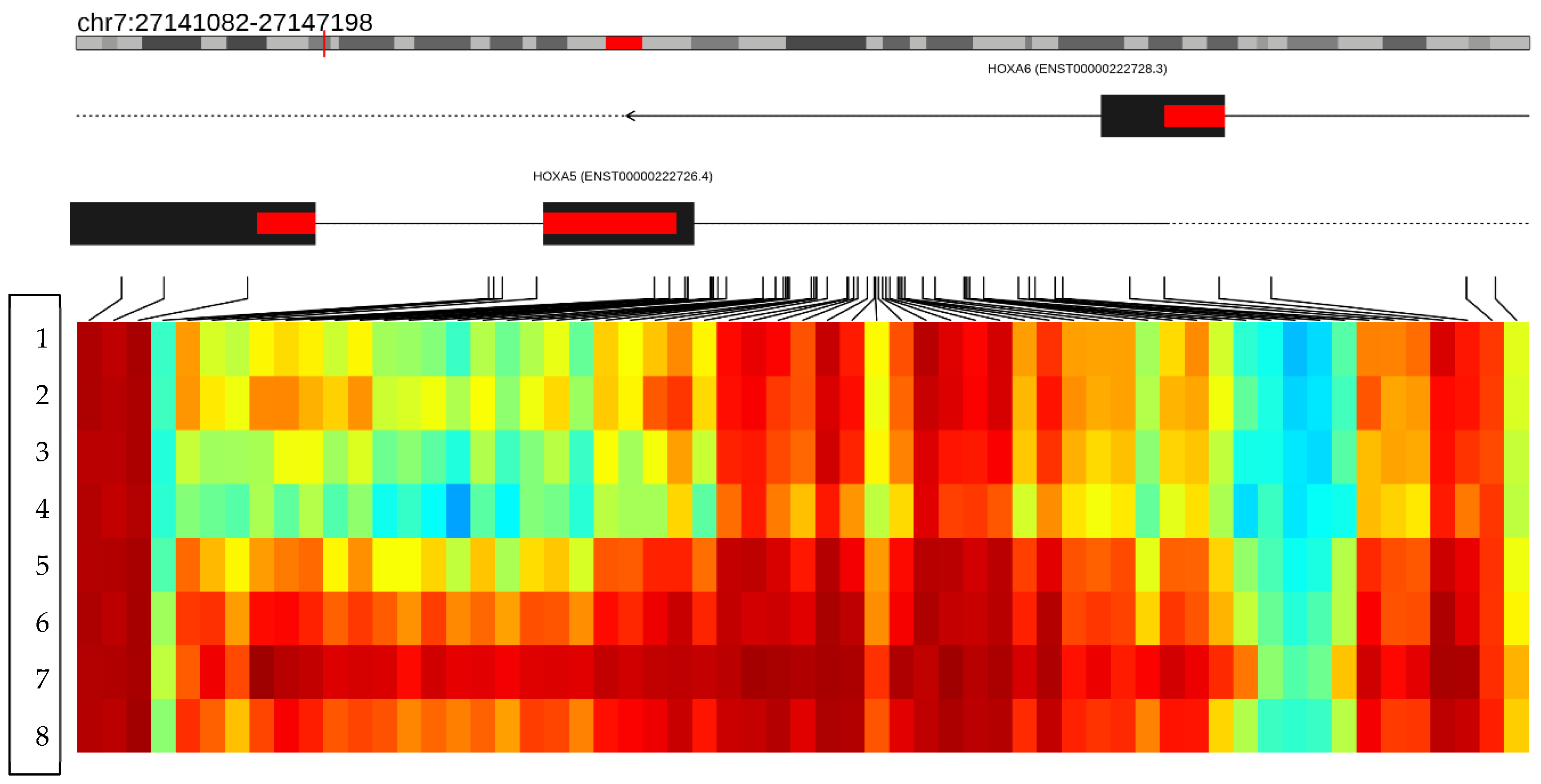
3.6. Epigenetic Sequencing of Human Methylation Profiling
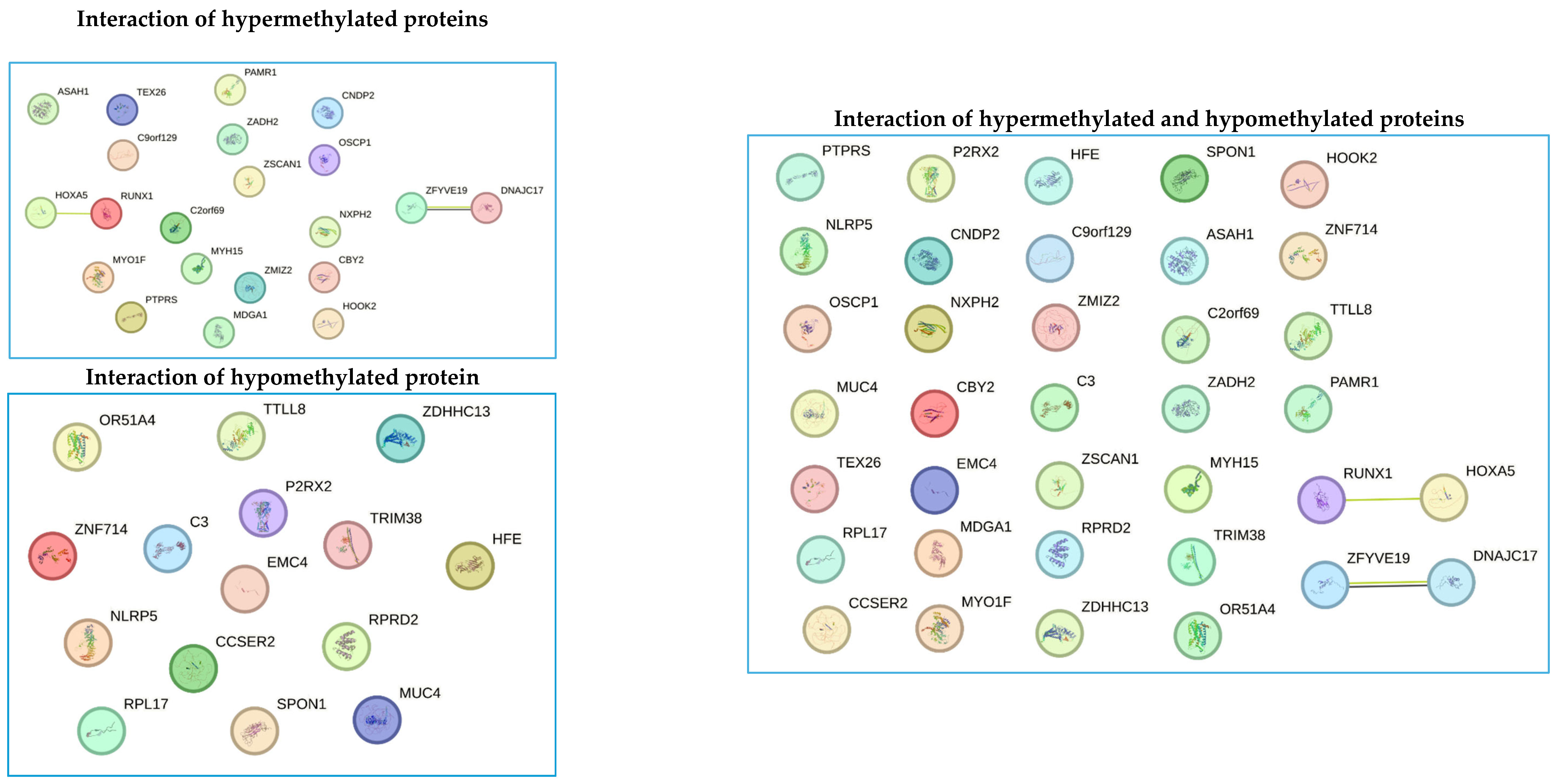
3.7. Bioinformatic Prediction of Protein Interactions
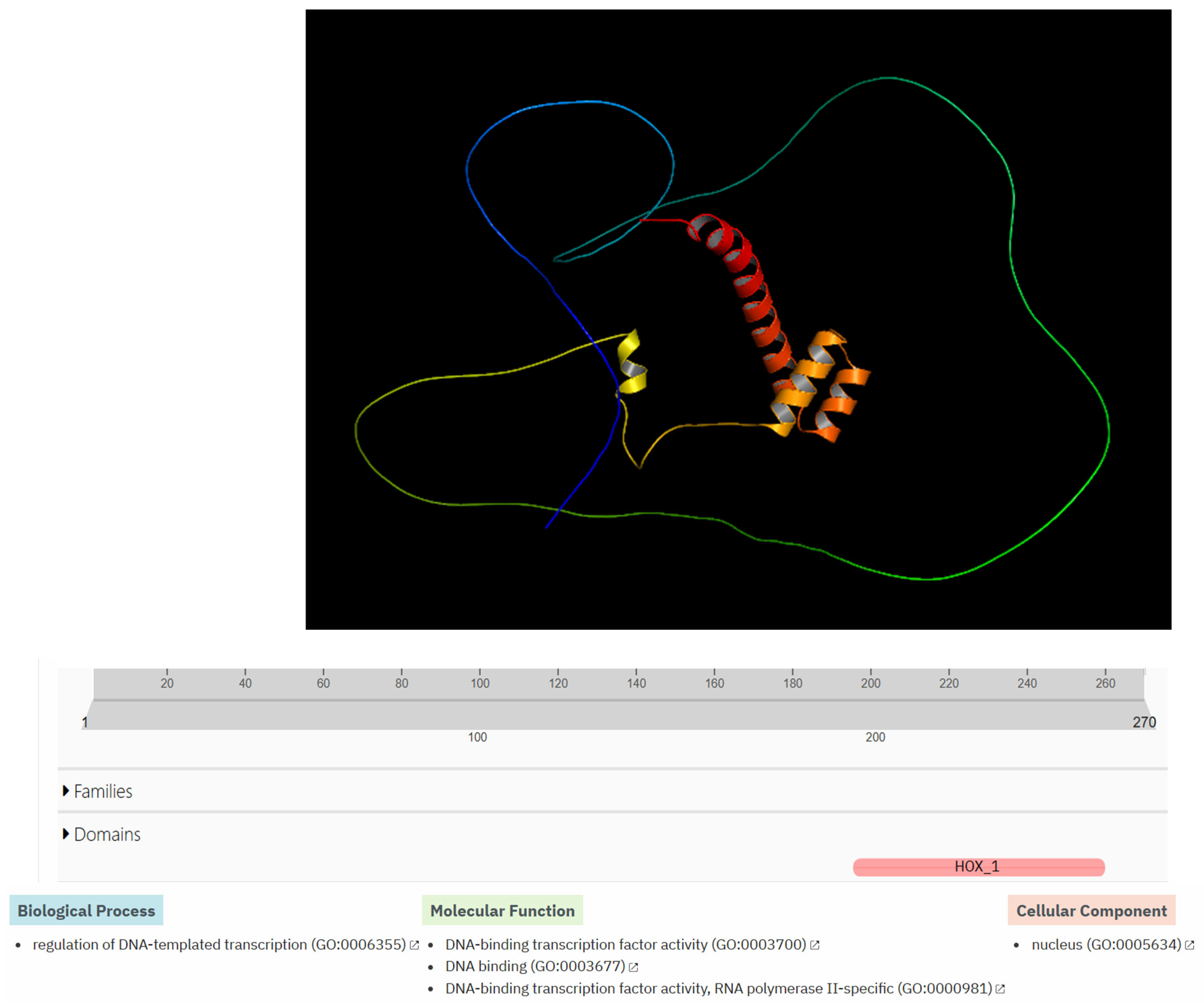
3.8. Structural Modeling and Functional Bioinformatics Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sociedad Española de EnSociedad Española de Endocrinología Pediátrica. Pequeño para la Edad Gestacional (peg): Desde el Período Prenatal Hasta la Adolescencia; Sociedad Española de EnSociedad Española de Endocrinología Pediátrica: Madrid, Spain, 2012. [Google Scholar]
- Organización Mundial de la Salud. Metas Mundiales de Nutrición 2025: Documento Normativo Sobre Bajo Peso al Nacer; World Health Organization: Geneva, Switzerland, 2014; p. 8. [Google Scholar]
- Peraza Roque, G.J.; Pérez Delgado, S.D.L.C.; Figueroa Barreto, Z.D.L.A. FigueroaBarreto. Factores asociados al bajo peso al nacer. Rev. Cuba. Med. Gen. Integral 2001, 17, 490–496. [Google Scholar]
- Hernández, M.I.; Mericq, V. Metabolic syndrome in children born small-for-gestational age. Arq. Bras. Endocrinol. Metabol. 2011, 55, 583–589. [Google Scholar] [CrossRef]
- Ramírez-Vélez, R.; Correa-Bautista, J.E.; Villa-González, E.; Martínez-Torres, J.; Hackney, A.C.; García-Hermoso, A. Effects of preterm birth and fetal growth retardation on life-course cardiovascular risk factors among schoolchildren from Colombia: The FUPRECOL study. Early Hum. Dev. 2017, 106–107, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.N.; Barker, D.J.P. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Jana, A.; Saha, U.R.; Reshmi, R.S.; Muhammad, T. Relationship between low birth weight and infant mortality: Evidence from National Family Health Survey 2019-21, India. Arch. Public Health 2023, 81, 28. [Google Scholar] [CrossRef] [PubMed]
- FAO; IFAD; PAHO; WFP; UNICEF. América Latina y el Caribe—Panorama Regional de la Seguridad Alimentaria y la Nutrición 2023; PAHO: Santiago, Chile, 2024; pp. 1–90. [Google Scholar]
- DANE. Análisis de los Nacimientos en Colombia; DANE: Bogotá, Colombia, 2023; p. 63. [Google Scholar]
- Oswiecimska, J.M.; Roczniak, W.; Roczniak, R.G.; Malczyk, Z.; Chyra, M.; Mazur, B.; Ziora, K. Serum adiponectin levels in adolescents and young adults with growth hormone deficiency. Neuro Endocrinol. Lett. 2017, 38, 107–116. [Google Scholar] [PubMed]
- Albert, S.G.; Haas, M.J.; Mooradian, A.D. The effects of recombinant human growth hormone (rhGH) supplementation on adipokines and C-reactive protein in obese subjects. Growth Horm. IGF Res. 2007, 17, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ariza Jiménez, A.B.; Martínez-Aedo Ollero, M.J.; López-Siguero, J.P. Eficacia y seguridad del tratamiento sustitutivo en el déficit aislado de hormona del crecimiento. An. Pediatría 2019, 90, 285–292. [Google Scholar] [CrossRef]
- Slattery, M.; Bredella, M.A.; Stanley, T.; Torriani, M.; Misra, M. Effects of recombinant human growth hormone (rhGH) administration on body composition and cardiovascular risk factors in obese adolescent girls. Int. J. Pediatr. Endocrinol. 2014, 2014, 22. [Google Scholar] [CrossRef] [PubMed]
- Kolarova, J.; Ammerpohl, O.; Gutwein, J.; Welzel, M.; Baus, I.; Riepe, F.G.; Eggermann, T.; Caliebe, A.; Holterhus, P.M.; Siebert, R.; et al. In vivo Investigations of the Effect of Short- and Long-Term Recombinant Growth Hormone Treatment on DNA-Methylation in Humans. PLoS ONE 2015, 10, e0120463. [Google Scholar] [CrossRef] [PubMed]
- Aurensanz Clemente, E.; Samper Villagrasa, P.; Ayerza Casas, A.; Ruiz Frontera, P.; Bueno Lozano, O.; Moreno Aznar, L.A.; Lozano, G.B. Modificaciones en variables antropométricas, analíticas de riesgo metabólico y composición corporal en pequeños para la edad gestacional en tratamiento con hormona de crecimiento. An. Pediatría 2017, 86, 240–248. [Google Scholar] [CrossRef]
- Braud, S.; Ciufolini, M.; Harosh, I. ‘Energy Expenditure Genes’ or ‘Energy Absorption Genes’: A New Target for the Treatment of Obesity and Type II Diabetes. Future Med. Chem. 2010, 2, 1777–1783. [Google Scholar] [CrossRef]
- González Jiménez, E. Genes and obesity: A cause and effect relationship. Endocrinol. Nutr. Engl. Ed. 2011, 58, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Samblas, M.; Milagro, F.I.; Martínez, A. DNA methylation markers in obesity, metabolic syndrome, and weight loss. Epigenetics 2019, 14, 421–444. [Google Scholar] [CrossRef]
- Herrera, B.M.; Keildson, S.; Lindgren, C.M. Genetics and epigenetics of obesity. Maturitas 2011, 69, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Rohde, K.; Keller, M.; la Cour Poulsen, L.; Blüher, M.; Kovacs, P.; Böttcher, Y. Genetics and epigenetics in obesity. Metabolism 2019, 92, 37–50. [Google Scholar] [CrossRef]
- Hwang, I.T. Efficacy and safety of growth hormone treatment for children born small for gestational age. Korean J. Pediatr. 2014, 57, 379. [Google Scholar] [CrossRef] [PubMed]
- Crabbé, R.; von Holtey, M.; Engrand, P.; Chatelain, P. Recombinant human growth hormone for children born small for gestational age: Meta-analysis confirms the consistent dose-effect relationship on catch-up growth. J. Endocrinol. Investig. 2008, 31, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Kühl, A.M.; Tortorella, C.C.S.; Almeida, C.C.B.; Gomes Dias, M.R.M.; Pereira, R.M. Growth hormone effect on body composition of children born small for gestational age: A systematic review. J. Pediatr. 2023, 99, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Ferruzzi, A.; Vrech, M.; Pietrobelli, A.; Cavarzere, P.; Zerman, N.; Guzzo, A.; Flodmark, C.E.; Piacentini, G.; Antoniazzi, F. The influence of growth hormone on pediatric body composition: A systematic review. Front. Endocrinol. 2023, 14, 1093691. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinformat. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef] [PubMed]
- Vourdoumpa, A.; Paltoglou, G.; Charmandari, E. The Genetic Basis of Childhood Obesity: A Systematic Review. Nutrients 2023, 15, 1416. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Kelly, T.; He, J. Genetic Epidemiology of Obesity. Epidemiol. Rev. 2007, 29, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Broken Energy Homeostasis and Obesity Pathogenesis: The Surrounding Concepts. J. Clin. Med. 2018, 7, 453. [Google Scholar] [CrossRef] [PubMed]
- Baars, J.; van den Broeck, J.; le Cessie, S.; Massa, G.; Wit, J.M. Body Mass Index in Growth Hormone Deficient Children before and during Growth Hormone Treatment. Horm. Res. 1998, 49, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Cho, S.Y.; Kwak, M.J.; Kim, S.J.; Park, S.W.; Jin, D.K.; Lee, J.E. Impact of BMI on peak growth hormone responses to provocative tests and therapeutic outcome in children with growth hormone deficiency. Sci. Rep. 2019, 9, 16181. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, A.; Bollepalli, S.; Ollikainen, M. The potential of DNA methylation as a biomarker for obesity and smoking. J. Intern. Med. 2022, 292, 390–408. [Google Scholar] [CrossRef]
- Chen, M.; Gan, D.; Luo, Y.; Rampersad, S.; Xu, L.; Yang, S.; Li, N.; Li, H. Effect of recombinant human growth hormone therapy on blood lipid and carotid intima-media thickness in children with growth hormone deficiency. Pediatr. Res. 2018, 83, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Kubo, T.; Furujo, M.; Takahashi, K.; Hyodo, Y.; Tsuchiya, H.; Hattori, M.; Fujinaga, S.; Urayama, K. Effects of Growth Hormone Treatment on Lipid Profiles. Indian J. Pediatr. 2018, 85, 261–265. [Google Scholar] [CrossRef]
- Mahmoud, A.M. An Overview of Epigenetics in Obesity: The Role of Lifestyle and Therapeutic Interventions. Int. J. Mol. Sci. 2022, 23, 1341. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Escribano, C.; Buonaiuto, V.; Ruiz-Moreno, M.I.; Vargas-Candela, A.; Vilches-Perez, A.; Benitez-Porres, J.; Romance-Garcia, A.R.; Ruiz-Moreno, A.; Gomez-Huelgas, R.; Bernal-Lopez, M.R. Epigenetic approach in obesity: DNA methylation in a prepubertal population which underwent a lifestyle modification. Clin. Epigenetics 2020, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Lastialno, M.; Bashari, M.; Ariyanto, E. Current Updates on the Understanding of the Role of DNA Methylation on Obesity. Diabetes Metab. Syndr. Obes. 2024, 17, 3177–3186. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, U.S.; Adiga, D.; Kabekkodu, S.P.; Hunter, K.D.; Radhakrishnan, R. Molecular implications of HOX genes targeting multiple signaling pathways in cancer. Cell Biol. Toxicol. 2022, 38, 1–30. [Google Scholar] [CrossRef]
- Bürglin, T.R.; Affolter, M. Homeodomain proteins: An update. Chromosoma 2016, 125, 497–521. [Google Scholar] [CrossRef]
- Carless, M.A.; Kulkarni, H.; Kos, M.Z.; Charlesworth, J.; Peralta, J.M.; Göring, H.H.H.; Curran, J.E.; Almasy, L.; Dyer, T.D.; Comuzzie, A.G.; et al. Genetic Effects on DNA Methylation and Its Potential Relevance for Obesity in Mexican Americans. Perry, J.R.B.; editor. PLoS ONE 2013, 8, e73950. [Google Scholar] [CrossRef] [PubMed]
- Gesta, S.; Blüher, M.; Yamamoto, Y.; Norris, A.W.; Berndt, J.; Kralisch, S.; Boucher, J.; Lewis, C.; Kahn, C.R. Evidence for a role of developmental genes in the origin of obesity and body fat distribution. Proc. Natl. Acad. Sci. USA 2006, 103, 6676–6681. [Google Scholar] [CrossRef] [PubMed]
- Parrillo, L.; Spinelli, R.; Longo, M.; Zatterale, F.; Santamaria, G.; Leone, A.; Campitelli, M.; Raciti, G.A.; Beguinot, F. The Transcription Factor HOXA5: Novel Insights into Metabolic Diseases and Adipose Tissue Dysfunction. Cells 2023, 12, 2090. [Google Scholar] [CrossRef] [PubMed]
- Rönn, T.; Perfilyev, A.; Jönsson, J.; Eriksson, K.F.; Jørgensen, S.W.; Brøns, C.; Gillberg, L.; Vaag, A.; Stener-Victorin, E.; Ling, C. Circulating triglycerides are associated with human adipose tissue DNA methylation of genes linked to metabolic disease. Hum. Mol. Genet. 2023, 32, 1875–1887. [Google Scholar] [CrossRef] [PubMed]
- Parrillo, L.; Spinelli, R.; Costanzo, M.; Florese, P.; Cabaro, S.; Desiderio, A.; Prevenzano, I.; Raciti, G.A.; Smith, U.; Miele, C.; et al. Epigenetic Dysregulation of the Homeobox A5 (HOXA5) Gene Associates with Subcutaneous Adipocyte Hypertrophy in Human Obesity. Cells 2022, 11, 728. [Google Scholar] [CrossRef]
- Parrillo, L.; Spinelli, R.; Nicolò, A.; Longo, M.; Mirra, P.; Raciti, G.A.; Miele, C.; Beguinot, F. Nutritional Factors, DNA Methylation, and Risk of Type 2 Diabetes and Obesity: Perspectives and Challenges. Int. J. Mol. Sci. 2019, 20, 2983. [Google Scholar] [CrossRef] [PubMed]
- Phizicky, E.M.; Fields, S. Protein-protein interactions: Methods for detection and analysis. Microbiol. Rev. 1995, 59, 94–123. [Google Scholar] [CrossRef]
- Rao, V.S.; Srinivas, K.; Sujini, G.N.; Kumar, G.N.S. Protein-Protein Interaction Detection: Methods and Analysis. Int. J. Proteomics 2014, 2014, 147648. [Google Scholar] [CrossRef] [PubMed]
- Skrabanek, L.; Saini, H.K.; Bader, G.D.; Enright, A.J. Computational Prediction of Protein–Protein Interactions. Mol. Biotechnol. 2008, 38, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Janin, J.; Bonvin, A.M. Protein–protein interactions. Curr. Opin. Struct. Biol. 2013, 23, 859–861. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Li, P.; Ma, Y.; Jiang, C.F.; Chen, Y.; Shi, Y.; Gupta, N.; Seifuddin, F.; Pirooznia, M.; Ohnishi, Y.; et al. Identification of human long non-coding RNAs associated with nonalcoholic fatty liver disease and metabolic homeostasis. J. Clin. Investig. 2021, 131, e136336. [Google Scholar] [CrossRef]
- Hennessy, E.J.; Van Solingen, C.; Scacalossi, K.R.; Ouimet, M.; Afonso, M.S.; Prins, J.; Koelwyn, G.J.; Sharma, M.; Ramkhelawon, B.; Carpenter, S.; et al. The long non-coding RNA CHROME regulates cholesterol homeostasis in primates. Nat. Metab. 2018, 1, 98–110. [Google Scholar] [CrossRef]
- Du, Q.; Hoover, A.R.; Dozmorov, I.; Raj, P.; Khan, S.; Molina, E.; Chang, T.C.; De La Morena, M.T.; Cleaver, O.B.; Mendell, J.T.; et al. MIR205HG Is a Long Non-coding RNA that Regulates Growth Hormone and Prolactin Production in the Anterior Pituitary. Dev. Cell 2019, 49, 618–631.e5. [Google Scholar] [CrossRef]
- Zhang, P.; Cao, L.; Fan, P.; Mei, Y.; Wu, M. Lnc RNA-MIF, a c-Myc-activated long non-coding RNA, suppresses glycolysis by promoting Fbxw7-mediated c-Myc degradation. EMBO Rep. 2016, 17, 1204–1220. [Google Scholar] [CrossRef]
- Zheng, X.; Han, H.; Liu, G.; Ma, Y.; Pan, R.; Sang, L.; Li, R.; Yang, L.; Marks, J.; Wang, W.; et al. Lnc RNA wires up Hippo and Hedgehog signaling to reprogramme glucose metabolism. EMBO J. 2017, 36, 3325–3335. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Category | Growth Hormone | p | |||
|---|---|---|---|---|---|---|
| Yes | No | |||||
| n | % | n | % | |||
| Sex | Female | 4 | 28.6 | 10 | 71.4 | |
| Male | 10 | 71.4 | 4 | 28.6 | ||
| SMGD | 1 | 7.70 | 1 | 7.70 | 0.95 | |
| Tobacco exposure | 1 | 7.10 | 0 | 0.00 | 0.30 | |
| Toxemia | 1 | 7.10 | 2 | 14.3 | 0.54 | |
| HBP | 1 | 7.10 | 1 | 7.10 | 1.00 | |
| GH | 2 | 14.3 | 1 | 7.10 | 0.54 | |
| TPB | 7 | 50.0 | 2 | 14.3 | 0.04 | |
| TA | 3 | 21.4 | 1 | 7.10 | 0.28 | |
| Mean | SD | Mean | SD | p | ||
| Birth weight | 1921.14 | 536.3 | 1949.21 | 483.21 | 0.88 | |
| MAD | 28 | 7.33 | 27.71 | 4.76 | 0.90 | |
| Pregnancies | 2.15 | 1.23 | 1.64 | 0.74 | 0.20 | |
| Gestational age | 33.5 | 3.79 | 34.57 | 2.31 | 0.37 | |
| Prenatal history | 6.57 | 4.01 | 7.78 | 1.05 | 0.29 | |
| Variable | Growth Hormone | p | |||
|---|---|---|---|---|---|
| Yes | No | ||||
| Mean | SD | Mean | SD | ||
| Waist circumference (cm) | 68.76 | 9.01 | 73.66 | 19.21 | 0.39 |
| Total cholesterol (mg/dL) | 156.92 | 17.3 | 155.42 | 36.55 | 0.89 |
| HDL cholesterol (mg/dL) | 53.66 | 13.1 | 46.43 | 9.12 | 0.1 |
| LDL cholesterol (mg/dL) | 94.92 | 16.55 | 97.02 | 33.93 | 0.83 |
| Triglycerides (mg/dL) | 86.14 | 36.44 | 139.57 | 95.16 | 0.06 |
| Glucose (mg/dL) | 91.42 | 7.42 | 89.92 | 9.23 | 0.64 |
| # | Sample Name | Purity (260/280) | Purity (260/230) | Concentration (ng/μL) | Volume (μL) | Total Amount (μg) |
|---|---|---|---|---|---|---|
| 1 | 11 (Control) | 1.85 | 1.81 | 82.23 | 25 | 2.06 |
| 2 | 32 (Control) | 1.87 | 1.99 | 104.33 | 14 | 1.46 |
| 3 | 9 (Control) | 1.91 | 1.85 | 81 | 30 | 2.43 |
| 4 | 3 (Control) | 1.87 | 1.97 | 42.1 | 20 | 0.84 |
| 5 | 8 (Control) | 1.91 | 1.85 | 22.59 | 30 | 0.68 |
| 6 | 14B (Control) | 2.02 | 2.1 | 28.27 | 30 | 0.85 |
| 7 | 21B (Control) | 1.98 | 2.14 | 44.77 | 30 | 1.34 |
| 8 | 5 (Control) | 1.94 | 1.86 | 46.78 | 30 | 1.4 |
| 9 | 20B (Case) | 1.81 | 1.95 | 13.86 | 175 | 2.42 |
| 10 | 6B (Case) | 1.84 | 1.85 | 14.91 | 180 | 2.68 |
| 11 | 5B (Case) | 1.91 | 1.91 | 13.12 | 180 | 2.36 |
| 12 | 19B (Case) | 1.82 | 1.81 | 18.57 | 180 | 3.34 |
| 13 | 17B (Case) | 1.85 | 1.86 | 13.31 | 185 | 2.46 |
| 14 | 18B (Case) | 2.08 | 2.04 | 23.09 | 30 | 0.69 |
| 15 | 10B (Case) | 2.05 | 2.02 | 21.75 | 30 | 0.65 |
| 16 | 11B (Case) | 2.01 | 1.86 | 19.21 | 30 | 0.58 |
| Gene Name | Gene Accession | Control Mean | Case Mean | Delta Mean | p-Value | Fold Change | Odds Ratio | Methylation Status |
|---|---|---|---|---|---|---|---|---|
| MDGA1 | NM_153487.4 | 0.4352 | 0.8272 | 0.3920 | 0.0325 | 2.2004 | 7.6790 | Hypermethylated |
| HOXA5 | NM_019102.4 | 0.4181 | 0.7375 | 0.3195 | 0.0102 | 1.7870 | 4.4147 | Hypermethylated |
| LINC01168 | NR_046231.1 | 0.4603 | 0.7336 | 0.2733 | 0.0383 | 1.5631 | 3.7230 | Hypermethylated |
| ZFYVE19 | NM_032850.5 | 0.5352 | 0.7693 | 0.2340 | 0.0002 | 1.4404 | 2.9114 | Hypermethylated |
| ASAH1 | NR_125429.1 | 0.4673 | 0.6945 | 0.2271 | 0.0182 | 1.4777 | 2.7093 | Hypermethylated |
| MYH15 | NM_014981.3 | 0.5901 | 0.8163 | 0.2261 | 0.0003 | 1.3881 | 3.0815 | Hypermethylated |
| DNAJC17 | NM_018163.3 | 0.2342 | 0.4550 | 0.2208 | 0.0000 | 1.9411 | 2.7305 | Hypermethylated |
| PAMR1 | NM_015430.4 | 0.5031 | 0.7166 | 0.2135 | 0.0373 | 1.4697 | 2.4634 | Hypermethylated |
| MROCKI | NR_038863.2 | 0.5322 | 0.7417 | 0.2095 | 0.0083 | 1.4047 | 2.5660 | Hypermethylated |
| CNDP2 | NM_001168499.2 | 0.4905 | 0.6950 | 0.2044 | 0.0349 | 1.4147 | 2.5076 | Hypermethylated |
| CBY2 | NM_001286342.2 | 0.2132 | 0.4163 | 0.2031 | 0.0052 | 2.2042 | 2.9562 | Hypermethylated |
| ZADH2 | NM_001306093.1 | 0.2021 | 0.4381 | 0.2360 | 0.023839 | 2.1549 | 3.1290 | Hypermethylated |
| HOOK2 | NM_001100176.2 | 0.1994 | 0.6461 | 0.4467 | 0.021309 | 4.5404 | 12.282 | Hypermethylated |
| C9orf129 | NR_166069.1 | 0.0545 | 0.2682 | 0.2137 | 0.042371 | 3.6805 | 4.8420 | Hypermethylated |
| NXPH2 | NM_007226.3 | 0.3943 | 0.6575 | 0.2632 | 0.025904 | 1.7214 | 3.1292 | Hypermethylated |
| OSCP1 | NM_001330493.2 | 0.2899 | 0.5103 | 0.2203 | 0.003039 | 1.8052 | 2.6168 | Hypermethylated |
| ZMIZ2 | NM_001300959.2 | 0.4532 | 0.7045 | 0.2513 | 0.027977 | 1.5689 | 3.1253 | Hypermethylated |
| RUNX1 | NM_001001890.3 | 0.2143 | 0.4373 | 0.2229 | 0.043655 | 2.3259 | 3.2752 | Hypermethylated |
| PTPRS | NM_001394011.1 | 0.5526 | 0.7648 | 0.2121 | 0.0000745 | 1.3850 | 2.6442 | Hypermethylated |
| TEX26 | NM_001353390.2 | 0.4745 | 0.7073 | 0.2327 | 0.026512 | 1.5207 | 2.7487 | Hypermethylated |
| EIF2A4K | NM_001013703.4 | 0.5625 | 0.7739 | 0.2114 | 0.044890 | 1.3523 | 3.1282 | Hypermethylated |
| MYO1F | NM_001348355.2 | 0.6045 | 0.8542 | 0.2497 | 0.013192 | 1.4221 | 5.1808 | Hypermethylated |
| C2orf69 | NM_153689.6 | 0.4277 | 0.6855 | 0.2578 | 0.041214 | 1.6343 | 3.3052 | Hypermethylated |
| ZSCAN1 | NM_182572.4 | 0.2962 | 0.5344 | 0.2382 | 0.033339 | 1.9793 | 2.9281 | Hypermethylated |
| C10orf71 | NR_108038.1 | 0.5094 | 0.3065 | −0.2029 | 0.0418 | −1.6093 | 0.4233 | Hypomethylated |
| ZDHHC13 | NM_001001483.3 | 0.6519 | 0.4462 | −0.2057 | 0.0136 | −1.4886 | 0.4246 | Hypomethylated |
| RPL17 | NM_001199342.3 | 0.5739 | 0.3681 | −0.2058 | 0.0192 | −1.6226 | 0.4230 | Hypomethylated |
| EMC4 | NM_001286420.2 | 0.7009 | 0.4912 | −0.2097 | 0.0387 | −1.4272 | 0.3798 | Hypomethylated |
| RPRD2 | NM_001387114.1 | 0.9296 | 0.7105 | −0.2190 | 0.0354 | −1.3311 | 0.2035 | Hypomethylated |
| OBSCN | NM_001098623.2 | 0.5200 | 0.3004 | −0.2195 | 0.0447 | −1.8213 | 0.3751 | Hypomethylated |
| ZNF714 | NM_182515.4 | 0.5595 | 0.3199 | −0.2395 | 0.0459 | −1.7464 | 0.3589 | Hypomethylated |
| MUC4 | NM_004532.6_8 | 0.7046 | 0.4643 | −0.2403 | 0.0422 | −1.5140 | 0.3101 | Hypomethylated |
| SUGT1P4 | NR_036526.1 | 0.8832 | 0.6321 | −0.2510 | 0.0054 | −1.4048 | 0.2032 | Hypomethylated |
| TRIM38 | NM_006355.5 | 0.7799 | 0.5188 | −0.2611 | 0.0000 | −1.5043 | 0.3026 | Hypomethylated |
| C3 | NM_000064.4 | 0.4268 | 0.1082 | −0.3186 | 0.0133 | −3.5811 | 0.1735 | Hypomethylated |
| SPON1 | NM_006108.4 | 0.4823 | 0.1468 | −0.3355 | 0.0424 | −2.8404 | 0.1901 | Hypomethylated |
| NGF-AS1 | NR_157569.1 | 0.5362 | 0.1052 | −0.4309 | 0.0330 | −4.1430 | 0.1059 | Hypomethylated |
| CCSER2 | NM_001284243.2 | 0.6073 | 0.2451 | −0.3622 | 0.0145040 | −2.51360 | 0.1880 | Hypomethylated |
| P2RX2 | NM_174872.3 | 0.7155 | 0.5131 | −0.2023 | 0.0307259 | −1.37424 | 0.3882 | Hypomethylated |
| LOC284379 | NR_002938.3 | 0.5250 | 0.285 | −0.2393 | 0.0289216 | −1.77223 | 0.3563 | Hypomethylated |
| GGTA1 | NM_001382584.1 | 0.9308 | 0.6773 | −0.2535 | 0.0320150 | −1.40730 | 0.2004 | Hypomethylated |
| NLRP5 | NM_153447.4 | 0.7447 | 0.5068 | −0.2378 | 0.0109644 | −1.49354 | 0.3446 | Hypomethylated |
| OR51A4 | NM_001005329.2 | 0.4572 | 0.1365 | −0.3206 | 0.0158866 | −6.11715 | 0.1048 | Hypomethylated |
| HLA-H | NR_001434.4_8 | 0.5794 | 0.3483 | −0.2310 | 0.0098659 | −1.65606 | 0.3809 | Hypomethylated |
| TTLL8 | NM_001350317.2 | 0.9795 | 0.7516 | −0.2278 | 0.0008761 | −1.308043 | 0.0649 | Hypomethylated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velásquez, J.M.A.; Vásquez Trespalacios, E.M.; Urrego, R.; Arroyave Toro, M.C.; Montilla Velásquez, M.d.P.; Soto, C.M.D.; Vélez, J.C.Z.; Jaramillo Henríquez, V.; Flórez, J.E.S.; Monroy, F.P.; et al. Effect of Recombinant Human Growth Hormone (rhGH) Use on Genetic Methylation Patterns and Their Relationship with Body Composition in Small-for-Gestational-Age (SGA) Newborns. Biomedicines 2025, 13, 1288. https://doi.org/10.3390/biomedicines13061288
Velásquez JMA, Vásquez Trespalacios EM, Urrego R, Arroyave Toro MC, Montilla Velásquez MdP, Soto CMD, Vélez JCZ, Jaramillo Henríquez V, Flórez JES, Monroy FP, et al. Effect of Recombinant Human Growth Hormone (rhGH) Use on Genetic Methylation Patterns and Their Relationship with Body Composition in Small-for-Gestational-Age (SGA) Newborns. Biomedicines. 2025; 13(6):1288. https://doi.org/10.3390/biomedicines13061288
Chicago/Turabian StyleVelásquez, Juan M. Alfaro, Elsa Maria Vásquez Trespalacios, Rodrigo Urrego, María C. Arroyave Toro, María del Pilar Montilla Velásquez, Cecilia Maria Díaz Soto, Juan C. Zuluaga Vélez, Verónica Jaramillo Henríquez, Jorge Emilio Salazar Flórez, Fernando P. Monroy, and et al. 2025. "Effect of Recombinant Human Growth Hormone (rhGH) Use on Genetic Methylation Patterns and Their Relationship with Body Composition in Small-for-Gestational-Age (SGA) Newborns" Biomedicines 13, no. 6: 1288. https://doi.org/10.3390/biomedicines13061288
APA StyleVelásquez, J. M. A., Vásquez Trespalacios, E. M., Urrego, R., Arroyave Toro, M. C., Montilla Velásquez, M. d. P., Soto, C. M. D., Vélez, J. C. Z., Jaramillo Henríquez, V., Flórez, J. E. S., Monroy, F. P., Palacio Mosquera, H. A., Vélez Gómez, S., & Pelaez Sánchez, R. G. (2025). Effect of Recombinant Human Growth Hormone (rhGH) Use on Genetic Methylation Patterns and Their Relationship with Body Composition in Small-for-Gestational-Age (SGA) Newborns. Biomedicines, 13(6), 1288. https://doi.org/10.3390/biomedicines13061288