
Proteome-Wide Analysis of Autoantibodies in Open-Angle Glaucoma in Japanese Population: A Pilot Study

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
- (1)
- Three or more points with p < 5%, except for the most peripheral inspection points in the pattern deviation probability plot that are adjacent to each other with p < 1%.
- (2)
- Pattern standard deviation with a probability of less than 5%.
- (3)
- Glaucoma hemifield test indicating that the field is outside normal limits.
2.2. Wet Proteome Analysis
2.2.1. Protein Expression In Vitro
2.2.2. Step 1. Measurement of Plasma Autoantibodies by Comprehensive Wet Protein Array (CWPA) in Initial Screening
2.2.3. Step 2. Plasma Autoantibody Assay with Custom-Designed WPA
2.2.4. Protein Array Data Analysis
2.3. Statistical Analysis
3. Results
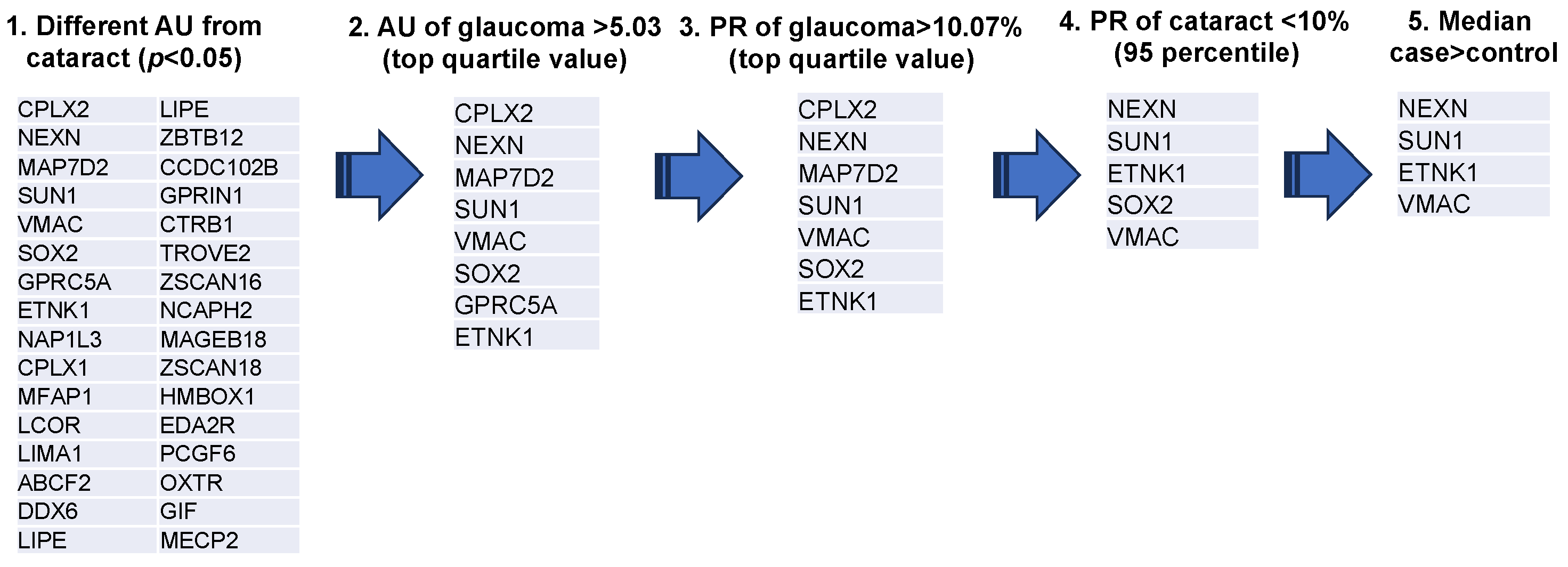
3.1. Extraction of Autoantibodies
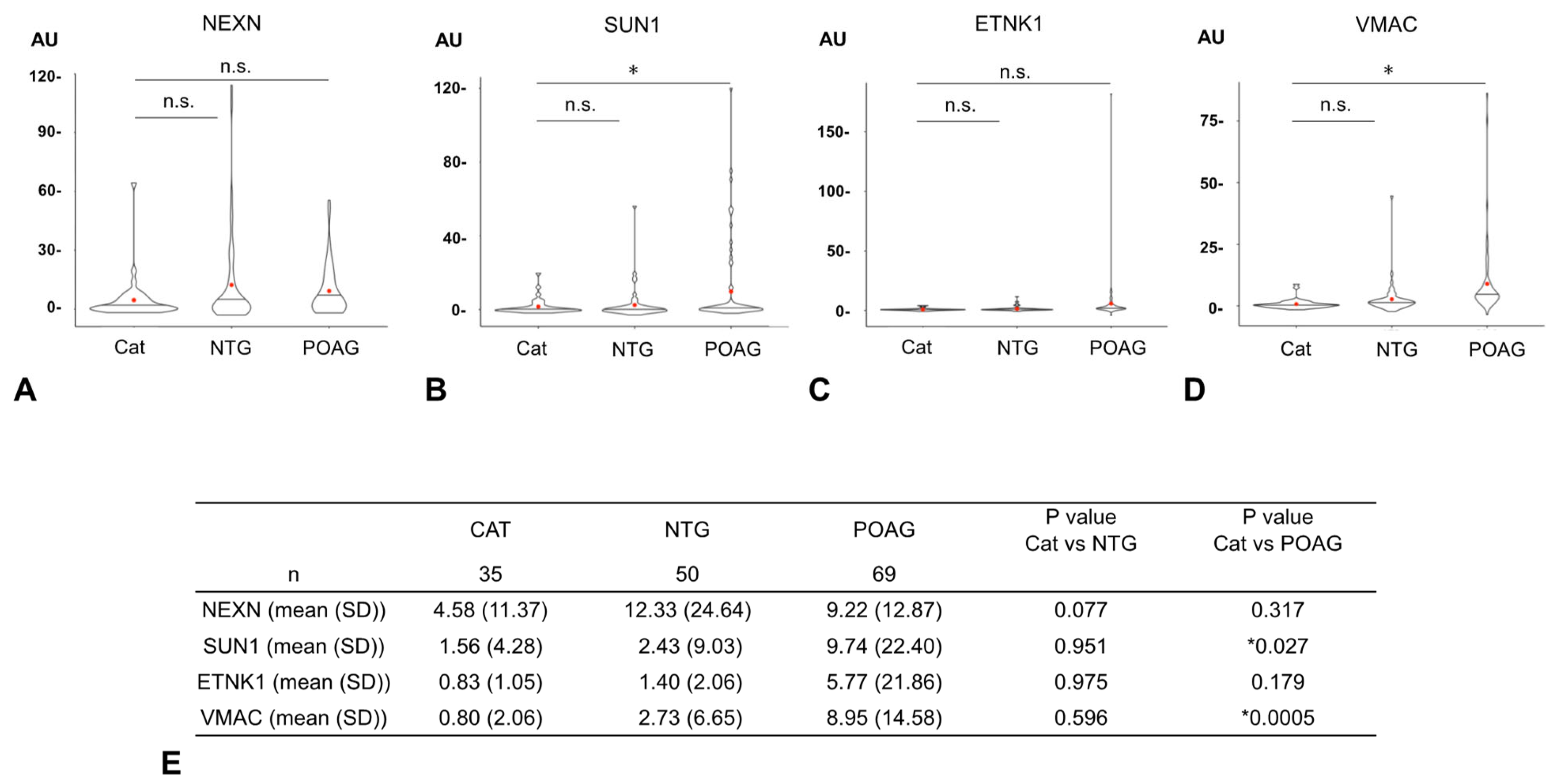
3.2. Comparison of Autoantibody Titers
3.3. Examination of the Ability of Each Autoantibody to Discriminate Between the Cataract and Glaucoma Groups
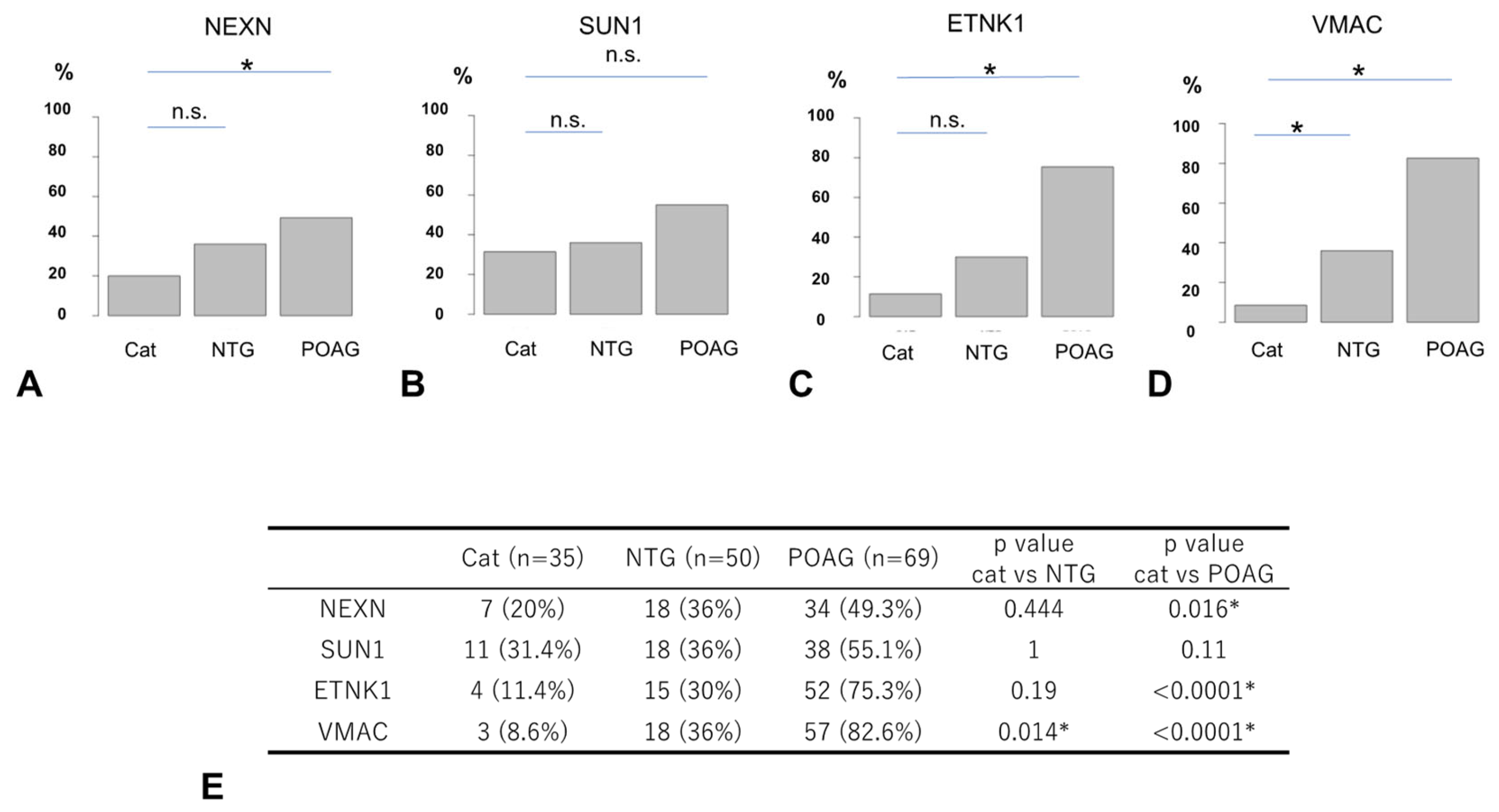
3.4. Comparison of Autoantibody Positivity Rates
3.5. Overlap of Autoantibodies
3.6. Relationship Between Autoantibodies and Clinical Findings
3.7. Logistic Regression Analysis to Predict Glaucoma Using a Set of Glaucoma-Associated Autoantibodies
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stein, J.D.; Khawaja, A.P.; Weizer, J.S. Glaucoma in Adults-Screening, Diagnosis, and Management: A Review. JAMA 2021, 325, 164–174. [Google Scholar] [CrossRef]
- Matoba, R.; Morimoto, N.; Kawasaki, R.; Fujiwara, M.; Kanenaga, K.; Yamashita, H.; Sakamoto, T.; Morizane, Y. A nationwide survey of newly certified visually impaired individuals in Japan for the fiscal year 2019: Impact of the revision of criteria for visual impairment certification. Jpn. J. Ophthalmol. 2023, 67, 346–352. [Google Scholar] [CrossRef]
- Iwase, A.; Suzuki, Y.; Araie, M.; Yamamoto, T.; Abe, H.; Shirato, S.; Kuwayama, Y.; Mishima, H.K.; Shimizu, H.; Tomita, G.; et al. The prevalence of primary open-angle glaucoma in Japanese: The Tajimi Study. Ophthalmology 2004, 111, 1641–1648. [Google Scholar] [CrossRef]
- Zhao, J.; Solano, M.M.; Oldenburg, C.E.; Liu, T.; Wang, Y.; Wang, N.; Lin, S.C. Prevalence of Normal-Tension Glaucoma in the Chinese Population: A Systematic Review and Meta-Analysis. Am. J. Ophthalmol. 2019, 199, 101–110. [Google Scholar] [CrossRef]
- Collaborative Normal-Tension Glaucoma Study Group. The effectiveness of intraocular pressure reduction in the treatment of normal-tension glaucoma. Am. J. Ophthalmol. 1998, 126, 498–505. [Google Scholar] [CrossRef]
- Kerrigan-Baumrind, L.A.; Quigley, H.A.; Pease, M.E.; Kerrigan, D.F.; Mitchell, R.S. Number of ganglion cells in glaucoma eyes compared with threshold visual field tests in the same persons. Investig. Ophthalmol. Vis. Sci. 2000, 41, 741–748. [Google Scholar]
- Wang, L.H.; Huang, C.H.; Lin, I.C. Advances in Neuroprotection in glaucoma: Pharmacological strategies and emerging technologies. Pharmaceuticals 2024, 17, 1261. [Google Scholar] [CrossRef]
- Becerra, C.M.C.; Funk, R.O.; Kohli, D.; Hodge, D.O.; Roddy, G.W. Evaluating the association between autoimmune disease and normal tension glaucoma: A retrospective case-control study. BMC Ophthalmol. 2025, 25, 73. [Google Scholar] [CrossRef]
- Liang, S. Role of T cell-induced autoimmune response in the pathogenesis of glaucoma. Int. Ophthalmol. 2024, 44, 241. [Google Scholar] [CrossRef]
- Wang, L.; Wei, X. T Cell-Mediated Autoimmunity in Glaucoma Neurodegeneration. Front. Immunol. 2021, 12, 803485. [Google Scholar] [CrossRef]
- Tezel, G.; Seigel, G.M.; Wax, M.B. Autoantibodies to small heat shock proteins in glaucoma. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2277–2287. [Google Scholar]
- Maruyama, I.; Ikeda, Y.; Nakazawa, M.; Ohguro, H. Clinical roles of serum autoantibody against neuron-specific enolase in glaucoma patients. Tohoku J. Exp. Med. 2002, 197, 125–132. [Google Scholar] [CrossRef]
- Shin, Y.J.; Kim, E.; Han, B.K.; Yi, K. Serum biomarkers for the diagnosis of glaucoma. Diagnostics 2020, 11, 20. [Google Scholar] [CrossRef]
- Auler, N.; Tonner, H.; Pfeiffer, N.; Gru, F.H. Antibody and Protein Profiles in Glaucoma: Screening of Biomarkers and Identification of Signaling Pathways. Biology 2021, 10, 1296. [Google Scholar] [CrossRef]
- Goshima, N.; Kawamura, Y.; Fukumoto, A.; Miura, A.; Honma, R.; Satoh, R.; Wakamatsu, A.; Yamamoto, J.; Kimura, K.; Nishikawa, T.; et al. Human protein factory for converting the transcriptome into an in vitro-expressed proteome. Nat. Methods 2008, 5, 1011–1017. [Google Scholar] [CrossRef]
- Fukuda, E.; Tanaka, H.; Yamaguchi, K.; Takasaka, M.; Kawamura, Y.; Okuda, H.; Isotani, A.; Ikawa, M.; Shapiro, V.S.; Tsuchida, J.; et al. Identification and characterization of the antigen recognized by the germ cell mAb TRA98 using a human comprehensive wet protein array. Genes Cells 2021, 26, 180–189. [Google Scholar] [CrossRef]
- Matsuda, K.M.; Yoshizaki, A.; Yamaguchi, K.; Fukuda, E.; Okumura, T.; Ogawa, K.; Ono, C.; Norimatsu, Y.; Kotani, H.; Hisamoto, T.; et al. Autoantibody landscape revealed by wet protein array: Sum of autoantibody levels reflects disease status. Front. Immunol. 2022, 13, 893086. [Google Scholar] [CrossRef]
- Matsuda, K.M.; Kotani, H.; Yamaguchi, K.; Okumura, T.; Fukuda, E.; Kono, M.; Hisamoto, T.; Kawanabe, R.; Norimatsu, Y.; Kuzumi, A.; et al. Significance of anti-transcobalamin receptor antibodies in cutaneous arteritis revealed by proteome-wide autoantibody screening. J. Autoimmun. 2023, 135, 102995. [Google Scholar] [CrossRef]
- Anderson, D.R.; Chauhan, B.; Johnson, C.; Katz, J.; Patella, V.M.; Drance, S.M. Criteria for progression of glaucoma in clinical management and in outcome studies. J. Ophthalmol. 2000, 130, 827–829. [Google Scholar] [CrossRef]
- Takahashi, N.; Omodaka, K.; Nakazawa, A.; Kikawa, T.; Ninomiya, T.; Kiyota, N.; Tsuda, S.; Himori, N.; Akiba, M.; Nakazawa, T. Correlation Between Enlargement of Retinal Nerve Fiber Defect Angle in En Face Imaging and Visual Field Progression. Transl. Vis. Sci. Technol. 2022, 11, 8. [Google Scholar] [CrossRef]
- Prokofyeva, E.; Wegener, A.; Zrenner, E. Cataract prevalence and prevention in Europe: A literature review. Acta Ophthalmol. 2013, 91, 395–405. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, J.; Yang, F.; Xu, B.; Tang, Y.; Lu, Y. Prevalence rates of cataract and cataract surgery in elderly Chinese people living in suburban Shanghai: The Pujiang Cataract Cohort Study. Br. J. Ophthalmol. 2023, 107, 683–689. [Google Scholar] [CrossRef]
- Lakk, M.; Krizaj, D. TRPV4-Rho signaling drives cytoskeletal and focal adhesion remodeling in trabecular meshwork cells. Am. J. Physiol. Cell Physiol. 2021, 320, C1013–C1030. [Google Scholar] [CrossRef]
- Steenbergen, R.; Nanowski, T.S.; Beigneux, A.; Kulinski, A.; Young, S.G.; Vance, J.E. Disruption of the phosphatidylserine decarboxylase gene in mice causes embryonic lethality and mitochondrial defects. J. Biol. Chem. 2005, 280, 40032–40040. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Xie, Z.; Chen, S.Y.; Zhang, X. Mitochondrial dysfunction in glaucomatous degeneration. Int. J. Ophthalmol. 2023, 16, 811–823. [Google Scholar] [CrossRef]
- Chaphalkar, R.M.; Kodati, B.; Maddineni, P.; He, S.; Brooks, C.D.; Stankowska, D.L.; Yang, S.; Zode, G.; Krishnamoorthy, R.R. A Reduction in Mitophagy Is Associated with Glaucomatous Neurodegeneration in Rodent Models of Glaucoma. Int. J. Mol. Sci. 2024, 25, 13040. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Irie, K.; Kurihara, H.; Sakai, T.; Takai, Y. Vmac: A novel protein associated with vimentin-type intermediate filament in podocytes of rat kidney. Biochem. Biophys. Res. Commun. 2004, 315, 1120–1125. [Google Scholar] [CrossRef]
- Rutigliani, C.; Tribble, J.R.; Hagström, A.; Lardner, E.; Jóhannesson, G.; Stålhammar, G.; Williams, P.A. Widespread retina and optic nerve neuroinflammation in enucleated eyes from glaucoma patients. Acta Neuropathol. Commun. 2022, 10, 118. [Google Scholar] [CrossRef]
- Jiravarnsirikul, A.; Belghith, A.; Rezapour, J.; Bowd, C.; Moghimi, S.; Jonas, J.B.; Christopher, M.; Fazio, M.A.; Yang, H.; Burgoyne, C.F. Evaluating glaucoma in myopic eyes: Challenges and opportunities. Surv. Ophthalmol. 2024, 70, 563–582. [Google Scholar] [CrossRef]
- Jiang, L.; Koh, J.H.Z.; Seah, S.H.Y.; Dan, Y.S.; Wang, Z.; Chan, X.; Zhou, L.; Barathi, V.A.; Hoang, Q.V. Key role for inflammation-related signaling in the pathogenesis of myopia based on evidence from proteomics analysis. Sci. Rep. 2024, 14, 23486. [Google Scholar] [CrossRef]
- Ueda, N.; Maekawa, M.; Matsui, T.S.; Deguchi, S.; Takata, T.; Katahira, J.; Higashiyama, S.; Hieda, M. Inner Nuclear Membrane Protein, SUN1, is Required for Cytoskeletal Force Generation and Focal Adhesion Maturation. Front. Cell Dev. Biol. 2022, 10, 885859. [Google Scholar] [CrossRef]
- Badrinarayanan, L.; Nagarajan, H.; Rishi, P.; Rishi, E.; George, R.J.; Chitipothu, S. Whole-genome sequencing unravels novel genetic determinants and regulatory pathways associated with triamcinolone acetonide-induced ocular hypertension. Mol. Genet. Genom. 2023, 298, 13–26. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, D.; Zhou, Q.; Zhao, F.; He, M.; Yang, Z.; Su, Y.; Zhai, Y.; Yan, J.; Zhang, G.; et al. Scleral HIF-1alpha is a prominent regulatory candidate for genetic and environmental interactions in human myopia pathogenesis. EBioMedicine 2020, 57, 102878. [Google Scholar] [CrossRef]
- Bao, B.; Liu, J.; Li, T.; Yang, Z.; Wang, G.; Xin, J.; Bi, H.; Guo, D. Elevated retinal fibrosis in experimental myopia is involved in the activation of the PI3K/AKT/ERK signaling pathway. Arch. Biochem. Biophys. 2023, 743, 109663. [Google Scholar] [CrossRef]
- Kiyota, N.; Shiga, Y.; Omodaka, K.; Pak, K.; Nakazawa, T. Time-Course Changes in Optic Nerve Head Blood Flow and Retinal Nerve Fiber Layer Thickness in Eyes with Open-angle Glaucoma. Ophthalmology 2021, 128, 663–671. [Google Scholar] [CrossRef]
- Sato, K.; Takada, N.; Fujioka, A.; Himori, N.; Yokoyama, Y.; Tsuda, S.; Omodaka, K.; Kirihara, T.; Ishikawa, M.; Kunikata, H.; et al. Reduced Plasma BDNF Levels in Normal Tension Glaucoma Compared to Open Angle Glaucoma. J. Glaucoma 2023, 32, 734–737. [Google Scholar] [CrossRef]
- Sato, K.; Saigusa, D.; Kokubun, T.; Fujioka, A.; Feng, Q.; Saito, R.; Uruno, A.; Matsukawa, N.; Ohno-Oishi, M.; Kunikata, H.; et al. Reduced glutathione level in the aqueous humor of patients with primary open-angle glaucoma and normal-tension glaucoma. PJ Aging 2023, 9, 28. [Google Scholar] [CrossRef]
- Von Thun Und Hohenstein-Blaul, N.; Kunst, S.; Pfeiffer, N.; Grus, F.H. Biomarkers for glaucoma: From the lab to the clinic. Eye 2017, 31, 225–231. [Google Scholar] [CrossRef]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Antibodies to alpha B-crystallin, vimentin, and heat shock protein 70 in patients with normal tension glaucoma and IgG antibody patterns against retinal antigen in aqueous humor. Curr. Eye Res. 2007, 32, 501–509. [Google Scholar] [CrossRef]
- Tsai, T.; Grotegut, P.; Reinehr, S.; Joachim, S.C. Role of Heat Shock Proteins in Glaucoma. Int. J. Mol. Sci. 2019, 20, 5160. [Google Scholar] [CrossRef]
- Vanags, D.; Williams, B.; Johnson, B.; Hall, S.; Nash, P.; Taylor, A.; Weiss, J.; Feeney, D. Therapeutic efficacy and safety of chaperonin 10 in patients with rheumatoid arthritis: A double-blind randomised trial. Lancet 2006, 368, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Gupta, V.B.; Chick, J.M.; Greco, T.M.; Wu, Y.; Chitranshi, N.; Wall, R.V.; Hone, E.; Deng, L.; Dheer, Y.; et al. Age-related neurodegenerative disease associated pathways identified in retinal and vitreous proteome from human glaucoma eyes. Sci. Rep. 2017, 7, 12685. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.; Wilding, C.; Funke, S.; Pfeiffer, N.; Grus, F.H. Protective effect of 14-3-3 antibodies on stressed neuroretinal cells via the mitochondrial apoptosis pathway. BMC Ophthalmol. 2015, 15, 64. [Google Scholar] [CrossRef] [PubMed]
- Yap, T.E.; Davis, B.M.; Guo, L.; Normando, E.M.; Cordeiro, M.F. Annexins in Glaucoma. Int. J. Mol. Sci. 2018, 19, 1218. [Google Scholar] [CrossRef]
- Joachim, S.C.; Reichelt, J.; Berneiser, S.; Pfeiffer, N.; Grus, F.H. Sera of glaucoma patients show autoantibodies against myelin basic protein and complex autoantibody profiles against human optic nerve antigens. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 573–580. [Google Scholar] [CrossRef]
- Beutgen, V.M.; Perumal, N.; Pfeiffer, N.; Grus, F.H. Autoantibody biomarker discovery in primary open angle glaucoma using serological proteome analysis (SERPA). Front. Immunol. 2019, 10, 381. [Google Scholar] [CrossRef]
- Maruyama, I.; Ohguro, H.; Ikeda, Y. Retinal ganglion cells recognized by serum autoantibody against gamma-enolase found in glaucoma patients. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1657–1665. [Google Scholar]
- Boehm, N.; Wolters, D.; Thiel, U.; Lossbrand, U.; Wietgel, N.; Pfeiffer, N.; Grus, F.H. New insights into autoantibody profiles from immune privileged sites in the eye: A glaucoma study. Brain Behav. Immun. 2012, 26, 96–102. [Google Scholar] [CrossRef]
- Latalska, M.; Gerkowicz, M.; Kosior-Jarecka, E.; Kozioł-Montewka, M.; Pietraś-Trzpiel, M. Antibodies to beta-2 glycoprotein I in serum and aqueous humor of patients with glaucoma and their influence on the static perimetry. Klin. Oczna 2004, 106, 160–161. [Google Scholar]
- Surgucheva, I.; McMahan, B.; Ahmed, F.; Tomarev, S.; Wax, M.B.; Surguchov, A. Synucleins in glaucoma: Implication of gamma-synuclein in glaucomatous alterations in the optic nerve. J. Neurosci. Res. 2002, 68, 97–106. [Google Scholar] [CrossRef]
- Saccà, S.C.; Centofanti, M.; Izzotti, A. New proteins as vascular biomarkers in primary open angle glaucomatous aqueous humor. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4242–4253. [Google Scholar] [CrossRef] [PubMed]
- Romano, C.; Barrett, D.A.; Li, Z.; Pestronk, A.; Wax, M.B. Anti-rhodopsin antibodies in sera from patients with normal-pressure glaucoma. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1968–1975. [Google Scholar] [PubMed]
- Grus, F.H.; Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Wax, M.B. Serum autoantibodies to alpha-fodrin are present in glaucoma patients from Germany and the United States. Investig. Opthalmol Vis. Sci. 2006, 47, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, A.; Sheng, M.H.; Constantine-Paton, M. Eye opening induces a rapid dendritic localization of PSD-95 in central visual neurons. Proc. Natl. Acad. Sci. USA 2003, 100, 1334–1339. [Google Scholar] [CrossRef]
- García-Barcina, J.M.; Matute, C. AMPA-selective glutamate receptor subunits in glial cells of adult bovine white matter. Brain Res. Mol. Brain Res. 1998, 53, 270–276. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Zhang, D.; Ozkaynak, E.; Wang, X.; Kasper, E.M.; Leguern, E.; Baulac, S.; Anderson, M.P. Epilepsy gene LGI1 regulates postnatal developmental remodeling of retinogeniculate synapses. J. Neurosci. 2012, 32, 903–910. [Google Scholar] [CrossRef]
- Lennon, V.A.; Wingerchuk, D.M.; Kryzer, T.J.; Pittock, S.J.; Lucchinetti, C.F.; Fujihara, K.; Nakashima, I.; Weinshenker, B.G. A serum autoantibody marker of neuromyelitis optica: Distinction from multiple sclerosis. Lancet 2004, 364, 2106–2112. [Google Scholar] [CrossRef]
- Kezuka, T.; Ishikawa, H. Diagnosis and treatment of anti-myelin oligodendrocyte glycoprotein antibody positive optic neuritis. Jpn. J. Ophthalmol. 2018, 62, 101–108. [Google Scholar] [CrossRef]
- Yasin, A.; Dudeck, L.; Redick, D.W.; Khodeiry, M.M.; Lam, B.L.; Jiang, H. Severe vision loss and optic disc edema associated with GAD-65 antibody positive Miller Fisher syndrome. J. Neuroophthalmol. 2022, 44, e40–e44. [Google Scholar] [CrossRef]
- Huntsman, M.M.; Jones, E.G. Expression of alpha3, beta3 and gamma1 GABA(A) receptor subunit messenger RNAs in visual cortex and lateral geniculate nucleus of normal and monocularly deprived monkeys. Neuroscience 1998, 87, 385–400. [Google Scholar] [CrossRef]
- Moldavan, M.G.; Allen, C.N. GABAB receptor-mediated frequency-dependent and circadian changes in synaptic plasticity modulate retinal input to the suprachiasmatic nucleus. J. Physiol. 2013, 591, 2475–24390. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Yang, H.; Zhu, F.; Wang, Q.; Shan, W. Clinical character of CASPR2 autoimmune encephalitis: A multiple center retrospective study. Front. Immunol. 2021, 12, 652864. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Jiang, M.; Xu, S.; Yang, L.; Yang, P.; Song, Y.; Zhu, H.; Wang, Y.; Sun, Y.; Yan, C.; et al. Mirror image pain mediated by D2 receptor regulation of astrocytic Cx43 phosphorylation and channel opening. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166657. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.A.; Rios, J.C.; Lu, Y.; Garcia-Fresco, G.P.; Ching, W.; Martin, M.S.; Li, J.; Einheber, S.; Chesler, M.; Rosenbluth, J.; et al. Axon-glia interactions and the domain organization of myelinated axons requires neurexin IV/Caspr/Paranodin. Neuron 2001, 30, 369–383. [Google Scholar] [CrossRef]
- Keeley, P.W.; Reese, B.E. DNER and NFIA are expressed by developing and mature AII amacrine cells in the mouse retina. J. Comp. Neurol. 2018, 526, 467–479. [Google Scholar] [CrossRef]
- Murali, S.S.; Napier, I.A.; Mohammadi, S.A.; Alewood, P.F.; Lewis, R.J.; Christie, M.J. High-voltage-activated calcium current subtypes in mouse DRG neurons adapt in a subpopulation-specific manner after nerve injury. J. Neurophysiol. 2015, 113, 1511–1519. [Google Scholar] [CrossRef]
- Butt, A.M.; Vanzulli, I.; Papanikolaou, M.; De La Rocha, I.C.; Hawkins, V.E. Metabotropic glutamate receptors protect oligodendrocytes from acute ischemia in mouse optic nerve. Neurochem. Res. 2017, 42, 2468–2478. [Google Scholar] [CrossRef]
- Simmonds, M.A. Depolarizing responses to glycine, beta-alanine and muscimol in isolated optic nerve and cuneate nucleus. Br. J. Pharmacol. 1983, 79, 799–806. [Google Scholar] [CrossRef]
- Pittock, S.J.; Lucchinetti, C.F.; Parisi, J.E.; Benarroch, E.E.; Mokri, B.; Stephan, C.L.; Kim, K.; Kilimann, M.W.; Lennon, V.A. Amphiphysin autoimmunity: Paraneoplastic accompaniments. Ann. Neurol. 2005, 58, 96–107. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cat | OAG | p Value cat vs. OAG | p Value NTG vs. POAG | |||
|---|---|---|---|---|---|---|
| Total | NTG | POAG | ||||
| N | 35 | 119 | 50 | 69 | ||
| Age (mean (SD)) | 69.97 (10.84) | 68.76 (7.03) | 68.48 (6.04) | 68.96 (7.71) | 0.433 | 0.717 |
| Sex = Male (%) | 18 (51.4) | 68 (57.1) | 25 (50.0) | 43 (62.3) | 0.686 | 0.249 |
| IOP (mean (SD)) | 14.06 (3.31) | 14.54 (4.43) | 12.67 (2.54) | 15.89 (4.99) | 0.553 | <0.001 * |
| MD (mean (SD)) | −17.50 (7.98) | −19.27 (8.93) | 0.268 | |||
| AL (mean (SD)) | 24.50 (2.29) | 25.24 (1.64) | 25.15 (1.45) | 25.31 (1.77) | 0.042 * | 0.616 |
| CCT (mean (SD)) | 509.00 (34.07) | 502.03 (35.83) | 494.46 (32.07) | 507.52 (37.61) | 0.332 | 0.049* |
| OAG | ||||
|---|---|---|---|---|
| ETNK1 | VMAC | NEXN | SUN1 | |
| MD | 0.057 | −0.020 | −0.182 | −0.011 |
| IOP | 0.095 | 0.128 | −0.022 | −0.005 |
| AL | −0.045 | −0.042 | 0.073 | 0.027 |
| CCT | 0.167 | 0.043 | 0.008 | 0.0117 |
| Age | −0.035 | −0.066 | 0.183 | 0.001 |
| NTG | ||||
| ETNK1 | VMAC | NEXN | SUN1 | |
| MD | 0.057 | −0.020 | −0.182 | −0.011 |
| IOP | 0.095 | 0.128 | −0.022 | −0.005 |
| AL | −0.045 | −0.042 | 0.073 | 0.027 |
| CCT | 0.167 | 0.043 | 0.008 | 0.011 |
| Age | −0.035 | −0.066 | 0.183 | 0.001 |
| POAG | ||||
| ETNK1 | VMAC | NEXN | SUN1 | |
| MD | 0.234 | 0.068 | −0.168 | 0.104 |
| IOP | −0.178 | −0.098 | −0.033 | −0.205 |
| AL | −0.079 | −0.040 | 0.038 | −0.006 |
| CCT | 0.123 | 0.055 | 0.015 | 0.112 |
| Age | −0.187 | −0.094 | 0.273 * | −0.077 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takada, N.; Ishikawa, M.; Sato, K.; Kunikata, H.; Ninomiya, T.; Hanyuda, A.; Fukuda, E.; Yamaguchi, K.; Ono, C.; Kirihara, T.; et al. Proteome-Wide Analysis of Autoantibodies in Open-Angle Glaucoma in Japanese Population: A Pilot Study. Biomedicines 2025, 13, 718. https://doi.org/10.3390/biomedicines13030718
Takada N, Ishikawa M, Sato K, Kunikata H, Ninomiya T, Hanyuda A, Fukuda E, Yamaguchi K, Ono C, Kirihara T, et al. Proteome-Wide Analysis of Autoantibodies in Open-Angle Glaucoma in Japanese Population: A Pilot Study. Biomedicines. 2025; 13(3):718. https://doi.org/10.3390/biomedicines13030718
Chicago/Turabian StyleTakada, Naoko, Makoto Ishikawa, Kota Sato, Hiroshi Kunikata, Takahiro Ninomiya, Akiko Hanyuda, Eriko Fukuda, Kei Yamaguchi, Chihiro Ono, Tomoko Kirihara, and et al. 2025. "Proteome-Wide Analysis of Autoantibodies in Open-Angle Glaucoma in Japanese Population: A Pilot Study" Biomedicines 13, no. 3: 718. https://doi.org/10.3390/biomedicines13030718
APA StyleTakada, N., Ishikawa, M., Sato, K., Kunikata, H., Ninomiya, T., Hanyuda, A., Fukuda, E., Yamaguchi, K., Ono, C., Kirihara, T., Shintani, C., Tsusu, C., Osanai, A., Goshima, N., Izumi, Y., Zorumski, C. F., & Nakazawa, T. (2025). Proteome-Wide Analysis of Autoantibodies in Open-Angle Glaucoma in Japanese Population: A Pilot Study. Biomedicines, 13(3), 718. https://doi.org/10.3390/biomedicines13030718