
Impact of Long-Lasting Environmental Factors on Regulation Mediated by the miR-34 Family



Abstract
1. Introduction
2. Functions of Small Non-Coding RNAs and Their Modulation by Environmental Factors
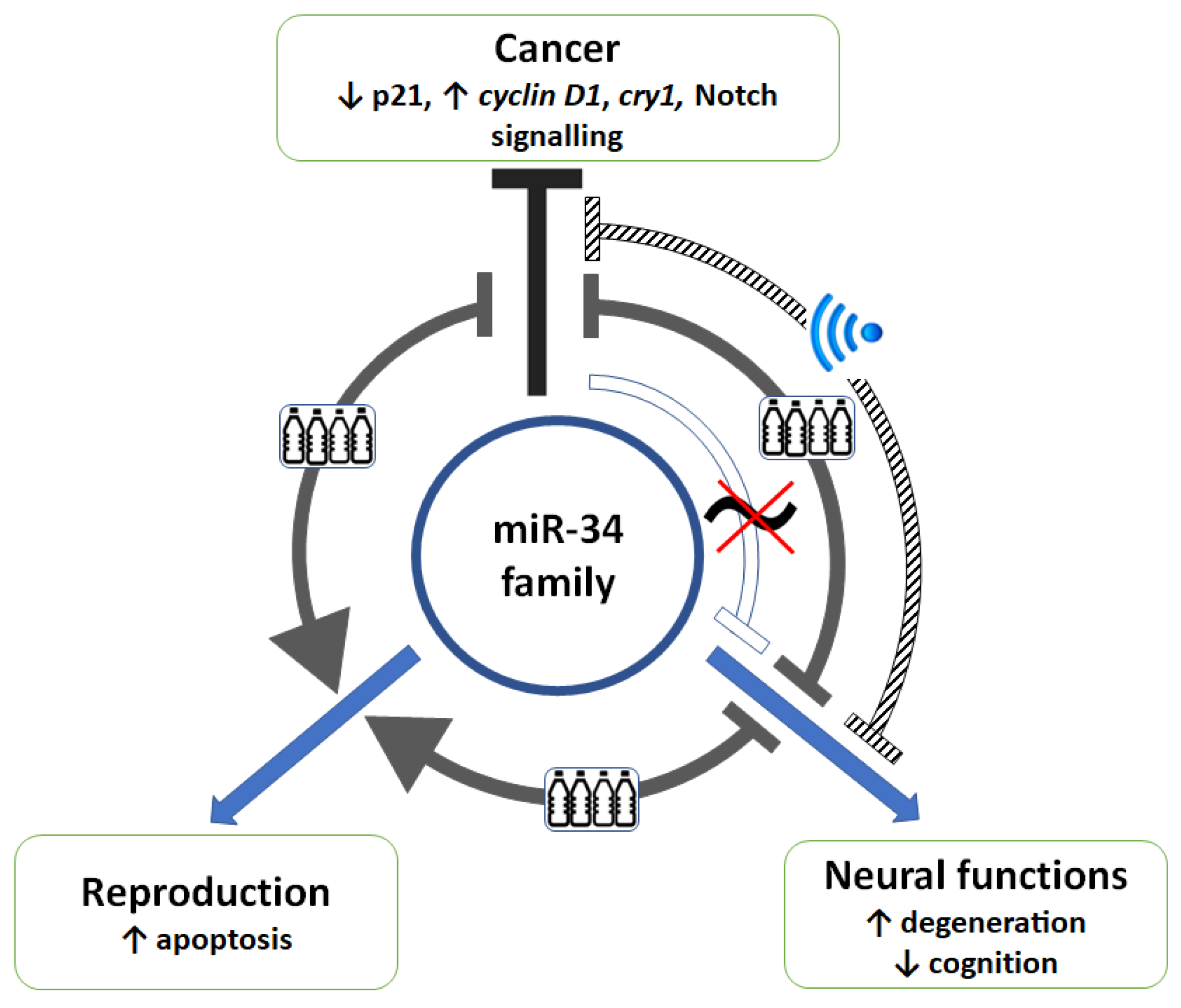
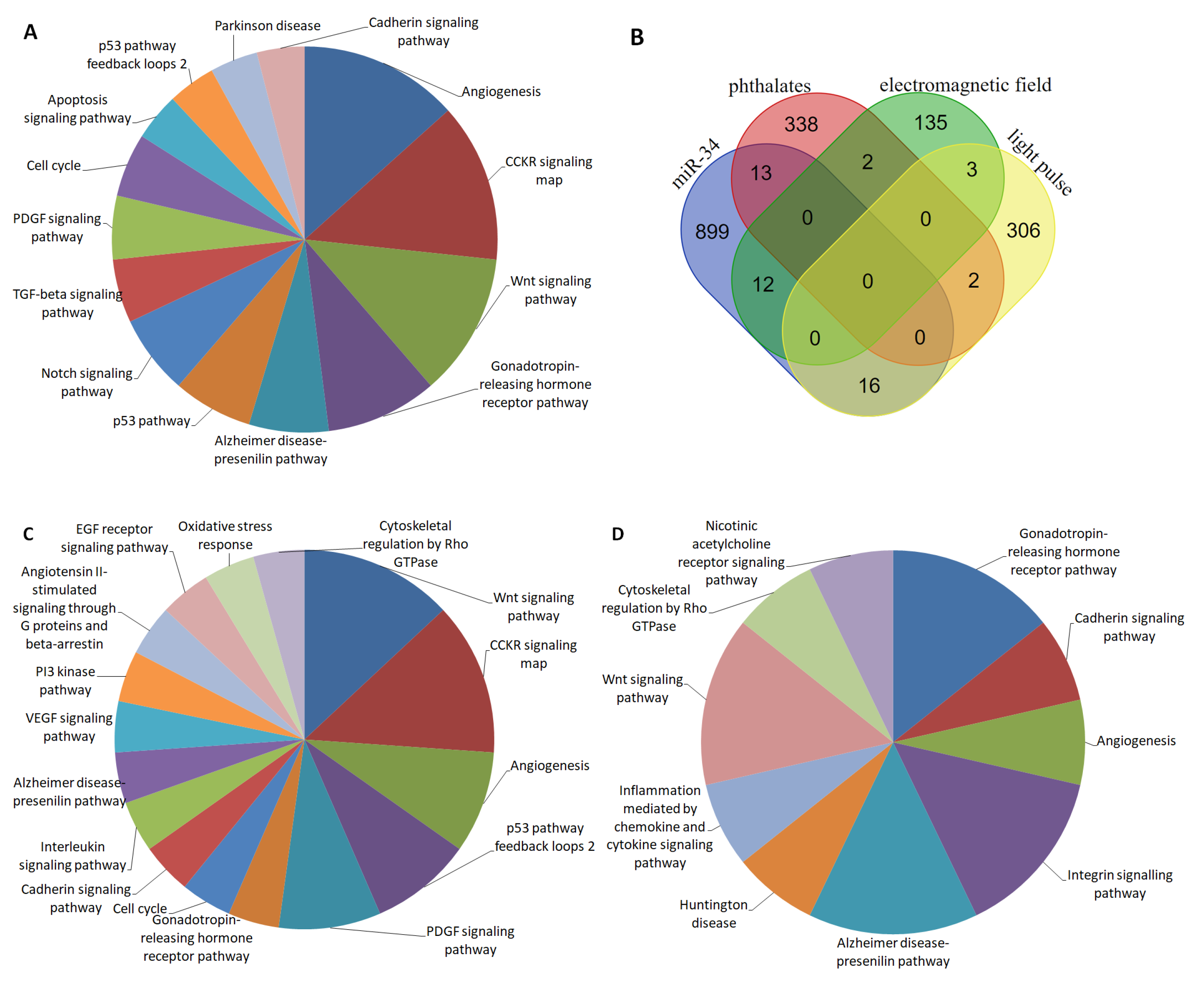
2.1. Interference of Phthalates on the Functions Executed by miR-34
2.2. Interference of an Electromagnetic Field on miR-34-Mediated Regulation
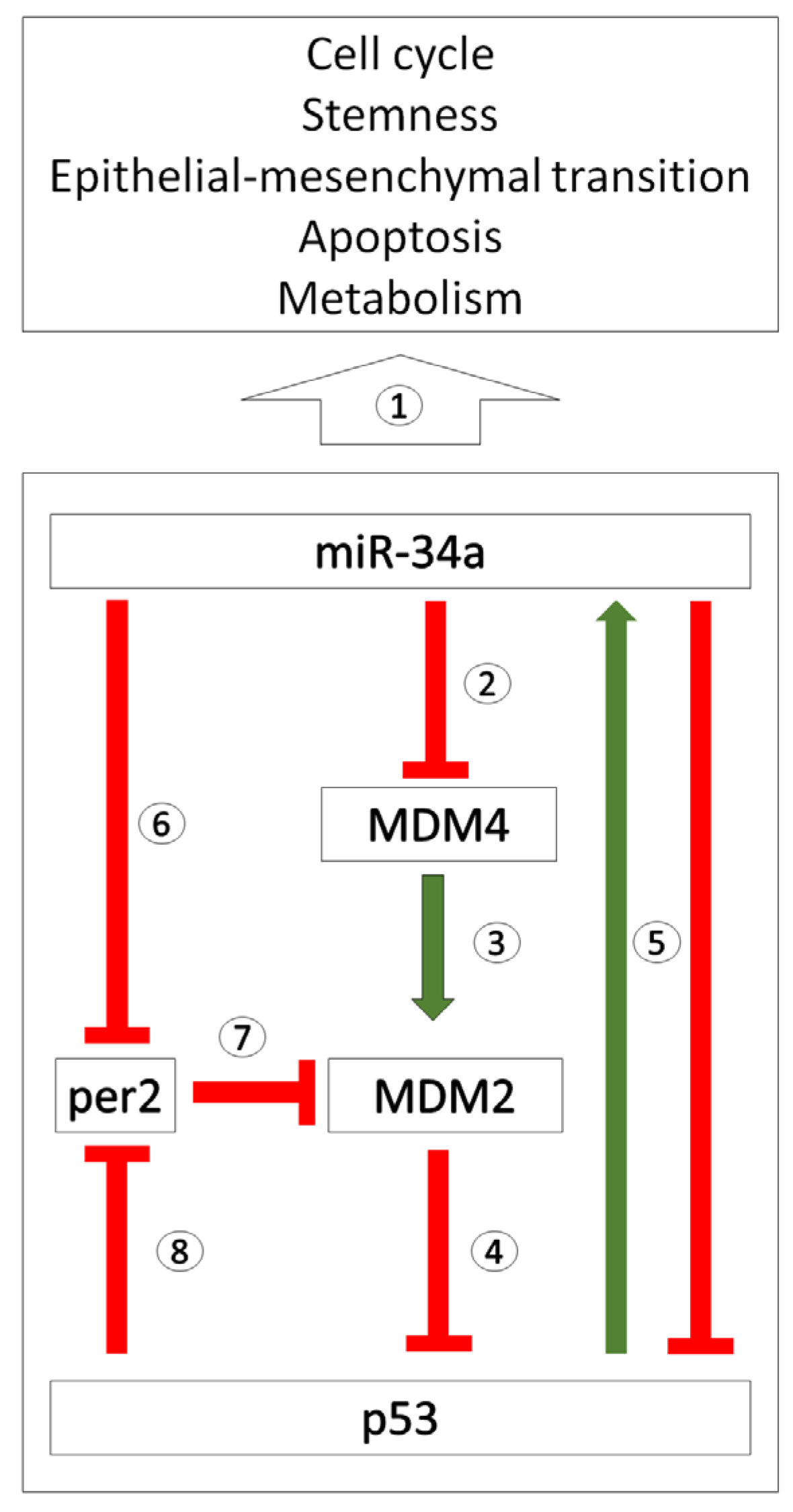
2.3. miR-34 Signalling and Its Interaction with the Circadian System
3. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef]
- Fu, Z.; Wang, L.; Li, S.; Chen, F.; Au-Yeung, K.K.; Shi, C. MicroRNA as an Important Target for Anticancer Drug Development. Front. Pharmacol. 2021, 12, 736323. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. The microRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genomics 2010, 11, 537–561. [Google Scholar] [CrossRef]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Bushati, N.; Cohen, S.M. microRNA functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef]
- Li, M.; Marin-Muller, C.; Bharadwaj, U.; Chow, K.H.; Yao, Q.; Chen, C. MicroRNAs: Control and loss of control in human physiology and disease. World J. Surg. 2009, 33, 667–684. [Google Scholar] [CrossRef]
- Voglova, K.; Bezakova, J.; Herichova, I. Micro RNAs: An arguable appraisal in medicine. Endocr. Regul. 2016, 50, 106–124. [Google Scholar] [CrossRef] [PubMed]
- Voglova, K.; Bezakova, J.; Herichova, I. Progress in micro RNA focused research in endocrinology. Endocr. Regul. 2016, 50, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Dhuppar, S.; Murugaiyan, G. miRNA effects on gut homeostasis: Therapeutic implications for inflammatory bowel disease. Trends Immunol. 2022, 43, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Gulyaeva, L.F.; Kushlinskiy, N.E. Regulatory mechanisms of microRNA expression. J. Transl. Med. 2016, 14, 143. [Google Scholar] [CrossRef]
- Pidíkova, P.; Reis, R.; Herichova, I. miRNA Clusters with Down-Regulated Expression in Human Colorectal Cancer and Their Regulation. Int. J. Mol. Sci. 2020, 21, 4633. [Google Scholar] [CrossRef]
- Pidíková, P.; Herichová, I. miRNA Clusters with Up-Regulated Expression in Colorectal Cancer. Cancers 2021, 13, 2979. [Google Scholar] [CrossRef]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. microRNA strand selection: Unwinding the rules. Wiley Interdiscip. Rev. RNA 2021, 12, e1627. [Google Scholar] [CrossRef] [PubMed]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Herichová, I. Angiotensin II in the Human Physiology: Novel Ways for Synthetic Compounds Utilization. Curr. Med. Chem. 2016, 23, 4735–4752. [Google Scholar] [CrossRef] [PubMed]
- Sadakierska-Chudy, A. MicroRNAs: Diverse Mechanisms of Action and Their Potential Applications as Cancer Epi-Therapeutics. Biomolecules 2020, 10, 1285. [Google Scholar] [CrossRef] [PubMed]
- Plotnikova, O.; Baranova, A.; Skoblov, M. Comprehensive Analysis of Human microRNA-mRNA Interactome. Front. Genet. 2019, 10, 933. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. miRTarBase update 2022: An informative resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef]
- Surendran, S.; Jideonwo, V.N.; Merchun, C.; Ahn, M.; Murray, J.; Ryan, J.; Dunn, K.W.; Kota, J.; Morral, N. Gene targets of mouse miR-709: Regulation of distinct pools. Sci. Rep. 2016, 6, 18958. [Google Scholar] [CrossRef]
- Loganathan, T.; Doss, C.G.P. Non-coding RNAs in human health and disease: Potential function as biomarkers and therapeutic targets. Funct. Integr. Genom. 2023, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, G. miRNAs: The Road from Bench to Bedside. Genes 2023, 14, 314. [Google Scholar] [CrossRef]
- Misso, G.; Di Martino, M.T.; De Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gullà, A.; Tagliaferri, P.; Tassone, P.; et al. Mir-34: A new weapon against cancer? Mol. Ther. Nucleic Acids 2014, 3, e194. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 family: A potential tumor suppressor and therapeutic candidate in cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, X.; Lim, L.P.; de Stanchina, E.; Xuan, Z.; Liang, Y.; Xue, W.; Zender, L.; Magnus, J.; Ridzon, D.; et al. A microRNA component of the p53 tumour suppressor network. Nature 2007, 447, 1130–1134. [Google Scholar] [CrossRef]
- Okada, N.; Lin, C.P.; Ribeiro, M.C.; Biton, A.; Lai, G.; He, X.; Bu, P.; Vogel, H.; Jablons, D.M.; Keller, A.C.; et al. A positive feedback between p53 and miR-34 miRNAs mediates tumor suppression. Genes Dev. 2014, 28, 438–450. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J. miR-34a repression of SIRT1 regulates apoptosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13421–13426. [Google Scholar] [CrossRef]
- Sargolzaei, J.; Etemadi, T.; Alyasin, A. The P53/microRNA network: A potential tumor suppressor with a role in anticancer therapy. Pharmacol. Res. 2020, 160, 105179. [Google Scholar] [CrossRef]
- Li, W.J.; Wang, Y.; Liu, R.; Kasinski, A.L.; Shen, H.; Slack, F.J.; Tang, D.G. MicroRNA-34a: Potent Tumor Suppressor, Cancer Stem Cell Inhibitor, and Potential Anticancer Therapeutic. Front. Cell Dev. Biol. 2021, 9, 640587. [Google Scholar] [CrossRef]
- Pan, W.; Chai, B.; Li, L.; Lu, Z.; Ma, Z. p53/MicroRNA-34 axis in cancer and beyond. Heliyon 2023, 9, e15155. [Google Scholar] [CrossRef]
- Cao, L.; Liu, Y.; Lu, J.B.; Miao, Y.; Du, X.Y.; Wang, R.; Yang, H.; Xu, W.; Li, J.Y.; Fan, L. A feedback circuit of miR-34a/MDM4/p53 regulates apoptosis in chronic lymphocytic leukemia cells. Transl. Cancer Res. 2020, 9, 6143–6153. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, H.; Hu, W. The regulation of MDM2 oncogene and its impact on human cancers. Acta Biochim. Biophys. Sin. 2014, 46, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Navarro, F.; Lieberman, J. miR-34 and p53: New Insights into a Complex Functional Relationship. PLoS ONE 2015, 10, e0132767. [Google Scholar] [CrossRef]
- Olejárová, S.; Moravčík, R.; Herichová, I. 2.4 GHz Electromagnetic Field Influences the Response of the Circadian Oscillator in the Colorectal Cancer Cell Line DLD1 to miR-34a-Mediated Regulation. Int. J. Mol. Sci. 2022, 23, 13210. [Google Scholar] [CrossRef] [PubMed]
- Moravčík, R.; Olejárová, S.; Zlacká, J.; Herichová, I. Effect of miR-34a on the expression of clock and clock-controlled genes in DLD1 and Lovo human cancer cells with different backgrounds with respect to p53 functionality and 17β-estradiol-mediated regulation. PLoS ONE 2023, 18, e0292880. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Pelicano, H.; Liu, J.; Huang, P.; Lee, C. The circadian gene Period2 plays an important role in tumor suppression and DNA damage response in vivo. Cell 2002, 111, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, T.; Vila-Caballer, M.; Santos, C.S.; Liu, J.; Yang, J.; Finkielstein, C.V. The circadian factor Period 2 modulates p53 stability and transcriptional activity in unstressed cells. Mol. Biol. Cell 2014, 25, 3081–3093. [Google Scholar] [CrossRef]
- Miki, T.; Matsumoto, T.; Zhao, Z.; Lee, C.C. p53 regulates Period2 expression and the circadian clock. Nat. Commun. 2013, 4, 2444. [Google Scholar] [CrossRef]
- Turkoglu, F.; Calisir, A.; Ozturk, B. Clinical importance of serum miRNA levels in breast cancer patients. Discov. Oncol. 2024, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Saffar, K.N.; Larypoor, M.; Torbati, M.B. Analyzing of colorectal cancerrelated genes and microRNAs expression profiles in response to probiotics Lactobacillus acidophilus and Saccharomyces cerevisiae in colon cancer cell lines. Mol. Biol. Rep. 2024, 51, 122. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhong, X.; Zhao, Y.; Shen, J.; Xiao, Z. Acacetin inhibited non-small-cell lung cancer (NSCLC) cell growth via upregulating miR-34a in vitro and in vivo. Sci. Rep. 2024, 14, 2348. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-F.; Lin, C.-S.; Shen, P.-C.; Chuang, S.-E.; Dai, Y.-H.; Huang, T.-W.; Lin, C.-Y.; Hung, Y.-J.; Shieh, Y.-S. MiR-34a functions as a tumor suppressor in oral cancer through the inhibition of the Axl/Akt/GSK-3β pathway. J. Dent. Sci. 2024, 19, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Alden, N.A.; Yeingst, T.J.; Pfeiffer, H.M.; Celik, N.; Arrizabalaga, J.H.; Helton, A.M.; Liu, Y.; Stairs, D.B.; Glick, A.B.; Goyal, N.; et al. Near-Infrared Induced miR-34a Delivery from Nanoparticles in Esophageal Cancer Treatment. Adv. Healthc. Mater. 2024, 12, e2303593. [Google Scholar] [CrossRef]
- Fu, J.; Imani, S.; Wu, M.Y.; Wu, R.C. MicroRNA-34 Family in Cancers: Role, Mechanism, and Therapeutic Potential. Cancers 2023, 15, 4723. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Mockly, S.; Houbron, É.; Seitz, H. A rationalized definition of general tumor suppressor microRNAs excludes miR-34a. Nucleic Acids Res. 2022, 50, 4703–4712. [Google Scholar] [CrossRef]
- Abdelaal, A.M.; Sohal, I.S.; Iyer, S.; Sudarshan, K.; Kothandaraman, H.; Lanman, N.A.; Low, P.S.; Kasinski, A.L. A first-in-class fully modified version of miR-34a with outstanding stability, activity, and anti-tumor efficacy. Oncogene 2023, 42, 2985–2999. [Google Scholar] [CrossRef]
- Hermeking, H. MicroRNAs in the p53 network: Micromanagement of tumour suppression. Nat. Rev. Cancer 2012, 12, 613–626. [Google Scholar] [CrossRef]
- Siemens, H.; Jackstadt, R.; Hünten, S.; Kaller, M.; Menssen, A.; Götz, U.; Hermeking, H. miR-34 and SNAIL form a double-negative feedback loop to regulate epithelial-mesenchymal transitions. Cell Cycle 2011, 10, 4256–4271. [Google Scholar] [CrossRef]
- Schettler, T. Human exposure to phthalates via consumer products. Int. J. Androl. 2006, 29, 134–139; discussion 181–185. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Sicińska, P.; Mokra, K.; Wozniak, K.; Michałowicz, J.; Bukowska, B. Genotoxic risk assessment and mechanism of DNA damage induced by phthalates and their metabolites in human peripheral blood mononuclear cells. Sci. Rep. 2021, 11, 1658. [Google Scholar] [CrossRef]
- Hu, G.; Li, J.; Shan, Y.; Li, X.; Zhu, Q.; Li, H.; Wang, Y.; Chen, X.; Lian, Q.; Ge, R.S. In utero combined di-(2-ethylhexyl) phthalate and diethyl phthalate exposure cumulatively impairs rat fetal Leydig cell development. Toxicology 2018, 395, 23–33. [Google Scholar] [CrossRef]
- Štefánik, P.; Michalec, J.; Morová, M.; Olexová, L.; Kršková, L. Prenatal and perinatal phthalate exposure is associated with sex-dependent changes in hippocampal miR-15b-5p and miR-34a-5p expression and changes in testicular morphology in rat offspring. Arh. Hig. Rada Toksikol. 2022, 73, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Kougias, D.G.; Sellinger, E.P.; Willing, J.; Juraska, J.M. Perinatal Exposure to an Environmentally Relevant Mixture of Phthalates Results in a Lower Number of Neurons and Synapses in the Medial Prefrontal Cortex and Decreased Cognitive Flexibility in Adult Male and Female Rats. J. Neurosci. 2018, 38, 6864–6872. [Google Scholar] [CrossRef] [PubMed]
- Morová, M.; Senko, T.; Olexová, L.; Dzirbíková, Z.; Kršková, L. A mixture of diethylhexyl, diisononyl and dibutyl phthalate decreased anogenital distance, postnatal testosterone levels, and changed social behavior in Wistar rats. Physiol. Res. 2020, 69 (Suppl. S3), S489–S498. [Google Scholar] [CrossRef] [PubMed]
- Day, D.B.; Collett, B.R.; Barrett, E.S.; Bush, N.R.; Swan, S.H.; Nguyen, R.H.N.; Szpiro, A.A.; Sathyanarayana, S. Phthalate mixtures in pregnancy, autistic traits, and adverse childhood behavioral outcomes. Environ. Int. 2021, 147, 106330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, M.; Liu, J.; Ni, J.; Jiao, Y.; Bai, C. Up regulation of IL-6 is involved in di (2-ethylhexyl) phthalate (DEHP) induced migration and invasion of non small cell lung cancer (NSCLC) cells. Biomed. Pharmacother. 2017, 89, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Wang, Y.; Wang, X.; Wu, J.; Song, Q.; Sun, Z.; Zhang, Y. In utero and lactational exposure of DEHP increases the susceptibility of prostate carcinogenesis in male offspring through PSCA hypomethylation. Toxicol. Lett. 2018, 292, 78–84. [Google Scholar] [CrossRef]
- Sarath Josh, M.K.; Pradeep, S.; Adarsh, V.K.; Vijayalekshmi Amma, K.S.; Sudha Devi, R.; Balachandran, S.; Sreejith, M.N.; Abdul Jaleel, U.C.; Benjamin, S. In silico evidences for the binding of phthalates onto human estrogen receptor α, β subtypes and human estrogen-related receptor γ. Mol. Simul. 2014, 40, 408–417. [Google Scholar] [CrossRef]
- Takeuchi, S.; Iida, M.; Kobayashi, S.; Jin, K.; Matsuda, T.; Kojima, H. Differential effects of phthalate esters on transcriptional activities via human estrogen receptors alpha and beta, and androgen receptor. Toxicology 2005, 210, 223–233. [Google Scholar] [CrossRef]
- Kambia, N.; Séverin, I.; Farce, A.; Dahbi, L.; Dine, T.; Moreau, E.; Sautou, V.; Chagnon, M.C. Comparative Effects of Di-(2-ethylhexyl)phthalate and Di-(2-ethylhexyl)terephthalate Metabolites on Thyroid Receptors: In Vitro and In Silico Studies. Metabolites 2021, 11, 94. [Google Scholar] [CrossRef]
- Sarath Josh, M.K.; Pradeep, S.; Vijayalekshmi Amma, K.S.; Balachandran, S.; Abdul Jaleel, U.C.; Doble, M.; Spener, F.; Benjamin, S. Phthalates efficiently bind to human peroxisome proliferator activated receptor and retinoid X receptor α, β, γ subtypes: An in silico approach. J. Appl. Toxicol. 2014, 34, 754–765. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef] [PubMed]
- Piché, C.D.; Sauvageau, D.; Vanlian, M.; Erythropel, H.C.; Robaire, B.; Leask, R.L. Effects of di-(2-ethylhexyl) phthalate and four of its metabolites on steroidogenesis in MA-10 cells. Ecotoxicol. Environ. Saf. 2012, 79, 108–115. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, L.; Wei, L.; Li, L. DEHP reduces thyroid hormones via interacting with hormone synthesis-related proteins, deiodinases, transthyretin, receptors, and hepatic enzymes in rats. Environ. Sci. Pollut. Res. Int. 2015, 22, 12711–12719. [Google Scholar] [CrossRef]
- Wen, H.J.; Chen, C.C.; Wu, M.T.; Chen, M.L.; Sun, C.W.; Wu, W.C.; Huang, I.W.; Huang, P.C.; Yu, T.Y.; Hsiung, C.A.; et al. Phthalate exposure and reproductive hormones and sex-hormone binding globulin before puberty–Phthalate contaminated-foodstuff episode in Taiwan. PLoS ONE 2017, 12, e0175536. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Wu, S.; Wang, Y.X.; Liu, L.; Zhang, J.; Shen, H.; Lu, Y.; Bao, H.; Huang, Q. Associations of environmental phthalate exposure with male steroid hormone synthesis and metabolism: An integrated epidemiology and toxicology study. J. Hazard. Mater. 2022, 436, 129213. [Google Scholar] [CrossRef] [PubMed]
- Zota, A.R.; Geller, R.J.; VanNoy, B.N.; Marfori, C.Q.; Tabbara, S.; Hu, L.Y.; Baccarelli, A.A.; Moawad, G.N. Phthalate Exposures and MicroRNA Expression in Uterine Fibroids: The FORGE Study. Epigenet Insights 2020, 13, 2516865720904057. [Google Scholar] [CrossRef]
- Amir, S.; Shah, S.T.A.; Mamoulakis, C.; Docea, A.O.; Kalantzi, O.I.; Zachariou, A.; Calina, D.; Carvalho, F.; Sofikitis, N.; Makrigiannakis, A.; et al. Endocrine Disruptors Acting on Estrogen and Androgen Pathways Cause Reproductive Disorders through Multiple Mechanisms: A Review. Int. J. Environ. Res. Public Health 2021, 18, 1464. [Google Scholar] [CrossRef]
- Buñay, J.; Larriba, E.; Moreno, R.D.; Del Mazo, J. Chronic low-dose exposure to a mixture of environmental endocrine disruptors induces microRNAs/isomiRs deregulation in mouse concomitant with intratesticular estradiol reduction. Sci. Rep. 2017, 7, 3373. [Google Scholar] [CrossRef] [PubMed]
- Scarano, W.R.; Bedrat, A.; Alonso-Costa, L.G.; Aquino, A.M.; Fantinatti, B.E.A.; Justulin, L.A.; Barbisan, L.F.; Freire, P.P.; Flaws, J.A.; Lemos, B. Exposure to an Environmentally Relevant Phthalate Mixture During Prostate Development Induces MicroRNA Upregulation and Transcriptome Modulation in Rats. Toxicol. Sci. 2019, 171, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, T.; Tao, Q.; Tan, W.; Chen, H.; Liu, W.; Lin, P.; Zhou, D.; Wang, A.; Jin, Y.; et al. Bta-miR-34b inhibits proliferation and promotes apoptosis via the MEK/ERK pathway by targeting MAP2K1 in bovine primary Sertoli cells. J. Anim. Sci. 2020, 98, skaa313. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wu, J.; Ma, X.; Huang, C.; Wu, R.; Zhu, W.; Li, X.; Liang, Z.; Deng, F.; Zhu, J.; et al. Butyl benzyl phthalate promotes prostate cancer cell proliferation through miR-34a downregulation. Toxicol. Vitr. 2019, 54, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, Y.; Chen, J.; Song, Y.; You, M.; Yang, G. Long-term co-exposure DBP and BaP causes imbalance in liver macrophages polarization via activation of Notch signaling regulated by miR-34a-5p in rats. Chem. Biol. Interact. 2022, 359, 109919. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Varol, M.; Tuorkey, M.; Sak, K.; Parashar, N.C.; Barwal, T.S.; Sharma, U.; Iqubal, A.; Parashar, G.; et al. NOTCH signaling: Journey of an evolutionarily conserved pathway in driving tumor progression and its modulation as a therapeutic target. Crit. Rev. Oncol. Hematol. 2021, 164, 103403. [Google Scholar] [CrossRef]
- Ferrante, M.; Cristaldi, A.; Oliveri Conti, G. Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review. J. Pers. Med. 2021, 11, 500. [Google Scholar] [CrossRef]
- Mohammadi, H.; Ashari, S. Mechanistic insight into toxicity of phthalates, the involved receptors, and the role of Nrf2, NF-κB, and PI3K/AKT signaling pathways. Environ. Sci. Pollut. Res. Int. 2021, 28, 35488–35527. [Google Scholar] [CrossRef]
- Basso, C.G.; de Araújo-Ramos, A.T.; Martino-Andrade, A.J. Exposure to phthalates and female reproductive health: A literature review. Reprod. Toxicol. 2022, 109, 61–79. [Google Scholar] [CrossRef]
- Egusquiza, R.J.; Blumberg, B. Environmental Obesogens and Their Impact on Susceptibility to Obesity: New Mechanisms and Chemicals. Endocrinology 2020, 161, bqaa024. [Google Scholar] [CrossRef]
- Meruvu, S.; Zhang, J.; Choudhury, M. Butyl Benzyl Phthalate Promotes Adipogenesis in 3T3-L1 Cells via the miRNA-34a-5p Signaling Pathway in the Absence of Exogenous Adipogenic Stimuli. Chem. Res. Toxicol. 2021, 34, 2251–2260. [Google Scholar] [CrossRef]
- Gharanei, S.; Shabir, K.; Brown, J.E.; Weickert, M.O.; Barber, T.M.; Kyrou, I.; Randeva, H.S. Regulatory microRNAs in Brown, Brite and White Adipose Tissue. Cells 2020, 9, 2489. [Google Scholar] [CrossRef]
- Schug, T.T.; Li, X. Sirtuin 1 in lipid metabolism and obesity. Ann. Med. 2011, 43, 198–211. [Google Scholar] [CrossRef]
- Nielsen, K.N.; Peics, J.; Ma, T.; Karavaeva, I.; Dall, M.; Chubanava, S.; Basse, A.L.; Dmytriyeva, O.; Treebak, J.T.; Gerhart-Hines, Z. NAMPT-mediated NAD+ biosynthesis is indispensable for adipose tissue plasticity and development of obesity. Mol. Metab. 2018, 11, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Yang, F.S.; Sivasankaran, V.P.; Lo, Y.L.; Wu, Y.T.; Chang, C.Y.; Chiu, C.C.; Liao, Z.X.; Wang, L.F. Comparing the Variants of Iron Oxide Nanoparticle-Mediated Delivery of miRNA34a for Efficiency in Silencing of PD-L1 Genes in Cancer Cells. Pharmaceutics 2023, 15, 215. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhao, H.; Jing, S.; Fan, Y.; Sheng, G.; Ding, Q.; Liu, C.; Wu, H.; Liu, Y. Magnetofection of miR-21 promoted by electromagnetic field and iron oxide nanoparticles via the p38 MAPK pathway contributes to osteogenesis and angiogenesis for intervertebral fusion. J. Nanobiotechnol. 2023, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Schladt, T.D.; Schneider, K.; Schild, H.; Tremel, W. Synthesis and bio-functionalization of magnetic nanoparticles for medical diagnosis and treatment. Dalton Trans. 2011, 40, 6315–6343. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lin, L.; Li, L.; Zhou, L.; Zhang, Y.; Hao, S.; Ding, Z. Exosomal small RNA sequencing uncovers the microRNA dose markers for power frequency electromagnetic field exposure. Biomarkers 2018, 23, 315–327. [Google Scholar] [CrossRef]
- Lamkowski, A.; Kreitlow, M.; Radunz, J.; Willenbockel, M.; Stiemer, M.; Fichte, L.O.; Rädel, C.F.; Majewski, M.; Ostheim, P.; Port, M.; et al. Analyzing the impact of 900 MHz EMF short-term exposure to the expression of 667 miRNAs in human peripheral blood cells. Sci. Rep. 2021, 11, 4444. [Google Scholar] [CrossRef] [PubMed]
- Aalami Zavareh, F.; Abdi, S.; Entezari, M. Up-regulation of miR-144 and miR-375 in the human gastric cancer cell line following the exposure to extremely low-frequency electromagnetic fields. Int. J. Radiat. Biol. 2021, 97, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Consales, C.; Cirotti, C.; Filomeni, G.; Panatta, M.; Butera, A.; Merla, C.; Lopresto, V.; Pinto, R.; Marino, C.; Benassi, B. Fifty-Hertz Magnetic Field Affects the Epigenetic Modulation of the miR-34b/c in Neuronal Cells. Mol. Neurobiol. 2018, 55, 5698–5714. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Meng, X.; Wu, J.; Pan, C.; Ying, X.; Zhou, Y.; Liu, R.; Huang, W. Cryptochrome 1 overexpression correlates with tumor progression and poor prognosis in patients with colorectal cancer. PLoS ONE 2013, 8, e61679. [Google Scholar] [CrossRef]
- Mazzoccoli, G.; Colangelo, T.; Panza, A.; Rubino, R.; De Cata, A.; Tiberio, C.; Valvano, M.R.; Pazienza, V.; Merla, G.; Augello, B.; et al. Deregulated expression of cryptochrome genes in human colorectal cancer. Mol. Cancer 2016, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Honma, S. The mammalian circadian system: A hierarchical multi-oscillator structure for generating circadian rhythm. J. Physiol. Sci. 2018, 68, 207–219. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Pilorz, V.; Astiz, M.; Heinen, K.O.; Rawashdeh, O.; Oster, H. The Concept of Coupling in the Mammalian Circadian Clock Network. J. Mol. Biol. 2020, 432, 3618–3638. [Google Scholar] [CrossRef]
- Lee, Y. Roles of circadian clocks in cancer pathogenesis and treatment. Exp. Mol. Med. 2021, 53, 1529–1538. [Google Scholar] [CrossRef]
- Anna, G.; Kannan, N.N. Post-transcriptional modulators and mediators of the circadian clock. Chronobiol. Int. 2021, 38, 1244–1261. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lazar, M.A. Transcriptional Control of Circadian Rhythms and Metabolism: A Matter of Time and Space. Endocr. Rev. 2020, 41, 707–732. [Google Scholar] [CrossRef]
- Mehta, N.; Cheng, H.Y. Micro-managing the circadian clock: The role of microRNAs in biological timekeeping. J. Mol. Biol. 2013, 425, 3609–3624. [Google Scholar] [CrossRef]
- Kinoshita, C.; Aoyama, K.; Nakaki, T. Neuroprotection afforded by circadian regulation of intracellular glutathione levels: A key role for miRNAs. Free Radic. Biol. Med. 2018, 119, 17–33. [Google Scholar] [CrossRef]
- Ma, Q.; Mo, G.; Tan, Y. Micro RNAs and the biological clock: A target for diseases associated with a loss of circadian regulation. Afr. Health Sci. 2020, 20, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Ray, I.; Goswami, S. Circadian rhythm genes in cancer: Insight into their functions and regulation involving noncoding RNAs. Chronobiol. Int. 2021, 38, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.Y.; Papp, J.W.; Varlamova, O.; Dziema, H.; Russell, B.; Curfman, J.P.; Nakazawa, T.; Shimizu, K.; Okamura, H.; Impey, S.; et al. microRNA modulation of circadian-clock period and entrainment. Neuron 2007, 54, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; D’Alessandro, M.; Lee, C. miRNAs are required for generating a time delay critical for the circadian oscillator. Curr. Biol. 2013, 23, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.H.; Kojima, S.; Shimomura, K.; Koike, N.; Buhr, E.D.; Furukawa, T.; Ko, C.H.; Gloston, G.; Ayoub, C.; Nohara, K.; et al. Period2 3’-UTR and microRNA-24 regulate circadian rhythms by repressing PERIOD2 protein accumulation. Proc. Natl. Acad. Sci. USA 2017, 114, E8855–E8864. [Google Scholar] [CrossRef] [PubMed]
- Pidíková, P.; Chovancová, B.; Mravec, B.; Herichová, I. The 24-h pattern of dgcr8, drosha, and dicer expression in the rat suprachiasmatic nuclei and peripheral tissues and its modulation by angiotensin II. Gen. Physiol. Biophys. 2022, 41, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Hasakova, K.; Reis, R.; Vician, M.; Zeman, M.; Herichova, I. Expression of miR-34a-5p is up-regulated in human colorectal cancer and correlates with survival and clock gene PER2 expression. PLoS ONE 2019, 14, e0224396. [Google Scholar] [CrossRef]
- Han, Y.; Meng, F.; Venter, J.; Wu, N.; Wan, Y.; Standeford, H.; Francis, H.; Meininger, C.; Greene, J., Jr.; Trzeciakowski, J.P.; et al. miR-34a-dependent overexpression of Per1 decreases cholangiocarcinoma growth. J. Hepatol. 2016, 64, 1295–1304. [Google Scholar] [CrossRef]
- Nagel, R.; Clijsters, L.; Agami, R. The miRNA-192/194 cluster regulates the Period gene family and the circadian clock. FEBS J. 2009, 276, 5447–5455. [Google Scholar] [CrossRef]
- Wang, H.; Fan, Z.; Zhao, M.; Li, J.; Lu, M.; Liu, W.; Ying, H.; Liu, M.; Yan, J. Oscillating primary transcripts harbor miRNAs with circadian functions. Sci. Rep. 2016, 6, 21598. [Google Scholar] [CrossRef]
- Hansen, K.F.; Sakamoto, K.; Obrietan, K. MicroRNAs: A potential interface between the circadian clock and human health. Genome Med. 2011, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Zhang, P.; Zhou, L.; Yin, B.; Pan, H.; Peng, X. Clock-controlled mir-142-3p can target its activator, Bmal1. BMC Mol. Biol. 2012, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Shende, V.R.; Neuendorff, N.; Earnest, D.J. Role of miR-142-3p in the post-transcriptional regulation of the clock gene Bmal1 in the mouse SCN. PLoS ONE 2013, 8, e65300. [Google Scholar] [CrossRef] [PubMed]
- Gatfield, D.; Le Martelot, G.; Vejnar, C.E.; Gerlach, D.; Schaad, O.; Fleury-Olela, F.; Ruskeepää, A.L.; Oresic, M.; Esau, C.C.; Zdobnov, E.M.; et al. Integration of microRNA miR-122 in hepatic circadian gene expression. Genes. Dev. 2009, 23, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Viveros, L.; Bouchard-Cannon, P.; Hegazi, S.; Cheng, A.H.; Pastore, S.; Cheng, H.M. Molecular modulators of the circadian clock: Lessons from flies and mice. Cell Mol. Life Sci. 2017, 74, 1035–1059. [Google Scholar] [CrossRef]
- Herichová, I.; Tesáková, B.; Kršková, L.; Olexová, L. Food reward induction of rhythmic clock gene expression in the prefrontal cortex of rats is accompanied by changes in miR-34a-5p expression. Eur. J. Neurosci. 2021, 54, 7476–7492. [Google Scholar] [CrossRef] [PubMed]
- Gantier, M.P.; McCoy, C.E.; Rusinova, I.; Saulep, D.; Wang, D.; Xu, D.; Irving, A.T.; Behlke, M.A.; Hertzog, P.J.; Mackay, F.; et al. Analysis of microRNA turnover in mammalian cells following Dicer1 ablation. Nucleic Acids Res. 2011, 39, 5692–5703. [Google Scholar] [CrossRef]
- Herichova, I. Changes of physiological functions induced by shift work. Endocr. Regul. 2013, 47, 159–170. [Google Scholar] [CrossRef]
- Sheikh-Ali, M.; Maharaj, J. Circadian clock desynchronisation and metabolic syndrome. Postgrad. Med. J. 2014, 90, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Lunn, R.M.; Blask, D.E.; Coogan, A.N.; Figueiro, M.G.; Gorman, M.R.; Hall, J.E.; Hansen, J.; Nelson, R.J.; Panda, S.; Smolensky, M.H.; et al. Health consequences of electric lighting practices in the modern world: A report on the National Toxicology Program’s workshop on shift work at night, artificial light at night, and circadian disruption. Sci. Total Environ. 2017, 607–608, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Okuliarova, M.; Rumanova, V.S.; Stebelova, K.; Zeman, M. Dim Light at Night Disturbs Molecular Pathways of Lipid Metabolism. Int. J. Mol. Sci. 2020, 18, 6919. [Google Scholar] [CrossRef] [PubMed]
- Okuliarova, M.; Mazgutova, N.; Majzunova, M.; Rumanova, V.S.; Zeman, M. Dim Light at Night Impairs Daily Variation of Circulating Immune Cells and Renal Immune Homeostasis. Front Immunol. 2021, 11, 614960. [Google Scholar] [CrossRef] [PubMed]
- Modi, P.K.; Jaiswal, S.; Sharma, P. Regulation of Neuronal Cell Cycle and Apoptosis by MicroRNA 34a. Mol. Cell. Biol. 2015, 36, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Chua, C.E.L.; Tang, B.L. miR-34a in Neurophysiology and Neuropathology. J. Mol. Neurosci. 2019, 67, 235–246. [Google Scholar] [CrossRef]
- Namgyal, D.; Chandan, K.; Sultan, A.; Aftab, M.; Ali, S.; Mehta, R.; El-Serehy, H.A.; Al-Misned, F.A.; Sarwat, M. Dim Light at Night Induced Neurodegeneration and Ameliorative Effect of Curcumin. Cells 2020, 9, 2093. [Google Scholar] [CrossRef]
- Andolina, D.; Di Segni, M.; Bisicchia, E.; D’Alessandro, F.; Cestari, V.; Ventura, A.; Concepcion, C.; Puglisi-Allegra, S.; Ventura, R. Effects of lack of microRNA-34 on the neural circuitry underlying the stress response and anxiety. Neuropharmacology 2016, 107, 305–316. [Google Scholar] [CrossRef]
- Mi, H.; Thomas, P. PANTHER Pathway: An Ontology-Based Pathway Database Coupled with Data Analysis Tools. In Protein Networks and Pathway Analysis; Nikolsky, Y., Bryant, J., Eds.; Methods in Molecular Biology Series; Humana Press: Totowa, NJ, USA, 2009; Volume 563. [Google Scholar]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef]
- Liu, K.; Lehmann, K.P.; Sar, M.; Young, S.S.; Gaido, K.W. Gene expression profiling following in utero exposure to phthalate esters reveals new gene targets in the etiology of testicular dysgenesis. Biol. Reprod. 2005, 73, 180–192. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim Ey Ryu, J.C. Identification of estrogenic genes responding to phthalate esters treatment in human MCF-7 cells. Mol. Cell. Toxicol. 2011, 7, 163–170. [Google Scholar] [CrossRef]
- Gopalakrishnan, K.; Aushev, V.N.; Manservisi, F.; Falcioni, L.; Panzacchi, S.; Belpoggi, F.; Parada, H., Jr.; Garbowski, G.; Hibshoosh, H.; Santella, R.M.; et al. Gene expression profiles for low-dose exposure to diethyl phthalate in rodents and humans: A translational study with implications for breast carcinogenesis. Sci. Rep. 2020, 10, 7067. [Google Scholar] [CrossRef]
- Remondini, D.; Nylund, R.; Reivinen, J.; Poulletier de Gannes, F.; Veyret, B.; Lagroye, I.; Haro, E.; Trillo, M.A.; Capri, M.; Franceschi, C.; et al. Gene expression changes in human cells after exposure to mobile phone microwaves. Proteomics 2006, 6, 4745–4754. [Google Scholar] [CrossRef]
- Huang, T.Q.; Lee, M.S.; Oh, E.H.; Kalinec, F.; Zhang, B.T.; Seo, J.S.; Park, W.Y. Characterization of biological effect of 1763 MHz radiofrequency exposure on auditory hair cells. Int. J. Radiat. Biol. 2008, 84, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Regalbuto, E.; Anselmo, A.; De Sanctis, S.; Franchini, V.; Lista, F.; Benvenuto, M.; Bei, R.; Masuelli, L.; D’Inzeo, G.; Paffi, A.; et al. Human Fibroblasts In Vitro Exposed to 2.45 GHz Continuous and Pulsed Wave Signals: Evaluation of Biological Effects with a Multimethodological Approach. Int. J. Mol. Sci. 2020, 21, 7069. [Google Scholar] [CrossRef] [PubMed]
- Kervezee, L.; Cuesta, M.; Cermakian, N.; Boivin, D.B. The Phase-Shifting Effect of Bright Light Exposure on Circadian Rhythmicity in the Human Transcriptome. J. Biol. Rhythms 2019, 34, 84–97. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Stress Factor | miRNA | Change in Expression | Tissue/Cell Line (Species) | Reference |
|---|---|---|---|---|
| Phtalates | pre-miR-34b miR-34b-5p | up | Testes (mouse) | [73] |
| miR-34a-5p | up | 3T3-L1 cells (mouse) | [83] | |
| miR-34a | down | PC-3 cells, LNCaP cells (human) | [76] | |
| miR-34a-5p | down | Female/CA3 and DG in Hippocampus (rat) | [56] | |
| miR-34a-5p | down | Liver (rat) | [77] | |
| EMF | miR-34b/c | down | SH-SY5Y cells (human) PCN cells (mouse) | [94] |
| circadian cycle deregulation/ cancer | pre-miR-34a | up | Kidney, Heart (rat) | [110] |
| miR-34a-5p | up | Colorectal cancer tissue (human) | [111] | |
| miR-34a-5p | aberrant | Cholangeocarcinoma cells (human) | [112] | |
| circadian cycle deregulation/neurodegeneration | miR-34a | down | Hippocampus (mouse) | [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štefánik, P.; Morová, M.; Herichová, I. Impact of Long-Lasting Environmental Factors on Regulation Mediated by the miR-34 Family. Biomedicines 2024, 12, 424. https://doi.org/10.3390/biomedicines12020424
Štefánik P, Morová M, Herichová I. Impact of Long-Lasting Environmental Factors on Regulation Mediated by the miR-34 Family. Biomedicines. 2024; 12(2):424. https://doi.org/10.3390/biomedicines12020424
Chicago/Turabian StyleŠtefánik, Peter, Martina Morová, and Iveta Herichová. 2024. "Impact of Long-Lasting Environmental Factors on Regulation Mediated by the miR-34 Family" Biomedicines 12, no. 2: 424. https://doi.org/10.3390/biomedicines12020424
APA StyleŠtefánik, P., Morová, M., & Herichová, I. (2024). Impact of Long-Lasting Environmental Factors on Regulation Mediated by the miR-34 Family. Biomedicines, 12(2), 424. https://doi.org/10.3390/biomedicines12020424