
Bile Acid Sequestration via Colesevelam Reduces Bile Acid Hydrophobicity and Improves Liver Pathology in Cyp2c70−/− Mice with a Human-like Bile Acid Composition

 , , and
, , and 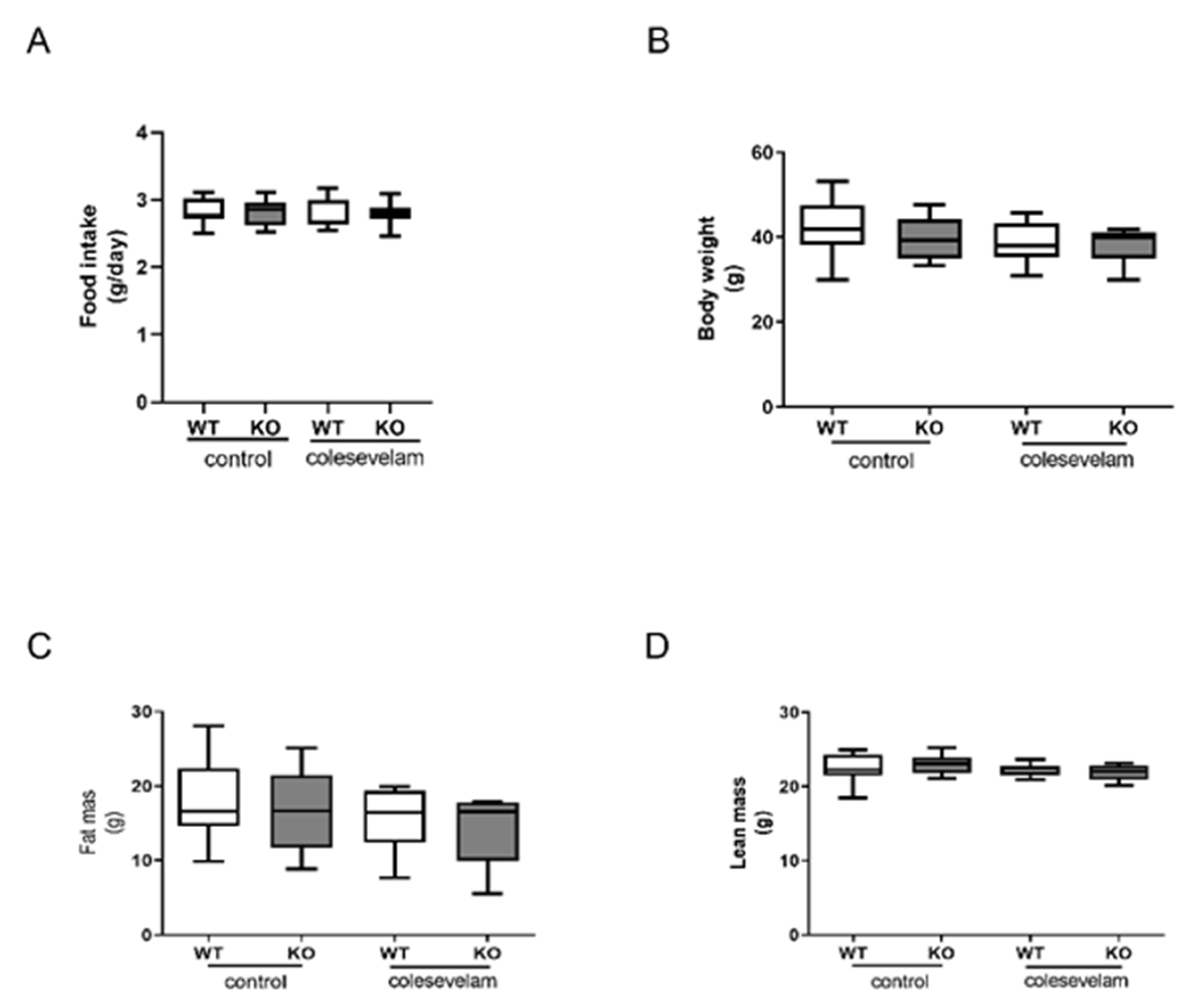{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Body Weight, Food Intake and Body Composition Analysis
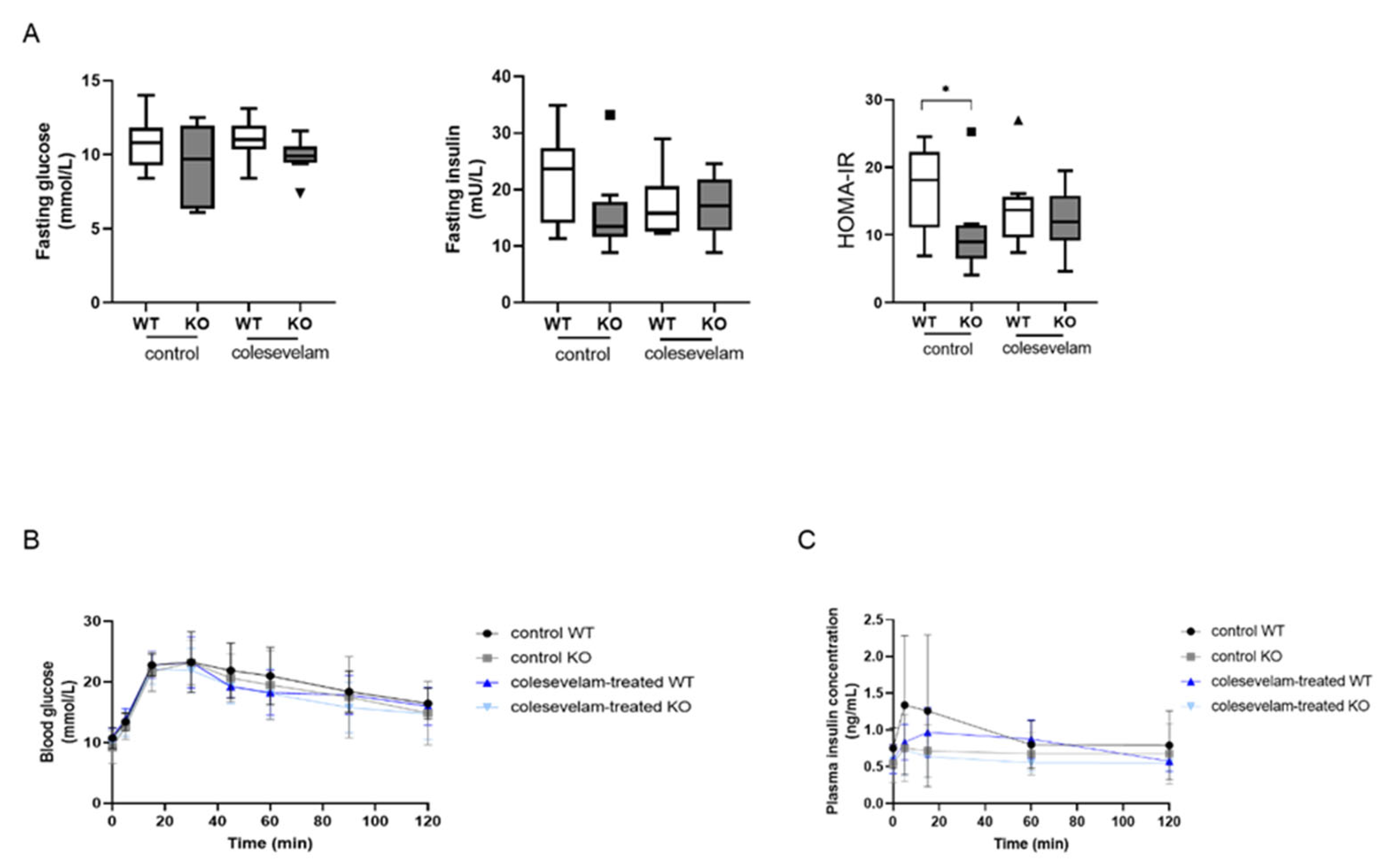
2.3. Assessment of Glucose Tolerance and Insulin Resistance
2.4. Plasma Biochemistry
2.5. Bile Acid Measurements
2.6. Hepatic Lipid Analyses
2.7. Fecal Neutral Sterol Analyses
2.8. Determination of mRNA Levels
2.9. Histology and Staining of Liver
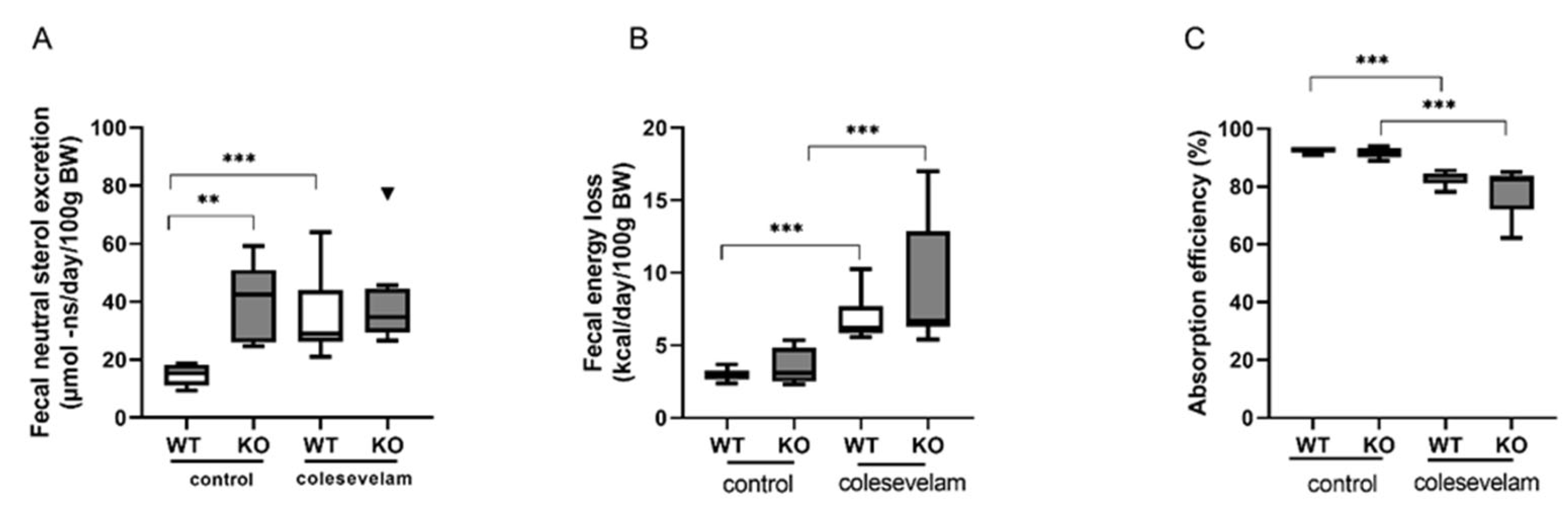
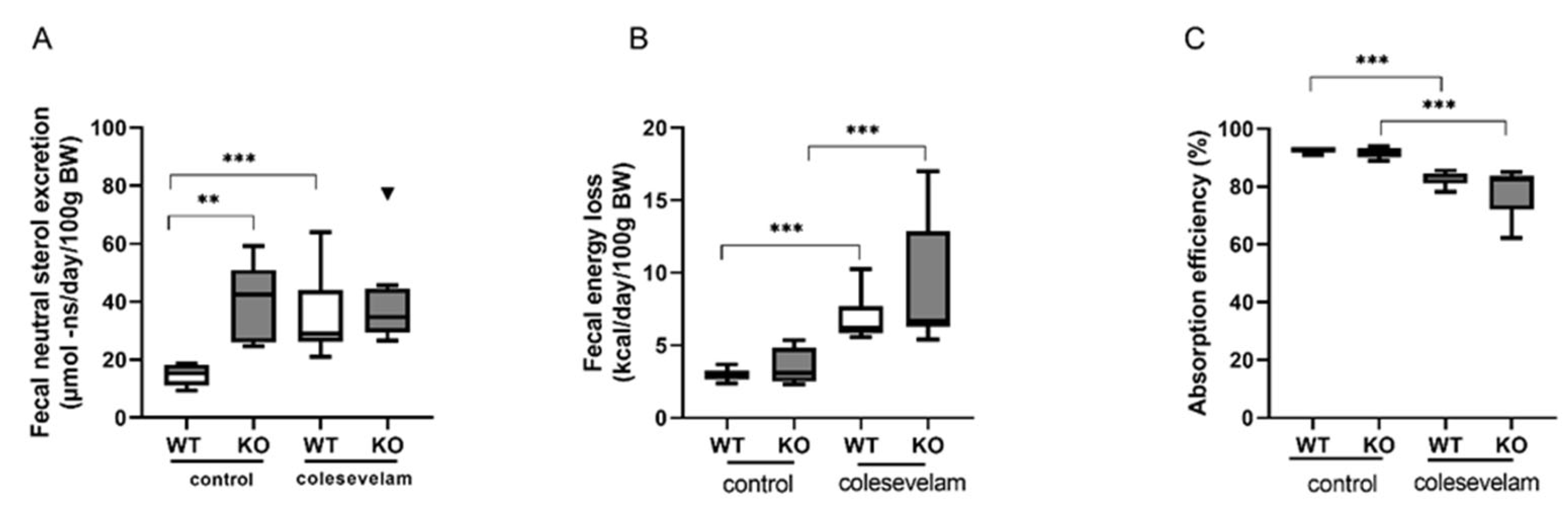
2.10. Determination of Fecal Energy Content and Energy Absorption
2.11. Statistical Analysis
3. Results
3.1. Colesevelam Does Not Affect the Body Weight of WTD-Fed Cyp2c70−/− Mice
3.2. Colesevelam Reduces the Hydrophobicity of the Bile Acid Pool in WTD-Fed Cyp2c70−/− Mice
3.3. Colesevelam Differentially Modulates WTD-Induced Hepatic Steatosis in WT and Cyp2c70−/− Mice
3.4. Colesevelam Ameliorates Liver Damage in WTD-Fed Cyp2c70−/− Mice
3.5. Colesevelam Modulates the Absorption Efficiency of Dietary Energy in WTD-Fed WT and Cyp2c70−/− Mice
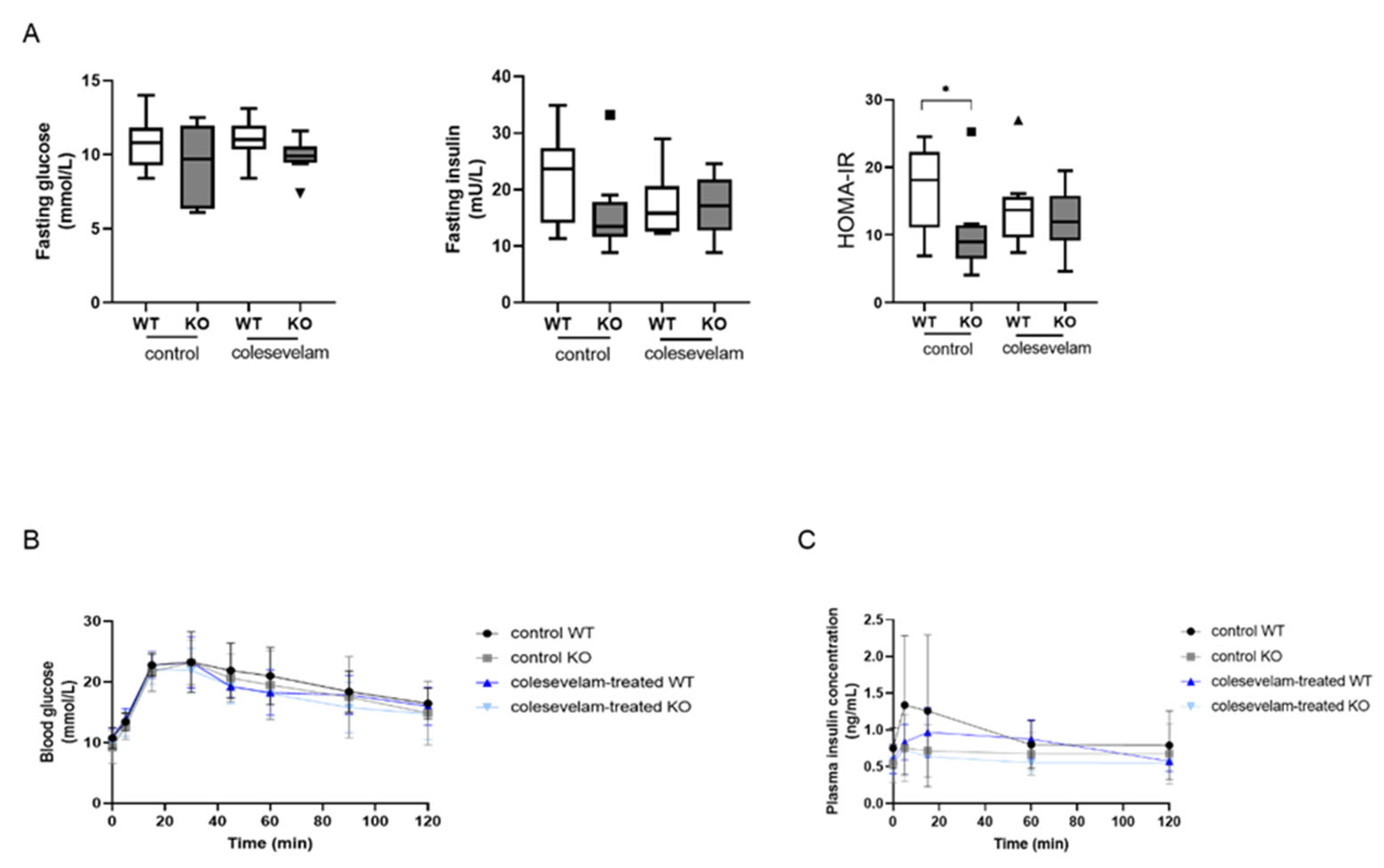
3.6. Colesevelam Does Not Affect Glucose Excursions and Insulin Sensitivity in WTD-Fed Cyp2c70−/− Mice upon 2 Weeks of Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, T.; Chiang, J.Y. Bile acids as metabolic regulators: An update. Curr. Opin. Gastroenterol. 2023, 39, 249–255. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acids as Metabolic Regulators and Nutrient Sensors. Annu. Rev. Nutr. 2019, 39, 175–200. [Google Scholar] [CrossRef]
- Floreani, A.; Mangini, C. Primary biliary cholangitis: Old and novel therapy. Eur. J. Intern. Med. 2018, 47, 20. [Google Scholar] [CrossRef]
- Verbeke, L.; Mannaerts, I.; Schierwagen, R.; Govaere, O.; Klein, S.; Elst, I.V.; Windmolders, P.; Farre, R.; Wenes, M.; Mazzone, M.; et al. FXR agonist obeticholic acid reduces hepatic inflammation and fibrosis in a rat model of toxic cirrhosis. Sci. Rep. 2016, 6, 33453. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-Y.; Ding, W.; Zhang, C.; Fu, L.; Xu, D.-X.; Chen, X. Obeticholic acid prevents carbon tetrachloride-induced liver fibrosis through interaction between farnesoid X receptor and Smad3. Int. Immunopharmacol. 2019, 77, 105911. [Google Scholar] [CrossRef]
- Kowdley, K.V.; Vuppalanchi, R.; Levy, C.; Floreani, A.; Andreone, P.; LaRusso, N.F.; Shrestha, R.; Trotter, J.; Goldberg, D.; Rushbrook, S.; et al. A randomized, placebo-controlled, phase II study of obeticholic acid for primary sclerosing cholangitis. J. Hepatol. 2020, 73, 94–101. [Google Scholar] [CrossRef]
- Nevens, F.; Andreone, P.; Mazzella, G.; Strasser, S.I.; Bowlus, C.; Invernizzi, P.; Drenth, J.P.; Pockros, P.J.; Regula, J.; Beuers, U.; et al. A Placebo-Controlled Trial of Obeticholic Acid in Primary Biliary Cholangitis. N. Engl. J. Med. 2016, 375, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Manne, V.; Kowdley, K.V. Obeticholic acid in primary biliary cholangitis: Where we stand. Curr. Opin. Gastroenterol. 2019, 35, 191–196. [Google Scholar] [CrossRef]
- Bowlus, C.L. Obeticholic acid for the treatment of primary biliary cholangitis in adult patients: Clinical utility and patient selection. Hepatic Med. Évid. Res. 2016, 8, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Kuipers, F.; de Boer, J.F.; Kuivenhoven, J.A. Modulation of Bile Acid Metabolism to Improve Plasma Lipid and Lipoprotein Profiles. J. Clin. Med. 2022, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.D.; Paumgartner, G.; Mlitz, V.; Kunczer, V.; Halilbasic, E.; Leditznig, N.; Wahlström, A.; Ståhlman, M.; Thüringer, A.; Kashofer, K.; et al. Colesevelam attenuates cholestatic liver and bile duct injury in Mdr2−/− mice by modulating composition, signalling and excretion of faecal bile acids. Gut 2018, 67, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Baghdasaryan, A.; Fuchs, C.D.; Österreicher, C.H.; Lemberger, U.J.; Halilbasic, E.; Påhlman, I.; Graffner, H.; Krones, E.; Fickert, P.; Wahlström, A.; et al. Inhibition of intestinal bile acid absorption improves cholestatic liver and bile duct injury in a mouse model of sclerosing cholangitis. J. Hepatol. 2016, 64, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E.; Goldberg, R.B.; Truitt, K.E.; Jones, M.R. Colesevelam Hydrochloride Therapy in Patients with Type 2 Diabetes Mellitus Treated with Metformin Glucose and Lipid Effects. Available online: https://jamanetwork.com/ (accessed on 13 October 2008).
- Zieve, F.J.; Kalin, M.F.; Schwartz, S.L.; Jones, M.R.; Bailey, W.L. Results of the glucose-lowering effect of WelChol study (GLOWS): A randomized, double-blind, placebo-controlled pilot study evaluating the effect of colesevelam hydrochloride on glycemic control in subjects with type 2 diabetes. Clin. Ther. 2007, 29, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Brufau, G.; Stellaard, F.; Prado, K.; Bloks, V.W.; Jonkers, E.; Boverhof, R.; Kuipers, F.; Murphy, E.J. Improved glycemic control with colesevelam treatment in patients with type 2 diabetes is not directly associated with changes in bile acid metabolism. Hepatology 2010, 52, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Meissner, M.; Herrema, H.; van Dijk, T.H.; Gerding, A.; Havinga, R.; Boer, T.; Müller, M.; Reijngoud, D.-J.; Groen, A.K.; Kuipers, F. Bile Acid Sequestration Reduces Plasma Glucose Levels in db/db Mice by Increasing Its Metabolic Clearance Rate. PLoS ONE 2011, 6, e24564. [Google Scholar] [CrossRef] [PubMed]
- Beil, U.; Crouse, J.R.; Einarsson, K.; Grundy, S.M. Effects of interruption of the enterohepatic circulation of bile acids on the transport of very low density-lipoprotein triglycerides. Metabolism 1982, 31, 438–444. [Google Scholar] [CrossRef]
- Crouse, J.R. Hypertriglyceridemia: A contraindication to the use of bile acid binding resins. Am. J. Med. 1987, 83, 243–248. [Google Scholar] [CrossRef]
- de Boer, J.F.; de Vries, H.D.; Palmiotti, A.; Li, R.; Doestzada, M.; Hoogerland, J.A.; Fu, J.; La Rose, A.M.; Westerterp, M.; Mulder, N.L.; et al. Cholangiopathy and Biliary Fibrosis in Cyp2c70-Deficient Mice Are Fully Reversed by Ursodeoxycholic Acid. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1045–1069. [Google Scholar] [CrossRef]
- Honda, A.; Miyazaki, T.; Iwamoto, J.; Hirayama, T.; Morishita, Y.; Monma, T.; Ueda, H.; Mizuno, S.; Sugiyama, F.; Takahashi, S.; et al. Regulation of bile acid metabolism in mouse models with hydrophobic bile acid composition. J. Lipid Res. 2020, 61, 54–69. [Google Scholar] [CrossRef]
- Straniero, S.; Laskar, A.; Savva, C.; HÄrdfeldt, J.; Angelin, B.; Rudling, M. Of mice and men: Murine bile acids explain species differences in the regulation of bile acid and cholesterol metabolism. J. Lipid Res. 2020, 61, 480–491. [Google Scholar] [CrossRef]
- Li, R.; Palmiotti, A.; de Vries, H.D.; Hovingh, M.V.; Koehorst, M.; Mulder, N.L.; Zhang, Y.; Kats, K.; Bloks, V.W.; Fu, J.; et al. Low production of 12α-hydroxylated bile acids prevents hepatic steatosis in Cyp2c70−/− mice by reducing fat absorption. J. Lipid Res. 2021, 62, 100134. [Google Scholar] [CrossRef] [PubMed]
- Dommerholt, M.B.; Blankestijn, M.; Vieira-Lara, M.A.; van Dijk, T.H.; Wolters, H.; Koster, M.H.; Gerding, A.; van Os, R.P.; Bloks, V.W.; Bakker, B.M.; et al. Short-term protein restriction at advanced age stimulates FGF21 signalling, energy expenditure and browning of white adipose tissue. FEBS J. 2021, 288, 2257–2277. [Google Scholar] [CrossRef] [PubMed]
- Eggink, H.M.; Tambyrajah, L.L.; Berg, R.v.D.; Mol, I.M.; Heuvel, J.K.v.D.; Koehorst, M.; Groen, A.K.; Boelen, A.; Kalsbeek, A.; A Romijn, J.; et al. Chronic infusion of taurolithocholate into the brain increases fat oxidation in mice. J. Endocrinol. 2018, 236, 85–97. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Stricker, D. BrightStat.com: Free statistics online. Comput. Methods Programs Biomed. 2008, 92, 135–143. [Google Scholar] [CrossRef] [PubMed]
- de Boer, J.F.; Verkade, E.; Mulder, N.L.; de Vries, H.D.; Huijkman, N.; Koehorst, M.; Boer, T.; Wolters, J.C.; Bloks, V.W.; van de Sluis, B.; et al. A human-like bile acid pool induced by deletion of hepatic Cyp2c70 modulates effects of FXR activation in mice. J. Lipid Res. 2020, 61, 291–305. [Google Scholar] [CrossRef]
- Truong, J.K.; Bennett, A.L.; Klindt, C.; Donepudi, A.C.; Malla, S.R.; Pachura, K.J.; Zaufel, A.; Moustafa, T.; Dawson, P.A.; Karpen, S.J. Ileal bile acid transporter inhibition in Cyp2c70 KO mice ameliorates cholestatic liver injury. J. Lipid Res. 2022, 63, 100261. [Google Scholar] [CrossRef]
- Mudaliar, S.; Henry, R.R.; Sanyal, A.J.; Morrow, L.; Marschall, H.; Kipnes, M.; Adorini, L.; Sciacca, C.I.; Clopton, P.; Castelloe, E.; et al. Efficacy and Safety of the Farnesoid X Receptor Agonist Obeticholic Acid in Patients with Type 2 Diabetes and Nonalcoholic Fatty Liver Disease. Gastroenterology 2013, 145, 574–582.e1. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Chèvre, R.; Mayan, D.C.; Corlianò, M.; Cochran, B.J.; Sem, K.P.; van Dijk, T.H.; Peng, J.; Tan, L.J.; Hartimath, S.V.; et al. Haploinsufficiency of CYP8B1 associates with increased insulin sensitivity in humans. J. Clin. Investig. 2022, 132, e152961. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmiotti, A.; de Vries, H.D.; Hovingh, M.V.; Koehorst, M.; Mulder, N.L.; Verkade, E.; Veentjer, M.K.; van Dijk, T.H.; Bloks, V.W.; Havinga, R.; et al. Bile Acid Sequestration via Colesevelam Reduces Bile Acid Hydrophobicity and Improves Liver Pathology in Cyp2c70−/− Mice with a Human-like Bile Acid Composition. Biomedicines 2023, 11, 2495. https://doi.org/10.3390/biomedicines11092495
Palmiotti A, de Vries HD, Hovingh MV, Koehorst M, Mulder NL, Verkade E, Veentjer MK, van Dijk TH, Bloks VW, Havinga R, et al. Bile Acid Sequestration via Colesevelam Reduces Bile Acid Hydrophobicity and Improves Liver Pathology in Cyp2c70−/− Mice with a Human-like Bile Acid Composition. Biomedicines. 2023; 11(9):2495. https://doi.org/10.3390/biomedicines11092495
Chicago/Turabian StylePalmiotti, Anna, Hilde D. de Vries, Milaine V. Hovingh, Martijn Koehorst, Niels L. Mulder, Esther Verkade, Melany K. Veentjer, Theo H. van Dijk, Vincent W. Bloks, Rick Havinga, and et al. 2023. "Bile Acid Sequestration via Colesevelam Reduces Bile Acid Hydrophobicity and Improves Liver Pathology in Cyp2c70−/− Mice with a Human-like Bile Acid Composition" Biomedicines 11, no. 9: 2495. https://doi.org/10.3390/biomedicines11092495
APA StylePalmiotti, A., de Vries, H. D., Hovingh, M. V., Koehorst, M., Mulder, N. L., Verkade, E., Veentjer, M. K., van Dijk, T. H., Bloks, V. W., Havinga, R., Verkade, H. J., de Boer, J. F., & Kuipers, F. (2023). Bile Acid Sequestration via Colesevelam Reduces Bile Acid Hydrophobicity and Improves Liver Pathology in Cyp2c70−/− Mice with a Human-like Bile Acid Composition. Biomedicines, 11(9), 2495. https://doi.org/10.3390/biomedicines11092495