
Examining the Interaction between Exercise, Gut Microbiota, and Neurodegeneration: Future Research Directions

 ,
,  ,
,  and
and 
Abstract
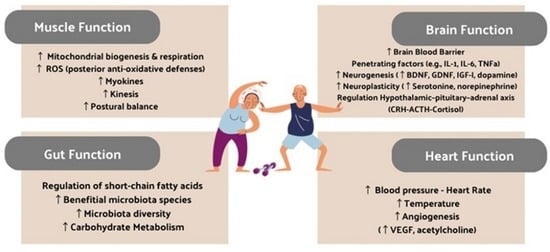
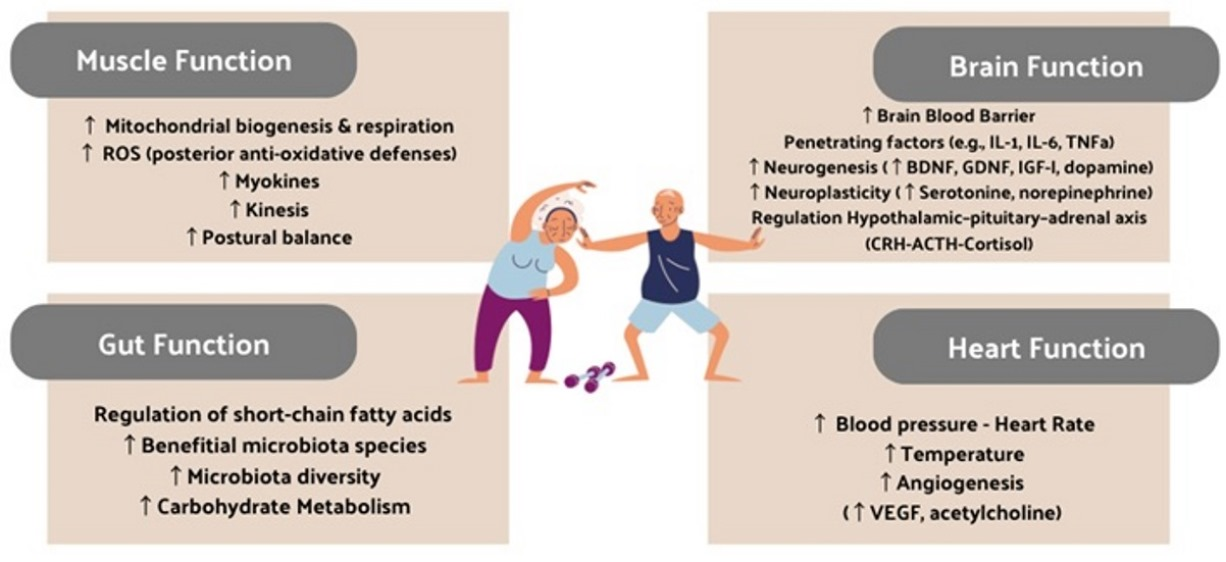
1. Introduction
2. Methods
2.1. Eligibility Criteria
2.2. Information Sources
2.3. Search Strategy
2.4. Findings Presentation
3. Physical Exercise–Gut Health Relationship
4. Physical Exercise as a Neuromodulator
5. The Role of Exercise in Neurodegeneration
6. Complex Interactions between Exercise, Neurodegeneration, and Gut Health
7. Future Research Directions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lanza, I.R.; Short, D.K.; Short, K.R.; Raghavakaimal, S.; Basu, R.; Joyner, M.J.; McConnell, J.P.; Nair, K.S. Endurance Exercise as a Countermeasure for Aging. Diabetes 2008, 57, 2933–2942. [Google Scholar] [CrossRef]
- Mcleod, J.C.; Stokes, T.; Phillips, S.M. Resistance Exercise Training as a Primary Countermeasure to Age-Related Chronic Disease. Front. Physiol. 2019, 10, 645. [Google Scholar] [CrossRef]
- Bell, D.S.H. Changes Seen in Gut Bacteria Content and Distribution with Obesity: Causation or Association? Postgrad. Med. 2015, 127, 863–868. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial Ecology: Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Carbajo-Pescador, S.; Porras, D.; García-Mediavilla, M.V.; Martínez-Flórez, S.; Juarez-Fernández, M.; Cuevas, M.J.; Mauriz, J.L.; González-Gallego, J.; Nistal, E.; Sánchez-Campos, S. Beneficial Effects of Exercise on Gut Microbiota Functionality and Barrier Integrity, and Gut-Liver Crosstalk in an in Vivo Model of Early Obesity and Non-Alcoholic Fatty Liver Disease. Dis. Models Mech. 2019, 12, dmm039206. [Google Scholar] [CrossRef]
- Hintikka, J.E.; Ahtiainen, J.P.; Permi, P.; Jalkanen, S.; Lehtonen, M.; Pekkala, S. Aerobic exercise training and gut microbiome-associated metabolic shifts in women with overweight: A multi-omic study. Sci. Rep. 2023, 13, 11228. [Google Scholar] [CrossRef]
- Aya, V.; Jimenez, P.; Muñoz, E.; Ramírez, J.D. Effects of exercise and physical activity on gut microbiota composition and function in older adults: A systematic review. BMC Geriatr. 2023, 23, 364. [Google Scholar] [CrossRef]
- Clauss, M.; Gérard, P.; Mosca, A.; Leclerc, M. Interplay between Exercise and Gut Microbiome in the Context of Human Health and Performance. Front. Nutr. 2021, 8, 637010. [Google Scholar] [CrossRef]
- Aya, V.; Flórez, A.; Perez, L.; Ramírez, J.D. Association between Physical Activity and Changes in Intestinal Microbiota Composition: A Systematic Review. PLoS ONE 2021, 16, e0247039. [Google Scholar] [CrossRef]
- Nieman, D.C.; Wentz, L.M. The Compelling Link between Physical Activity and the Body’s Defense System. J. Sport Health Sci. 2019, 8, 201–217. [Google Scholar] [CrossRef]
- Keirns, B.H.; Koemel, N.A.; Sciarrillo, C.M.; Anderson, K.L.; Emerson, S.R. Exercise and Intestinal Permeability: Another Form of Exercise-Induced Hormesis? Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G512–G518. [Google Scholar] [CrossRef]
- Camilleri, M. The Leaky Gut: Mechanisms, Measurement and Clinical Implications in Humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Mach, N.; Fuster-Botella, D. Endurance Exercise and Gut Microbiota: A Review. J. Sport Health Sci. 2017, 6, 179–197. [Google Scholar] [CrossRef]
- Mailing, L.J.; Allen, J.M.; Buford, T.W.; Fields, C.J.; Woods, J.A. Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc. Sport Sci. Rev. 2019, 47, 75. [Google Scholar] [CrossRef]
- Bonomini-Gnutzmann, R.; Plaza-Díaz, J.; Jorquera-Aguilera, C.; Rodríguez-Rodríguez, A.; Rodríguez-Rodríguez, F. Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 9518. [Google Scholar] [CrossRef]
- Bonilla, D.A.; Pérez-Idárraga, A.; Odriozola-Martínez, A.; Kreider, R.B. The 4R’s Framework of Nutritional Strategies for Post-Exercise Recovery: A Review with Emphasis on New Generation of Carbohydrates. Int. J. Environ. Res. Public Health 2020, 18, 103. [Google Scholar] [CrossRef]
- van Wijck, K.; Lenaerts, K.; Grootjans, J.; Wijnands, K.A.P.; Poeze, M.; van Loon, L.J.C.; Dejong, C.H.C.; Buurman, W.A. Physiology and Pathophysiology of Splanchnic Hypoperfusion and Intestinal Injury during Exercise: Strategies for Evaluation and Prevention. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G155–G168. [Google Scholar] [CrossRef]
- Wegierska, A.E.; Charitos, I.A.; Topi, S.; Potenza, M.A.; Montagnani, M.; Santacroce, L. The Connection between Physical Exercise and Gut Microbiota: Implications for Competitive Sports Athletes. Sports Med. 2022, 52, 2355–2369. [Google Scholar] [CrossRef]
- Ribeiro, F.M.; Petriz, B.; Marques, G.; Kamilla, L.H.; Franco, O.L. Is There an Exercise-Intensity Threshold Capable of Avoiding the Leaky Gut? Front. Nutr. 2021, 8, 627289. [Google Scholar] [CrossRef]
- Clark, A.; Mach, N. Exercise-Induced Stress Behavior, Gut-Microbiota-Brain Axis and Diet: A Systematic Review for Athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef]
- Zhu, X.; Li, B.; Lou, P.; Dai, T.; Chen, Y.; Zhuge, A.; Yuan, Y.; Li, L. The Relationship between the Gut Microbiome and Neurodegenerative Diseases. Neurosci. Bull. 2021, 37, 1510–1522. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Y.; Wang, Z.; Xie, G.; Liu, M.; Yuan, B.; Chai, H.; Wang, W.; Cheng, P. Implications of Gut Microbiota in Neurodegenerative Diseases. Front. Immunol. 2022, 13, 785644. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Li, B.; He, B.; Li, L.; Nice, E.C.; Zhang, W.; Xu, J. New Insights into the Gut Microbiota in Neurodegenerative Diseases from the Perspective of Redox Homeostasis. Antioxidants 2022, 11, 2287. [Google Scholar] [CrossRef]
- Monda, V.; Villano, I.; Messina, A.; Valenzano, A.; Esposito, T.; Moscatelli, F.; Viggiano, A.; Cibelli, G.; Chieffi, S.; Monda, M.; et al. Exercise Modifies the Gut Microbiota with Positive Health Effects. Oxidative Med. Cell. Longev. 2017, 2017, 3831972. [Google Scholar] [CrossRef]
- Dixon-Woods, M.; Bonas, S.; Booth, A.; Jones, D.R.; Miller, T.; Sutton, A.J.; Shaw, R.L.; Smith, J.A.; Young, B. How Can Systematic Reviews Incorporate Qualitative Research? A Critical Perspective. Qual. Res. 2006, 6, 27–44. [Google Scholar] [CrossRef]
- Popay, J.; Roberts, H.; Sowden, A.; Petticrew, M.; Arai, L.; Rodgers, M.; Britten, N.; Roen, K.; Duffy, S. ESRC Methods Programme; Version 1; b92; ESRC: Swindon, UK, 2006.
- Scheer, V.; Tiller, N.; Doutrleau, S.; Khodaee, M.; Knechtle, B.; Pasternak, A.; Rojas-Valverde, D. Potential Long-Term Health Problems Associated with Ultra-Endurance Running: A Narrative Review. Sport Med. 2021; in press. [Google Scholar]
- Sterling, P. Allostasis: A Model of Predictive Regulation. Physiol. Behav. 2012, 106, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Bobba-Alves, N.; Juster, R.-P.; Picard, M. The Energetic Cost of Allostasis and Allostatic Load. Psychoneuroendocrinology 2022, 146, 105951. [Google Scholar] [CrossRef]
- Parker, H.W.; Abreu, A.M.; Sullivan, M.C.; Vadiveloo, M.K. Allostatic Load and Mortality: A Systematic Review and Meta-Analysis. Am. J. Prev. Med. 2022, 63, 131–140. [Google Scholar] [CrossRef]
- Christensen, D.S.; Zachariae, R.; Amidi, A.; Wu, L.M. Sleep and Allostatic Load: A Systematic Review and Meta-Analysis. Sleep Med. Rev. 2022, 64, 101650. [Google Scholar] [CrossRef]
- Lenart-Bugla, M.; Szcześniak, D.; Bugla, B.; Kowalski, K.; Niwa, S.; Rymaszewska, J.; Misiak, B. The Association between Allostatic Load and Brain: A Systematic Review. Psychoneuroendocrinology 2022, 145, 105917. [Google Scholar] [CrossRef]
- Longpré-Poirier, C.; Dougoud, J.; Jacmin-Park, S.; Moussaoui, F.; Vilme, J.; Desjardins, G.; Cartier, L.; Cipriani, E.; Kerr, P.; Le Page, C.; et al. Sex and Gender and Allostatic Mechanisms of Cardiovascular Risk and Disease. Can. J. Cardiol. 2022, 38, 1812–1827. [Google Scholar] [CrossRef]
- Wang, Q.; Qian, W.; Xu, X.; Bajpai, A.; Guan, K.; Zhang, Z.; Chen, R.; Flamini, V.; Chen, W. Energy-Mediated Machinery Drives Cellular Mechanical Allostasis. Adv. Mater. 2019, 31, e1900453. [Google Scholar] [CrossRef]
- Chow, L.S.; Gerszten, R.E.; Taylor, J.M.; Pedersen, B.K.; van Praag, H.; Trappe, S.; Febbraio, M.A.; Galis, Z.S.; Gao, Y.; Haus, J.M.; et al. Exerkines in Health, Resilience and Disease. Nat. Rev. Endocrinol. 2022, 18, 273–289. [Google Scholar] [CrossRef]
- Hamer, M.; Sabia, S.; Batty, G.D.; Shipley, M.J.; Tabák, A.G.; Singh-Manoux, A.; Kivimaki, M. Physical Activity and Inflammatory Markers over 10 Years Follow up in Men and Women from the Whitehall II Cohort Study. Circulation 2012, 126, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Safdar, A.; Saleem, A.; Tarnopolsky, M.A. The potential of endurance exercise-derived exosomes to treat metabolic diseases. Nat. Rev. Endocrinol. 2016, 12, 504–517. [Google Scholar] [CrossRef]
- Sanford, J.A.; Nogiec, C.D.; Lindholm, M.E.; Adkins, J.N.; Amar, D.; Dasari, S.; Drugan, J.K.; Fernández, F.M.; Radom-Aizik, S.; Schenk, S.; et al. Molecular Transducers of Physical Activity Consortium (MoTrPAC): Mapping the dynamic responses to exercise. Cell 2020, 181, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- National Heart, Lung, and Blood Institute. NHLBI and NIDDK Workshop: Exerkines in Health, Resilience, and Diseases Executive Summary. 2020. Available online: https://www.nhlbi.nih.gov/events/2020/nhlbiand-niddk-workshop-exerkines-health-resilience-and-diseases-executive-summary (accessed on 2 August 2023).
- Leuchtmann, A.B.; Adak, V.; Dilbaz, S.; Handschin, C. The role of the skeletal muscle secretome in mediating endurance and resistance training adaptations. Front. Physiol. 2021, 12, 709807. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, J. Metabolic Shifts during Aging and Pathology. Compr. Physiol. 2015, 5, 667–686. [Google Scholar] [CrossRef] [PubMed]
- Matijašić, M.; Meštrović, T.; Paljetak, H.Č.; Perić, M.; Barešić, A.; Verbanac, D. Gut Microbiota beyond Bacteria-Mycobiome, Virome, Archaeome, and Eukaryotic Parasites in IBD. Int. J. Mol. Sci. 2020, 21, 2668. [Google Scholar] [CrossRef]
- Elias-Oliveira, J.; Leite, J.A.; Pereira, Í.S.; Guimarães, J.B.; Manso, G.M.D.C.; Silva, J.S.; Tostes, R.C.; Carlos, D. NLR and Intestinal Dysbiosis-Associated Inflammatory Illness: Drivers or Dampers? Front. Immunol. 2020, 11, 1810. [Google Scholar] [CrossRef] [PubMed]
- Dziewiecka, H.; Buttar, H.S.; Kasperska, A.; Ostapiuk–Karolczuk, J.; Domagalska, M.; Cichoń, J.; Skarpańska-Stejnborn, A. Physical Activity Induced Alterations of Gut Microbiota in Humans: A Systematic Review. BMC Sports Sci. Med. Rehabil. 2022, 14, 122. [Google Scholar] [CrossRef]
- Sales, K.M.; Reimer, R.A. Unlocking a Novel Determinant of Athletic Performance: The Role of the Gut Microbiota, Short-Chain Fatty Acids, and “Biotics” in Exercise. J. Sport Health Sci. 2022, 12, 36–44. [Google Scholar] [CrossRef]
- Lobionda, S.; Sittipo, P.; Kwon, H.Y.; Lee, Y.K. The Role of Gut Microbiota in Intestinal Inflammation with Respect to Diet and Extrinsic Stressors. Microorganisms 2019, 7, 271. [Google Scholar] [CrossRef]
- Aragón-Vela, J.; Solis-Urra, P.; Ruiz-Ojeda, F.J.; Álvarez-Mercado, A.I.; Olivares-Arancibia, J.; Plaza-Diaz, J. Impact of Exercise on Gut Microbiota in Obesity. Nutrients 2021, 13, 3999. [Google Scholar] [CrossRef]
- Glancy, B.; Kane, D.A.; Kavazis, A.N.; Goodwin, M.L.; Willis, W.T.; Gladden, L.B. Mitochondrial Lactate Metabolism: History and Implications for Exercise and Disease. J. Physiol. 2021, 599, 863–888. [Google Scholar] [CrossRef]
- Brooks, G.A.; Osmond, A.D.; Arevalo, J.A.; Duong, J.J.; Curl, C.C.; Moreno-Santillan, D.D.; Leija, R.G. Lactate as a Myokine and Exerkine: Drivers and Signals of Physiology and Metabolism. J. Appl. Physiol. 2023, 134, 529–548. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.-N.; Luo, Y.; Yang, Y.-H.; Fu, J.-T.; Geng, X.-M.; Shi, J.-P.; Yang, J. Lactylation, a Novel Metabolic Reprogramming Code: Current Status and Prospects. Front. Immunol. 2021, 12, 688910. [Google Scholar] [CrossRef]
- Thomas, C.; Bishop, D.J.; Lambert, K.; Mercier, J.; Brooks, G.A. Effects of Acute and Chronic Exercise on Sarcolemmal MCT1 and MCT4 Contents in Human Skeletal Muscles: Current Status. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R1–R14. [Google Scholar] [CrossRef]
- Ramírez de la Piscina-Viúdez, X.; Álvarez-Herms, J.; Bonilla, D.A.; Castañeda-Babarro, A.; Larruskain, J.; Díaz-Ramírez, J.; Ahmetov, I.I.; Martínez-Ascensión, A.; Kreider, R.B.; Odriozola-Martínez, A. Putative Role of MCT1 Rs1049434 Polymorphism in High-Intensity Endurance Performance: Concept and Basis to Understand Possible Individualization Stimulus. Sports 2021, 9, 143. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-Utilizing Bacteria, Isolated from Human Feces, That Produce Butyrate as a Major Fermentation Product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef]
- van der Vossen, E.W.J.; de Goffau, M.C.; Levin, E.; Nieuwdorp, M. Recent Insights into the Role of Microbiome in the Pathogenesis of Obesity. Therap. Adv. Gastroenterol. 2022, 15, 17562848221115320. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-L.; Zhang, L.-Q.; Yang, Y.; Yin, B.-C.; Ye, B.-C.; Zhou, Y. Advances in the Role and Mechanism of Lactic Acid Bacteria in Treating Obesity. Food Bioeng. 2022, 1, 101–115. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-Omics Analysis of Elite Athletes Identifies a Performance-Enhancing Microbe That Functions via Lactate Metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Van Rymenant, E.; Abrankó, L.; Tumova, S.; Grootaert, C.; Van Camp, J.; Williamson, G.; Kerimi, A. Chronic Exposure to Short-Chain Fatty Acids Modulates Transport and Metabolism of Microbiome-Derived Phenolics in Human Intestinal Cells. J. Nutr. Biochem. 2017, 39, 156–168. [Google Scholar] [CrossRef]
- Hinton, J.; Mahmoudi, M. The Role of Exercise in Cardiac Disorders. Medicine 2018, 46, 516–519. [Google Scholar] [CrossRef]
- Tian, D.; Meng, J. Exercise for Prevention and Relief of Cardiovascular Disease: Prognoses, Mechanisms, and Approaches. Oxidative Med. Cell. Longev. 2019, 2019, e3756750. [Google Scholar] [CrossRef]
- Bussotti, M.; Di Marco, S.; Marchese, G. Respiratory Disorders in Endurance Athletes—How Much Do They Really Have to Endure? Open Access J. Sports Med. 2014, 5, 47–63. [Google Scholar] [CrossRef]
- Gosselink, R. Physical Therapy in Adults with Respiratory Disorders: Where Are We? Braz. J. Phys. Ther. 2006, 10, 361–372. [Google Scholar] [CrossRef][Green Version]
- Pedersen, B.K.; Fischer, C.P. Beneficial Health Effects of Exercise—The Role of IL-6 as a Myokine. Trends Pharmacol. Sci. 2007, 28, 152–156. [Google Scholar] [CrossRef]
- Young, J.C. Exercise Prescription for Individuals with Metabolic Disorders. Sports Med. 1995, 19, 43–54. [Google Scholar] [CrossRef]
- Montesi, L.; Moscatiello, S.; Malavolti, M.; Marzocchi, R.; Marchesini, G. Physical Activity for the Prevention and Treatment of Metabolic Disorders. Intern. Emerg. Med. 2013, 8, 655–666. [Google Scholar] [CrossRef]
- Serra, M.V.G.B.; Camargo, P.R.; Zaia, J.E.; Tonello, M.G.M.; Quemelo, P.R.V. Effects of Physical Exercise on Musculoskeletal Disorders, Stress and Quality of Life in Workers. Int. J. Occup. Saf. Ergon. 2018, 24, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Tyler, M.E.; Kaczmarek, K.A.; Rust, K.L.; Subbotin, A.M.; Skinner, K.L.; Danilov, Y.P. Non-Invasive Neuromodulation to Improve Gait in Chronic Multiple Sclerosis: A Randomized Double Blind Controlled Pilot Trial. J. NeuroEng. Rehabil. 2014, 11, 79. [Google Scholar] [CrossRef]
- Nay, K.; Smiles, W.J.; Kaiser, J.; McAloon, L.M.; Loh, K.; Galic, S.; Oakhill, J.S.; Gundlach, A.L.; Scott, J.W. Molecular Mechanisms Underlying the Beneficial Effects of Exercise on Brain Function and Neurological Disorders. Int. J. Mol. Sci. 2021, 22, 4052. [Google Scholar] [CrossRef]
- McDonnell, M.N.; Smith, A.E.; Mackintosh, S.F. Aerobic Exercise to Improve Cognitive Function in Adults with Neurological Disorders: A Systematic Review. Arch. Phys. Med. Rehabil. 2011, 92, 1044–1052. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Behan, P.O. Fatigue in Neurological Disorders. Lancet 2004, 363, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Budde, H.; Velasques, B.; Ribeiro, P.; Soya, H. Editorial: Neuromodulation of Exercise: Impact on Different Kinds of Behavior. Front. Neurosci. 2020, 14, 455. [Google Scholar]
- Gronwald, T.; Budde, H. Commentary: Physical Exercise as Personalized Medicine for Dementia Prevention? Front. Physiol. 2019, 10, 1358. [Google Scholar] [PubMed]
- Müllers, P.; Taubert, M.; Müller, N.G. Physical Exercise as Personalized Medicine for Dementia Prevention? Front. Physiol. 2019, 10, 672. [Google Scholar]
- Gouarné, C.; Groussard, C.; Gratas-Delamarche, A.; Delamarche, P.; Duclos, M. Overnight Urinary Cortisol and Cortisone Add New Insights into Adaptation to Training. Med. Sci. Sports Exerc. 2005, 37, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Ehlert, U.; Gaab, J.; Heinrichs, M. Psychoneuroendocrinological Contributions to the Etiology of Depression, Posttraumatic Stress Disorder, and Stress-Related Bodily Disorders: The Role of the Hypothalamus–Pituitary–Adrenal Axis. Biol. Psychol. 2001, 57, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Valverde, D. Potential Role of Cannabidiol on Sports Recovery: A Narrative Review. Front. Physiol. 2021, 12, 722550. [Google Scholar] [CrossRef]
- Raichlen, D.A.; Foster, A.D.; Seillier, A.; Giuffrida, A.; Gerdeman, G.L. Exercise-Induced Endocannabinoid Signaling Is Modulated by Intensity. Eur. J. Appl. Physiol. 2013, 113, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Gunduz-Cinar, O.; Hill, M.N.; McEwen, B.S.; Holmes, A. Amygdala FAAH and Anandamide: Mediating Protection and Recovery from Stress. Trends Pharmacol. Sci. 2013, 34, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.M.; Vecchiarelli, H.A.; Morena, M.; Lee, T.T.Y.; Hermanson, D.J.; Kim, A.B.; McLaughlin, R.J.; Hassan, K.I.; Kühne, C.; Wotjak, C.T.; et al. Corticotropin-Releasing Hormone Drives Anandamide Hydrolysis in the Amygdala to Promote Anxiety. J. Neurosci. 2015, 35, 3879–3892. [Google Scholar] [CrossRef]
- Heijnen, S.; Hommel, B.; Kibele, A.; Colzato, L.S. Neuromodulation of Aerobic Exercise—A Review. Front. Psychol. 2016, 6, 1890. [Google Scholar] [CrossRef]
- Heyman, E.; Gamelin, F.-X.; Goekint, M.; Piscitelli, F.; Roelands, B.; Leclair, E.; Di Marzo, V.; Meeusen, R. Intense Exercise Increases Circulating Endocannabinoid and BDNF Levels in Humans--Possible Implications for Reward and Depression. Psychoneuroendocrinology 2012, 37, 844–851. [Google Scholar] [CrossRef]
- Leckie, R.L.; Oberlin, L.E.; Voss, M.W.; Prakash, R.S.; Szabo-Reed, A.; Chaddock-Heyman, L.; Phillips, S.M.; Gothe, N.P.; Mailey, E.; Vieira-Potter, V.J.; et al. BDNF Mediates Improvements in Executive Function Following a 1-Year Exercise Intervention. Front. Hum. Neurosci. 2014, 8, 985. [Google Scholar]
- Patrick, R.P.; Ames, B.N. Vitamin D and the Omega-3 Fatty Acids Control Serotonin Synthesis and Action, Part 2: Relevance for ADHD, Bipolar Disorder, Schizophrenia, and Impulsive Behavior. FASEB J. 2015, 29, 2207–2222. [Google Scholar] [CrossRef]
- Mills, K.C. Serotonin Syndrome. A Clinical Update. Crit. Care Clin. 1997, 13, 763–783. [Google Scholar] [CrossRef] [PubMed]
- Birmes, P.; Coppin, D.; Schmitt, L.; Lauque, D. Serotonin Syndrome: A Brief Review. CMAJ 2003, 168, 1439–1442. [Google Scholar] [PubMed]
- Nieoullon, A. Dopamine and the Regulation of Cognition and Attention. Prog. Neurobiol. 2002, 67, 53–83. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, N.; Alghasham, A. Central Dopaminergic System and Its Implications in Stress-Mediated Neurological Disorders and Gastric Ulcers: Short Review. Adv. Pharmacol. Sci. 2012, 2012, 182671. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Reyes-Resina, I.; Navarro, G. Dopamine in Health and Disease: Much More Than a Neurotransmitter. Biomedicines 2021, 9, 109. [Google Scholar] [CrossRef]
- Weaver, S.R.; Skinner, B.D.; Furlong, R.; Lucas, R.A.I.; Cable, N.T.; Rendeiro, C.; McGettrick, H.M.; Lucas, S.J.E. Cerebral Hemodynamic and Neurotrophic Factor Responses Are Dependent on the Type of Exercise. Front. Physiol. 2021, 11, 609935. [Google Scholar] [CrossRef]
- Gutiérrez-Vargas, R.; Ugalde-Ramírez, A.; Rico-González, M.; Pino-Ortega, J.; González-Hernández, J.; Rojas-Valverde, D. A Systematic Review of the Effects of Football Playing on Changes in Serum Brain-Derived Neurotrophic Factor Level. Appl. Sci. 2021, 11, 11828. [Google Scholar] [CrossRef]
- Heyn, P.; Abreu, B.C.; Ottenbacher, K.J. The Effects of Exercise Training on Elderly Persons with Cognitive Impairment and Dementia: A Meta-Analysis. Arch. Phys. Med. Rehabil. 2004, 85, 1694–1704. [Google Scholar] [CrossRef]
- Arêas, F.Z.D.S.; Nakamura-Palacios, E.M.; Boening, A.; Arêas, G.P.T.; Nascimento, L.R. Does Neuromodulation Transcranial Direct Current Stimulation (TDCS) Associated with Peripheral Stimulation through Exercise to Walk Have an Impact on Falls in People with Parkinson’s Disease? Med. Hypotheses 2020, 144, 109916. [Google Scholar] [CrossRef]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Goetz, C.G.; Marin, C.; Kordower, J.H.; Rodriguez, M.; Hirsch, E.C.; Farrer, M.; Schapira, A.H.V.; Halliday, G. Missing Pieces in the Parkinson’s Disease Puzzle. Nat. Med. 2010, 16, 653–661. [Google Scholar] [CrossRef]
- Davie, C.A. A Review of Parkinson’s Disease. Br. Med. Bull. 2008, 86, 109–127. [Google Scholar] [CrossRef]
- Cugusi, L.; Manca, A.; Bergamin, M.; Di Blasio, A.; Monticone, M.; Deriu, F.; Mercuro, G. Aquatic Exercise Improves Motor Impairments in People with Parkinson’s Disease, with Similar or Greater Benefits than Land-Based Exercise: A Systematic Review. J. Physiother. 2019, 65, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Wong-Yu, I.S.K.; Mak, M.K.Y. Effects of Exercise on Falls, Balance, and Gait Ability in Parkinson’s Disease: A Meta-Analysis. Neurorehabil. Neural Repair 2016, 30, 512–527. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.-F.; Yang, T.; Yu, S.-X.; Huang, H.-D.; Jiang, L.-L.; Gu, J.-W.; Kuang, Y.-Q. Aerobic Exercise for Parkinson’s Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. PLoS ONE 2014, 9, e100503. [Google Scholar] [CrossRef]
- Ni, X.; Liu, S.; Lu, F.; Shi, X.; Guo, X. Efficacy and Safety of Tai Chi for Parkinson’s Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. PLoS ONE 2014, 9, e99377. [Google Scholar] [CrossRef]
- Kabra, A.; Sharma, R.; Kabra, R.; Baghel, U.S. Emerging and Alternative Therapies For Parkinson Disease: An Updated Review. Curr. Pharm. Des. 2018, 24, 2573–2582. [Google Scholar] [CrossRef]
- Shanahan, J.; Morris, M.E.; Bhriain, O.N.; Saunders, J.; Clifford, A.M. Dance for People With Parkinson Disease: What Is the Evidence Telling Us? Arch. Phys. Med. Rehabil. 2015, 96, 141–153. [Google Scholar] [CrossRef]
- De Santis, K.K.; Kaplan, I. The Motor and the Non-Motor Outcomes of Nordic Walking in Parkinson’s Disease: A Systematic Review. J. Bodyw. Mov. Ther. 2020, 24, 4–10. [Google Scholar] [CrossRef]
- Chung, C.L.H.; Thilarajah, S.; Tan, D. Effectiveness of Resistance Training on Muscle Strength and Physical Function in People with Parkinson’s Disease: A Systematic Review and Meta-Analysis. Clin. Rehabil. 2016, 30, 11–23. [Google Scholar] [CrossRef]
- Cass, S.P. Alzheimer’s Disease and Exercise: A Literature Review. Curr. Sports Med. Rep. 2017, 16, 19–22. [Google Scholar] [CrossRef]
- Meng, Q.; Lin, M.-S.; Tzeng, I.-S. Relationship Between Exercise and Alzheimer’s Disease: A Narrative Literature Review. Front. Neurosci. 2020, 14, 131. [Google Scholar] [CrossRef]
- Boyle, C.P.; Raji, C.A.; Erickson, K.I.; Lopez, O.L.; Becker, J.T.; Gach, H.M.; Longstreth, W.T.; Teverovskiy, L.; Kuller, L.H.; Carmichael, O.T.; et al. Physical Activity, Body Mass Index, and Brain Atrophy in Alzheimer’s Disease. Neurobiol. Aging 2015, 36, S194–S202. [Google Scholar] [CrossRef] [PubMed]
- Braskie, M.N.; Boyle, C.P.; Rajagopalan, P.; Gutman, B.A.; Toga, A.W.; Raji, C.A.; Tracy, R.P.; Kuller, L.H.; Becker, J.T.; Lopez, O.L.; et al. Physical Activity, Inflammation, and Volume of the Aging Brain. Neuroscience 2014, 273, 199–209. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, A.; Olaso-Gonzalez, G.; Arc-Chagnaud, C.; Millan, F.; Salvador-Pascual, A.; García-Lucerga, C.; Blasco-Lafarga, C.; Garcia-Dominguez, E.; Carretero, A.; Correas, A.G.; et al. Physical Exercise in the Prevention and Treatment of Alzheimer’s Disease. J. Sport. Health Sci. 2020, 9, 394–404. [Google Scholar] [CrossRef]
- Hoffmann, K.; Sobol, N.A.; Frederiksen, K.S.; Beyer, N.; Vogel, A.; Vestergaard, K.; Brændgaard, H.; Gottrup, H.; Lolk, A.; Wermuth, L.; et al. Moderate-to-High Intensity Physical Exercise in Patients with Alzheimer’s Disease: A Randomized Controlled Trial. J. Alzheimers Dis. 2016, 50, 443–453. [Google Scholar] [CrossRef]
- Agüera Sánchez, M.Á.; Barbancho Ma, M.Á.; García-Casares, N. Effect of physical exercise on Alzheimer’s disease. A sistematic review. Aten. Primaria 2020, 52, 307–318. [Google Scholar] [CrossRef]
- Motl, R.W.; McAuley, E.; Snook, E.M. Physical Activity and Multiple Sclerosis: A Meta-Analysis. Mult. Scler. 2005, 11, 459–463. [Google Scholar] [CrossRef]
- Shannon, J.B. Multiple Sclerosis Sourcebook; Omnigraphics, Inc.: Detroit, MI, USA, 2007; ISBN 978-0-7808-0998-7. [Google Scholar]
- Snook, E.M.; Motl, R.W. Effect of Exercise Training on Walking Mobility in Multiple Sclerosis: A Meta-Analysis. Neurorehabil. Neural Repair 2009, 23, 108–116. [Google Scholar] [CrossRef]
- Heine, M.; van de Port, I.; Rietberg, M.B.; van Wegen, E.E.H.; Kwakkel, G. Exercise Therapy for Fatigue in Multiple Sclerosis. Cochrane Database Syst. Rev. 2015, 2015, CD009956. [Google Scholar] [CrossRef] [PubMed]
- Bello-Haas, V.D. Physical Therapy for Individuals with Amyotrophic Lateral Sclerosis: Current Insights. Degener. Neurol. Neuromuscul. Dis. 2018, 8, 45–54. [Google Scholar] [CrossRef]
- Paganoni, S.; Karam, C.; Joyce, N.; Bedlack, R.; Carter, G.T. Comprehensive Rehabilitative Care across the Spectrum of Amyotrophic Lateral Sclerosis. NeuroRehabilitation 2015, 37, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Li, X.; Li, C.; Tsang, R.C.C.; Chen, Y.; Ge, Y.; Gao, Q. Effects of Exercise in Patients With Amyotrophic Lateral Sclerosis: A Systematic Review and Meta-Analysis. Am. J. Phys. Med. Rehabil. 2020, 99, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Lisle, S.; Tennison, M. Amyotrophic Lateral Sclerosis: The Role of Exercise. Curr. Sports Med. Rep. 2015, 14, 45–46. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.C.; Wisniewski, P.J.; Noji, M.; McGuinness, L.R.; Häggblom, M.M.; Lightfoot, S.A.; Joseph, L.B.; Kerkhof, L.J. The Effect of Diet and Exercise on Intestinal Integrity and Microbial Diversity in Mice. PLoS ONE 2016, 11, e0150502. [Google Scholar] [CrossRef]
- Gubert, C.; Kong, G.; Renoir, T.; Hannan, A.J. Exercise, Diet and Stress as Modulators of Gut Microbiota: Implications for Neurodegenerative Diseases. Neurobiol. Dis. 2020, 134, 104621. [Google Scholar] [CrossRef]
- Vijay, N.; Morris, M.E. Role of monocarboxylate transporters in drug delivery to the brain. Curr. Pharm. Des. 2014, 20, 1487–1498. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Felice, V.D.; Nally, K.; Savignac, H.M.; Claesson, M.J.; Scully, P.; Woznicki, J.; Hyland, N.P.; Shanahan, F.; Quigley, E.M.; et al. Disturbance of the gut microbiota in early-life selectively affects visceral pain in adulthood without impacting cognitive or anxiety-related behaviors in male rats. Neuroscience 2014, 277, 885–901. [Google Scholar] [CrossRef]
- Cenit, M.C.; Sanz, Y.; Codoñer-Franch, P. Influence of Gut Microbiota on Neuropsychiatric Disorders. World J. Gastroenterol. 2017, 23, 5486–5498. [Google Scholar] [CrossRef]
- Stefano, G.B.; Pilonis, N.; Ptacek, R.; Raboch, J.; Vnukova, M.; Kream, R.M. Gut, Microbiome, and Brain Regulatory Axis: Relevance to Neurodegenerative and Psychiatric Disorders. Cell. Mol. Neurobiol. 2018, 38, 1197–1206. [Google Scholar] [CrossRef]
- Spielman, L.J.; Gibson, D.L.; Klegeris, A. Unhealthy Gut, Unhealthy Brain: The Role of the Intestinal Microbiota in Neurodegenerative Diseases. Neurochem. Int. 2018, 120, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.; Lomis, N.; Kahouli, I.; Dia, S.Y.; Singh, S.P.; Prakash, S. Microbiome, Probiotics and Neurodegenerative Diseases: Deciphering the Gut Brain Axis. Cell. Mol. Life Sci. 2017, 74, 3769–3787. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G. Chapter Eleven—Cognitive Function and the Microbiome. In International Review of Neurobiology; Cryan, J.F., Clarke, G., Eds.; Gut Microbiome and Behavior; Academic Press: Cambridge, MA, USA, 2016; Volume 131, pp. 227–246. [Google Scholar]
- Evans, J.; Morris, L.; Marchesi, J. The Gut Microbiome: The Role of a Virtual Organ in the Endocrinology of the Host. J. Endocrinol. 2013, 218, R37–R47. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Berg Miller, M.E.; Pence, B.D.; Whitlock, K.; Nehra, V.; Gaskins, H.R.; White, B.A.; Fryer, J.D.; Woods, J.A. Voluntary and Forced Exercise Differentially Alters the Gut Microbiome in C57BL/6J Mice. J. Appl. Physiol. 2015, 118, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Batacan, R.B.; Fenning, A.S.; Dalbo, V.J.; Scanlan, A.T.; Duncan, M.J.; Moore, R.J.; Stanley, D. A Gut Reaction: The Combined Influence of Exercise and Diet on Gastrointestinal Microbiota in Rats. J. Appl. Microbiol. 2017, 122, 1627–1638. [Google Scholar] [CrossRef]
- Matsumoto, M.; Inoue, R.; Tsukahara, T.; Ushida, K.; Chiji, H.; Matsubara, N.; Hara, H. Voluntary Running Exercise Alters Microbiota Composition and Increases N-Butyrate Concentration in the Rat Cecum. Biosci. Biotechnol. Biochem. 2008, 72, 572–576. [Google Scholar] [CrossRef]
- Allen, J.M.; Mailing, L.J.; Cohrs, J.; Salmonson, C.; Fryer, J.D.; Nehra, V.; Hale, V.L.; Kashyap, P.; White, B.A.; Woods, J.A. Exercise Training-Induced Modification of the Gut Microbiota Persists after Microbiota Colonization and Attenuates the Response to Chemically-Induced Colitis in Gnotobiotic Mice. Gut Microbes 2018, 9, 115–130. [Google Scholar] [CrossRef]
- Yin, Y.; Guo, Q.; Zhou, X.; Duan, Y.; Yang, Y.; Gong, S.; Han, M.; Liu, Y.; Yang, Z.; Chen, Q.; et al. Role of Brain-Gut-Muscle Axis in Human Health and Energy Homeostasis. Front. Nutr. 2022, 9, 947033. [Google Scholar] [CrossRef]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Front. Physiol. 2016, 7, 51. [Google Scholar]
- Hoffman-Goetz, L.; Pervaiz, N.; Packer, N.; Guan, J. Freewheel Training Decreases Pro- and Increases Anti-Inflammatory Cytokine Expression in Mouse Intestinal Lymphocytes. Brain Behav. Immun. 2010, 24, 1105–1115. [Google Scholar] [CrossRef]
- Hoffman-Goetz, L.; Quadrilatero, J. Treadmill Exercise in Mice Increases Intestinal Lymphocyte Loss via Apoptosis. Acta Physiol. Scand. 2003, 179, 289–297. [Google Scholar] [CrossRef]
- Chapp, A.D.; Behnke, J.E.; Driscoll, K.M.; Hahka, T.; LaLonde, Z.; Shan, Z.; Chen, Q.-H. Elevated L-Lactate Promotes Major Cellular Pathologies Associated with Neurodegenerative Diseases. Neurosci. Bull. 2021, 37, 380–384. [Google Scholar] [CrossRef]
- Mason, S. A Novel, Multi-Faceted Perception of Lactate in Neurology. Front. Neurosci. 2020, 14, 460. [Google Scholar] [CrossRef]
- Ross, J.M.; Öberg, J.; Brené, S.; Coppotelli, G.; Terzioglu, M.; Pernold, K.; Goiny, M.; Sitnikov, R.; Kehr, J.; Trifunovic, A.; et al. High Brain Lactate Is a Hallmark of Aging and Caused by a Shift in the Lactate Dehydrogenase A/B Ratio. Proc. Natl. Acad. Sci. USA 2010, 107, 20087–20092. [Google Scholar] [CrossRef]
- Liguori, C.; Stefani, A.; Sancesario, G.; Sancesario, G.M.; Marciani, M.G.; Pierantozzi, M. CSF Lactate Levels, τ Proteins, Cognitive Decline: A Dynamic Relationship in Alzheimer’s Disease. J. Neurol. Neurosurg. Psychiatry 2015, 86, 655–659. [Google Scholar] [CrossRef]
- Messonnier, L.; Freund, H.; Féasson, L.; Prieur, F.; Castells, J.; Denis, C.; Linossier, M.T.; Geyssant, A.; Lacour, J.R. Blood Lactate Exchange and Removal Abilities after Relative High-Intensity Exercise: Effects of Training in Normoxia and Hypoxia. Eur. J. Appl. Physiol. 2001, 84, 403–412. [Google Scholar] [CrossRef]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef]
- Drake, J.C.; Wilson, R.J.; Yan, Z. Molecular mechanisms for mitochondrial adaptation to exercise training in skeletal muscle. FASEB J. 2016, 30, 13–22. [Google Scholar] [CrossRef]
- Denou, E.; Marcinko, K.; Surette, M.G.; Steinberg, G.R.; Schertzer, J.D. High-intensity exercise training increases the diversity and metabolic capacity of the mouse distal gut microbiota during diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E982–E993. [Google Scholar] [CrossRef]
- Martínez-Guardado, I.; Arboleya, S.; Grijota, F.J.; Kaliszewska, A.; Gueimonde, M.; Arias, N. The Therapeutic Role of Exercise and Probiotics in Stressful Brain Conditions. Int. J. Mol. Sci. 2022, 23, 3610. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
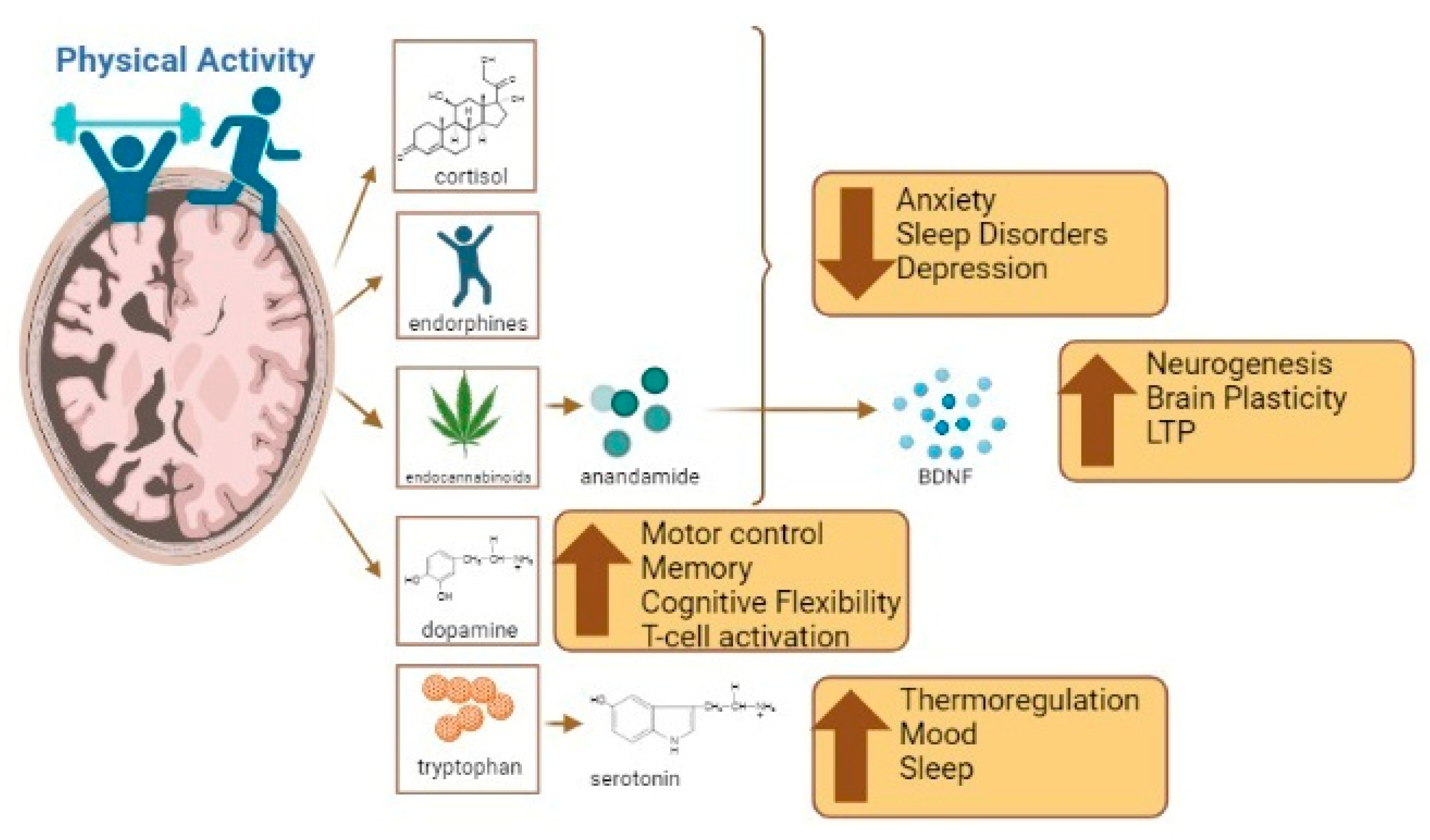
| Gut Microbiota Changes | Brain Changes |
|---|---|
| - Increases Firmicutes and Actinobacteria | - Decreases anxiety and depression |
| - Increases butyrate-producing bacteria, such as Roseburia hominis, Faecalibacterium pausnitzii, and Ruminococcaceae | - Improves mood |
| - Increases butyrate concentration | - Improves motor control |
| - Reduces transient stool time in the gastrointestinal tract | - Decreases inflammation through t-cell activation |
| - Increases key antioxidant enzymes (catalase and glutathione peroxidase), anti-inflammatory cytokines (including IL-10), and antiapoptotic proteins (including Bcl-2) in intestinal lymphocytes | - Improves memory, long-term potentiation and cognitive flexibility |
| - Decreases proinflammatory cytokines (TNF-α and IL-17) and proapoptotic proteins (caspase 3 and 7), leading to an overall reduction in gut inflammation | - Improves sleep |
| - Increases SCFAs, followed by a decrease in Bacterioides and an increase in Faecalibacterium and Lachnospira | - Improves neurogenesis and brain plasticity |
| - Modulates gastrointestinal motility | - Improves brain metabolism through mitochondria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas-Valverde, D.; Bonilla, D.A.; Gómez-Miranda, L.M.; Calleja-Núñez, J.J.; Arias, N.; Martínez-Guardado, I. Examining the Interaction between Exercise, Gut Microbiota, and Neurodegeneration: Future Research Directions. Biomedicines 2023, 11, 2267. https://doi.org/10.3390/biomedicines11082267
Rojas-Valverde D, Bonilla DA, Gómez-Miranda LM, Calleja-Núñez JJ, Arias N, Martínez-Guardado I. Examining the Interaction between Exercise, Gut Microbiota, and Neurodegeneration: Future Research Directions. Biomedicines. 2023; 11(8):2267. https://doi.org/10.3390/biomedicines11082267
Chicago/Turabian StyleRojas-Valverde, Daniel, Diego A. Bonilla, Luis M. Gómez-Miranda, Juan J. Calleja-Núñez, Natalia Arias, and Ismael Martínez-Guardado. 2023. "Examining the Interaction between Exercise, Gut Microbiota, and Neurodegeneration: Future Research Directions" Biomedicines 11, no. 8: 2267. https://doi.org/10.3390/biomedicines11082267
APA StyleRojas-Valverde, D., Bonilla, D. A., Gómez-Miranda, L. M., Calleja-Núñez, J. J., Arias, N., & Martínez-Guardado, I. (2023). Examining the Interaction between Exercise, Gut Microbiota, and Neurodegeneration: Future Research Directions. Biomedicines, 11(8), 2267. https://doi.org/10.3390/biomedicines11082267