
The Role of Adipokines in Health and Disease

 ,
,  ,
,  ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Methods
3. Obesity and Adipose Tissue Dysregulation
3.1. Obesity Pandemic
3.2. White Adipocytes
3.3. Brown Adipocytes
3.4. Beige Adipocytes
4. Adipokines and Inflammation
4.1. Proinflammatory Adipokines
4.2. Antiinflamatory Adipokines
5. Adipokines and Cardiovascular Disease
6. Adipokines and Atherosclerosis
7. Adipokines and Mental Disease
8. Adipokines and Eating Behaviors
9. Adipokines and Metabolic Disease
10. Adipokines and Cancer
11. The Role of Microbiota in Adipokines
12. The Role of Nutrition in Adipokines
13. The Role of Physical Activity in Adipokines
14. Practical Applications
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fasshauer, M.; Blüher, M. Adipokines in Health and Disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, P. The Role of Adipokines in Chronic Inflammation. Immunotargets Ther. 2016, 5, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Mattu, H.S.; Randeva, H.S. Role of Adipokines in Cardiovascular Disease. J. Endocrinol. 2013, 216, T17–T36. [Google Scholar] [CrossRef]
- Lau, W.B.; Ohashi, K.; Wang, Y.; Ogawa, H.; Murohara, T.; Ma, X.-L.; Ouchi, N. Role of Adipokines in Cardiovascular Disease. Circ. J. 2017, 81, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Farkhondeh, T.; Llorens, S.; Pourbagher-Shahri, A.M.; Ashrafizadeh, M.; Talebi, M.; Shakibaei, M.; Samarghandian, S. An Overview of the Role of Adipokines in Cardiometabolic Diseases. Molecules 2020, 25, 5218. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Rocha, D.Q.C.; McIntyre, R.S.; Mesquita, L.M.; Köhler, C.A.; Hyphantis, T.N.; Sales, P.M.G.; Machado-Vieira, R.; Berk, M. Adipokines as Emerging Depression Biomarkers: A Systematic Review and Meta-Analysis. J. Psychiatr. Res. 2014, 59, 28–37. [Google Scholar] [CrossRef]
- Morris, A.A.; Ahmed, Y.; Stoyanova, N.; Hooper, W.C.; De Staerke, C.; Gibbons, G.; Quyyumi, A.; Vaccarino, V. The Association between Depression and Leptin Is Mediated by Adiposity. Psychosom. Med. 2012, 74, 483–488. [Google Scholar] [CrossRef]
- Yang, J.L.; Liu, D.X.; Jiang, H.; Pan, F.; Ho, C.S.; Ho, R.C. The Effects of High-Fat-Diet Combined with Chronic Unpredictable Mild Stress on Depression-like Behavior and Leptin/LepRb in Male Rats. Sci. Rep. 2016, 6, 35239. [Google Scholar] [CrossRef]
- Balistreri, C.R.; Caruso, C.; Candore, G. The Role of Adipose Tissue and Adipokines in Obesity-Related Inflammatory Diseases. Mediat. Inflamm. 2010, 2010, 802078. [Google Scholar] [CrossRef]
- Sahu, B.; Bal, N.C. Adipokines from White Adipose Tissue in Regulation of Whole Body Energy Homeostasis. Biochimie 2023, 204, 92–107. [Google Scholar] [CrossRef]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a Key Adipokine in Obesity Related Liver Diseases. World J. Gastroenterol. 2011, 17, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M. Adipose Tissue, Obesity and Adipokines: Role in Cancer Promotion. Horm. Mol. Biol. Clin. Investig. 2015, 21, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Suriano, F.; Van Hul, M.; Cani, P.D. Gut Microbiota and Regulation of Myokine-Adipokine Function. Curr. Opin. Pharmacol. 2020, 52, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Grases-Pintó, B.; Abril-Gil, M.; Castell, M.; Rodríguez-Lagunas, M.J.; Burleigh, S.; Fåk Hållenius, F.; Prykhodko, O.; Pérez-Cano, F.J.; Franch, À. Influence of Leptin and Adiponectin Supplementation on Intraepithelial Lymphocyte and Microbiota Composition in Suckling Rats. Front. Immunol. 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Kedenko, L.; Antonielli, L.; Kedenko, I.; Gemeier, C.; Leitner, M.; Kautzky-Willer, A.; Paulweber, B.; Hackl, E. Gut Microbiota Dysbiosis Associated with Glucose Metabolism Disorders and the Metabolic Syndrome in Older Adults. Benef. Microbes 2017, 8, 545–556. [Google Scholar] [CrossRef]
- Czaja-Stolc, S.; Potrykus, M.; Stankiewicz, M.; Kaska, Ł.; Małgorzewicz, S. Pro-Inflammatory Profile of Adipokines in Obesity Contributes to Pathogenesis, Nutritional Disorders, and Cardiovascular Risk in Chronic Kidney Disease. Nutrients 2022, 14, 1457. [Google Scholar] [CrossRef]
- Huang, F.; Del-Río-Navarro, B.E.; Leija-Martinez, J.; Torres-Alcantara, S.; Ruiz-Bedolla, E.; Hernández-Cadena, L.; Barraza-Villarreal, A.; Romero-Nava, R.; Sanchéz-Muñoz, F.; Villafaña, S.; et al. Effect of Omega-3 Fatty Acids Supplementation Combined with Lifestyle Intervention on Adipokines and Biomarkers of Endothelial Dysfunction in Obese Adolescents with Hypertriglyceridemia. J. Nutr. Biochem. 2019, 64, 162–169. [Google Scholar] [CrossRef]
- Senesi, P.; Luzi, L.; Terruzzi, I. Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions. Int. J. Mol. Sci. 2020, 21, 8372. [Google Scholar] [CrossRef]
- Sirico, F.; Bianco, A.; D’Alicandro, G.; Castaldo, C.; Montagnani, S.; Spera, R.; Di Meglio, F.; Nurzynska, D. Effects of Physical Exercise on Adiponectin, Leptin, and Inflammatory Markers in Childhood Obesity: Systematic Review and Meta-Analysis. Child. Obes. 2018, 14, 207–217. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Mottillo, E.P.; Granneman, J.G. Adipose Tissue Plasticity from WAT to BAT and in Between. Biochim. Biophys. Acta 2014, 1842, 358–369. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. What We Talk about When We Talk about Fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, A.; Buoncristiano, M.; Kovacs, V.A.; Yngve, A.; Spiroski, I.; Obreja, G.; Starc, G.; Pérez, N.; Rito, A.I.; Kunešová, M.; et al. Prevalence of Severe Obesity among Primary School Children in 21 European Countries. Obes. Facts 2019, 12, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.K.; Teymourian, Y.; Tursini, M.S. The Effect of Sugar and Processed Food Imports on the Prevalence of Overweight and Obesity in 172 Countries. Glob. Health 2018, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Blaak, E.E. Adipose Tissue Dysfunction and Impaired Metabolic Health in Human Obesity: A Matter of Oxygen? Front. Endocrinol. 2015, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.L.; Bensley, J.G.; Wood-Bradley, R.J.; Cullen-McEwen, L.A.; Bertram, J.F.; Armitage, J.A. White Adipocytes: More than Just Fat Depots. Int. J. Biochem. Cell. Biol. 2012, 44, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M. Leptin and the Endocrine Control of Energy Balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Hassan-Zadeh, V. IL-6 Signalling Pathways and the Development of Type 2 Diabetes. Inflammopharmacology 2018, 26, 685–698. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscles, Exercise and Obesity: Skeletal Muscle as a Secretory Organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Koper-Lenkiewicz, O.M.; Sutkowska, K.; Wawrusiewicz-Kurylonek, N.; Kowalewska, E.; Matowicka-Karna, J. Proinflammatory Cytokines (IL-1, -6, -8, -15, -17, -18, -23, TNF-α) Single Nucleotide Polymorphisms in Rheumatoid Arthritis-A Literature Review. Int. J. Mol. Sci. 2022, 23, 2106. [Google Scholar] [CrossRef]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [PubMed]
- Haider, D.G.; Holzer, G.; Schaller, G.; Weghuber, D.; Widhalm, K.; Wagner, O.; Kapiotis, S.; Wolzt, M. The Adipokine Visfatin Is Markedly Elevated in Obese Children. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 548–549. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Frontini, A.; Cinti, S. Distribution and Development of Brown Adipocytes in the Murine and Human Adipose Organ. Cell Metab. 2010, 11, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Fisher, F.M.; Chui, P.C.; Antonellis, P.J.; Bina, H.A.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E. Obesity Is a Fibroblast Growth Factor 21 (FGF21)-Resistant State. Diabetes 2010, 59, 2781–2789. [Google Scholar] [CrossRef]
- Vijgen, G.H.E.J.; Bouvy, N.D.; Teule, G.J.J.; Brans, B.; Schrauwen, P.; van Marken Lichtenbelt, W.D. Brown Adipose Tissue in Morbidly Obese Subjects. PLoS ONE 2011, 6, e17247. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N. The Physiological and Pathophysiological Role of Adiponectin and Adiponectin Receptors in the Peripheral Tissues and CNS. FEBS Lett. 2008, 582, 74–80. [Google Scholar] [CrossRef]
- Chen, Y.; Siegel, F.; Kipschull, S.; Haas, B.; Fröhlich, H.; Meister, G.; Pfeifer, A. MiR-155 Regulates Differentiation of Brown and Beige Adipocytes via a Bistable Circuit. Nat. Commun. 2013, 4, 1769. [Google Scholar] [CrossRef]
- Ikeda, K.; Maretich, P.; Kajimura, S. The Common and Distinct Features of Brown and Beige Adipocytes. Trends Endocrinol. Metab. 2018, 29, 191–200. [Google Scholar] [CrossRef]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Fabbiano, S.; Suárez-Zamorano, N.; Rigo, D.; Veyrat-Durebex, C.; Stevanovic Dokic, A.; Colin, D.J.; Trajkovski, M. Caloric Restriction Leads to Browning of White Adipose Tissue through Type 2 Immune Signaling. Cell Metab. 2016, 24, 434–446. [Google Scholar] [CrossRef]
- Kajimura, S.; Spiegelman, B.M.; Seale, P. Brown and Beige Fat: Physiological Roles beyond Heat Generation. Cell Metab. 2015, 22, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Guerre-Millo, M. Adipose Tissue and Adipokines: For Better or Worse. Diabetes Metab. 2004, 30, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Francisco, V.; Pino, J.; Gonzalez-Gay, M.A.; Mera, A.; Lago, F.; Gómez, R.; Mobasheri, A.; Gualillo, O. Adipokines and Inflammation: Is It a Question of Weight? Br. J. Pharmacol. 2018, 175, 1569–1579. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators Linking Adipose Tissue, Inflammation and Immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity Induces a Phenotypic Switch in Adipose Tissue Macrophage Polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef]
- Deiuliis, J.; Shah, Z.; Shah, N.; Needleman, B.; Mikami, D.; Narula, V.; Perry, K.; Hazey, J.; Kampfrath, T.; Kollengode, M.; et al. Visceral Adipose Inflammation in Obesity Is Associated with Critical Alterations in Tregulatory Cell Numbers. PLoS ONE 2011, 6, e16376. [Google Scholar] [CrossRef]
- Ohmura, K.; Ishimori, N.; Ohmura, Y.; Tokuhara, S.; Nozawa, A.; Horii, S.; Andoh, Y.; Fujii, S.; Iwabuchi, K.; Onoé, K.; et al. Natural Killer T Cells Are Involved in Adipose Tissues Inflammation and Glucose Intolerance in Diet-Induced Obese Mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 193–199. [Google Scholar] [CrossRef]
- Rodríguez, L.; Graniel, J.; Ortiz, R. Effect of Leptin on Activation and Cytokine Synthesis in Peripheral Blood Lymphocytes of Malnourished Infected Children. Clin. Exp. Immunol. 2007, 148, 478–485. [Google Scholar] [CrossRef]
- Grewal, T.; Enrich, C.; Rentero, C.; Buechler, C. Annexins in Adipose Tissue: Novel Players in Obesity. Int. J. Mol. Sci. 2019, 20, 3449. [Google Scholar] [CrossRef] [PubMed]
- Freitas Lima, L.C.; de Braga, V.A.; do Socorro de França Silva, M.; de Cruz, J.C.; Sousa Santos, S.H.; de Oliveira Monteiro, M.M.; de Moura Balarini, C. Adipokines, Diabetes and Atherosclerosis: An Inflammatory Association. Front. Physiol. 2015, 6, 304. [Google Scholar] [CrossRef] [PubMed]
- Pivtorak, K.V.; Shevchuk, N.A.; Pivtorak, N.A.; Fedzhaga, I.V. Correction of Adipocyte Secretion Disorders in Patients with Non-Alcoholic Fatty Liver Disease with Overweight and Obesity. Wiad. Lek. 2019, 72, 1477–1480. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Hernández, A.; Beneit, N.; Díaz-Castroverde, S.; Escribano, Ó. Differential Role of Adipose Tissues in Obesity and Related Metabolic and Vascular Complications. Int. J. Endocrinol. 2016, 2016, 1216783. [Google Scholar] [CrossRef] [PubMed]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; de la Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, H.; Yin, C.; Lan, X.; Wu, L.; Du, X.; Griffiths, H.R.; Gao, D. Adipokines, Hepatokines and Myokines: Focus on Their Role and Molecular Mechanisms in Adipose Tissue Inflammation. Front. Endocrinol. 2022, 13, 873699. [Google Scholar] [CrossRef]
- Kwon, O.; Kim, K.W.; Kim, M.-S. Leptin Signalling Pathways in Hypothalamic Neurons. Cell Mol. Life Sci. 2016, 73, 1457–1477. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Sánchez-Jiménez, F.; Vilariño-García, T.; Sánchez-Margalet, V. Role of Leptin in Inflammation and Vice Versa. Int. J. Mol. Sci. 2020, 21, 5887. [Google Scholar] [CrossRef]
- La Cava, A. Leptin in Inflammation and Autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef]
- Matarese, G.; La Cava, A.; Sanna, V.; Lord, G.M.; Lechler, R.I.; Fontana, S.; Zappacosta, S. Balancing Susceptibility to Infection and Autoimmunity: A Role for Leptin? Trends Immunol. 2002, 23, 182–187. [Google Scholar] [CrossRef]
- Lee, S.; Lee, H.-C.; Kwon, Y.-W.; Lee, S.E.; Cho, Y.; Kim, J.; Lee, S.; Kim, J.-Y.; Lee, J.; Yang, H.-M.; et al. Adenylyl Cyclase-Associated Protein 1 Is a Receptor for Human Resistin and Mediates Inflammatory Actions of Human Monocytes. Cell Metab. 2014, 19, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.S.; Le, Y.Y. Role of Resistin in Inflammation and Inflammation-Related Diseases. Cell Mol. Immunol. 2006, 3, 29–34. [Google Scholar] [PubMed]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. Towards an Integrative Approach to Understanding the Role of Chemerin in Human Health and Disease. Obes. Rev. 2013, 14, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Moraes-Vieira, P.M.; Yore, M.M.; Dwyer, P.M.; Syed, I.; Aryal, P.; Kahn, B.B. RBP4 Activates Antigen-Presenting Cells, Leading to Adipose Tissue Inflammation and Systemic Insulin Resistance. Cell Metab. 2014, 19, 512–526. [Google Scholar] [CrossRef]
- Wang, H.-H.; Wu, M.-M.; Chan, M.W.Y.; Pu, Y.-S.; Chen, C.-J.; Lee, T.-C. Long-Term Low-Dose Exposure of Human Urothelial Cells to Sodium Arsenite Activates Lipocalin-2 via Promoter Hypomethylation. Arch. Toxicol. 2014, 88, 1549–1559. [Google Scholar] [CrossRef]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of Anti-Inflammatory Adipokines in Obesity-Related Diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef]
- Tan, Y.-L.; Zheng, X.-L.; Tang, C.-K. The Protective Functions of Omentin in Cardiovascular Diseases. Clin. Chim. Acta 2015, 448, 98–106. [Google Scholar] [CrossRef]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the Past Two Decades. J. Mol. Cell Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Walsh, K. Obesity, Adiponectin and Vascular Inflammatory Disease. Curr. Opin. Lipidol. 2003, 14, 561–566. [Google Scholar] [CrossRef]
- Yoshida, S.; Fuster, J.J.; Walsh, K. Adiponectin Attenuates Abdominal Aortic Aneurysm Formation in Hyperlipidemic Mice. Atherosclerosis 2014, 235, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Schindler, M.; Pendzialek, M.; Grybel, K.J.; Seeling, T.; Gürke, J.; Fischer, B.; Navarrete Santos, A. Adiponectin Stimulates Lipid Metabolism via AMPK in Rabbit Blastocysts. Hum. Reprod. 2017, 32, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Peterson, J.M.; Wong, G.W. Metabolic Regulation by C1q/TNF-Related Protein-13 (CTRP13): Activation OF AMP-Activated Protein Kinase and Suppression of Fatty Acid-Induced JNK Signaling. J. Biol. Chem. 2011, 286, 15652–15665. [Google Scholar] [CrossRef]
- Kopp, A.; Bala, M.; Buechler, C.; Falk, W.; Gross, P.; Neumeier, M.; Schölmerich, J.; Schäffler, A. C1q/TNF-Related Protein-3 Represents a Novel and Endogenous Lipopolysaccharide Antagonist of the Adipose Tissue. Endocrinology 2010, 151, 5267–5278. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Sano, S.; Fuster, J.J.; Kikuchi, R.; Shimizu, I.; Ohshima, K.; Katanasaka, Y.; Ouchi, N.; Walsh, K. Secreted Frizzled-Related Protein 5 Diminishes Cardiac Inflammation and Protects the Heart from Ischemia/Reperfusion Injury. J. Biol. Chem. 2016, 291, 2566–2575. [Google Scholar] [CrossRef]
- Matsuzawa, Y.; Funahashi, T.; Nakamura, T. The Concept of Metabolic Syndrome: Contribution of Visceral Fat Accumulation and Its Molecular Mechanism. J. Atheroscler. Thromb. 2011, 18, 629–639. [Google Scholar] [CrossRef]
- GBD 2019 Diseases and Injuries Collaborators Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [CrossRef]
- Kumada, M.; Kihara, S.; Sumitsuji, S.; Kawamoto, T.; Matsumoto, S.; Ouchi, N.; Arita, Y.; Okamoto, Y.; Shimomura, I.; Hiraoka, H.; et al. Association of Hypoadiponectinemia with Coronary Artery Disease in Men. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 85–89. [Google Scholar] [CrossRef]
- Ebrahimi-Mamaeghani, M.; Mohammadi, S.; Arefhosseini, S.R.; Fallah, P.; Bazi, Z. Adiponectin as a Potential Biomarker of Vascular Disease. Vasc. Health Risk Manag. 2015, 11, 55–70. [Google Scholar] [CrossRef]
- Mitsuhashi, H.; Yatsuya, H.; Tamakoshi, K.; Matsushita, K.; Otsuka, R.; Wada, K.; Sugiura, K.; Takefuji, S.; Hotta, Y.; Kondo, T.; et al. Adiponectin Level and Left Ventricular Hypertrophy in Japanese Men. Hypertension 2007, 49, 1448–1454. [Google Scholar] [CrossRef][Green Version]
- Imatoh, T.; Miyazaki, M.; Momose, Y.; Tanihara, S.; Une, H. Adiponectin Levels Associated with the Development of Hypertension: A Prospective Study. Hypertens. Res. 2008, 31, 229–233. [Google Scholar] [CrossRef]
- Leon, B.M.; Maddox, T.M. Diabetes and Cardiovascular Disease: Epidemiology, Biological Mechanisms, Treatment Recommendations and Future Research. World J. Diabetes 2015, 6, 1246–1258. [Google Scholar] [CrossRef]
- Yang, L.; Li, B.; Zhao, Y.; Zhang, Z. Prognostic Value of Adiponectin Level in Patients with Coronary Artery Disease: A Systematic Review and Meta-Analysis. Lipids Health Dis. 2019, 18, 227. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, L.; Wang, J. Adiponectin and the Risk of New-Onset Atrial Fibrillation: A Meta-Analysis of Prospective Cohort Studies. Biosci. Rep. 2019, 39, BSR20182284. [Google Scholar] [CrossRef]
- Bai, W.; Huang, J.; Zhu, M.; Liu, X.; Tao, J. Association between Elevated Adiponectin Level and Adverse Outcomes in Patients with Heart Failure: A Systematic Review and Meta-Analysis. Braz. J. Med. Biol. Res. 2019, 52, e8416. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.-P.; Sun, L.; Li, X.-H.; Che, F.-G.; Zhu, X.-Q.; Yang, F.; Han, J.; Jia, C.-Y.; Yang, Z. Association of Adiponectin Polymorphism with Metabolic Syndrome Risk and Adiponectin Level with Stroke Risk: A Meta-Analysis. Sci. Rep. 2016, 6, 31945. [Google Scholar] [CrossRef]
- Arregui, M.; Buijsse, B.; Fritsche, A.; di Giuseppe, R.; Schulze, M.B.; Westphal, S.; Isermann, B.; Boeing, H.; Weikert, C. Adiponectin and Risk of Stroke: Prospective Study and Meta-Analysis. Stroke 2014, 45, 10–17. [Google Scholar] [CrossRef]
- Farooqi, I.S.; O’Rahilly, S. Leptin: A Pivotal Regulator of Human Energy Homeostasis. Am. J. Clin. Nutr. 2009, 89, 980S–984S. [Google Scholar] [CrossRef]
- Blum, W.F.; Englaro, P.; Hanitsch, S.; Juul, A.; Hertel, N.T.; Müller, J.; Skakkebaek, N.E.; Heiman, M.L.; Birkett, M.; Attanasio, A.M.; et al. Plasma Leptin Levels in Healthy Children and Adolescents: Dependence on Body Mass Index, Body Fat Mass, Gender, Pubertal Stage, and Testosterone. J. Clin. Endocrinol. Metab. 1997, 82, 2904–2910. [Google Scholar] [CrossRef] [PubMed]
- Liberale, L.; Bonaventura, A.; Vecchiè, A.; Casula, M.; Dallegri, F.; Montecucco, F.; Carbone, F. The Role of Adipocytokines in Coronary Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 10. [Google Scholar] [CrossRef]
- Khafaji, H.A.R.H.; Bener, A.B.; Rizk, N.M.; Al Suwaidi, J. Elevated Serum Leptin Levels in Patients with Acute Myocardial Infarction; Correlation with Coronary Angiographic and Echocardiographic Findings. BMC Res. Notes 2012, 5, 262. [Google Scholar] [CrossRef] [PubMed]
- Montazerifar, F.; Bolouri, A.; Paghalea, R.S.; Mahani, M.K.; Karajibani, M. Obesity, Serum Resistin and Leptin Levels Linked to Coronary Artery Disease. Arq. Bras. Cardiol. 2016, 107, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Xu, C.-H.; Xu, Y.-N.; Wang, Y.-L.; Wang, M. Association of Leptin Levels with Pathogenetic Risk of Coronary Heart Disease and Stroke: A Meta-Analysis. Arq. Bras. Endocrinol. Metabol. 2014, 58, 817–823. [Google Scholar] [CrossRef]
- Bickel, C.; Schnabel, R.B.; Zeller, T.; Lackner, K.J.; Rupprecht, H.J.; Blankenberg, S.; Sinning, C.; Westermann, D. Predictors of Leptin Concentration and Association with Cardiovascular Risk in Patients with Coronary Artery Disease: Results from the AtheroGene Study. Biomarkers 2017, 22, 210–218. [Google Scholar] [CrossRef]
- Bigalke, B.; Stellos, K.; Geisler, T.; Seizer, P.; Mozes, V.; Gawaz, M. High Plasma Levels of Adipocytokines Are Associated with Platelet Activation in Patients with Coronary Artery Disease. Platelets 2010, 21, 11–19. [Google Scholar] [CrossRef]
- Cirillo, P.; Angri, V.; De Rosa, S.; Calì, G.; Petrillo, G.; Maresca, F.; D’Ascoli, G.-L.; Maietta, P.; Brevetti, L.; Chiariello, M. Pro-Atherothrombotic Effects of Leptin in Human Coronary Endothelial Cells. Thromb. Haemost. 2010, 103, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Guo, W.; Li, J.; Cao, S.; Zhang, J.; Pan, J.; Wang, Z.; Wen, P.; Shi, X.; Zhang, S. Leptin Concentration and Risk of Coronary Heart Disease and Stroke: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0166360. [Google Scholar] [CrossRef]
- Simiti, L.A.V.; Todor, I.; Stoia, M.A.; Goidescu, C.M.; Anton, F.P.; Farcas, A.D. Better Prognosis in Overweight/Obese Coronary Heart Disease Patients with High Plasma Levels of Leptin. Clujul Med. 2016, 89, 65–71. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Curran, J.E.; Stocker, C.J.; Zaibi, M.S.; Segal, D.; Konstantopoulos, N.; Morrison, S.; Carless, M.; Dyer, T.D.; Cole, S.A.; et al. Chemerin, a Novel Adipokine in the Regulation of Angiogenesis. J. Clin. Endocrinol. Metab. 2010, 95, 2476–2485. [Google Scholar] [CrossRef]
- Aksan, G.; İnci, S.; Nar, G.; Soylu, K.; Gedikli, Ö.; Yüksel, S.; Özdemir, M.; Nar, R.; Meriç, M.; Şahin, M. Association of Serum Chemerin Levels with the Severity of Coronary Artery Disease in Patients with Metabolic Syndrome. Int. J. Clin. Exp. Med. 2014, 7, 5461–5468. [Google Scholar]
- Gu, P.; Cheng, M.; Hui, X.; Lu, B.; Jiang, W.; Shi, Z. Elevating Circulation Chemerin Level Is Associated with Endothelial Dysfunction and Early Atherosclerotic Changes in Essential Hypertensive Patients. J. Hypertens. 2015, 33, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, O.; Ji, Q.; Lin, Y.; Wang, Z.; Huang, Y.; Lu, W.; Liu, X.; Zhang, J.; Liu, Y.; Zhou, Y. Circulating Chemerin Levels Elevated in Dilated Cardiomyopathy Patients with Overt Heart Failure. Clin. Chim. Acta 2015, 448, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Venteclef, N.; Guglielmi, V.; Balse, E.; Gaborit, B.; Cotillard, A.; Atassi, F.; Amour, J.; Leprince, P.; Dutour, A.; Clément, K.; et al. Human Epicardial Adipose Tissue Induces Fibrosis of the Atrial Myocardium through the Secretion of Adipo-Fibrokines. Eur. Heart J. 2015, 36, 795–805. [Google Scholar] [CrossRef]
- Codoñer-Franch, P.; Alonso-Iglesias, E. Resistin: Insulin Resistance to Malignancy. Clin. Chim. Acta 2015, 438, 46–54. [Google Scholar] [CrossRef]
- Henning, R.J. Obesity and Obesity-Induced Inflammatory Disease Contribute to Atherosclerosis: A Review of the Pathophysiology and Treatment of Obesity. Am. J. Cardiovasc. Dis. 2021, 11, 504–529. [Google Scholar]
- Muse, E.D.; Feldman, D.I.; Blaha, M.J.; Dardari, Z.A.; Blumenthal, R.S.; Budoff, M.J.; Nasir, K.; Criqui, M.H.; Cushman, M.; McClelland, R.L.; et al. The Association of Resistin with Cardiovascular Disease in the Multi-Ethnic Study of Atherosclerosis. Atherosclerosis 2015, 239, 101–108. [Google Scholar] [CrossRef]
- Gao, W.; Wang, H.; Zhang, L.; Cao, Y.; Bao, J.-Z.; Liu, Z.-X.; Wang, L.-S.; Yang, Q.; Lu, X. Retinol-Binding Protein 4 Induces Cardiomyocyte Hypertrophy by Activating TLR4/MyD88 Pathway. Endocrinology 2016, 157, 2282–2293. [Google Scholar] [CrossRef] [PubMed]
- Chavarria, N.; Kato, T.S.; Khan, R.; Chokshi, A.; Collado, E.; Akashi, H.; Takayama, H.; Naka, Y.; Farr, M.; Mancini, D.; et al. Increased Levels of Retinol Binding Protein 4 in Patients with Advanced Heart Failure Correct after Hemodynamic Improvement through Ventricular Assist Device Placement. Circ. J. 2012, 76, 2148–2152. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Z.; Zhang, K.-Z.; Yan, J.-J.; Wang, L.; Wang, Y.; Shen, X.-Y.; Sun, H.-X.; Liu, L.; Zhao, C.; He, H.-W.; et al. Serum Retinol-Binding Protein 4 as a Predictor of Cardiovascular Events in Elderly Patients with Chronic Heart Failure. ESC Heart Fail. 2020, 7, 542–550. [Google Scholar] [CrossRef]
- von Jeinsen, B.; Short, M.I.; Xanthakis, V.; Carneiro, H.; Cheng, S.; Mitchell, G.F.; Vasan, R.S. Association of Circulating Adipokines with Echocardiographic Measures of Cardiac Structure and Function in a Community-Based Cohort. J. Am. Heart Assoc. 2018, 7, e008997. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, Y.; Yidilisi, A.; Xu, Y.; Dong, Q.; Jiang, J. Causal Associations Between Circulating Adipokines and Cardiovascular Disease: A Mendelian Randomization Study. J. Clin. Endocrinol. Metab. 2022, 107, e2572–e2580. [Google Scholar] [CrossRef] [PubMed]
- Björkegren, J.L.M.; Lusis, A.J. Atherosclerosis: Recent Developments. Cell 2022, 185, 1630–1645. [Google Scholar] [CrossRef] [PubMed]
- Yuxiang, L.; Fujiu, K. Human Resistin and Cardiovascular Disease. Int. Heart J. 2020, 61, 421–423. [Google Scholar] [CrossRef]
- Eichelmann, F.; Schulze, M.B.; Wittenbecher, C.; Menzel, J.; Weikert, C.; di Giuseppe, R.; Biemann, R.; Isermann, B.; Fritsche, A.; Boeing, H.; et al. Chemerin as a Biomarker Linking Inflammation and Cardiovascular Diseases. J. Am. Coll. Cardiol. 2019, 73, 378–379. [Google Scholar] [CrossRef]
- Kim, D.H.; Vanella, L.; Inoue, K.; Burgess, A.; Gotlinger, K.; Manthati, V.L.; Koduru, S.R.; Zeldin, D.C.; Falck, J.R.; Schwartzman, M.L.; et al. Epoxyeicosatrienoic Acid Agonist Regulates Human Mesenchymal Stem Cell-Derived Adipocytes through Activation of HO-1-PAKT Signaling and a Decrease in PPARγ. Stem Cells Dev. 2010, 19, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, R.G.; Masui, Y.; Fang, F.; Korman, B.; Lord, G.; Lee, J.; Lakota, K.; Wei, J.; Scherer, P.E.; Otvos, L.; et al. Adiponectin Is an Endogenous Anti-Fibrotic Mediator and Therapeutic Target. Sci. Rep. 2017, 7, 4397. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Takeda, Y.; Nakanishi, K.; Tachibana, I.; Kumanogoh, A. Adiponectin: A Novel Link between Adipocytes and COPD. Vitam. Horm. 2012, 90, 419–435. [Google Scholar] [CrossRef]
- Fruebis, J.; Tsao, T.S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic Cleavage Product of 30-KDa Adipocyte Complement-Related Protein Increases Fatty Acid Oxidation in Muscle and Causes Weight Loss in Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 2005–2010. [Google Scholar] [CrossRef]
- Martin, L.J.; Woo, J.G.; Daniels, S.R.; Goodman, E.; Dolan, L.M. The Relationships of Adiponectin with Insulin and Lipids Are Strengthened with Increasing Adiposity. J. Clin. Endocrinol. Metab. 2005, 90, 4255–4259. [Google Scholar] [CrossRef]
- Hasan-Ali, H.; Abd El-Mottaleb, N.A.; Hamed, H.B.; Abd-Elsayed, A. Serum Adiponectin and Leptin as Predictors of the Presence and Degree of Coronary Atherosclerosis. Coron. Artery Dis. 2011, 22, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Katsiki, N.; Mikhailidis, D.P.; Banach, M. Leptin, Cardiovascular Diseases and Type 2 Diabetes Mellitus. Acta Pharmacol. Sin. 2018, 39, 1176–1188. [Google Scholar] [CrossRef]
- Payne, G.A.; Tune, J.D.; Knudson, J.D. Leptin-Induced Endothelial Dysfunction: A Target for Therapeutic Interventions. Curr. Pharm. Des. 2014, 20, 603–608. [Google Scholar] [CrossRef]
- Leptin and Endothelial Function in the Elderly: The Prospective Investigation of the Vasculature in Uppsala Seniors (PIVUS) Study—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/23591414/ (accessed on 16 March 2023).
- Fang, C.; Lei, J.; Zhou, S.; Zhang, Y.; Yuan, G.; Wang, J. Association of Higher Resistin Levels with Inflammatory Activation and Endothelial Dysfunction in Patients with Essential Hypertension. Chin. Med. J. 2013, 126, 646–649. [Google Scholar] [PubMed]
- Liang, W.; Ye, D.D. The Potential of Adipokines as Biomarkers and Therapeutic Agents for Vascular Complications in Type 2 Diabetes Mellitus. Cytokine Growth Factor Rev. 2019, 48, 32–39. [Google Scholar] [CrossRef]
- Joksic, J.; Sopic, M.; Spasojevic-Kalimanovska, V.; Kalimanovska-Ostric, D.; Andjelkovic, K.; Jelic-Ivanovic, Z. Circulating Resistin Protein and MRNA Concentrations and Clinical Severity of Coronary Artery Disease. Biochem. Med. 2015, 25, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Jiang, J.; Lü, J.-M.; Chai, H.; Wang, X.; Lin, P.H.; Yao, Q. Resistin Decreases Expression of Endothelial Nitric Oxide Synthase through Oxidative Stress in Human Coronary Artery Endothelial Cells. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H193–H201. [Google Scholar] [CrossRef]
- Yoo, H.J.; Choi, H.Y.; Yang, S.J.; Kim, H.Y.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Choi, K.M.; Choi, D.S.; Baik, S.H. Circulating Chemerin Level Is Independently Correlated with Arterial Stiffness. J. Atheroscler. Thromb. 2012, 19, 59–66, discussion 67–68. [Google Scholar] [CrossRef]
- Landgraf, K.; Friebe, D.; Ullrich, T.; Kratzsch, J.; Dittrich, K.; Herberth, G.; Adams, V.; Kiess, W.; Erbs, S.; Körner, A. Chemerin as a Mediator between Obesity and Vascular Inflammation in Children. J. Clin. Endocrinol. Metab. 2012, 97, E556–E564. [Google Scholar] [CrossRef]
- Fatima, S.S.; Rehman, R.; Baig, M.; Khan, T.A. New Roles of the Multidimensional Adipokine: Chemerin. Peptides 2014, 62, 15–20. [Google Scholar] [CrossRef]
- Zylla, S.; Dörr, M.; Völzke, H.; Schminke, U.; Felix, S.B.; Nauck, M.; Friedrich, N. Association of Circulating Chemerin with Subclinical Parameters of Atherosclerosis: Results of a Population-Based Study. Arterioscler. Thromb Vasc. Biol. 2018, 38, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ye, S.-D.; Wang, W. Elevated Retinol Binding Protein 4 Levels Are Associated with Atherosclerosis in Diabetic Rats via JAK2/STAT3 Signaling Pathway. World J. Diabetes 2021, 12, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhong, Y.; Chen, H.; Wang, D.; Wang, M.; Ou, J.-S.; Xia, M. Retinol-Binding Protein-Dependent Cholesterol Uptake Regulates Macrophage Foam Cell Formation and Promotes Atherosclerosis. Circulation 2017, 135, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Setayesh, L.; Ebrahimi, R.; Pooyan, S.; Yarizadeh, H.; Rashidbeygi, E.; Badrooj, N.; Imani, H.; Mirzaei, K. The Possible Mediatory Role of Adipokines in the Association between Low Carbohydrate Diet and Depressive Symptoms among Overweight and Obese Women. PLoS ONE 2021, 16, e0257275. [Google Scholar] [CrossRef] [PubMed]
- Ouerghi, N.; Ben Fradj, M.K.; Talbi, E.; Bezrati, I.; Feki, M.; Bouassida, A. Association of Selected Adipokines with Metabolic Syndrome and Cardio-Metabolic Risk Factors in Young Males. Cytokine 2020, 133, 155170. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Lee, D.K.; Liu, M.; Portincasa, P.; Wang, D.Q.-H. Novel Insights into the Pathogenesis and Management of the Metabolic Syndrome. Pediatr. Gastroenterol. Hepatol. Nutr. 2020, 23, 189–230. [Google Scholar] [CrossRef]
- Prasun, P. Mitochondrial Dysfunction in Metabolic Syndrome. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165838. [Google Scholar] [CrossRef]
- Ventriglio, A.; Baldessarini, R.J.; Vitrani, G.; Bonfitto, I.; Cecere, A.C.; Rinaldi, A.; Petito, A.; Bellomo, A. Metabolic Syndrome in Psychotic Disorder Patients Treated with Oral and Long-Acting Injected Antipsychotics. Front. Psychiatry 2018, 9, 744. [Google Scholar] [CrossRef]
- Garrido-Torres, N.; Rocha-Gonzalez, I.; Alameda, L.; Rodriguez-Gangoso, A.; Vilches, A.; Canal-Rivero, M.; Crespo-Facorro, B.; Ruiz-Veguilla, M. Metabolic Syndrome in Antipsychotic-Naïve Patients with First-Episode Psychosis: A Systematic Review and Meta-Analysis. Psychol. Med. 2021, 51, 2307–2320. [Google Scholar] [CrossRef]
- Dmitrzak-Weglarz, M.; Tyszkiewicz-Nwafor, M.; Rybakowski, F.; Permoda-Pachuta, A. Changes in Adipokine Concentrations in Antidepressant-Resistant Bipolar Depression after Ketamine Infusion and Electroconvulsive Therapy. World J. Biol. Psychiatry 2022, 24, 267–279. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Bustamante-Sanchez, Á.; Tornero-Aguilera, J.F.; Ruisoto, P.; Mielgo-Ayuso, J. Inflammation in COVID-19 and the Effects of Non-Pharmacological Interventions during the Pandemic: A Review. Int. J. Mol. Sci. 2022, 23, 15584. [Google Scholar] [CrossRef]
- Carli, M.; Kolachalam, S.; Longoni, B.; Pintaudi, A.; Baldini, M.; Aringhieri, S.; Fasciani, I.; Annibale, P.; Maggio, R.; Scarselli, M. Atypical Antipsychotics and Metabolic Syndrome: From Molecular Mechanisms to Clinical Differences. Pharmaceuticals 2021, 14, 238. [Google Scholar] [CrossRef] [PubMed]
- Raue, S.; Wedekind, D.; Wiltfang, J.; Schmidt, U. The Role of Proopiomelanocortin and α-Melanocyte-Stimulating Hormone in the Metabolic Syndrome in Psychiatric Disorders: A Narrative Mini-Review. Front. Psychiatry 2019, 10, 834. [Google Scholar] [CrossRef] [PubMed]
- Çakici, N.; Sutterland, A.L.; Penninx, B.W.J.H.; Dalm, V.A.; de Haan, L.; van Beveren, N.J.M. Altered Peripheral Blood Compounds in Drug-Naïve First-Episode Patients with Either Schizophrenia or Major Depressive Disorder: A Meta-Analysis. Brain Behav. Immun. 2020, 88, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Painold, A.; Mörkl, S.; Kashofer, K.; Halwachs, B.; Dalkner, N.; Bengesser, S.; Birner, A.; Fellendorf, F.; Platzer, M.; Queissner, R.; et al. A Step Ahead: Exploring the Gut Microbiota in Inpatients with Bipolar Disorder during a Depressive Episode. Bipolar Disord. 2019, 21, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Dobrowolny, H.; Guest, P.C.; Bernstein, H.-G.; Fuchs, D.; Roeser, J.; Summergrad, P.; Oxenkrug, G.F. Plasma Anthranilic Acid and Leptin Levels Predict HAM-D Scores in Depressed Women. Int. J. Tryptophan Res. 2021, 14, 11786469211016474. [Google Scholar] [CrossRef]
- Bhatt, S.; Nagappa, A.N.; Patil, C.R. Role of Oxidative Stress in Depression. Drug Discov. Today 2020, 25, 1270–1276. [Google Scholar] [CrossRef]
- da Silva Souza, S.V.; da Rosa, P.B.; Neis, V.B.; Moreira, J.D.; Rodrigues, A.L.S.; Moretti, M. Effects of Cholecalciferol on Behavior and Production of Reactive Oxygen Species in Female Mice Subjected to Corticosterone-Induced Model of Depression. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 111–120. [Google Scholar] [CrossRef]
- The Features of Psychological State, Eating Behavior, Hormonal and Adipokine Regulation of Metabolism in Men with Subcutaneous and Abdominal Fat Distribution|Mokhova|Obesity and Metabolism. Available online: https://www.omet-endojournals.ru/jour/article/view/12100/en_US?locale=en_US (accessed on 16 March 2023).
- Extreme Eating Behaviours|Frontiers Research Topic. Available online: https://www.frontiersin.org/research-topics/9836/extreme-eating-behaviours (accessed on 16 March 2023).
- Arouca, A. Interplay of Diet with Adiposity, Sedentary Behavior, and Genetic Polymorphisms towards Inflammation in Adolescents. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2021. [Google Scholar]
- Himmerich, H.; Dalton, B.; Patsalos, O.; Schmidt, U.; Campbell, I.C. Cytokines and Water Distribution in Anorexia Nervosa. Mediat. Inflamm. 2021, 2021, 8811051. [Google Scholar] [CrossRef]
- Zhou, J.C.; Rifas-Shiman, S.L.; Haines, J.; Jones, K.; Oken, E. Adolescent Overeating and Binge Eating Behavior in Relation to Subsequent Cardiometabolic Risk Outcomes: A Prospective Cohort Study. J. Eat. Disord. 2022, 10, 140. [Google Scholar] [CrossRef]
- Westfall, S.; Caracci, F.; Zhao, D.; Wu, Q.-L.; Frolinger, T.; Simon, J.; Pasinetti, G.M. Microbiota Metabolites Modulate the T Helper 17 to Regulatory T Cell (Th17/Treg) Imbalance Promoting Resilience to Stress-Induced Anxiety- and Depressive-like Behaviors. Brain Behav. Immun. 2021, 91, 350–368. [Google Scholar] [CrossRef] [PubMed]
- Solmi, M.; Suresh Sharma, M.; Osimo, E.F.; Fornaro, M.; Bortolato, B.; Croatto, G.; Miola, A.; Vieta, E.; Pariante, C.M.; Smith, L.; et al. Peripheral Levels of C-Reactive Protein, Tumor Necrosis Factor-α, Interleukin-6, and Interleukin-1β across the Mood Spectrum in Bipolar Disorder: A Meta-Analysis of Mean Differences and Variability. Brain Behav. Immun. 2021, 97, 193–203. [Google Scholar] [CrossRef] [PubMed]
- do Rosário Caldas, N.; Braulio, V.B.; Brasil, M.A.A.; Furtado, V.C.S.; de Carvalho, D.P.; Cotrik, E.M.; Dantas, J.R.; Zajdenverg, L. Binge Eating Disorder, Frequency of Depression, and Systemic Inflammatory State in Individuals with Obesity—A Cross Sectional Study. Arch. Endocrinol. Metab. 2022, 66, 489–497. [Google Scholar] [CrossRef]
- Wang, H.; Ren, X.; Tan, H.; Wang, Y. Influence of A Recombinant Human Soluble Tumor Necrosis Factor Receptor FC Fusion Protein on ACR Scale, C-Reactive Protein, Interleukin-6 And Erythrocyte Sedimentation Rate in Children with Juvenile Idiopathic Arthritis. Rev. Argent. Clín. Psicol. 2020, 29, 446–453. [Google Scholar] [CrossRef]
- Hussein, B.J.; Issa, I.H.; Khaleel, A.M.; Al-Dahhan, N.A.A. Salivary Levels of Interleukin-1beta, Tumour Necrosis Factor-α, and C-Reactive Proteins in Smokers Patients with Severe Chronic Periodontitis. Syst. Rev. Pharm. 2021, 12, 1804–1809. [Google Scholar]
- Beaudry, J.L.; Kaur, K.D.; Varin, E.M.; Baggio, L.L.; Cao, X.; Mulvihill, E.E.; Stern, J.H.; Campbell, J.E.; Scherer, P.E.; Drucker, D.J. The Brown Adipose Tissue Glucagon Receptor Is Functional but Not Essential for Control of Energy Homeostasis in Mice. Mol. Metab. 2019, 22, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Biglari, N.; Gaziano, I.; Schumacher, J.; Radermacher, J.; Paeger, L.; Klemm, P.; Chen, W.; Corneliussen, S.; Wunderlich, C.M.; Sue, M.; et al. Functionally Distinct POMC-Expressing Neuron Subpopulations in Hypothalamus Revealed by Intersectional Targeting. Nat. Neurosci. 2021, 24, 913–929. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, I.; Fricker, L.D. Obesity, POMC, and POMC-Processing Enzymes: Surprising Results from Animal Models. Endocrinology 2021, 162, bqab155. [Google Scholar] [CrossRef]
- Rau, A.R.; Hentges, S.T. GABAergic Inputs to POMC Neurons Originating from the Dorsomedial Hypothalamus Are Regulated by Energy State. J. Neurosci. 2019, 39, 6449–6459. [Google Scholar] [CrossRef]
- Kim, J.; Lee, N.; Suh, S.B.; Jang, S.; Kim, S.; Kim, D.-G.; Park, J.K.; Lee, K.-W.; Choi, S.Y.; Lee, C.H. Metformin Ameliorates Olanzapine-Induced Disturbances in POMC Neuron Number, Axonal Projection, and Hypothalamic Leptin Resistance. BMB Rep. 2022, 55, 293–298. [Google Scholar] [CrossRef]
- Jais, A.; Brüning, J.C. Arcuate Nucleus-Dependent Regulation of Metabolism-Pathways to Obesity and Diabetes Mellitus. Endocr. Rev. 2022, 43, 314–328. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Rauf, N.; Nabi, G.; Yi, S.; Yu-Dong, Z.; Fu, J. Mechanistic Insight into High-Fat Diet-Induced Metabolic Inflammation in the Arcuate Nucleus of the Hypothalamus. Biomed. Pharmacother. 2021, 142, 112012. [Google Scholar] [CrossRef]
- Carreira-Míguez, M.; Ramos-Campo, D.J.; Clemente-Suárez, V.J. Differences in Nutritional and Psychological Habits in Hypertension Patients. BioMed Res. Int. 2022, 2022, e1920996. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J. Multidisciplinary Intervention in the Treatment of Mixed Anxiety and Depression Disorder. Physiol. Behav. 2020, 219, 112858. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose Tissue Inflammation and Metabolic Dysfunction in Obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Kim, J.-S.; Jo, M.-J.; Cho, E.; Ahn, S.-Y.; Kwon, Y.-J.; Ko, G.-J. The Roles and Associated Mechanisms of Adipokines in Development of Metabolic Syndrome. Molecules 2022, 27, 334. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhai, Y.; Zhao, J.; He, H.; Li, Y.; Liu, Y.; Feng, A.; Li, L.; Huang, T.; Xu, A.; et al. Impact of Metabolic Syndrome and it’s Components on Prognosis in Patients with Cardiovascular Diseases: A Meta-Analysis. Front. Cardiovasc. Med. 2021, 8, 704145. [Google Scholar] [CrossRef]
- Kahn, C.R.; Wang, G.; Lee, K.Y. Altered Adipose Tissue and Adipocyte Function in the Pathogenesis of Metabolic Syndrome. J Clin. Investig. 2019, 129, 3990–4000. [Google Scholar] [CrossRef]
- Szydło, B.; Kiczmer, P.; Świętochowska, E.; Ostrowska, Z. Role of Omentin and Chemerin in Metabolic Syndrome and Tumor Diseases. Postepy Hig. Med. Dosw. 2016, 70, 844–849. [Google Scholar] [CrossRef]
- Sun, X.; Harris, E.N. New Aspects of Hepatic Endothelial Cells in Physiology and Nonalcoholic Fatty Liver Disease. Am. J. Physiol. Cell Physiol. 2020, 318, C1200–C1213. [Google Scholar] [CrossRef]
- Azzu, V.; Vacca, M.; Virtue, S.; Allison, M.; Vidal-Puig, A. Adipose Tissue-Liver Cross Talk in the Control of Whole-Body Metabolism: Implications in Nonalcoholic Fatty Liver Disease. Gastroenterology 2020, 158, 1899–1912. [Google Scholar] [CrossRef]
- Renehan, A.G.; Roberts, D.L.; Dive, C. Obesity and Cancer: Pathophysiological and Biological Mechanisms. Arch. Physiol. Biochem. 2008, 114, 71–83. [Google Scholar] [CrossRef]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-Reducing Effects of the Plasma Protein Encoded by the Obese Gene. Science 1995, 269, 543–546. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Baskin, D.G.; Bukowski, T.R.; Kuijper, J.L.; Foster, D.; Lasser, G.; Prunkard, D.E.; Porte, D.; Woods, S.C.; Seeley, R.J.; et al. Specificity of Leptin Action on Elevated Blood Glucose Levels and Hypothalamic Neuropeptide Y Gene Expression in Ob/Ob Mice. Diabetes 1996, 45, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Tanida, M.; Niijima, A.; Nagai, K. In Vivo Effects of Leptin on Autonomic Nerve Activity and Lipolysis in Rats. Neurosci. Lett. 2007, 416, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Shin, D. Prospective Associations of Serum Adiponectin, Leptin, and Leptin-Adiponectin Ratio with Incidence of Metabolic Syndrome: The Korean Genome and Epidemiology Study. Int. J. Environ. Res. Public Health 2020, 17, 3287. [Google Scholar] [CrossRef]
- McNeely, M.J.; Boyko, E.J.; Weigle, D.S.; Shofer, J.B.; Chessler, S.D.; Leonnetti, D.L.; Fujimoto, W.Y. Association between Baseline Plasma Leptin Levels and Subsequent Development of Diabetes in Japanese Americans. Diabetes Care 1999, 22, 65–70. [Google Scholar] [CrossRef]
- de Luis, D.A.; Izaola, O.; Primo, D.; Aller, R. Relation of a Variant in Adiponectin Gene (Rs266729) with Metabolic Syndrome and Diabetes Mellitus Type 2 in Adult Obese Subjects. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10646–10652. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xu, Y.; Zhang, Y.; Ji, T.; Li, Y. Lower Levels of Circulating Adiponectin in Elderly Patients with Metabolic Inflammatory Syndrome: A Cross-Sectional Study. Diabetes Metab. Syndr. Obes. 2020, 13, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Silha, J.V.; Krsek, M.; Skrha, J.V.; Sucharda, P.; Nyomba, B.L.G.; Murphy, L.J. Plasma Resistin, Adiponectin and Leptin Levels in Lean and Obese Subjects: Correlations with Insulin Resistance. Eur. J. Endocrinol. 2003, 149, 331–335. [Google Scholar] [CrossRef]
- Shim, Y.S.; Kang, M.J.; Oh, Y.J.; Baek, J.W.; Yang, S.; Hwang, I.T. Fetuin-A as an Alternative Marker for Insulin Resistance and Cardiovascular Risk in Prepubertal Children. J. Atheroscler. Thromb 2017, 24, 1031–1038. [Google Scholar] [CrossRef]
- Assessment of Plasminogen Activator Inhibitor-1 in Obese Egyptian Children|Egyptian Pediatric Association Gazette|Full Text. Available online: https://epag.springeropen.com/articles/10.1186/s43054-019-0012-8 (accessed on 5 February 2023).
- Currò, D.; Vergani, E.; Bruno, C.; Comi, S.; D’Abate, C.; Mancini, A. Plasmatic Lipocalin-2 Levels in Chronic Low-Grade Inflammation Syndromes: Comparison between Metabolic Syndrome, Total and Partial Adult Growth Hormone Deficiency. Biofactors 2020, 46, 629–636. [Google Scholar] [CrossRef]
- de Luis, D.A.; Aller, R.; Gonzalez Sagrado, M.; Conde, R.; Izaola, O.; de la Fuente, B. Serum Visfatin Levels and Metabolic Syndrome Criteria in Obese Female Subjects. Diabetes Metab. Res. Rev. 2013, 29, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Peng, D. Adipokines as Novel Biomarkers of Cardio-Metabolic Disorders. Clin. Chim. Acta 2020, 507, 31–38. [Google Scholar] [CrossRef]
- Auguet, T.; Quintero, Y.; Riesco, D.; Morancho, B.; Terra, X.; Crescenti, A.; Broch, M.; Aguilar, C.; Olona, M.; Porras, J.A.; et al. New Adipokines Vaspin and Omentin. Circulating Levels and Gene Expression in Adipose Tissue from Morbidly Obese Women. BMC Med. Genet. 2011, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qu, H.; Xiong, X.; Qiu, Y.; Liao, Y.; Chen, Y.; Zheng, Y.; Zheng, H. Plasma Asprosin Concentrations Are Increased in Individuals with Glucose Dysregulation and Correlated with Insulin Resistance and First-Phase Insulin Secretion. Mediat. Inflamm. 2018, 2018, 9471583. [Google Scholar] [CrossRef]
- Tian, Q.-P.; Liu, M.-L.; Tang, C.-S.; Xue, L.; Pang, Y.-Z.; Qi, Y.-F. Association of Circulating Neuregulin-4 with Presence and Severity of Coronary Artery Disease. Int. Heart J. 2019, 60, 45–49. [Google Scholar] [CrossRef]
- Smekal, A.; Vaclavik, J. Adipokines and Cardiovascular Disease: A Comprehensive Review. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech Repub. 2017, 161, 31–40. [Google Scholar] [CrossRef]
- Mechanick, J.I.; Zhao, S.; Garvey, W.T. The Adipokine-Cardiovascular-Lifestyle Network: Translation to Clinical Practice. J. Am. Coll. Cardiol. 2016, 68, 1785–1803. [Google Scholar] [CrossRef] [PubMed]
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 16 March 2023).
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. International Agency for Research on Cancer Handbook Working Group Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef]
- Yoon, Y.S.; Kwon, A.R.; Lee, Y.K.; Oh, S.W. Circulating Adipokines and Risk of Obesity Related Cancers: A Systematic Review and Meta-Analysis. Obes. Res. Clin. Pract. 2019, 13, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.E.; Kim, J.M.; Joung, K.H.; Lee, J.H.; You, B.R.; Choi, M.J.; Ryu, M.J.; Ko, Y.B.; Lee, M.A.; Lee, J.; et al. The Roles of Adipokines, Proinflammatory Cytokines, and Adipose Tissue Macrophages in Obesity-Associated Insulin Resistance in Modest Obesity and Early Metabolic Dysfunction. PLoS ONE 2016, 11, e0154003. [Google Scholar] [CrossRef]
- Doerstling, S.S.; O’Flanagan, C.H.; Hursting, S.D. Obesity and Cancer Metabolism: A Perspective on Interacting Tumor-Intrinsic and Extrinsic Factors. Front. Oncol. 2017, 7, 216. [Google Scholar] [CrossRef]
- Christodoulatos, G.S.; Spyrou, N.; Kadillari, J.; Psallida, S.; Dalamaga, M. The Role of Adipokines in Breast Cancer: Current Evidence and Perspectives. Curr. Obes. Rep. 2019, 8, 413–433. [Google Scholar] [CrossRef] [PubMed]
- García-Miranda, A.; Garcia-Hernandez, A.; Castañeda-Saucedo, E.; Navarro-Tito, N.; Maycotte, P. Adipokines as Regulators of Autophagy in Obesity-Linked Cancer. Cells 2022, 11, 3230. [Google Scholar] [CrossRef]
- Sánchez-Jiménez, F.; Pérez-Pérez, A.; de la Cruz-Merino, L.; Sánchez-Margalet, V. Obesity and Breast Cancer: Role of Leptin. Front. Oncol. 2019, 9, 596. [Google Scholar] [CrossRef]
- Harwood, H.J. The Adipocyte as an Endocrine Organ in the Regulation of Metabolic Homeostasis. Neuropharmacology 2012, 63, 57–75. [Google Scholar] [CrossRef]
- Zhang, H.-P.; Zou, J.; Xu, Z.-Q.; Ruan, J.; Yang, S.-D.; Yin, Y.; Mu, H.-J. Association of Leptin, Visfatin, Apelin, Resistin and Adiponectin with Clear Cell Renal Cell Carcinoma. Oncol. Lett. 2017, 13, 463–468. [Google Scholar] [CrossRef][Green Version]
- Pan, H.; Deng, L.-L.; Cui, J.-Q.; Shi, L.; Yang, Y.-C.; Luo, J.-H.; Qin, D.; Wang, L. Association between Serum Leptin Levels and Breast Cancer Risk: An Updated Systematic Review and Meta-Analysis. Medicine 2018, 97, e11345. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Pan, Q.; Chen, X.; Xu, S.; Luo, X.; Chen, L. The Association between Obesity Related Adipokines and Risk of Breast Cancer: A Meta-Analysis. Oncotarget 2017, 8, 75389–75399. [Google Scholar] [CrossRef] [PubMed]
- Harris, H.R.; Tworoger, S.S.; Hankinson, S.E.; Rosner, B.A.; Michels, K.B. Plasma Leptin Levels and Risk of Breast Cancer in Premenopausal Women. Cancer Prev. Res. 2011, 4, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Stattin, P.; Söderberg, S.; Hallmans, G.; Bylund, A.; Kaaks, R.; Stenman, U.H.; Bergh, A.; Olsson, T. Leptin Is Associated with Increased Prostate Cancer Risk: A Nested Case-Referent Study. J. Clin. Endocrinol. Metab. 2001, 86, 1341–1345. [Google Scholar] [CrossRef]
- Lee, C.H.; Woo, Y.C.; Wang, Y.; Yeung, C.Y.; Xu, A.; Lam, K.S.L. Obesity, Adipokines and Cancer: An Update. Clin. Endocrinol. 2015, 83, 147–156. [Google Scholar] [CrossRef]
- Ye, J.; Jia, J.; Dong, S.; Zhang, C.; Yu, S.; Li, L.; Mao, C.; Wang, D.; Chen, J.; Yuan, G. Circulating Adiponectin Levels and the Risk of Breast Cancer: A Meta-Analysis. Eur. J. Cancer Prev. 2014, 23, 158–165. [Google Scholar] [CrossRef]
- Erdogan, S.; Sezer, S.; Baser, E.; Gun-Eryilmaz, O.; Gungor, T.; Uysal, S.; Yilmaz, F.M. Evaluating Vaspin and Adiponectin in Postmenopausal Women with Endometrial Cancer. Endocr. Relat. Cancer 2013, 20, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Gonullu, G.; Kahraman, H.; Bedir, A.; Bektas, A.; Yücel, I. Association between Adiponectin, Resistin, Insulin Resistance, and Colorectal Tumors. Int. J. Colorectal. Dis. 2010, 25, 205–212. [Google Scholar] [CrossRef]
- Dalamaga, M. Resistin as a Biomarker Linking Obesity and Inflammation to Cancer: Potential Clinical Perspectives. Biomark. Med. 2014, 8, 107–118. [Google Scholar] [CrossRef]
- Park, H.K.; Kwak, M.K.; Kim, H.J.; Ahima, R.S. Linking Resistin, Inflammation, and Cardiometabolic Diseases. Korean J. Intern. Med. 2017, 32, 239–247. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Spyrou, N.; Avgerinos, K.I.; Mantzoros, C.S.; Dalamaga, M. Classic and Novel Adipocytokines at the Intersection of Obesity and Cancer: Diagnostic and Therapeutic Strategies. Curr. Obes. Rep. 2018, 7, 260–275. [Google Scholar] [CrossRef]
- Lehrke, M.; Reilly, M.P.; Millington, S.C.; Iqbal, N.; Rader, D.J.; Lazar, M.A. An Inflammatory Cascade Leading to Hyperresistinemia in Humans. PLoS Med. 2004, 1, e45. [Google Scholar] [CrossRef]
- Gong, W.-J.; Zheng, W.; Xiao, L.; Tan, L.-M.; Song, J.; Li, X.-P.; Xiao, D.; Cui, J.-J.; Li, X.; Zhou, H.-H.; et al. Circulating Resistin Levels and Obesity-Related Cancer Risk: A Meta-Analysis. Oncotarget 2016, 7, 57694–57704. [Google Scholar] [CrossRef]
- Reizes, O.; Berger, N.A. (Eds.) Adipocytokines, Energy Balance, and Cancer; Energy Balance and Cancer; Springer International Publishing: Cham, Switzerland, 2017; Volume 12, ISBN 978-3-319-41675-5. [Google Scholar]
- Lee, J.O.; Kim, N.; Lee, H.J.; Lee, Y.W.; Kim, S.J.; Park, S.H.; Kim, H.S. Resistin, a Fat-Derived Secretory Factor, Promotes Metastasis of MDA-MB-231 Human Breast Cancer Cells through ERM Activation. Sci. Rep. 2016, 6, 18923. [Google Scholar] [CrossRef] [PubMed]
- Danese, E.; Montagnana, M.; Minicozzi, A.M.; Bonafini, S.; Ruzzenente, O.; Gelati, M.; De Manzoni, G.; Lippi, G.; Guidi, G.C. The Role of Resistin in Colorectal Cancer. Clin. Chim. Acta 2012, 413, 760–764. [Google Scholar] [CrossRef]
- Mohammadi, M.; Mianabadi, F.; Mehrad-Majd, H. Circulating Visfatin Levels and Cancers Risk: A Systematic Review and Meta-Analysis. J. Cell Physiol. 2019, 234, 5011–5022. [Google Scholar] [CrossRef] [PubMed]
- Curat, C.A.; Wegner, V.; Sengenès, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumié, A. Macrophages in Human Visceral Adipose Tissue: Increased Accumulation in Obesity and a Source of Resistin and Visfatin. Diabetologia 2006, 49, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Gallí, M.; Van Gool, F.; Rongvaux, A.; Andris, F.; Leo, O. The Nicotinamide Phosphoribosyltransferase: A Molecular Link between Metabolism, Inflammation, and Cancer. Cancer Res. 2010, 70, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Yoshitake, N.; Fukui, H.; Yamagishi, H.; Sekikawa, A.; Fujii, S.; Tomita, S.; Ichikawa, K.; Imura, J.; Hiraishi, H.; Fujimori, T. Expression of SDF-1 Alpha and Nuclear CXCR4 Predicts Lymph Node Metastasis in Colorectal Cancer. Br. J. Cancer 2008, 98, 1682–1689. [Google Scholar] [CrossRef]
- Ghaemmaghami, S.; Mohaddes, S.M.; Hedayati, M.; Gorgian Mohammadi, M.; Dehbashi, G. Resistin and Visfatin Expression in HCT-116 Colorectal Cancer Cell Line. Int. J. Mol. Cell Med. 2013, 2, 143–150. [Google Scholar]
- Dalamaga, M. Nicotinamide Phosphoribosyl-Transferase/Visfatin: A Missing Link between Overweight/Obesity and Postmenopausal Breast Cancer? Potential Preventive and Therapeutic Perspectives and Challenges. Med. Hypotheses 2012, 79, 617–621. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Yang, Y.-H.; Su, J.-H.; Chang, H.-L.; Hou, M.-F.; Yuan, S.-S.F. High Visfatin Expression in Breast Cancer Tissue Is Associated with Poor Survival. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1892–1901. [Google Scholar] [CrossRef]
- Berta, J.; Hoda, M.A.; Laszlo, V.; Rozsas, A.; Garay, T.; Torok, S.; Grusch, M.; Berger, W.; Paku, S.; Renyi-Vamos, F.; et al. Apelin Promotes Lymphangiogenesis and Lymph Node Metastasis. Oncotarget 2014, 5, 4426–4437. [Google Scholar] [CrossRef] [PubMed]
- Kunduzova, O.; Alet, N.; Delesque-Touchard, N.; Millet, L.; Castan-Laurell, I.; Muller, C.; Dray, C.; Schaeffer, P.; Herault, J.P.; Savi, P.; et al. Apelin/APJ Signaling System: A Potential Link between Adipose Tissue and Endothelial Angiogenic Processes. FASEB J. 2008, 22, 4146–4153. [Google Scholar] [CrossRef] [PubMed]
- Heo, K.; Kim, Y.H.; Sung, H.J.; Li, H.Y.; Yoo, C.W.; Kim, J.Y.; Park, J.Y.; Lee, U.L.; Nam, B.H.; Kim, E.O.; et al. Hypoxia-Induced up-Regulation of Apelin Is Associated with a Poor Prognosis in Oral Squamous Cell Carcinoma Patients. Oral Oncol. 2012, 48, 500–506. [Google Scholar] [CrossRef]
- Yang, G.; Fan, W.; Luo, B.; Xu, Z.; Wang, P.; Tang, S.; Xu, P.; Yu, M. Circulating Resistin Levels and Risk of Colorectal Cancer: A Meta-Analysis. Biomed. Res. Int. 2016, 2016, 7367485. [Google Scholar] [CrossRef]
- Altinkaya, S.O.; Nergiz, S.; Küçük, M.; Yüksel, H. Apelin Levels Are Higher in Obese Patients with Endometrial Cancer. J. Obstet. Gynaecol. Res. 2015, 41, 294–300. [Google Scholar] [CrossRef]
- Hu, D.; Zhu, W.F.; Shen, W.C.; Xia, Y.; Wu, X.F.; Zhang, H.J.; Liu, W.; Cui, Z.L.; Zheng, X.W.; Chen, G. Expression of Apelin and Snail protein in breast cancer and their prognostic significance. Zhonghua Bing Li Xue Za Zhi 2018, 47, 743–746. [Google Scholar] [CrossRef]
- Salman, T.; Demir, L.; Varol, U.; Akyol, M.; Oflazoglu, U.; Yildiz, Y.; Taskaynatan, H.; Cengiz, H.; Guvendi, G.; Kucukzeybek, Y.; et al. Serum Apelin Levels and Body Composition Changes in Breast Cancer Patients Treated with an Aromatase Inhibitor. J. Buon. 2016, 21, 1419–1424. [Google Scholar]
- Kelly, A.S.; Steinberger, J.; Olson, T.P.; Dengel, D.R. In the Absence of Weight Loss, Exercise Training Does Not Improve Adipokines or Oxidative Stress in Overweight Children. Metabolism 2007, 56, 1005–1009. [Google Scholar] [CrossRef]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Front. Physiol. 2016, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Zaibi, M.S.; Stocker, C.J.; O’Dowd, J.; Davies, A.; Bellahcene, M.; Cawthorne, M.A.; Brown, A.J.H.; Smith, D.M.; Arch, J.R.S. Roles of GPR41 and GPR43 in Leptin Secretory Responses of Murine Adipocytes to Short Chain Fatty Acids. FEBS Lett. 2010, 584, 2381–2386. [Google Scholar] [CrossRef]
- Pos0329 Gastrointestinal Involvement in Systemic Sclerosis: Pathogenetic Role of Gut Microbiome, Cytokines and Adipokines|Annals of the Rheumatic Diseases. Available online: https://ard.bmj.com/content/80/Suppl_1/392.1 (accessed on 17 March 2023).
- Agranyoni, O.; Meninger-Mordechay, S.; Uzan, A.; Ziv, O.; Salmon-Divon, M.; Rodin, D.; Raz, O.; Koman, I.; Koren, O.; Pinhasov, A.; et al. Gut Microbiota Determines the Social Behavior of Mice and Induces Metabolic and Inflammatory Changes in Their Adipose Tissue. NPJ Biofilms Microbiomes 2021, 7, 28. [Google Scholar] [CrossRef]
- Nagpal, R.; Newman, T.M.; Wang, S.; Jain, S.; Lovato, J.F.; Yadav, H. Obesity-Linked Gut Microbiome Dysbiosis Associated with Derangements in Gut Permeability and Intestinal Cellular Homeostasis Independent of Diet. J. Diabetes Res. 2018, 2018, 3462092. [Google Scholar] [CrossRef] [PubMed]
- Sefcíková, Z.; Kmet, V.; Bujnáková, D.; Racek, L.; Mozes, S. Development of Gut Microflora in Obese and Lean Rats. Folia Microbiol. 2010, 55, 373–375. [Google Scholar] [CrossRef]
- de La Serre, C.B.; Ellis, C.L.; Lee, J.; Hartman, A.L.; Rutledge, J.C.; Raybould, H.E. Propensity to High-Fat Diet-Induced Obesity in Rats Is Associated with Changes in the Gut Microbiota and Gut Inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G440–G448. [Google Scholar] [CrossRef]
- Hamilton, M.K.; Boudry, G.; Lemay, D.G.; Raybould, H.E. Changes in Intestinal Barrier Function and Gut Microbiota in High-Fat Diet-Fed Rats Are Dynamic and Region Dependent. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G840–G851. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.N.; Olofsson, L.E. Gut Microbiota-Dependent Modulation of Energy Metabolism. J. Innate Immun. 2018, 10, 163–171. [Google Scholar] [CrossRef]
- Cunningham, A.L.; Stephens, J.W.; Harris, D.A. A Review on Gut Microbiota: A Central Factor in the Pathophysiology of Obesity. Lipids Health Dis. 2021, 20, 65. [Google Scholar] [CrossRef]
- Virtue, A.T.; McCright, S.J.; Wright, J.M.; Jimenez, M.T.; Mowel, W.K.; Kotzin, J.J.; Joannas, L.; Basavappa, M.G.; Spencer, S.P.; Clark, M.L.; et al. The Gut Microbiota Regulates White Adipose Tissue Inflammation and Obesity via a Family of MicroRNAs. Sci. Transl. Med. 2019, 11, eaav1892. [Google Scholar] [CrossRef]
- Puddu, A.; Sanguineti, R.; Montecucco, F.; Viviani, G.L. Evidence for the Gut Microbiota Short-Chain Fatty Acids as Key Pathophysiological Molecules Improving Diabetes. Mediat. Inflamm. 2014, 2014, 162021. [Google Scholar] [CrossRef]
- Sanz, Y.; Santacruz, A.; Gauffin, P. Gut Microbiota in Obesity and Metabolic Disorders. Proc. Nutr. Soc. 2010, 69, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Zamorano, N.; Fabbiano, S.; Chevalier, C.; Stojanović, O.; Colin, D.J.; Stevanović, A.; Veyrat-Durebex, C.; Tarallo, V.; Rigo, D.; Germain, S.; et al. Microbiota Depletion Promotes Browning of White Adipose Tissue and Reduces Obesity. Nat. Med. 2015, 21, 1497–1501. [Google Scholar] [CrossRef]
- Hong, J.; Jia, Y.; Pan, S.; Jia, L.; Li, H.; Han, Z.; Cai, D.; Zhao, R. Butyrate Alleviates High Fat Diet-Induced Obesity through Activation of Adiponectin-Mediated Pathway and Stimulation of Mitochondrial Function in the Skeletal Muscle of Mice. Oncotarget 2016, 7, 56071–56082. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.K.; Davenport, E.R.; Beaumont, M.; Jackson, M.A.; Knight, R.; Ober, C.; Spector, T.D.; Bell, J.T.; Clark, A.G.; Ley, R.E. Genetic Determinants of the Gut Microbiome in UK Twins. Cell Host Microbe 2016, 19, 731–743. [Google Scholar] [CrossRef]
- Li, G.; Xie, C.; Lu, S.; Nichols, R.G.; Tian, Y.; Li, L.; Patel, D.; Ma, Y.; Brocker, C.N.; Yan, T.; et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell Metab. 2017, 26, 672–685.e4. [Google Scholar] [CrossRef] [PubMed]
- Jocken, J.W.E.; González Hernández, M.A.; Hoebers, N.T.H.; van der Beek, C.M.; Essers, Y.P.G.; Blaak, E.E.; Canfora, E.E. Short-Chain Fatty Acids Differentially Affect Intracellular Lipolysis in a Human White Adipocyte Model. Front. Endocrinol. 2017, 8, 372. [Google Scholar] [CrossRef] [PubMed]
- Romacho, T.; Glosse, P.; Richter, I.; Elsen, M.; Schoemaker, M.H.; Van Tol, E.A.; Eckel, J. Nutritional Ingredients Modulate Adipokine Secretion and Inflammation in Human Primary Adipocytes. Nutrients 2015, 7, 865–886. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in Inflammation and Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the Pleiotropic Role of White Adipose Tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef]
- Gove, M.E.; Fantuzzi, G. Adipokines, Nutrition, and Obesity. In Preventive Nutrition: The Comprehensive Guide for Health Professionals; Bendich, A., Deckelbaum, R.J., Eds.; Nutrition and Health; Humana Press: Totowa, NJ, USA, 2010; pp. 419–432. ISBN 978-1-60327-542-2. [Google Scholar]
- Andreoli, M.F.; Donato, J.; Cakir, I.; Perello, M. Leptin Resensitisation: A Reversion of Leptin-Resistant States. J. Endocrinol. 2019, 241, R81–R96. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Herrera, K.; Florio, A.A.; Moore, M.; Marrero, A.; Tamez, M.; Bhupathiraju, S.N.; Mattei, J. The Leptin System and Diet: A Mini Review of the Current Evidence. Front. Endocrinol. 2021, 12, 749050. [Google Scholar] [CrossRef]
- Sumithran, P.; Prendergast, L.A.; Delbridge, E.; Purcell, K.; Shulkes, A.; Kriketos, A.; Proietto, J. Long-Term Persistence of Hormonal Adaptations to Weight Loss. N. Engl. J. Med. 2011, 365, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Latheef, S.K.; Dadar, M.; Samad, H.A.; Munjal, A.; Khandia, R.; Karthik, K.; Tiwari, R.; Yatoo, M.I.; Bhatt, P.; et al. Biomarkers in Stress Related Diseases/Disorders: Diagnostic, Prognostic, and Therapeutic Values. Front. Mol. Biosci. 2019, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Chen, X.; Mozzoli, M.; Ryan, I. Effect of Fasting on Serum Leptin in Normal Human Subjects. J. Clin. Endocrinol. Metab. 1996, 81, 3419–3423. [Google Scholar] [CrossRef]
- Mars, M.; de Graaf, C.; de Groot, L.C.P.G.M.; Kok, F.J. Decreases in Fasting Leptin and Insulin Concentrations after Acute Energy Restriction and Subsequent Compensation in Food Intake. Am. J. Clin. Nutr. 2005, 81, 570–577. [Google Scholar] [CrossRef]
- Wadden, T.A.; Considine, R.V.; Foster, G.D.; Anderson, D.A.; Sarwer, D.B.; Caro, J.S. Short- and Long-Term Changes in Serum Leptin Dieting Obese Women: Effects of Caloric Restriction and Weight Loss. J. Clin. Endocrinol. Metab. 1998, 83, 214–218. [Google Scholar] [CrossRef]
- Varkaneh Kord, H.; Tinsley, G.M.; Santos, H.O.; Zand, H.; Nazary, A.; Fatahi, S.; Mokhtari, Z.; Salehi-Sahlabadi, A.; Tan, S.C.; Rahmani, J.; et al. The Influence of Fasting and Energy-Restricted Diets on Leptin and Adiponectin Levels in Humans: A Systematic Review and Meta-Analysis. Clin. Nutr. 2021, 40, 1811–1821. [Google Scholar] [CrossRef]
- Cheng, Y.-C.; Liu, J.-R. Effect of Lactobacillus Rhamnosus GG on Energy Metabolism, Leptin Resistance, and Gut Microbiota in Mice with Diet-Induced Obesity. Nutrients 2020, 12, 2557. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Vilariño-García, T.; Guadix, P.; Dueñas, J.L.; Sánchez-Margalet, V. Leptin and Nutrition in Gestational Diabetes. Nutrients 2020, 12, 1970. [Google Scholar] [CrossRef] [PubMed]
- Al-muzafar, H.M.; Amin, K.A. Probiotic Mixture Improves Fatty Liver Disease by Virtue of Its Action on Lipid Profiles, Leptin, and Inflammatory Biomarkers. BMC Complement. Altern. Med. 2017, 17, 43. [Google Scholar] [CrossRef] [PubMed]
- Bagarolli, R.A.; Tobar, N.; Oliveira, A.G.; Araújo, T.G.; Carvalho, B.M.; Rocha, G.Z.; Vecina, J.F.; Calisto, K.; Guadagnini, D.; Prada, P.O.; et al. Probiotics Modulate Gut Microbiota and Improve Insulin Sensitivity in DIO Mice. J. Nutr. Biochem. 2017, 50, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Rouhani, M.H.; Hadi, A.; Ghaedi, E.; Salehi, M.; Mahdavi, A.; Mohammadi, H. Do Probiotics, Prebiotics and Synbiotics Affect Adiponectin and Leptin in Adults? A Systematic Review and Meta-Analysis of Clinical Trials. Clin. Nutr. 2019, 38, 2031–2037. [Google Scholar] [CrossRef]
- Sabour, H.; Norouzi Javidan, A.; Latifi, S.; Shidfar, F.; Heshmat, R.; Emami Razavi, S.-H.; Vafa, M.R.; Larijani, B. Omega-3 Fatty Acids’ Effect on Leptin and Adiponectin Concentrations in Patients with Spinal Cord Injury: A Double-Blinded Randomized Clinical Trial. J. Spinal Cord Med. 2015, 38, 599–606. [Google Scholar] [CrossRef]
- Sedláček, P.; Plavinová, I.; Langmajerová, J.; Dvořáková, J.; Novák, J.; Trefil, L.; Müller, L.; Buňatová, P.; Zeman, V.; Müllerová, D. Effect of N-3 Fatty Acids Supplementation during Life Style Modification in Women with Overweight. Cent. Eur. J. Public Health 2018, 26, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Batista, E.S.; da Silva Rios, T.; Muñoz, V.R.; Jesus, J.S.; Vasconcelos, M.M.; da Cunha, D.T.; Marques-Rocha, J.L.; Nakandakari, S.C.B.R.; Lara, R.; da Silva, A.S.R.; et al. Omega-3 Mechanism of Action in Inflammation and Endoplasmic Reticulum Stress in Mononuclear Cells from Overweight Non-Alcoholic Fatty Liver Disease Participants: Study Protocol for the “Brazilian Omega Study” (BROS)-a Randomized Controlled Trial. Trials 2021, 22, 927. [Google Scholar] [CrossRef] [PubMed]
- Balakrishna, R.; Bjørnerud, T.; Bemanian, M.; Aune, D.; Fadnes, L.T. Consumption of Nuts and Seeds and Health Outcomes Including Cardiovascular Disease, Diabetes and Metabolic Disease, Cancer, and Mortality: An Umbrella Review. Adv. Nutr. 2022, 13, 2136–2148. [Google Scholar] [CrossRef]
- Abbaspour, N.; Roberts, T.; Hooshmand, S.; Kern, M.; Hong, M.Y. Mixed Nut Consumption May Improve Cardiovascular Disease Risk Factors in Overweight and Obese Adults. Nutrients 2019, 11, 1488. [Google Scholar] [CrossRef]
- Liao, P.-J.; Ting, M.-K.; Wu, I.-W.; Chen, S.-W.; Yang, N.-I.; Hsu, K.-H. Higher Leptin-to-Adiponectin Ratio Strengthens the Association Between Body Measurements and Occurrence of Type 2 Diabetes Mellitus. Front. Public Health 2021, 9, 678681. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Yang, S.; Xiao, H.; Wang, M.; Ye, J.; Cao, L.; Sun, G. Role of Adiponectin in Cardiovascular Diseases Related to Glucose and Lipid Metabolism Disorders. Int. J. Mol. Sci. 2022, 23, 15627. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, P.; Kiss, T.; Tarantini, S.; Nyúl-Tóth, Á.; Ahire, C.; Yabluchanskiy, A.; Csipo, T.; Lipecz, A.; Tabak, A.; Institoris, A.; et al. Obesity-Induced Cognitive Impairment in Older Adults: A Microvascular Perspective. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H740–H761. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. Adiponectin-Resistance in Obesity. Adv. Exp. Med. Biol. 2017, 960, 415–441. [Google Scholar] [CrossRef]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Kroeger, C.M.; Trepanowski, J.F.; Haus, J.M.; Hoddy, K.K.; Calvo, Y. Alternate Day Fasting for Weight Loss in Normal Weight and Overweight Subjects: A Randomized Controlled Trial. Nutr. J. 2013, 12, 146. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.; Mirzaei, K.; Abdurahman, A.A.; Keshavarz, S.A. Adipokines May Mediate the Relationship between Resting Metabolic Rates and Bone Mineral Densities in Obese Women. Osteoporos. Int. 2017, 28, 1619–1629. [Google Scholar] [CrossRef]
- Löllgen, H.; Böckenhoff, A.; Knapp, G. Physical Activity and All-Cause Mortality: An Updated Meta-Analysis with Different Intensity Categories. Int. J. Sports Med. 2009, 30, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.W.; Pedersen, B.K. The Anti-Inflammatory Effect of Exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Monzillo, L.U.; Hamdy, O.; Horton, E.S.; Ledbury, S.; Mullooly, C.; Jarema, C.; Porter, S.; Ovalle, K.; Moussa, A.; Mantzoros, C.S. Effect of Lifestyle Modification on Adipokine Levels in Obese Subjects with Insulin Resistance. Obes. Res. 2003, 11, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Nassis, G.P.; Papantakou, K.; Skenderi, K.; Triandafillopoulou, M.; Kavouras, S.A.; Yannakoulia, M.; Chrousos, G.P.; Sidossis, L.S. Aerobic Exercise Training Improves Insulin Sensitivity without Changes in Body Weight, Body Fat, Adiponectin, and Inflammatory Markers in Overweight and Obese Girls. Metabolism 2005, 54, 1472–1479. [Google Scholar] [CrossRef]
- Jadhav, R.A.; Maiya, G.A.; Hombali, A.; Umakanth, S.; Shivashankar, K.N. Effect of Physical Activity Promotion on Adiponectin, Leptin and Other Inflammatory Markers in Prediabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Acta Diabetol. 2021, 58, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Hopps, E.; Canino, B.; Caimi, G. Effects of Exercise on Inflammation Markers in Type 2 Diabetic Subjects. Acta Diabetol. 2011, 48, 183–189. [Google Scholar] [CrossRef]
- Bouassida, A.; Zalleg, D.; Bouassida, S.; Zaouali, M.; Feki, Y.; Zbidi, A.; Tabka, Z. Leptin, Its Implication in Physical Exercise and Training: A Short Review. J. Sport. Sci. Med. 2006, 5, 172–181. [Google Scholar]
- Bouassida, A.; Zalleg, D.; Zaouali, M.; Gharbi, N.; Fekih, Y.; Richalet, J.P.; Tabka, Z. Effets d’un exercice supra-maximal sur les concentrations de la leptine plasmatique. Sci. Sport. 2004, 19, 136–138. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Konturek, S.J.; Duda, K.; Majerczak, J.; Sliwowski, Z.; Grandys, M.; Bielanski, W. Effect of Moderate Incremental Exercise, Performed in Fed and Fasted State on Cardio-Respiratory Variables and Leptin and Ghrelin Concentrations in Young Healthy Men. J. Physiol. Pharmacol. 2005, 56, 63–85. [Google Scholar]
- Kraemer, R.R.; Chu, H.; Castracane, V.D. Leptin and Exercise. Exp. Biol. Med. 2002, 227, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, M.; Ermolao, A.; Roi, G.S.; Englaro, P.; Tegon, G.; Varnier, M. Leptin Reduction after Endurance Races Differing in Duration and Energy Expenditure. Eur. J. Appl. Physiol. 2002, 87, 108–111. [Google Scholar] [CrossRef]
- Zafeiridis, A.; Smilios, I.; Considine, R.V.; Tokmakidis, S.P. Serum Leptin Responses after Acute Resistance Exercise Protocols. J. Appl. Physiol. 2003, 94, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Racil, G.; Zouhal, H.; Elmontassar, W.; Ben Abderrahmane, A.; De Sousa, M.V.; Chamari, K.; Amri, M.; Coquart, J.B. Plyometric Exercise Combined with High-Intensity Interval Training Improves Metabolic Abnormalities in Young Obese Females More so than Interval Training Alone. Appl. Physiol. Nutr. Metab. 2016, 41, 103–109. [Google Scholar] [CrossRef]
- Yaspelkis, B.B.; Singh, M.K.; Krisan, A.D.; Collins, D.E.; Kwong, C.C.; Bernard, J.R.; Crain, A.M. Chronic Leptin Treatment Enhances Insulin-Stimulated Glucose Disposal in Skeletal Muscle of High-Fat Fed Rodents. Life Sci. 2004, 74, 1801–1816. [Google Scholar] [CrossRef]
- Metcalf, B.S.; Jeffery, A.N.; Hosking, J.; Voss, L.D.; Sattar, N.; Wilkin, T.J. Objectively Measured Physical Activity and Its Association with Adiponectin and Other Novel Metabolic Markers: A Longitudinal Study in Children (EarlyBird 38). Diabetes Care 2009, 32, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Bunt, J.C.; Salbe, A.D.; Funahashi, T.; Matsuzawa, Y.; Tataranni, P.A. Plasma Adiponectin Concentrations in Children: Relationships with Obesity and Insulinemia. J. Clin. Endocrinol. Metab. 2002, 87, 4652–4656. [Google Scholar] [CrossRef]
- Shehzad, A.; Iqbal, W.; Shehzad, O.; Lee, Y.S. Adiponectin: Regulation of Its Production and Its Role in Human Diseases. Hormones 2012, 11, 8–20. [Google Scholar] [CrossRef]
- SciELO—Brazil—Efeito Do Treinamento Concorrente e Destreinamento Sobre o Biomarcador Anti-Inflamatório e Níveis de Condicionamento Físico Em Crianças Obesas Efeito Do Treinamento Concorrente e Destreinamento Sobre o Biomarcador Anti-Inflamatório e Níveis de Condicionamento Físico Em Crianças Obesas. Available online: https://www.scielo.br/j/rbme/a/H4BHxv76F7SK3YhpQYKDs5H/?format=html&lang=en (accessed on 17 March 2023).
- Kim, E.S.; Im, J.-A.; Kim, K.C.; Park, J.H.; Suh, S.-H.; Kang, E.S.; Kim, S.H.; Jekal, Y.; Lee, C.W.; Yoon, Y.-J.; et al. Improved Insulin Sensitivity and Adiponectin Level after Exercise Training in Obese Korean Youth. Obesity 2007, 15, 3023–3030. [Google Scholar] [CrossRef] [PubMed]
- Vasconcellos, F.; Seabra, A.; Cunha, F.; Montenegro, R.; Penha, J.; Bouskela, E.; Nogueira Neto, J.F.; Collett-Solberg, P.; Farinatti, P. Health Markers in Obese Adolescents Improved by a 12-Week Recreational Soccer Program: A Randomised Controlled Trial. J. Sports Sci. 2016, 34, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Marcell, T.J.; McAuley, K.A.; Traustadóttir, T.; Reaven, P.D. Exercise Training Is Not Associated with Improved Levels of C-Reactive Protein or Adiponectin. Metabolism 2005, 54, 533–541. [Google Scholar] [CrossRef]
- Kraemer, R.R.; Aboudehen, K.S.; Carruth, A.K.; Durand, R.T.J.; Acevedo, E.O.; Hebert, E.P.; Johnson, L.G.; Castracane, V.D. Adiponectin Responses to Continuous and Progressively Intense Intermittent Exercise. Med. Sci. Sports Exerc. 2003, 35, 1320–1325. [Google Scholar] [CrossRef]
- Jürimäe, J.; Purge, P.; Jürimäe, T. Adiponectin Is Altered after Maximal Exercise in Highly Trained Male Rowers. Eur. J. Appl. Physiol. 2005, 93, 502–505. [Google Scholar] [CrossRef]
- Hulver, M.W.; Zheng, D.; Tanner, C.J.; Houmard, J.A.; Kraus, W.E.; Slentz, C.A.; Sinha, M.K.; Pories, W.J.; MacDonald, K.G.; Dohm, G.L. Adiponectin Is Not Altered with Exercise Training despite Enhanced Insulin Action. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E861–E865. [Google Scholar] [CrossRef] [PubMed]
- Ring-Dimitriou, S.; Paulweber, B.; von Duvillard, S.P.; Stadlmann, M.; LeMura, L.M.; Lang, J.; Müller, E. The Effect of Physical Activity and Physical Fitness on Plasma Adiponectin in Adults with Predisposition to Metabolic Syndrome. Eur. J. Appl. Physiol. 2006, 98, 472–481. [Google Scholar] [CrossRef]
- Zhang, M.H.; Na, B.; Schiller, N.B.; Whooley, M.A. Resistin, Exercise Capacity, and Inducible Ischemia in Patients with Stable Coronary Heart Disease: Data from the Heart and Soul Study. Atherosclerosis 2010, 213, 604–610. [Google Scholar] [CrossRef]
- Kojta, I.; Chacińska, M.; Błachnio-Zabielska, A. Obesity, Bioactive Lipids, and Adipose Tissue Inflammation in Insulin Resistance. Nutrients 2020, 12, 1305. [Google Scholar] [CrossRef]
- Ben Ounis, O.; Elloumi, M.; Lac, G.; Makni, E.; Van Praagh, E.; Zouhal, H.; Tabka, Z.; Amri, M. Two-Month Effects of Individualized Exercise Training with or without Caloric Restriction on Plasma Adipocytokine Levels in Obese Female Adolescents. Ann. Endocrinol. 2009, 70, 235–241. [Google Scholar] [CrossRef] [PubMed]
- García-Hermoso, A.; Ceballos-Ceballos, R.J.M.; Poblete-Aro, C.E.; Hackney, A.C.; Mota, J.; Ramírez-Vélez, R. Exercise, Adipokines and Pediatric Obesity: A Meta-Analysis of Randomized Controlled Trials. Int. J. Obes. 2017, 41, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Jamurtas, A.Z. Exercise-Induced Muscle Damage and Oxidative Stress. Antioxidants 2018, 7, 50. [Google Scholar] [CrossRef]
- Anti-Inflammatory Effect of Exercise Training in Subjects with Type 2 Diabetes and the Metabolic Syndrome Is Dependent on Exercise Modalities and Independent of Weight Loss—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/19695853/ (accessed on 17 March 2023).
- Jorge, M.L.M.P.; de Oliveira, V.N.; Resende, N.M.; Paraiso, L.F.; Calixto, A.; Diniz, A.L.D.; Resende, E.S.; Ropelle, E.R.; Carvalheira, J.B.; Espindola, F.S.; et al. The Effects of Aerobic, Resistance, and Combined Exercise on Metabolic Control, Inflammatory Markers, Adipocytokines, and Muscle Insulin Signaling in Patients with Type 2 Diabetes Mellitus. Metabolism 2011, 60, 1244–1252. [Google Scholar] [CrossRef]
- Humińska-Lisowska, K.; Mieszkowski, J.; Kochanowicz, A.; Bojarczuk, A.; Niespodziński, B.; Brzezińska, P.; Stankiewicz, B.; Michałowska-Sawczyn, M.; Grzywacz, A.; Petr, M.; et al. Implications of Adipose Tissue Content for Changes in Serum Levels of Exercise-Induced Adipokines: A Quasi-Experimental Study. Int. J. Environ. Res. Public Health 2022, 19, 8782. [Google Scholar] [CrossRef]
- Fukuhara, A.; Matsuda, M.; Nishizawa, M.; Segawa, K.; Tanaka, M.; Kishimoto, K.; Matsuki, Y.; Murakami, M.; Ichisaka, T.; Murakami, H.; et al. Visfatin: A Protein Secreted by Visceral Fat That Mimics the Effects of Insulin. Science 2005, 307, 426–430. [Google Scholar] [CrossRef]
- Lee, K.-J.; Shin, Y.-A.; Lee, K.-Y.; Jun, T.-W.; Song, W. Aerobic Exercise Training-Induced Decrease in Plasma Visfatin and Insulin Resistance in Obese Female Adolescents. Int. J. Sport. Nutr. Exerc. Metab. 2010, 20, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M.; Kim, J.H.; Cho, G.J.; Baik, S.H.; Park, H.S.; Kim, S.M. Effect of Exercise Training on Plasma Visfatin and Eotaxin Levels. Eur. J. Endocrinol. 2007, 157, 437–442. [Google Scholar] [CrossRef]
- Seo, D.-I.; So, W.-Y.; Ha, S.; Yoo, E.-J.; Kim, D.; Singh, H.; Fahs, C.A.; Rossow, L.; Bemben, D.A.; Bemben, M.G.; et al. Effects of 12 Weeks of Combined Exercise Training on Visfatin and Metabolic Syndrome Factors in Obese Middle-Aged Women. J. Sport. Sci. Med. 2011, 10, 222–226. [Google Scholar]
- Ghanbari-Niaki, A.; Saghebjoo, M.; Soltani, R.; Kirwan, J.P. Plasma Visfatin is Increased after High-Intensity Exercise. Ann. Nutr. Metab. 2010, 57, 3–8. [Google Scholar] [CrossRef]
- Kadoglou, N.P.E.; Fotiadis, G.; Kapelouzou, A.; Kostakis, A.; Liapis, C.D.; Vrabas, I.S. The Differential Anti-Inflammatory Effects of Exercise Modalities and Their Association with Early Carotid Atherosclerosis Progression in Patients with Type 2 Diabetes. Diabet. Med. 2013, 30, e41–e50. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Fernandez, I.D.; Meng, Y.; Zhao, W.; Groth, S.W. Gut Hormones, Adipokines, and pro- and Anti-Inflammatory Cytokines/Markers in Loss of Control Eating: A Scoping Review. Appetite 2021, 166, 105442. [Google Scholar] [CrossRef] [PubMed]
- Pigatto, E.; Schiesaro, M.; Caputo, M.; Beggio, M.; Galozzi, P.; Cozzi, F.; Sfriso, P. POS0329 gastrointestinal involvement in systemic sclerosis: Pathogenetic role of gut microbiome, cytokines and adipokines. Ann. Rheum. Dis. 2021, 80, 392. [Google Scholar] [CrossRef]




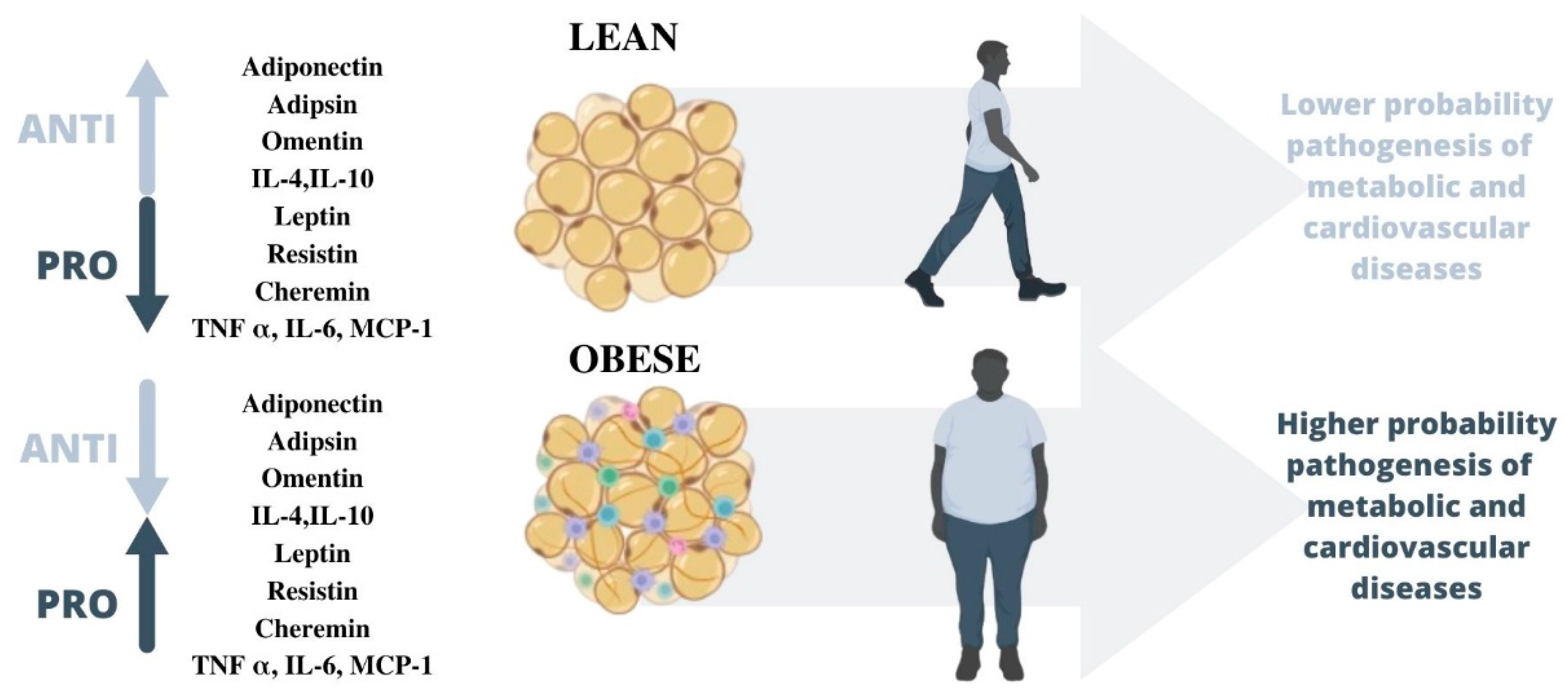{kind=link}
{kind=link}
{kind=link}
| Authors and Year | Study Title | Aim of Study | Main Outcomes | Section |
|---|---|---|---|---|
| Tilg et al. (2006) [46] | Adipocytokines: Mediators Linking Adipose Tissue, Inflammation, and Immunity | Understand adipocyte-derived mediators. | Adipocytokines—mediators are mainly synthesized by adipocytes in white adipose tissue. Adiponectin and leptin are the two most abundant adipocytokines. | Adipokines and inflammation |
| Ohashi et al. (2014) [66] | Role of Anti-Inflammatory Adipokines in Obesity-Related Diseases. Leptin in inflammation and autoimmunity | Understand the role of the anti-inflammatory adipokines. | Adiponectin exerts beneficial actions on obesity complications. | |
| La Cava (2018) [59] | Analyzes the known implications of leptin in multiple inflammatory conditions, including autoimmune diseases. | Leptin contributes to the generation and maintenance of low-grade inflammation. | ||
| Mitsuhashi et al. (2007) [80] | Adiponectin Level and Left Ventricular Hypertrophy in Japanese Men | Analyze the relationship that might exist between the serum adiponectin level and electrocardiographically diagnosed left ventricular hypertrophy (ECG-LVH) | Adiponectin concentration was inversely and independently associated with ECG-LVH. | Adipokines and cardiovascular disease |
| Khafaji et al. (2012) [91] | Elevated Serum Leptin Levels in Patients with Acute Myocardial Infarction; Correlation with Coronary Angiographic and Echocardiographic Findings | Assess the relationship between serial serum leptin levels in patients with acute myocardial infarction (AMI) | There was a trend for an increase in the mean serum leptin levels with an increasing number of diseased vessels. | |
| Hassan-Ali et al. (2011) [121] | Serum Adiponectin and Leptin as Predictors of the Presence and Degree of Coronary Atherosclerosis | Determine the relationship between serum adiponectin and leptin with the presence and degree of coronary atherosclerosis. | Both serum adiponectin and leptin might play an important pathogenic role not only in the occurrence of coronary artery disease patients. | Adipokines and atherosclerosis |
| Satayesh et al. (2021) [135] | The Possible Mediatory Role of Adipokines in the Association between Low Carbohydrate Diet and Depressive Symptoms among Overweight and Obese Women | Explore the possible mediatory role of adipokines Galectin-3, transforming growth factor-beta (TGF-β), and endothelial plasminogen activator inhibitor (PAI-1) in the association between low carbohydrate diet (LCD) and depressive symptoms. | Higher adherence to Low Carbohydrate Diet is probably associated with a lower prevalence of depressive symptoms in obese adults through the mediatory role of adipokines. | Adipokines and mental disease |
| Yu et al. (2021) [323] | Gut Hormones, Adipokines, and pro- and Anti-Inflammatory Cytokines/Markers in Loss of Control Eating: A Scoping Review | Synthesize research that has investigated these biomarkers with loss of control eating. | The use of glucagon-like peptide 1 analog was able to decrease binge eating. | Adipokines and eating behaviors |
| Authors and Year | Study Title | Aim of Study | Main Outcomes | Section |
|---|---|---|---|---|
| Ma et al. (2020) [183] | Lower Levels of Circulating Adiponectin in Elderly Patients with Metabolic Inflammatory Syndrome: A Cross-Sectional Study. | Assess whether circulating adiponectin can be used as an indicator for the metabolic inflammatory syndrome (MIS) in elderly adults. | Lower adiponectin levels in serum are associated with MIS | Adipokines and metabolic diseases |
| Sánchez-Jiménez et al. (2019) [202] | Obesity and Breast Cancer: Role of Leptin | Know a better knowledge of the molecular mechanisms that mediate leptin action may be helpful to understand the underlying processes which link obesity to breast cancer in postmenopausal women. | The activation of leptin signaling results in the concurrent activation of multiple oncogenic pathways leading to increased proliferation. | Adipokines and cancer |
| Stattin et al. (2001) [208] | Leptin Is Associated with Increased Prostate Cancer Risk: A Nested Case-Referent Study | Study the association between obesity and prostate cancer | A critical fat mass related to an interior milieu favorable for prostate cancer development seems to exist because intermediate but not high leptin levels are related to prostate cancer risk. | |
| Hong et al. (2016) [252] | Butyrate Alleviates High Fat Diet-Induced Obesity through Activation of Adiponectin-Mediated Pathway and Stimulation of Mitochondrial Function in the Skeletal Muscle of Mice | Explore whether the adiponectin-mediated pathway is involved in the anti-obesity action of butyrate | Short-term oral administration of SB can alleviate diet-induced obesity and insulin resistance in mice through the activation of the adiponectin-mediated pathway | The role of microbiota in adipokines |
| Pigatto et al. (2021) [324] | Gastrointestinal involvement in systemic sclerosis: Pathogenic role of the gut microbiome, cytokines, and adipokines | Evaluate serum levels of adipokines and cytokines involved in the pathogenesis of Systemic Sclerosis and their role. | Serum levels of IL12 and IL10 were found to correlate with specific bacteria alterations. | |
| Mendoza-Herrera et al. (2021) [261] | The Leptin System and Diet: A Mini Review of the Current Evidence | Review the literature on the relationship between diet and leptin, which suggests that addressing leptin resistance through dietary interventions can contribute to counteracting obesity | The link between nutritional components and leptin resistance, as well as research indicating that this condition is reversible, emphasizes the potential of diet to recover sensitivity to this hormone. | The role of nutrition in adipokines |
| Jadhav et al. (2021) [288] | Effect of Physical Activity Promotion on Adiponectin, Leptin and Other Inflammatory Markers in Prediabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials | Strengthen the evidence on the impact of physical activity promotion on inflammatory markers in prediabetes. | Meta-analysis found that physical activity with or without dietary or lifestyle modification reduces the level of leptin. | The role of physical activity in adipokines |
| Ghanbari-Niaki et al. (2010) [321] | Plasma Visfatin Is Increased after High-Intensity Exercise | Investigate the effects of repeated short bouts of high-intensity exercise on plasma visfatin and related metabolic responses | The elevation in plasma visfatin, together with increased plasma glucose and insulin concentrations immediately after high-intensity exercise |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. https://doi.org/10.3390/biomedicines11051290
Clemente-Suárez VJ, Redondo-Flórez L, Beltrán-Velasco AI, Martín-Rodríguez A, Martínez-Guardado I, Navarro-Jiménez E, Laborde-Cárdenas CC, Tornero-Aguilera JF. The Role of Adipokines in Health and Disease. Biomedicines. 2023; 11(5):1290. https://doi.org/10.3390/biomedicines11051290
Chicago/Turabian StyleClemente-Suárez, Vicente Javier, Laura Redondo-Flórez, Ana Isabel Beltrán-Velasco, Alexandra Martín-Rodríguez, Ismael Martínez-Guardado, Eduardo Navarro-Jiménez, Carmen Cecilia Laborde-Cárdenas, and José Francisco Tornero-Aguilera. 2023. "The Role of Adipokines in Health and Disease" Biomedicines 11, no. 5: 1290. https://doi.org/10.3390/biomedicines11051290
APA StyleClemente-Suárez, V. J., Redondo-Flórez, L., Beltrán-Velasco, A. I., Martín-Rodríguez, A., Martínez-Guardado, I., Navarro-Jiménez, E., Laborde-Cárdenas, C. C., & Tornero-Aguilera, J. F. (2023). The Role of Adipokines in Health and Disease. Biomedicines, 11(5), 1290. https://doi.org/10.3390/biomedicines11051290