
Adipokines as Diagnostic and Prognostic Markers for the Severity of COVID-19



Abstract
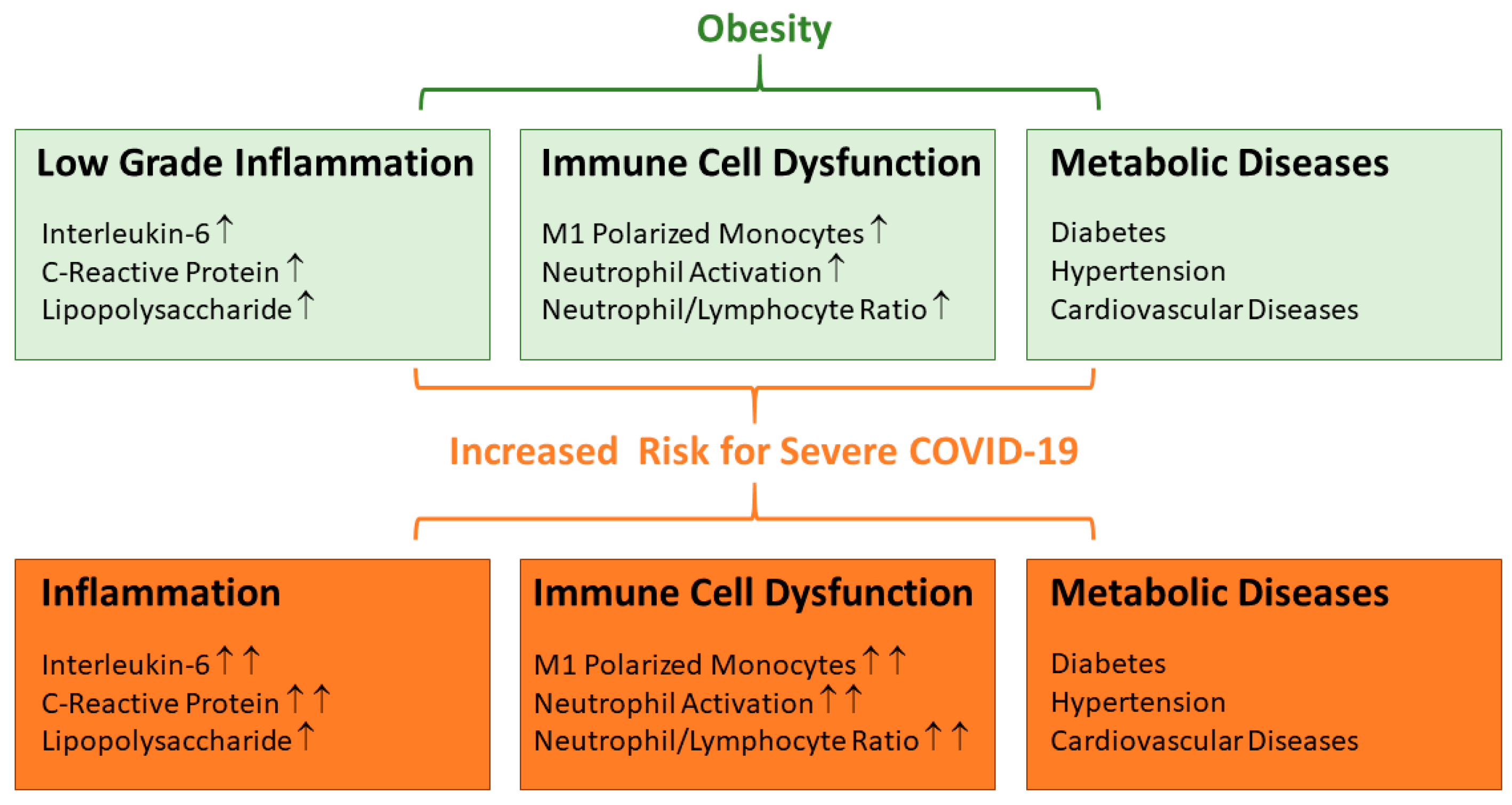
1. Introduction
2. Adipokines
2.1. Chemerin
2.2. Adiponectin
2.3. Leptin
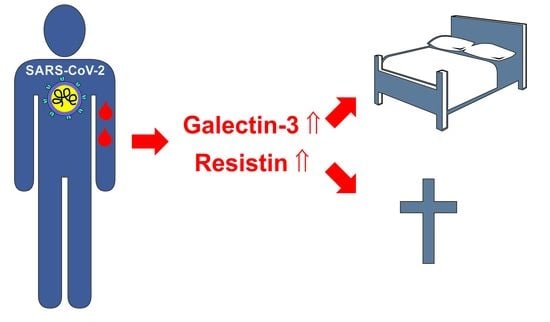
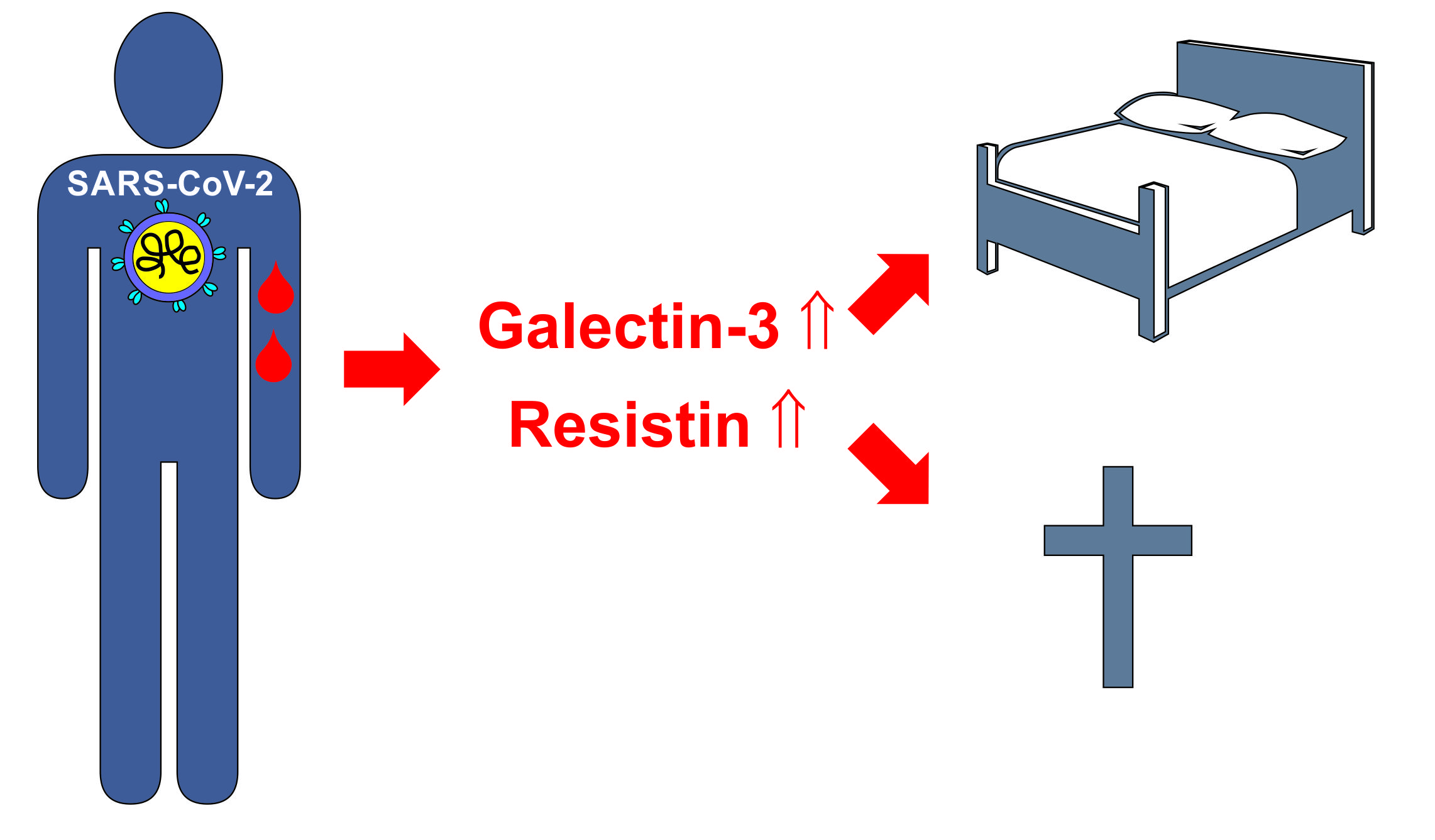
2.4. Resistin
2.5. Galectin-3
2.6. Visfatin
2.7. Apelin
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khot, W.Y.; Nadkar, M.Y. The 2019 Novel Coronavirus Outbreak—A Global Threat. J. Assoc. Physicians India 2020, 68, 67–71. [Google Scholar] [PubMed]
- Zsichla, L.; Muller, V. Risk Factors of Severe COVID-19: A Review of Host, Viral and Environmental Factors. Viruses 2023, 15, 175. [Google Scholar] [CrossRef] [PubMed]
- Aluganti Narasimhulu, C.; Singla, D.K. Mechanisms of COVID-19 pathogenesis in diabetes. Am. J. Physiol. Heart Circ. Physiol. 2022, 323, H403–H420. [Google Scholar] [CrossRef] [PubMed]
- Jeeyavudeen, M.S.; Chaudhari, R.; Pappachan, J.M.; Fouda, S. Clinical implications of COVID-19 in patients with metabolic-associated fatty liver disease. World J. Gastroenterol. 2023, 29, 487–502. [Google Scholar] [CrossRef]
- Hornung, F.; Rogal, J.; Loskill, P.; Loffler, B.; Deinhardt-Emmer, S. The Inflammatory Profile of Obesity and the Role on Pulmonary Bacterial and Viral Infections. Int. J. Mol. Sci. 2021, 22, 3456. [Google Scholar] [CrossRef]
- Fezeu, L.; Julia, C.; Henegar, A.; Bitu, J.; Hu, F.B.; Grobbee, D.E.; Kengne, A.P.; Hercberg, S.; Czernichow, S. Obesity is associated with higher risk of intensive care unit admission and death in influenza A (H1N1) patients: A systematic review and meta-analysis. Obes. Rev. 2011, 12, 653–659. [Google Scholar] [CrossRef]
- Moser, J.S.; Galindo-Fraga, A.; Ortiz-Hernandez, A.A.; Gu, W.; Hunsberger, S.; Galan-Herrera, J.F.; Guerrero, M.L.; Ruiz-Palacios, G.M.; Beigel, J.H.; La Red ILI 002 Study Group; et al. Underweight, overweight, and obesity as independent risk factors for hospitalization in adults and children from influenza and other respiratory viruses. Influenza Other Respir. Viruses 2019, 13, 3–9. [Google Scholar] [CrossRef]
- Alqahtani, F.Y.; Aleanizy, F.S.; Ali El Hadi Mohamed, R.; Alanazi, M.S.; Mohamed, N.; Alrasheed, M.M.; Abanmy, N.; Alhawassi, T. Prevalence of comorbidities in cases of Middle East respiratory syndrome coronavirus: A retrospective study. Epidemiol. Infect. 2018, 147, e35. [Google Scholar] [CrossRef]
- Frank, A.P.; de Souza Santos, R.; Palmer, B.F.; Clegg, D.J. Determinants of body fat distribution in humans may provide insight about obesity-related health risks. J. Lipid Res. 2019, 60, 1710–1719. [Google Scholar] [CrossRef]
- Alarcon, P.C.; Damen, M.; Madan, R.; Deepe, G.S., Jr.; Spearman, P.; Way, S.S.; Divanovic, S. Adipocyte inflammation and pathogenesis of viral pneumonias: An overlooked contribution. Mucosal Immunol. 2021, 14, 1224–1234. [Google Scholar] [CrossRef]
- Paich, H.A.; Sheridan, P.A.; Handy, J.; Karlsson, E.A.; Schultz-Cherry, S.; Hudgens, M.G.; Noah, T.L.; Weir, S.S.; Beck, M.A. Overweight and obese adult humans have a defective cellular immune response to pandemic H1N1 influenza A virus. Obesity 2013, 21, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Pranata, R.; Lim, M.A.; Yonas, E.; Vania, R.; Lukito, A.A.; Siswanto, B.B.; Meyer, M. Body mass index and outcome in patients with COVID-19: A dose-response meta-analysis. Diabetes Metab. 2021, 47, 101178. [Google Scholar] [CrossRef] [PubMed]
- Hafez, W.; Abdelshakor, M.; Kishk, S.; Gebril, A.; Gador, M.; Osman, S.; Abuelsaoud, H.M.; Abdelrahman, A. Body Mass Index and Clinical Outcomes in Adult COVID-19 Patients of Diverse Ethnicities. Healthcare 2022, 10, 2575. [Google Scholar] [CrossRef] [PubMed]
- Hajifathalian, K.; Kumar, S.; Newberry, C.; Shah, S.; Fortune, B.; Krisko, T.; Ortiz-Pujols, S.; Zhou, X.K.; Dannenberg, A.J.; Kumar, R.; et al. Obesity is Associated with Worse Outcomes in COVID-19: Analysis of Early Data from New York City. Obesity 2020, 28, 1606–1612. [Google Scholar] [CrossRef]
- Kalligeros, M.; Shehadeh, F.; Mylona, E.K.; Benitez, G.; Beckwith, C.G.; Chan, P.A.; Mylonakis, E. Association of Obesity with Disease Severity Among Patients with Coronavirus Disease 2019. Obesity 2020, 28, 1200–1204. [Google Scholar] [CrossRef]
- Ghanim, H.; Aljada, A.; Hofmeyer, D.; Syed, T.; Mohanty, P.; Dandona, P. Circulating mononuclear cells in the obese are in a proinflammatory state. Circulation 2004, 110, 1564–1571. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils Actively Contribute to Obesity-Associated Inflammation and Pathological Complications. Cells 2022, 11, 1883. [Google Scholar] [CrossRef]
- Yildirim, K.; Karabicak, I.; Ulu, E.K.; Aktimur, R.; Gursel, M.F.; Malazgirt, Z. Does Neutrophil-Lymphocyte Ratio Correlate with the Improvement of Hepatosteatosis after Laparoscopic Sleeve Gastrectomy? Obes. Facts 2022, 15, 711–716. [Google Scholar] [CrossRef]
- Coomes, E.A.; Haghbayan, H. Interleukin-6 in COVID-19: A systematic review and meta-analysis. Rev. Med. Virol. 2020, 30, 1–9. [Google Scholar] [CrossRef]
- Ikeagwulonu, R.C.; Ugwu, N.I.; Ezeonu, C.T.; Ikeagwulonu, Z.C.; Uro-Chukwu, H.C.; Asiegbu, U.V.; Obu, D.C.; Briggs, D.C. C-Reactive Protein and COVID-19 Severity: A Systematic Review. West. Afr. J. Med. 2021, 38, 1011–1023. [Google Scholar]
- Teixeira, P.C.; Dorneles, G.P.; Santana Filho, P.C.; da Silva, I.M.; Schipper, L.L.; Postiga, I.A.L.; Neves, C.A.M.; Rodrigues Junior, L.C.; Peres, A.; Souto, J.T.; et al. Increased LPS levels coexist with systemic inflammation and result in monocyte activation in severe COVID-19 patients. Int. Immunopharmacol. 2021, 100, 108125. [Google Scholar] [CrossRef] [PubMed]
- McKenna, E.; Wubben, R.; Isaza-Correa, J.M.; Melo, A.M.; Mhaonaigh, A.U.; Conlon, N.; O’Donnell, J.S.; Ni Cheallaigh, C.; Hurley, T.; Stevenson, N.J.; et al. Neutrophils in COVID-19: Not Innocent Bystanders. Front. Immunol. 2022, 13, 864387. [Google Scholar] [CrossRef]
- Wang Chau, C.; Sugimura, R. Locked in a pro-inflammatory state. eLife 2022, 11, e80699. [Google Scholar] [CrossRef] [PubMed]
- de Almeida-Pititto, B.; Dualib, P.M.; Zajdenverg, L.; Dantas, J.R.; de Souza, F.D.; Rodacki, M.; Bertoluci, M.C.; Brazilian Diabetes Society Study Group (SBD). Severity and mortality of COVID 19 in patients with diabetes, hypertension and cardiovascular disease: A meta-analysis. Diabetol. Metab. Syndr. 2020, 12, 75. [Google Scholar] [CrossRef]
- Cooper, A.J.; Gupta, S.R.; Moustafa, A.F.; Chao, A.M. Sex/Gender Differences in Obesity Prevalence, Comorbidities, and Treatment. Curr. Obes. Rep. 2021, 10, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Schaffler, A. Does global gene expression analysis in type 2 diabetes provide an opportunity to identify highly promising drug targets? Endocr. Metab. Immune Disord. Drug. Targets 2007, 7, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Eagon, J.C.; Trujillo, M.E.; Scherer, P.E.; Klein, S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef]
- Wiest, R.; Weigert, J.; Wanninger, J.; Neumeier, M.; Bauer, S.; Schmidhofer, S.; Farkas, S.; Scherer, M.N.; Schaffler, A.; Scholmerich, J.; et al. Impaired hepatic removal of interleukin-6 in patients with liver cirrhosis. Cytokine 2011, 53, 178–183. [Google Scholar] [CrossRef]
- Yu, J.Y.; Choi, W.J.; Lee, H.S.; Lee, J.W. Relationship between inflammatory markers and visceral obesity in obese and overweight Korean adults: An observational study. Medicine 2019, 98, e14740. [Google Scholar] [CrossRef]
- Battisti, S.; Pedone, C.; Napoli, N.; Russo, E.; Agnoletti, V.; Nigra, S.G.; Dengo, C.; Mughetti, M.; Conte, C.; Pozzilli, P.; et al. Computed Tomography Highlights Increased Visceral Adiposity Associated With Critical Illness in COVID-19. Diabetes Care 2020, 43, e129–e130. [Google Scholar] [CrossRef] [PubMed]
- Pranata, R.; Lim, M.A.; Huang, I.; Yonas, E.; Henrina, J.; Vania, R.; Lukito, A.A.; Nasution, S.A.; Alwi, I.; Siswanto, B.B. Visceral adiposity, subcutaneous adiposity, and severe coronavirus disease-2019 (COVID-19): Systematic review and meta-analysis. Clin. Nutr. ESPEN 2021, 43, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Goehler, A.; Hsu, T.H.; Seiglie, J.A.; Siedner, M.J.; Lo, J.; Triant, V.; Hsu, J.; Foulkes, A.; Bassett, I.; Khorasani, R.; et al. Visceral Adiposity and Severe COVID-19 Disease: Application of an Artificial Intelligence Algorithm to Improve Clinical Risk Prediction. Open. Forum Infect. Dis. 2021, 8, ofab275. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sun, X.; Han, D.; Zhong, J.; Zhang, H.; Zheng, L. Visceral adipose tissue and risk of COVID-19 susceptibility, hospitalization, and severity: A Mendelian randomization study. Front. Public Health 2022, 10, 1023935. [Google Scholar] [CrossRef]
- Camhi, S.M.; Bray, G.A.; Bouchard, C.; Greenway, F.L.; Johnson, W.D.; Newton, R.L.; Ravussin, E.; Ryan, D.H.; Smith, S.R.; Katzmarzyk, P.T. The relationship of waist circumference and BMI to visceral, subcutaneous, and total body fat: Sex and race differences. Obesity 2011, 19, 402–408. [Google Scholar] [CrossRef]
- Marcelin, G.; Gautier, E.L.; Clement, K. Adipose Tissue Fibrosis in Obesity: Etiology and Challenges. Annu. Rev. Physiol. 2022, 84, 135–155. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adiponectin in inflammatory and immune-mediated diseases. Cytokine 2013, 64, 1–10. [Google Scholar] [CrossRef]
- Ernst, M.C.; Sinal, C.J. Chemerin: At the crossroads of inflammation and obesity. Trends Endocrinol. Metab. 2010, 21, 660–667. [Google Scholar] [CrossRef]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gomez-Reino, J.J.; Mera, A.; Lago, F.; Gomez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef]
- Reiterer, M.; Rajan, M.; Gomez-Banoy, N.; Lau, J.D.; Gomez-Escobar, L.G.; Ma, L.; Gilani, A.; Alvarez-Mulett, S.; Sholle, E.T.; Chandar, V.; et al. Hyperglycemia in acute COVID-19 is characterized by insulin resistance and adipose tissue infectivity by SARS-CoV-2. Cell Metab. 2021, 33, 2174–2188.e2175. [Google Scholar] [CrossRef]
- Martinez-Colon, G.J.; Ratnasiri, K.; Chen, H.; Jiang, S.; Zanley, E.; Rustagi, A.; Verma, R.; Chen, H.; Andrews, J.R.; Mertz, K.D.; et al. SARS-CoV-2 infection drives an inflammatory response in human adipose tissue through infection of adipocytes and macrophages. Sci. Transl. Med. 2022, 14, eabm9151. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Feder, S.; Haberl, E.M.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mol. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef]
- Parlee, S.D.; McNeil, J.O.; Muruganandan, S.; Sinal, C.J.; Goralski, K.B. Elastase and tryptase govern TNFalpha-mediated production of active chemerin by adipocytes. PLoS ONE 2012, 7, e51072. [Google Scholar] [CrossRef] [PubMed]
- Cash, J.L.; Hart, R.; Russ, A.; Dixon, J.P.; Colledge, W.H.; Doran, J.; Hendrick, A.G.; Carlton, M.B.; Greaves, D.R. Synthetic chemerin-derived peptides suppress inflammation through ChemR23. J. Exp. Med. 2008, 205, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Bondue, B.; Vosters, O.; de Nadai, P.; Glineur, S.; De Henau, O.; Luangsay, S.; Van Gool, F.; Communi, D.; De Vuyst, P.; Desmecht, D.; et al. ChemR23 dampens lung inflammation and enhances anti-viral immunity in a mouse model of acute viral pneumonia. PLoS Pathog. 2011, 7, e1002358. [Google Scholar] [CrossRef]
- Lee, C.H. Role of specialized pro-resolving lipid mediators and their receptors in virus infection: A promising therapeutic strategy for SARS-CoV-2 cytokine storm. Arch. Pharm. Res. 2021, 44, 84–98. [Google Scholar] [CrossRef]
- Rajasagi, N.K.; Reddy, P.B.; Suryawanshi, A.; Mulik, S.; Gjorstrup, P.; Rouse, B.T. Controlling herpes simplex virus-induced ocular inflammatory lesions with the lipid-derived mediator resolvin E1. J. Immunol. 2011, 186, 1735–1746. [Google Scholar] [CrossRef]
- Serhan, C.N.; Libreros, S.; Nshimiyimana, R. E-series resolvin metabolome, biosynthesis and critical role of stereochemistry of specialized pro-resolving mediators (SPMs) in inflammation-resolution: Preparing SPMs for long COVID-19, human clinical trials, and targeted precision nutrition. Semin. Immunol. 2022, 59, 101597. [Google Scholar] [CrossRef]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. CMKLR1 and GPR1 mediate chemerin signaling through the RhoA/ROCK pathway. Mol. Cell. Endocrinol. 2015, 417, 36–51. [Google Scholar] [CrossRef]
- Haberl, E.M.; Pohl, R.; Rein-Fischboeck, L.; Feder, S.; Eisinger, K.; Krautbauer, S.; Sinal, C.J.; Buechler, C. Ex vivo analysis of serum chemerin activity in murine models of obesity. Cytokine 2018, 104, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Toulany, J.; Parlee, S.D.; Sinal, C.J.; Slayter, K.; McNeil, S.; Goralski, K.B. CMKLR1 activation ex vivo does not increase proportionally to serum total chemerin in obese humans. Endocr. Connect. 2016, 5, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Barnea, G.; Strapps, W.; Herrada, G.; Berman, Y.; Ong, J.; Kloss, B.; Axel, R.; Lee, K.J. The genetic design of signaling cascades to record receptor activation. Proc. Natl. Acad. Sci. USA 2008, 105, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Banas, M.; Zabieglo, K.; Kasetty, G.; Kapinska-Mrowiecka, M.; Borowczyk, J.; Drukala, J.; Murzyn, K.; Zabel, B.A.; Butcher, E.C.; Schroeder, J.M.; et al. Chemerin is an antimicrobial agent in human epidermis. PLoS ONE 2013, 8, e58709. [Google Scholar] [CrossRef]
- Chang, S.S.; Eisenberg, D.; Zhao, L.; Adams, C.; Leib, R.; Morser, J.; Leung, L. Chemerin activation in human obesity. Obesity 2016, 24, 1522–1529. [Google Scholar] [CrossRef]
- Shin, H.Y.; Lee, D.C.; Chu, S.H.; Jeon, J.Y.; Lee, M.K.; Im, J.A.; Lee, J.W. Chemerin levels are positively correlated with abdominal visceral fat accumulation. Clin. Endocrinol. 2012, 77, 47–50. [Google Scholar] [CrossRef]
- Acewicz, M.; Kasacka, I. Chemerin activity in selected pathological states of human body—A systematic review. Adv. Med. Sci. 2021, 66, 270–278. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Z.; Han, X.; Du, H.; Cao, Y.; Liu, Y.; Wang, W. Association between Congenital Cytomegalovirus Infection and Brain Injury in Neonates: A Meta-Analysis of Cohort Studies. Behav. Neurol. 2021, 2021, 9603660. [Google Scholar] [CrossRef]
- Vorontsov, O.; Levitt, L.; Lilleri, D.; Vainer, G.W.; Kaplan, O.; Schreiber, L.; Arossa, A.; Spinillo, A.; Furione, M.; Alfi, O.; et al. Amniotic fluid biomarkers predict the severity of congenital cytomegalovirus infection. J. Clin. Investig. 2022, 132, e157415. [Google Scholar] [CrossRef]
- Peschel, G.; Grimm, J.; Gulow, K.; Muller, M.; Buechler, C.; Weigand, K. Chemerin Is a Valuable Biomarker in Patients with HCV Infection and Correlates with Liver Injury. Diagnostics 2020, 10, 974. [Google Scholar] [CrossRef]
- Kukla, M.; Menzyk, T.; Dembinski, M.; Winiarski, M.; Garlicki, A.; Bociaga-Jasik, M.; Skonieczna, M.; Hudy, D.; Maziarz, B.; Kusnierz-Cabala, B.; et al. Anti-inflammatory adipokines: Chemerin, vaspin, omentin concentrations and SARS-CoV-2 outcomes. Sci. Rep. 2021, 11, 21514. [Google Scholar] [CrossRef] [PubMed]
- Lavis, P.; Morra, S.; Orte Cano, C.; Albayrak, N.; Corbiere, V.; Olislagers, V.; Dauby, N.; Del Marmol, V.; Marchant, A.; Decaestecker, C.; et al. Chemerin plasma levels are increased in COVID-19 patients and are an independent risk factor of mortality. Front. Immunol. 2022, 13, 941663. [Google Scholar] [CrossRef] [PubMed]
- Sulicka-Grodzicka, J.; Surdacki, A.; Surmiak, M.; Sanak, M.; Wizner, B.; Sydor, W.; Bociaga-Jasik, M.; Strach, M.; Korkosz, M.; Skladany, L.; et al. Chemerin as a Potential Marker of Resolution of Inflammation in COVID-19 Infection. Biomedicines 2022, 10, 2462. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiao, B.; Qu, L.; Yang, D.; Liu, R. The development of COVID-19 treatment. Front. Immunol. 2023, 14, 1125246. [Google Scholar] [CrossRef] [PubMed]
- Blot, M.; David, M.; Nguyen, M.; Bourredjem, A.; Group, L.S.; Binquet, C.; Piroth, L. Are adipokines the missing link between obesity, immune response, and outcomes in severe COVID-19? Int. J. Obes. 2021, 45, 2126–2131. [Google Scholar] [CrossRef]
- Caterino, M.; Gelzo, M.; Sol, S.; Fedele, R.; Annunziata, A.; Calabrese, C.; Fiorentino, G.; D’Abbraccio, M.; Dell’Isola, C.; Fusco, F.M.; et al. Dysregulation of lipid metabolism and pathological inflammation in patients with COVID-19. Sci. Rep. 2021, 11, 2941. [Google Scholar] [CrossRef]
- de Nooijer, A.H.; Kooistra, E.J.; Grondman, I.; Janssen, N.A.F.; Joosten, L.A.B.; van de Veerdonk, F.L.; Kox, M.; Pickkers, P.; Netea, M.G.; RCI-COVID-19 Study Group. Adipocytokine plasma concentrations reflect influence of inflammation but not body mass index (BMI) on clinical outcomes of COVID-19 patients: A prospective observational study from the Netherlands. Clin. Obes. 2022, 13, e12568. [Google Scholar] [CrossRef]
- Di Filippo, L.; De Lorenzo, R.; Sciorati, C.; Capobianco, A.; Lore, N.I.; Giustina, A.; Manfredi, A.A.; Rovere-Querini, P.; Conte, C. Adiponectin to leptin ratio reflects inflammatory burden and survival in COVID-19. Diabetes Metab. 2021, 47, 101268. [Google Scholar] [CrossRef]
- Minuzzi, L.G.; Seelaender, M.; Silva, B.S.A.; Cunha, E.; Deus, M.C.; Vasconcellos, F.T.F.; Marqueze, L.F.B.; Gadotti, A.C.; Baena, C.P.; Pereira, T.; et al. COVID-19 Outcome Relates with Circulating BDNF, According to Patient Adiposity and Age. Front. Nutr. 2021, 8, 784429. [Google Scholar] [CrossRef]
- Ryrso, C.K.; Dungu, A.M.; Hegelund, M.H.; Jensen, A.V.; Sejdic, A.; Faurholt-Jepsen, D.; Krogh-Madsen, R.; Lindegaard, B. Body composition, physical capacity, and immuno-metabolic profile in community-acquired pneumonia caused by COVID-19, influenza, and bacteria: A prospective cohort study. Int. J. Obes. 2022, 46, 817–824. [Google Scholar] [CrossRef]
- Tonon, F.; Di Bella, S.; Giudici, F.; Zerbato, V.; Segat, L.; Koncan, R.; Misin, A.; Toffoli, B.; D’Agaro, P.; Luzzati, R.; et al. Discriminatory Value of Adiponectin to Leptin Ratio for COVID-19 Pneumonia. Int. J. Endocrinol. 2022, 2022, 9908450. [Google Scholar] [CrossRef] [PubMed]
- Beltrao, F.E.L.; Beltrao, D.C.A.; Carvalhal, G.; Beltrao, F.N.L.; de Aquino, I.M.; Brito, T.D.S.; Paulino, B.C.; Aires, E.; Viegas, D.; Hecht, F.; et al. Low muscle mass and high visceral fat mass predict mortality in patients hospitalized with moderate-to-severe COVID-19: A prospective study. Endocr. Connect. 2022, 11, e220290. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A.; Lipcsey, M.; Hultstrom, M.; Frithiof, R.; Eriksson, M. Plasma Leptin Is Increased in Intensive Care Patients with COVID-19—An Investigation Performed in the PronMed-Cohort. Biomedicines 2021, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Perrotta, F.; Scialo, F.; Mallardo, M.; Signoriello, G.; D’Agnano, V.; Bianco, A.; Daniele, A.; Nigro, E. Adiponectin, Leptin, and Resistin Are Dysregulated in Patients Infected by SARS-CoV-2. Int. J. Mol. Sci. 2023, 24, 1131. [Google Scholar] [CrossRef]
- Ebihara, T.; Matsumoto, H.; Matsubara, T.; Togami, Y.; Nakao, S.; Matsuura, H.; Onishi, S.; Kojima, T.; Sugihara, F.; Okuzaki, D.; et al. Resistin Associated with Cytokines and Endothelial Cell Adhesion Molecules Is Related to Worse Outcome in COVID-19. Front. Immunol. 2022, 13, 830061. [Google Scholar] [CrossRef]
- Flikweert, A.W.; Kobold, A.C.M.; van der Sar-van der Brugge, S.; Heeringa, P.; Rodenhuis-Zybert, I.A.; Bijzet, J.; Tami, A.; van der Gun, B.T.F.; Wold, K.I.; Huckriede, A.; et al. Circulating adipokine levels and COVID-19 severity in hospitalized patients. Int. J. Obes. 2023, 47, 126–137. [Google Scholar] [CrossRef]
- Perpinan, C.; Bertran, L.; Terra, X.; Aguilar, C.; Binetti, J.; Lopez-Dupla, M.; Rull, A.; Reverte, L.; Yeregui, E.; Gomez-Bertomeu, F.; et al. Resistin and IL-15 as Predictors of Invasive Mechanical Ventilation in COVID-19 Pneumonia Irrespective of the Presence of Obesity and Metabolic Syndrome. J. Pers. Med. 2022, 12, 391. [Google Scholar] [CrossRef]
- Baykiz, D.; Emet, S.; Ayduk-Govdeli, E.; Kaytaz, M.; Yavuz, M.L.; Karaca-Ozer, P.; Karaayvaz, E.B.; Medetalibeyoglu, A.; Elitok, A.; Genc, S.; et al. Galectin-3 as a Novel Biomarker for Predicting Clinical Outcomes in Hospitalized COVID-19 Patients. Clin. Lab. 2022, 68. [Google Scholar] [CrossRef]
- Berber, N.K.; Geckil, A.A.; Altan, N.O.; Kiran, T.R.; Otlu, O.; Erdem, M.; In, E. Efficacy of Serum Apelin and Galectin-3 as Potential Predictors of Mortality in Severe COVID-19 Patients. J. Med. Virol. 2023, 95, e28494. [Google Scholar] [CrossRef]
- Bruni, F.; Charitos, P.; Lampart, M.; Moser, S.; Siegemund, M.; Bingisser, R.; Osswald, S.; Bassetti, S.; Twerenbold, R.; Trendelenburg, M.; et al. Complement and endothelial cell activation in COVID-19 patients compared to controls with suspected SARS-CoV-2 infection: A prospective cohort study. Front. Immunol. 2022, 13, 941742. [Google Scholar] [CrossRef]
- Caniglia, J.L.; Asuthkar, S.; Tsung, A.J.; Guda, M.R.; Velpula, K.K. Immunopathology of galectin-3: An increasingly promising target in COVID-19. F1000Resarch 2020, 9, 1078. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Alvarez, E.; la Rosa, N.L.; la Mora, M.S.; Valdez-Sandoval, P.; Palacios-Jimenez, M.; Rodriguez-Alvarez, F.; Vera-Maldonado, B.I.; Aguirre-Aguilar, E.; Escobar-Valderrama, J.M.; Alanis-Mendizabal, J.; et al. Galectin-3 as a potential prognostic biomarker of severe COVID-19 in SARS-CoV-2 infected patients. Sci. Rep. 2022, 12, 1856. [Google Scholar] [CrossRef] [PubMed]
- Gajovic, N.; Markovic, S.S.; Jurisevic, M.; Jovanovic, M.; Arsenijevic, N.; Mijailovic, Z.; Jovanovic, M.; Jovanovic, I. Galectin-3 as an important prognostic marker for COVID-19 severity. Sci. Rep. 2023, 13, 1460. [Google Scholar] [CrossRef]
- Kartal Baykan, E.; Sebin, E.; Karasahin, O.; Baykan, A.R.; Cerrah, S.; Gogebakan, H.; Sevinc, C.; Kahraman, M.; Yavuz, Y.C. Galectin-3: Can it be a diagnostic tool for pneumonia in COVID-19 patients? Turk. J. Med. Sci. 2021, 51, 2256–2262. [Google Scholar] [CrossRef] [PubMed]
- Kazancioglu, S.; Yilmaz, F.M.; Bastug, A.; Ozbay, B.O.; Aydos, O.; Yucel, C.; Bodur, H.; Yilmaz, G. Assessment of Galectin-1, Galectin-3, and Prostaglandin E2 Levels in Patients with COVID-19. Jpn. J. Infect. Dis. 2021, 74, 530–536. [Google Scholar] [CrossRef]
- Kusnierz-Cabala, B.; Maziarz, B.; Dumnicka, P.; Dembinski, M.; Kapusta, M.; Bociaga-Jasik, M.; Winiarski, M.; Garlicki, A.; Grodzicki, T.; Kukla, M. Diagnostic Significance of Serum Galectin-3 in Hospitalized Patients with COVID-19-A Preliminary Study. Biomolecules 2021, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, S.; Donmez, E.; Yavuz, S.T.; Ziyrek, M.; Ince, O.; Kucuk, H.S.; Tasdemir, Z.A.; Yilmaz, I.; Varol, S.; Sahin, I.; et al. Prognostic significance of serum galectin-3 in hospitalized patients with COVID-19. Cytokine 2022, 158, 155970. [Google Scholar] [CrossRef]
- Portacci, A.; Diaferia, F.; Santomasi, C.; Dragonieri, S.; Boniello, E.; Di Serio, F.; Carpagnano, G.E. Galectin-3 as prognostic biomarker in patients with COVID-19 acute respiratory failure. Respir. Med. 2021, 187, 106556. [Google Scholar] [CrossRef]
- Tawiah, K.; Jackson, L.; Omosule, C.; Ballman, C.; Shahideh, B.; Scott, M.G.; Murtagh, G.; Farnsworth, C.W. Serial cardiac biomarkers for risk stratification of patients with COVID-19. Clin. Biochem. 2022, 107, 24–32. [Google Scholar] [CrossRef]
- Turnic, T.N.; Popadic, V.; Klasnja, S.; Sekulic, A.; Nikolic, N.; Zivkovic, V.; Jeremic, N.; Andjic, M.; Draginic, N.; Srejovic, I.; et al. Bradykinin and Galectin-3 in Survived and Deceased Patients with COVID-19 Pneumonia: An Increasingly Promising Biochemical Target. Oxid. Med. Cell. Longev. 2022, 2022, 7920915. [Google Scholar] [CrossRef]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a key adipokine in obesity related liver diseases. World J. Gastroenterol. 2011, 17, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Walsh, K. Obesity, adiponectin and vascular inflammatory disease. Curr. Opin. Lipidol. 2003, 14, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Turer, A.T.; Scherer, P.E. Adiponectin: Mechanistic insights and clinical implications. Diabetologia 2012, 55, 2319–2326. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adiponectin: Friend or foe in obesity and inflammation. Med. Rev. 2022, 2, 349–362. [Google Scholar] [CrossRef]
- Tian, L.; Luo, N.; Zhu, X.; Chung, B.H.; Garvey, W.T.; Fu, Y. Adiponectin-AdipoR1/2-APPL1 signaling axis suppresses human foam cell formation: Differential ability of AdipoR1 and AdipoR2 to regulate inflammatory cytokine responses. Atherosclerosis 2012, 221, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Wulster-Radcliffe, M.C.; Ajuwon, K.M.; Wang, J.; Christian, J.A.; Spurlock, M.E. Adiponectin differentially regulates cytokines in porcine macrophages. Biochem. Biophys. Res. Commun. 2004, 316, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Kumada, M.; Kihara, S.; Ouchi, N.; Kobayashi, H.; Okamoto, Y.; Ohashi, K.; Maeda, K.; Nagaretani, H.; Kishida, K.; Maeda, N.; et al. Adiponectin specifically increased tissue inhibitor of metalloproteinase-1 through interleukin-10 expression in human macrophages. Circulation 2004, 109, 2046–2049. [Google Scholar] [CrossRef]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Nakamura, T.; Nishida, M.; Kumada, M.; Okamoto, Y.; Ohashi, K.; Nagaretani, H.; Kishida, K.; et al. Reciprocal association of C-reactive protein with adiponectin in blood stream and adipose tissue. Circulation 2003, 107, 671–674. [Google Scholar] [CrossRef]
- Salvator, H.; Grassin-Delyle, S.; Naline, E.; Brollo, M.; Fournier, C.; Couderc, L.J.; Devillier, P. Contrasting Effects of Adipokines on the Cytokine Production by Primary Human Bronchial Epithelial Cells: Inhibitory Effects of Adiponectin. Front. Pharmacol. 2020, 11, 56. [Google Scholar] [CrossRef]
- Abke, S.; Neumeier, M.; Weigert, J.; Wehrwein, G.; Eggenhofer, E.; Schaffler, A.; Maier, K.; Aslanidis, C.; Scholmerich, J.; Buechler, C. Adiponectin-induced secretion of interleukin-6 (IL-6), monocyte chemotactic protein-1 (MCP-1, CCL2) and interleukin-8 (IL-8, CXCL8) is impaired in monocytes from patients with type I diabetes. Cardiovasc. Diabetol. 2006, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Weigert, J.; Obermeier, F.; Neumeier, M.; Wanninger, J.; Filarsky, M.; Bauer, S.; Aslanidis, C.; Rogler, G.; Ott, C.; Schaffler, A.; et al. Circulating levels of chemerin and adiponectin are higher in ulcerative colitis and chemerin is elevated in Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Menzaghi, C.; Trischitta, V. The Adiponectin Paradox for All-Cause and Cardiovascular Mortality. Diabetes 2018, 67, 12–22. [Google Scholar] [CrossRef]
- Walkey, A.J.; Rice, T.W.; Konter, J.; Ouchi, N.; Shibata, R.; Walsh, K.; deBoisblanc, B.P.; Summer, R. Plasma adiponectin and mortality in critically ill subjects with acute respiratory failure. Crit. Care Med. 2010, 38, 2329–2334. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.L.; Hu, J.H.; Pao, L.H.; Lin, M.S.; Kuo, C.J.; Chen, S.C.; Fan, C.M.; Chang, M.Y.; Chien, R.N. Critical role of triglycerides for adiponectin levels in hepatitis C: A joint study of human and HCV core transgenic mice. BMC Immunol. 2021, 22, 54. [Google Scholar] [CrossRef]
- Durazzo, M.; Belci, P.; Niro, G.; Collo, A.; Grisoglio, E.; Ambrogio, V.; Spandre, M.; Fontana, R.; Gambino, R.; Cassader, M.; et al. Variations of serum levels of adiponectin and resistin in chronic viral hepatitis. J. Endocrinol. Investig. 2013, 36, 600–605. [Google Scholar] [CrossRef]
- Tsatsanis, C.; Margioris, A.N.; Kontoyiannis, D.P. Association between H1N1 infection severity and obesity-adiponectin as a potential etiologic factor. J. Infect. Dis. 2010, 202, 459–460. [Google Scholar] [CrossRef]
- Spirina, L.V.; Masunov, V.N.; Dyakov, D.A.; Akbasheva, O.E.; Kebekbayeva, A.Y.; Shuvalov, I.Y.; Masunova, N.V.; Kovaleva, I.V.; Dagbaeva, Y. SARS-CoV-2 Induced Biochemical Mechanisms in Liver Damage and Intestinal Lesions. Indian. J. Clin. Biochem. 2022, 12. [Google Scholar] [CrossRef]
- Kearns, S.M.; Ahern, K.W.; Patrie, J.T.; Horton, W.B.; Harris, T.E.; Kadl, A. Reduced adiponectin levels in patients with COVID-19 acute respiratory failure: A case-control study. Physiol. Rep. 2021, 9, e14843. [Google Scholar] [CrossRef]
- Perez-Perez, A.; Vilarino-Garcia, T.; Fernandez-Riejos, P.; Martin-Gonzalez, J.; Segura-Egea, J.J.; Sanchez-Margalet, V. Role of leptin as a link between metabolism and the immune system. Cytokine Growth Factor. Rev. 2017, 35, 71–84. [Google Scholar] [CrossRef]
- Muskiet, F.A.J.; Carrera-Bastos, P.; Pruimboom, L.; Lucia, A.; Furman, D. Obesity and Leptin Resistance in the Regulation of the Type I Interferon Early Response and the Increased Risk for Severe COVID-19. Nutrients 2022, 14, 1388. [Google Scholar] [CrossRef]
- Tschop, J.; Nogueiras, R.; Haas-Lockie, S.; Kasten, K.R.; Castaneda, T.R.; Huber, N.; Guanciale, K.; Perez-Tilve, D.; Habegger, K.; Ottaway, N.; et al. CNS leptin action modulates immune response and survival in sepsis. J. Neurosci. 2010, 30, 6036–6047. [Google Scholar] [CrossRef]
- Mastronardi, C.A.; Yu, W.H.; Srivastava, V.K.; Dees, W.L.; McCann, S.M. Lipopolysaccharide-induced leptin release is neurally controlled. Proc. Natl. Acad. Sci. USA 2001, 98, 14720–14725. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Preas, H.L.; Chrousos, G.P.; Suffredini, A.F. Circulating leptin levels during acute experimental endotoxemia and antiinflammatory therapy in humans. J. Infect. Dis. 1998, 178, 887–890. [Google Scholar] [CrossRef]
- Landman, R.E.; Puder, J.J.; Xiao, E.; Freda, P.U.; Ferin, M.; Wardlaw, S.L. Endotoxin stimulates leptin in the human and nonhuman primate. J. Clin. Endocrinol. Metab. 2003, 88, 1285–1291. [Google Scholar] [CrossRef]
- Jacobsson, S.; Larsson, P.; Johansson, G.; Norberg, M.; Wadell, G.; Hallmans, G.; Winso, O.; Soderberg, S. Leptin independently predicts development of sepsis and its outcome. J. Inflamm. 2017, 14, 19. [Google Scholar] [CrossRef]
- Karampela, I.; Kandri, E.; Chrysanthopoulou, E.; Skyllas, G.; Christodoulatos, G.S.; Antonakos, G.; Vogiatzakis, E.; Armaganidis, A.; Dalamaga, M. Circulating total leptin as a diagnostic and prognostic biomarker in sepsis. Eur. Respir. J. 2019, 54, PA2177. [Google Scholar] [CrossRef]
- Singh, R.; Hemati, H.; Bajpai, M.; Yadav, P.; Maheshwari, A.; Kumar, S.; Agrawal, S.; Sevak, J.K.; Islam, M.; Mars, J.S.; et al. Sustained expression of inflammatory monocytes and activated T cells in COVID-19 patients and recovered convalescent plasma donors. Immun. Inflamm. Dis. 2021, 9, 1279–1290. [Google Scholar] [CrossRef]
- Wang, J.; Xu, Y.; Zhang, X.; Wang, S.; Peng, Z.; Guo, J.; Jiang, H.; Liu, J.; Xie, Y.; Wang, J.; et al. Leptin correlates with monocytes activation and severe condition in COVID-19 patients. J. Leukoc. Biol. 2021, 110, 9–20. [Google Scholar] [CrossRef]
- Rocco, J.M.; Laghetti, P.; Di Stefano, M.; Sereti, I.; Ortega-Villa, A.; Wang, J.; Rupert, A.; Chironna, M.; Ye, L.; Liu, X.; et al. Impact of Innate Immunity, Endothelial Damage, and Metabolic Biomarkers on COVID-19 Severity and Mortality. Open. Forum Infect. Dis. 2022, 9, ofac427. [Google Scholar] [CrossRef]
- van der Voort, P.H.J.; Moser, J.; Zandstra, D.F.; Muller Kobold, A.C.; Knoester, M.; Calkhoven, C.F.; Hamming, I.; van Meurs, M. Leptin levels in SARS-CoV-2 infection related respiratory failure: A cross-sectional study and a pathophysiological framework on the role of fat tissue. Heliyon 2020, 6, e04696. [Google Scholar] [CrossRef]
- Khreefa, Z.; Barbier, M.T.; Koksal, A.R.; Love, G.; Del Valle, L. Pathogenesis and Mechanisms of SARS-CoV-2 Infection in the Intestine, Liver, and Pancreas. Cells 2023, 12, 262. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Catalan, V.; Rodriguez, A.; Gomez-Ambrosi, J. Adiponectin-leptin ratio: A promising index to estimate adipose tissue dysfunction. Relation with obesity-associated cardiometabolic risk. Adipocyte 2018, 7, 57–62. [Google Scholar] [CrossRef]
- Park, H.K.; Ahima, R.S. Resistin in rodents and humans. Diabetes Metab. J. 2013, 37, 404–414. [Google Scholar] [CrossRef]
- Ghosh, S.; Singh, A.K.; Aruna, B.; Mukhopadhyay, S.; Ehtesham, N.Z. The genomic organization of mouse resistin reveals major differences from the human resistin: Functional implications. Gene 2003, 305, 27–34. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Q.; Cai, D.; Guo, H.; Fang, J.; Cui, H.; Gou, L.; Deng, J.; Wang, Z.; Zuo, Z. Resistin, a Novel Host Defense Peptide of Innate Immunity. Front. Immunol. 2021, 12, 699807. [Google Scholar] [CrossRef]
- Bostrom, E.A.; Tarkowski, A.; Bokarewa, M. Resistin is stored in neutrophil granules being released upon challenge with inflammatory stimuli. Biochim. Biophys. Acta 2009, 1793, 1894–1900. [Google Scholar] [CrossRef]
- Mantula, P.S.; Outinen, T.K.; Jaatinen, P.; Hamalainen, M.; Huhtala, H.; Porsti, I.H.; Vaheri, A.; Mustonen, J.T.; Makela, S.M. High plasma resistin associates with severe acute kidney injury in Puumala hantavirus infection. PLoS ONE 2018, 13, e0208017. [Google Scholar] [CrossRef]
- Meng, Z.; Zhang, Y.; Wei, Z.; Liu, P.; Kang, J.; Zhang, Y.; Ma, D.; Ke, C.; Chen, Y.; Luo, J.; et al. High serum resistin associates with intrahepatic inflammation and necrosis: An index of disease severity for patients with chronic HBV infection. BMC Gastroenterol. 2017, 17, 6. [Google Scholar] [CrossRef]
- Morace, C.; Spadaro, A.; Cucunato, M.; Tortorella, V.; Consolo, P.; Luigiano, C.; Stabile, G.; Bonfiglio, C.; Bellerone, R.; Fortiguerra, A.; et al. High serum resistin in chronic viral hepatitis is not a marker of metabolic disorder. Hepatogastroenterology 2010, 57, 1215–1219. [Google Scholar]
- Mera, K.; Uto, H.; Mawatari, S.; Ido, A.; Yoshimine, Y.; Nosaki, T.; Oda, K.; Tabu, K.; Kumagai, K.; Tamai, T.; et al. Serum levels of apoptosis inhibitor of macrophage are associated with hepatic fibrosis in patients with chronic hepatitis C. BMC Gastroenterol. 2014, 14, 27. [Google Scholar] [CrossRef]
- Chang, M.L.; Liang, K.H.; Ku, C.L.; Lo, C.C.; Cheng, Y.T.; Hsu, C.M.; Yeh, C.T.; Chiu, C.T. Resistin reinforces interferon lambda-3 to eliminate hepatitis C virus with fine-tuning from RETN single-nucleotide polymorphisms. Sci. Rep. 2016, 6, 30799. [Google Scholar] [CrossRef]
- Solbach, P.; Wedemeyer, H. The New Era of Interferon-Free Treatment of Chronic Hepatitis C. Viszeralmedizin 2015, 31, 290–296. [Google Scholar] [CrossRef]
- Ebihara, T.; Matsumoto, H.; Matsubara, T.; Matsuura, H.; Hirose, T.; Shimizu, K.; Ogura, H.; Kang, S.; Tanaka, T.; Shimazu, T. Adipocytokine Profile Reveals Resistin Forming a Prognostic-Related Cytokine Network in the Acute Phase of Sepsis. Shock 2021, 56, 718–726. [Google Scholar] [CrossRef]
- Meizlish, M.L.; Pine, A.B.; Bishai, J.D.; Goshua, G.; Nadelmann, E.R.; Simonov, M.; Chang, C.H.; Zhang, H.; Shallow, M.; Bahel, P.; et al. A neutrophil activation signature predicts critical illness and mortality in COVID-19. Blood Adv. 2021, 5, 1164–1177. [Google Scholar] [CrossRef]
- Menini, S.; Iacobini, C.; Blasetti Fantauzzi, C.; Pesce, C.M.; Pugliese, G. Role of Galectin-3 in Obesity and Impaired Glucose Homeostasis. Oxid. Med. Cell. Longev. 2016, 2016, 9618092. [Google Scholar] [CrossRef]
- Henderson, N.C.; Sethi, T. The regulation of inflammation by galectin-3. Immunol. Rev. 2009, 230, 160–171. [Google Scholar] [CrossRef]
- Wang, W.H.; Lin, C.Y.; Chang, M.R.; Urbina, A.N.; Assavalapsakul, W.; Thitithanyanont, A.; Chen, Y.H.; Liu, F.T.; Wang, S.F. The role of galectins in virus infection—A systemic literature review. J. Microbiol. Immunol. Infect. 2020, 53, 925–935. [Google Scholar] [CrossRef]
- Weigert, J.; Neumeier, M.; Wanninger, J.; Bauer, S.; Farkas, S.; Scherer, M.N.; Schnitzbauer, A.; Schaffler, A.; Aslanidis, C.; Scholmerich, J.; et al. Serum galectin-3 is elevated in obesity and negatively correlates with glycosylated hemoglobin in type 2 diabetes. J. Clin. Endocrinol. Metab. 2010, 95, 1404–1411. [Google Scholar] [CrossRef] [PubMed]
- Humphries, D.C.; Mills, R.; Dobie, R.; Henderson, N.C.; Sethi, T.; Mackinnon, A.C. Selective Myeloid Depletion of Galectin-3 Offers Protection Against Acute and Chronic Lung Injury. Front. Pharmacol. 2021, 12, 715986. [Google Scholar] [CrossRef]
- Laforge, M.; Elbim, C.; Frere, C.; Hemadi, M.; Massaad, C.; Nuss, P.; Benoliel, J.J.; Becker, C. Tissue damage from neutrophil-induced oxidative stress in COVID-19. Nat. Rev. Immunol. 2020, 20, 515–516. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; Mainardi, A.; d’Alessandro, M.; Cameli, P.; Bennett, D.; Bargagli, E.; Sestini, P. Common Molecular Pathways Between Post-COVID19 Syndrome and Lung Fibrosis: A Scoping Review. Front. Pharmacol. 2022, 13, 748931. [Google Scholar] [CrossRef]
- Weigand, K.; Peschel, G.; Grimm, J.; Muller, M.; Buechler, C. Serum Galectin-3 in Hepatitis C Virus Infection Declines after Successful Virus Eradication by Direct-Acting Antiviral Therapy. J. Gastrointestin. Liver Dis. 2022, 31, 444–452. [Google Scholar] [CrossRef]
- Nishi, Y.; Sano, H.; Kawashima, T.; Okada, T.; Kuroda, T.; Kikkawa, K.; Kawashima, S.; Tanabe, M.; Goto, T.; Matsuzawa, Y.; et al. Role of galectin-3 in human pulmonary fibrosis. Allergol. Int. 2007, 56, 57–65. [Google Scholar] [CrossRef]
- Chen, P.K.; Lan, J.L.; Huang, P.H.; Hsu, J.L.; Chang, C.K.; Tien, N.; Lin, H.J.; Chen, D.Y. Interleukin-18 Is a Potential Biomarker to Discriminate Active Adult-Onset Still’s Disease from COVID-19. Front. Immunol. 2021, 12, 719544. [Google Scholar] [CrossRef]
- Gaughan, E.E.; Quinn, T.M.; Mills, A.; Bruce, A.M.; Antonelli, J.; MacKinnon, A.C.; Aslanis, V.; Li, F.; O’Connor, R.; Boz, C.; et al. An Inhaled Galectin-3 Inhibitor in COVID-19 Pneumonitis: A Phase Ib/IIa Randomized Controlled Clinical Trial (DEFINE). Am. J. Respir. Crit. Care Med. 2023, 207, 138–149. [Google Scholar] [CrossRef]
- Karsli, E.; Anabarli Metin, D.; Canacik, O.; Sabirli, R.; Kaymaz, B.; Kurt, O.; Koseler, A. Galectin-3 as a Potential Prognostic Biomarker for COVID-19 Disease: A Case-Control Study. Cureus 2022, 14, e28805. [Google Scholar] [CrossRef]
- Audrito, V.; Messana, V.G.; Deaglio, S. NAMPT and NAPRT: Two Metabolic Enzymes with Key Roles in Inflammation. Front. Oncol. 2020, 10, 358. [Google Scholar] [CrossRef]
- Buechler, C.; Haberl, E.M.; Rein-Fischboeck, L.; Aslanidis, C. Adipokines in Liver Cirrhosis. Int. J. Mol. Sci. 2017, 18, 1392. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.L.; Men, R.; Zhu, Y.; Yuan, C.; Wei, Y.; Liu, X.; Yang, L. Visfatin: An adipokine activator of rat hepatic stellate cells. Mol. Med. Rep. 2015, 11, 1073–1078. [Google Scholar] [CrossRef]
- Sawicka, K.; Michalska-Jakubus, M.; Potembska, E.; Kowal, M.; Pietrzak, A.; Krasowska, D. Visfatin and chemerin levels correspond with inflammation and might reflect the bridge between metabolism, inflammation and fibrosis in patients with systemic sclerosis. Postepy Dermatol. Alergol. 2019, 36, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Chang, D.M.; Lin, K.C.; Shin, S.J.; Lee, Y.J. Visfatin in overweight/obesity, type 2 diabetes mellitus, insulin resistance, metabolic syndrome and cardiovascular diseases: A meta-analysis and systemic review. Diabetes Metab. Res. Rev. 2011, 27, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Berndt, J.; Kloting, N.; Kralisch, S.; Kovacs, P.; Fasshauer, M.; Schon, M.R.; Stumvoll, M.; Bluher, M. Plasma visfatin concentrations and fat depot-specific mRNA expression in humans. Diabetes 2005, 54, 2911–2916. [Google Scholar] [CrossRef]
- Quijada, H.; Bermudez, T.; Kempf, C.L.; Valera, D.G.; Garcia, A.N.; Camp, S.M.; Song, J.H.; Franco, E.; Burt, J.K.; Sun, B.; et al. Endothelial eNAMPT amplifies pre-clinical acute lung injury: Efficacy of an eNAMPT-neutralising monoclonal antibody. Eur. Respir. J. 2021, 57, 2002536. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, T.; Sammani, S.; Song, J.H.; Hernon, V.R.; Kempf, C.L.; Garcia, A.N.; Burt, J.; Hufford, M.; Camp, S.M.; Cress, A.E.; et al. eNAMPT neutralization reduces preclinical ARDS severity via rectified NFkB and Akt/mTORC2 signaling. Sci. Rep. 2022, 12, 696. [Google Scholar] [CrossRef]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin-APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 820002. [Google Scholar] [CrossRef]
- Than, A.; He, H.L.; Chua, S.H.; Xu, D.; Sun, L.; Leow, M.K.; Chen, P. Apelin Enhances Brown Adipogenesis and Browning of White Adipocytes. J. Biol. Chem. 2015, 290, 14679–14691. [Google Scholar] [CrossRef]
- Oz, M.; Lorke, D.E. Multifunctional angiotensin converting enzyme 2, the SARS-CoV-2 entry receptor, and critical appraisal of its role in acute lung injury. Biomed. Pharmacother. 2021, 136, 111193. [Google Scholar] [CrossRef]
- Wang, W.; McKinnie, S.M.; Farhan, M.; Paul, M.; McDonald, T.; McLean, B.; Llorens-Cortes, C.; Hazra, S.; Murray, A.G.; Vederas, J.C.; et al. Angiotensin-Converting Enzyme 2 Metabolizes and Partially Inactivates Pyr-Apelin-13 and Apelin-17: Physiological Effects in the Cardiovascular System. Hypertension 2016, 68, 365–377. [Google Scholar] [CrossRef]
- Park, J.; Park, M.Y.; Kim, Y.; Jun, Y.; Lee, U.; Oh, C.M. Apelin as a new therapeutic target for COVID-19 treatment. QJM Int. J. Med. 2023, 116, 197–204. [Google Scholar] [CrossRef]
- Salles, E.L.; Khodadadi, H.; Jarrahi, A.; Ahluwalia, M.; Paffaro, V.A., Jr.; Costigliola, V.; Yu, J.C.; Hess, D.C.; Dhandapani, K.M.; Baban, B. Cannabidiol (CBD) modulation of apelin in acute respiratory distress syndrome. J. Cell. Mol. Med. 2020, 24, 12869–12872. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Toprak, S.; Ortmann, O.; Buechler, C.; Treeck, O. The Complex Roles of Adipokines in Polycystic Ovary Syndrome and Endometriosis. Biomedicines 2022, 10, 2503. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Song, Y.; Xie, H.; Dong, M. An update on brown adipose tissue and obesity intervention: Function, regulation and therapeutic implications. Front. Endocrinol. 2022, 13, 1065263. [Google Scholar] [CrossRef] [PubMed]
- Erdem, G.; Dogru, T.; Tasci, I.; Sonmez, A.; Tapan, S. Low plasma apelin levels in newly diagnosed type 2 diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2008, 116, 289–292. [Google Scholar] [CrossRef]
- Sonmez, A.; Celebi, G.; Erdem, G.; Tapan, S.; Genc, H.; Tasci, I.; Ercin, C.N.; Dogru, T.; Kilic, S.; Uckaya, G.; et al. Plasma apelin and ADMA Levels in patients with essential hypertension. Clin. Exp. Hypertens. 2010, 32, 179–183. [Google Scholar] [CrossRef]
- Rostamzadeh, F.; Najafipour, H.; Yazdani, R.; Nakhaei, S.; Alinaghi Langari, A. Changes in serum levels of apelin and nitric oxide in hospitalized patients with COVID-19: Association with hypertension, diabetes, obesity, and severity of disease. Eur. J. Med. Res. 2022, 27, 243. [Google Scholar] [CrossRef]
- Tsiotra, P.C.; Boutati, E.; Dimitriadis, G.; Raptis, S.A. High insulin and leptin increase resistin and inflammatory cytokine production from human mononuclear cells. Biomed. Res. Int. 2013, 2013, 487081. [Google Scholar] [CrossRef]
- Martinez-Martinez, E.; Jurado-Lopez, R.; Valero-Munoz, M.; Bartolome, M.V.; Ballesteros, S.; Luaces, M.; Briones, A.M.; Lopez-Andres, N.; Miana, M.; Cachofeiro, V. Leptin induces cardiac fibrosis through galectin-3, mTOR and oxidative stress: Potential role in obesity. J. Hypertens. 2014, 32, 1104–1114; discussion 1114. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Adipokine | Elevated in COVID-19 | Related to Disease Severity | Prognostic for Admission to Intensive Care | Prognostic for Survival | References |
|---|---|---|---|---|---|
| Chemerin | ? | No | No | No | [62,63] |
| Adiponectin | No | No | No | No | [65,66,67,68,69,70,71] |
| Leptin | Yes | No | No | No | [63,67,68,69,71,72,73,74] |
| Adiponectin/Leptin | Yes | ? | Yes | Yes | [68,71] |
| Resistin | Yes | Yes | Yes | Yes | [63,67,74,75,76,77] |
| Galectin-3 | Yes | Yes | Yes | Yes | [75,78,79,80,81,82,83,84,85,86,87,88,89,90] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grewal, T.; Buechler, C. Adipokines as Diagnostic and Prognostic Markers for the Severity of COVID-19. Biomedicines 2023, 11, 1302. https://doi.org/10.3390/biomedicines11051302
Grewal T, Buechler C. Adipokines as Diagnostic and Prognostic Markers for the Severity of COVID-19. Biomedicines. 2023; 11(5):1302. https://doi.org/10.3390/biomedicines11051302
Chicago/Turabian StyleGrewal, Thomas, and Christa Buechler. 2023. "Adipokines as Diagnostic and Prognostic Markers for the Severity of COVID-19" Biomedicines 11, no. 5: 1302. https://doi.org/10.3390/biomedicines11051302
APA StyleGrewal, T., & Buechler, C. (2023). Adipokines as Diagnostic and Prognostic Markers for the Severity of COVID-19. Biomedicines, 11(5), 1302. https://doi.org/10.3390/biomedicines11051302