
Vitamin K and Calcium Chelation in Vascular Health



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
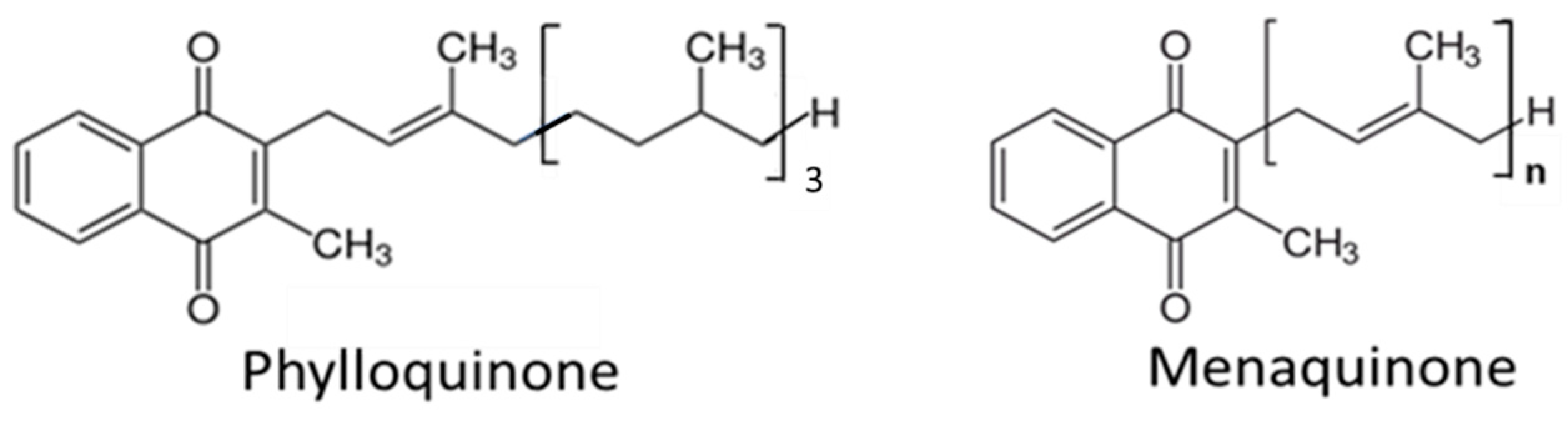
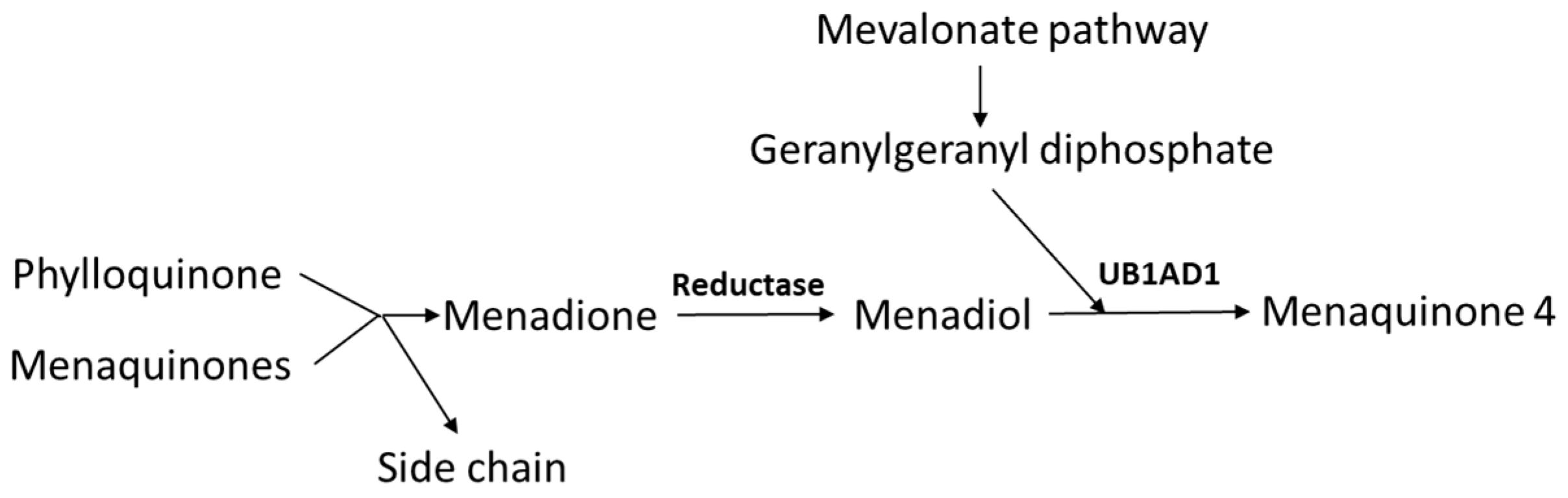
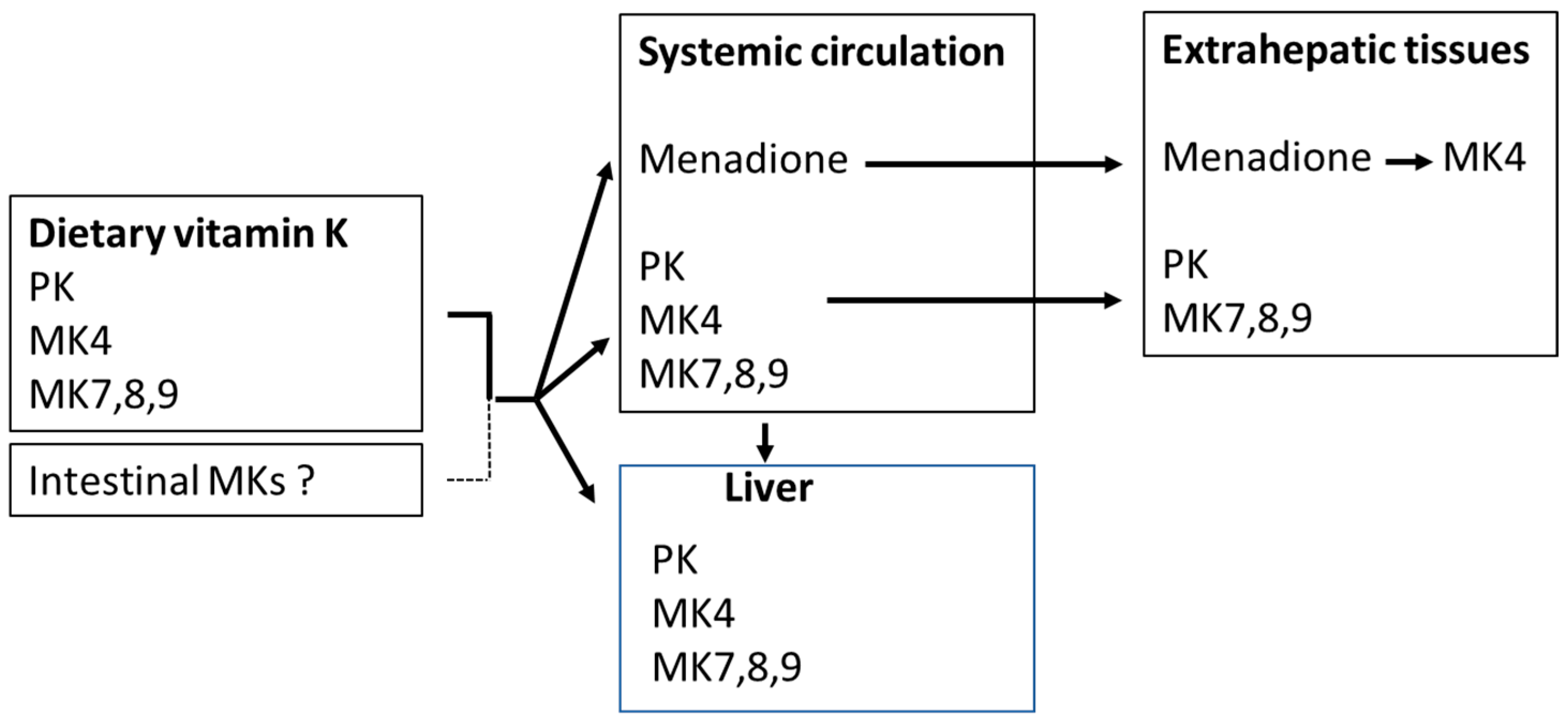
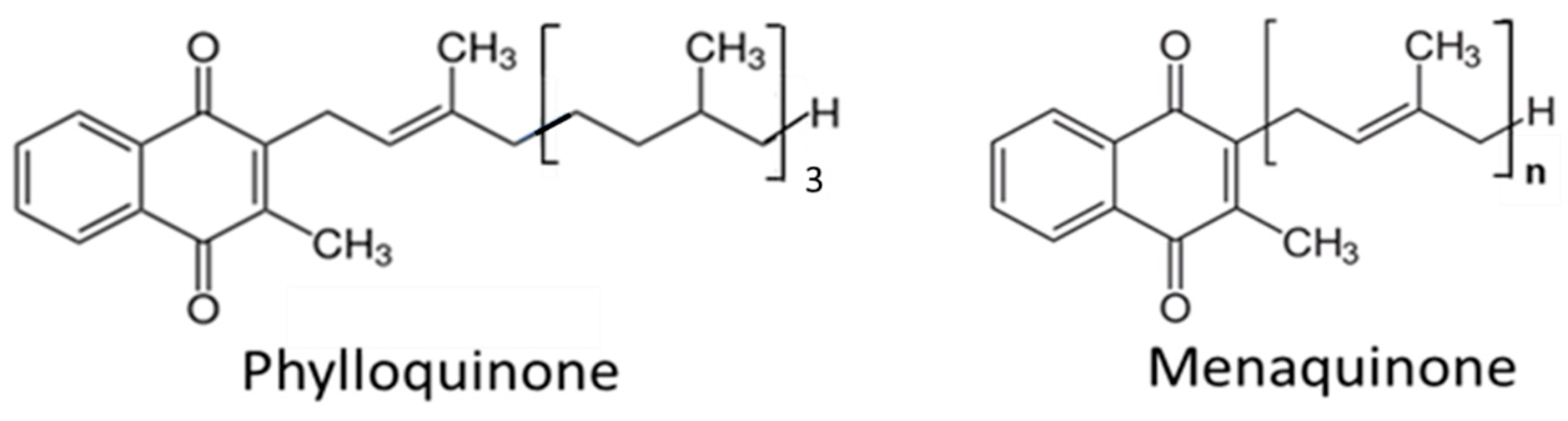
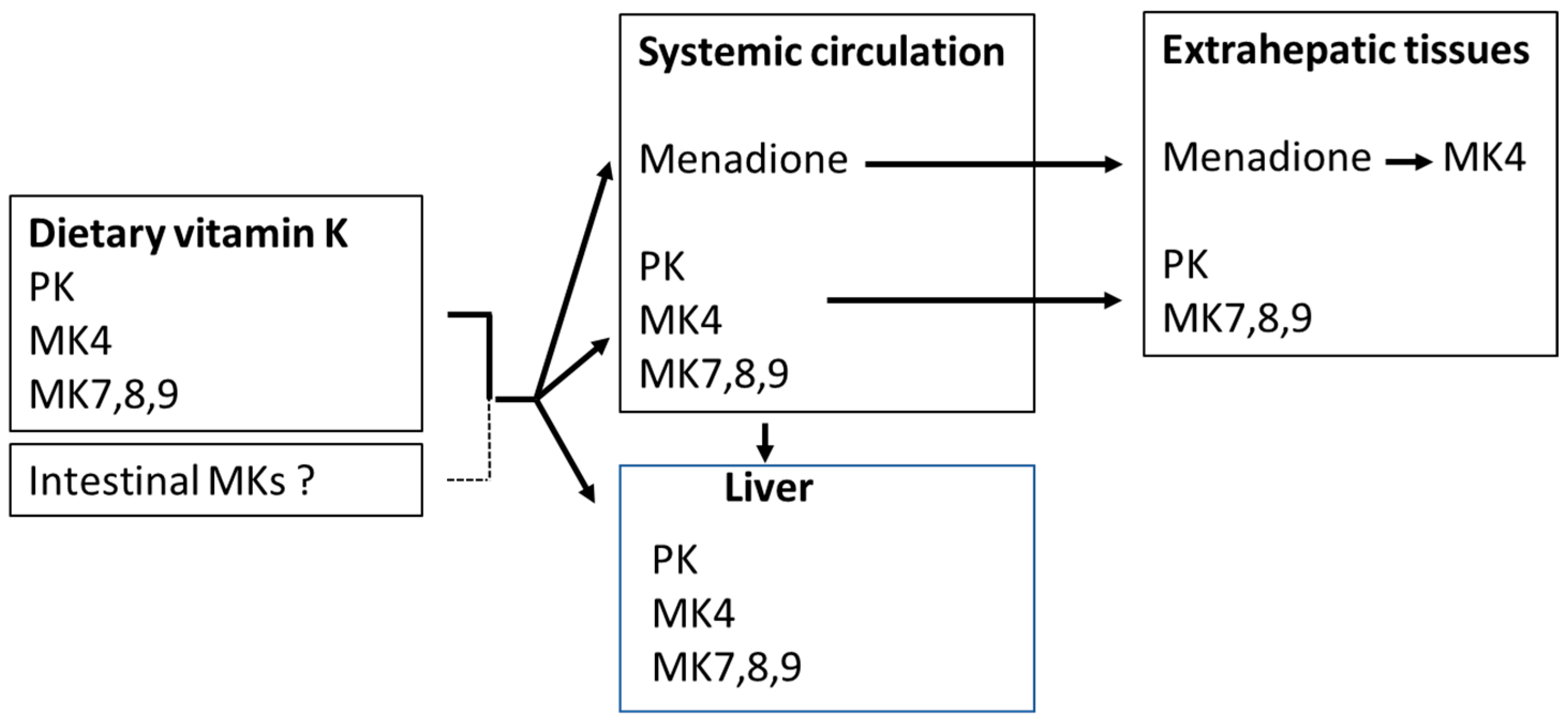
2. Sources and Kinetics of Vitamin K
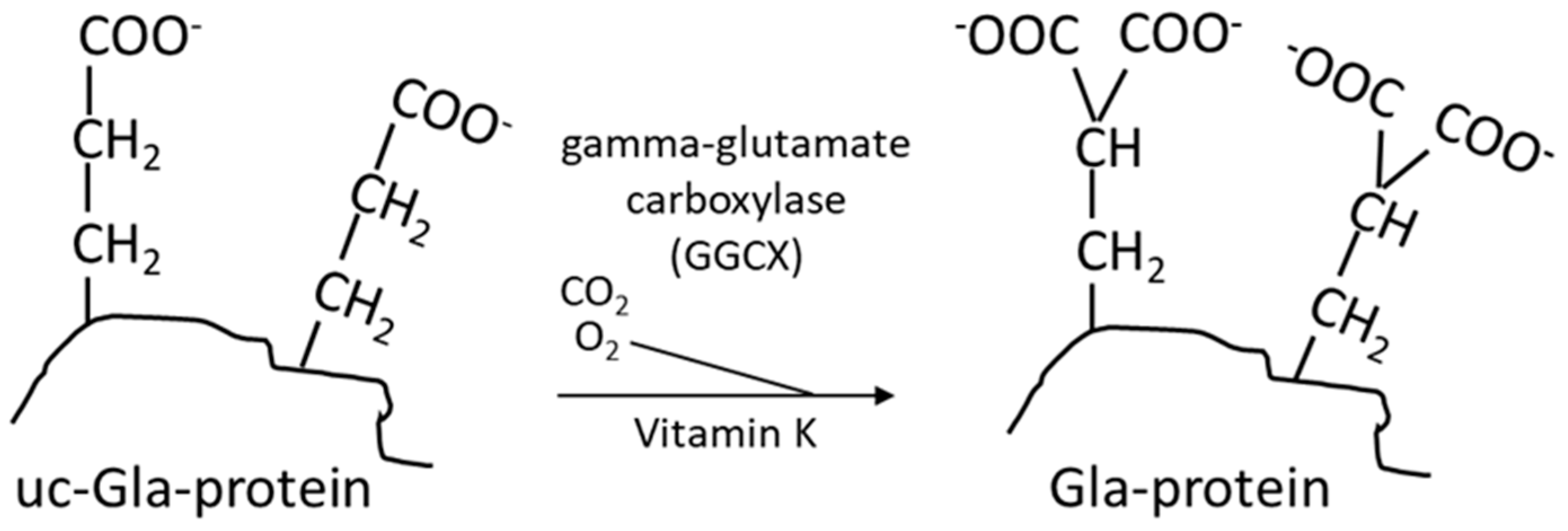
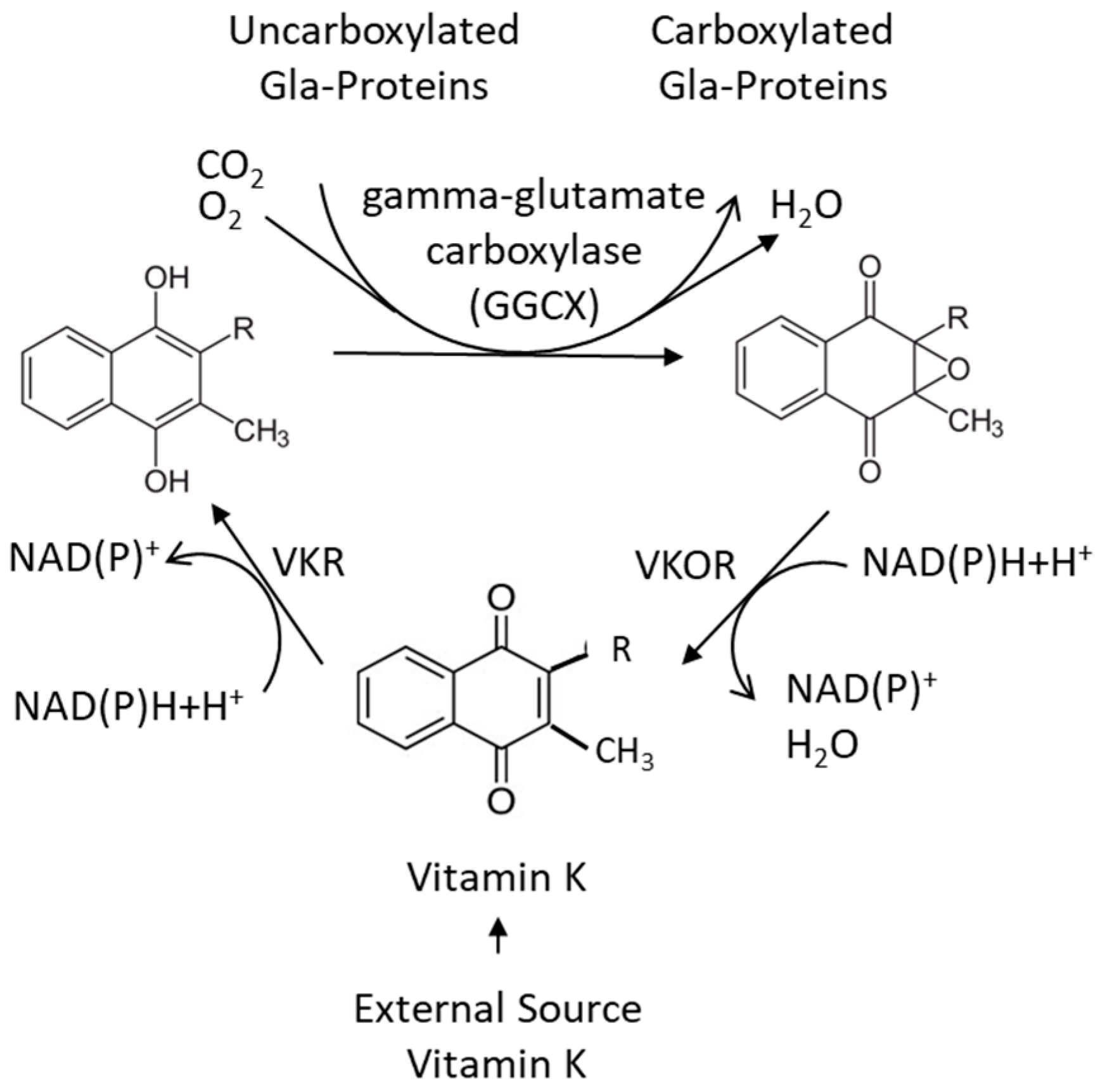
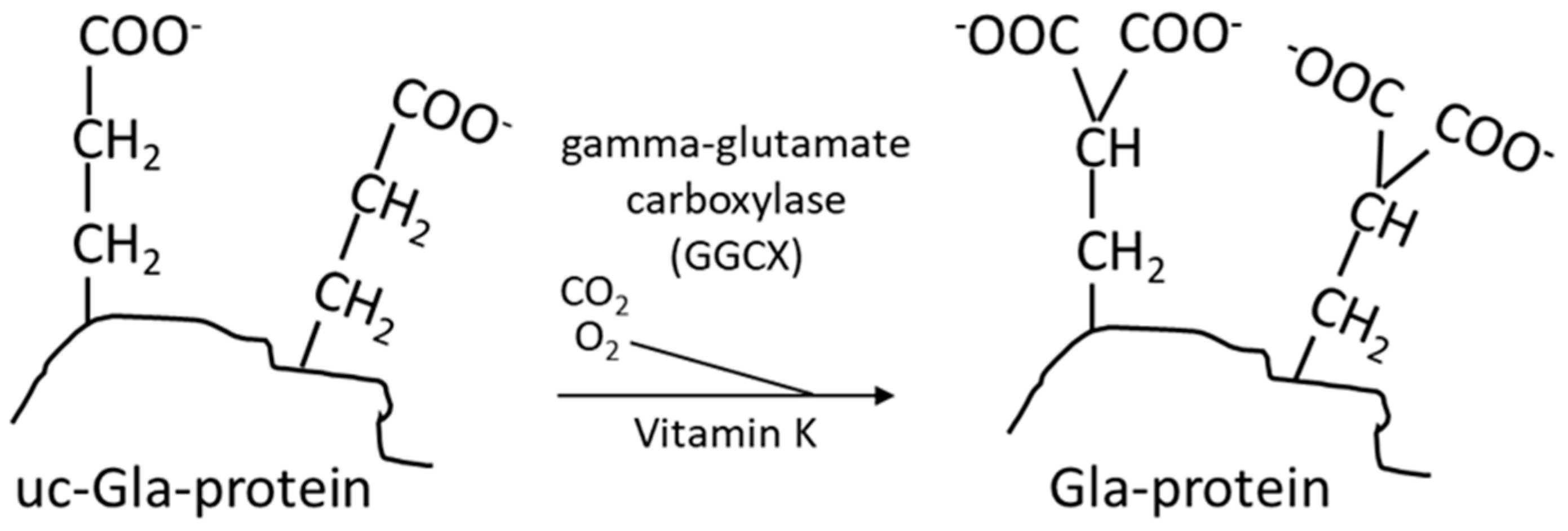
3. The Vitamin-K-Dependent Carboxylation, the Vitamin K Cycle, and Pharmaceuticals Affecting Vitamin K

4. Vitamin K, Human Steroid and Xenobiotic Receptor (SXR), and Inflammation
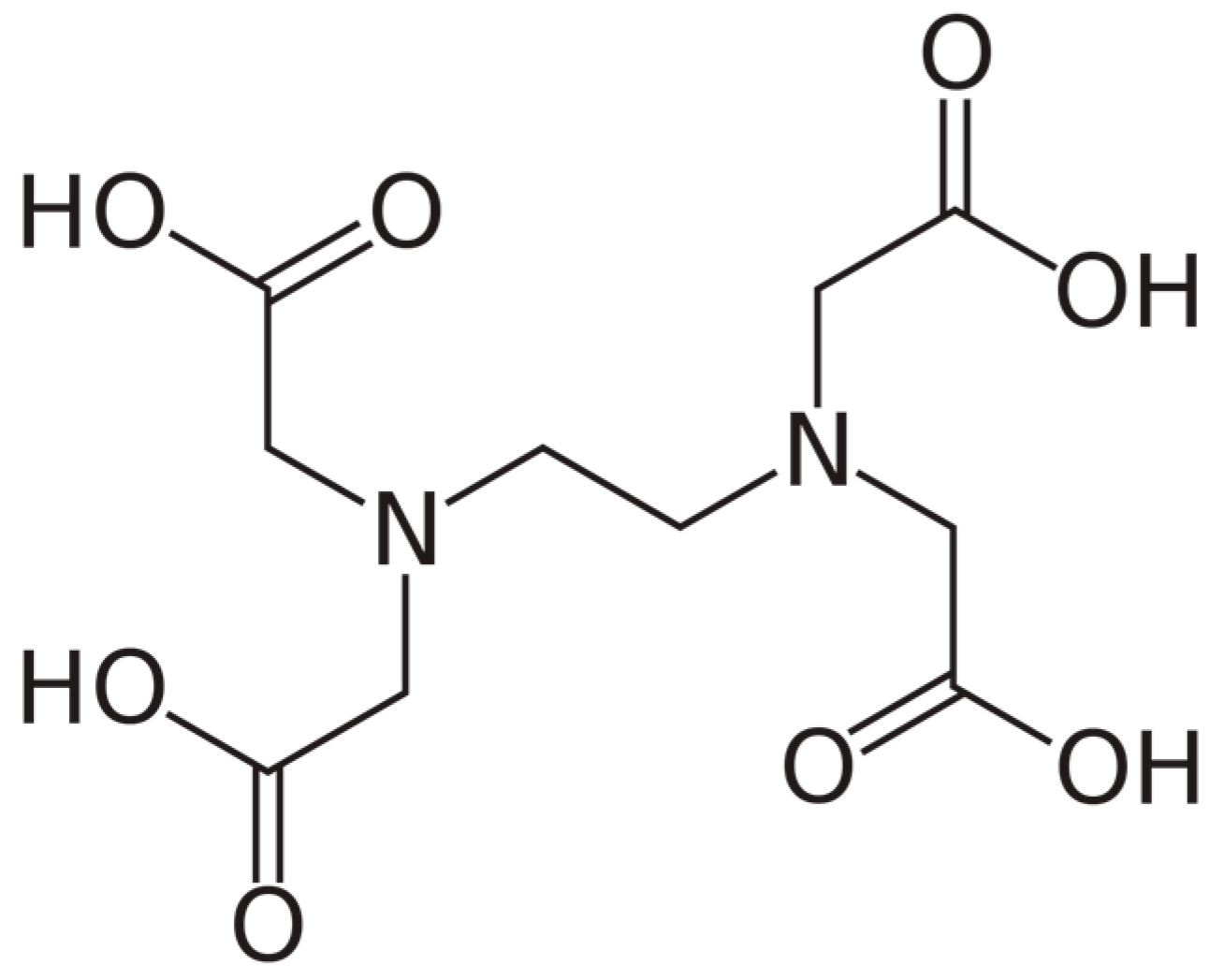
5. Calcium Chelation Therapy—With Endogenous or Exogenous Chelators
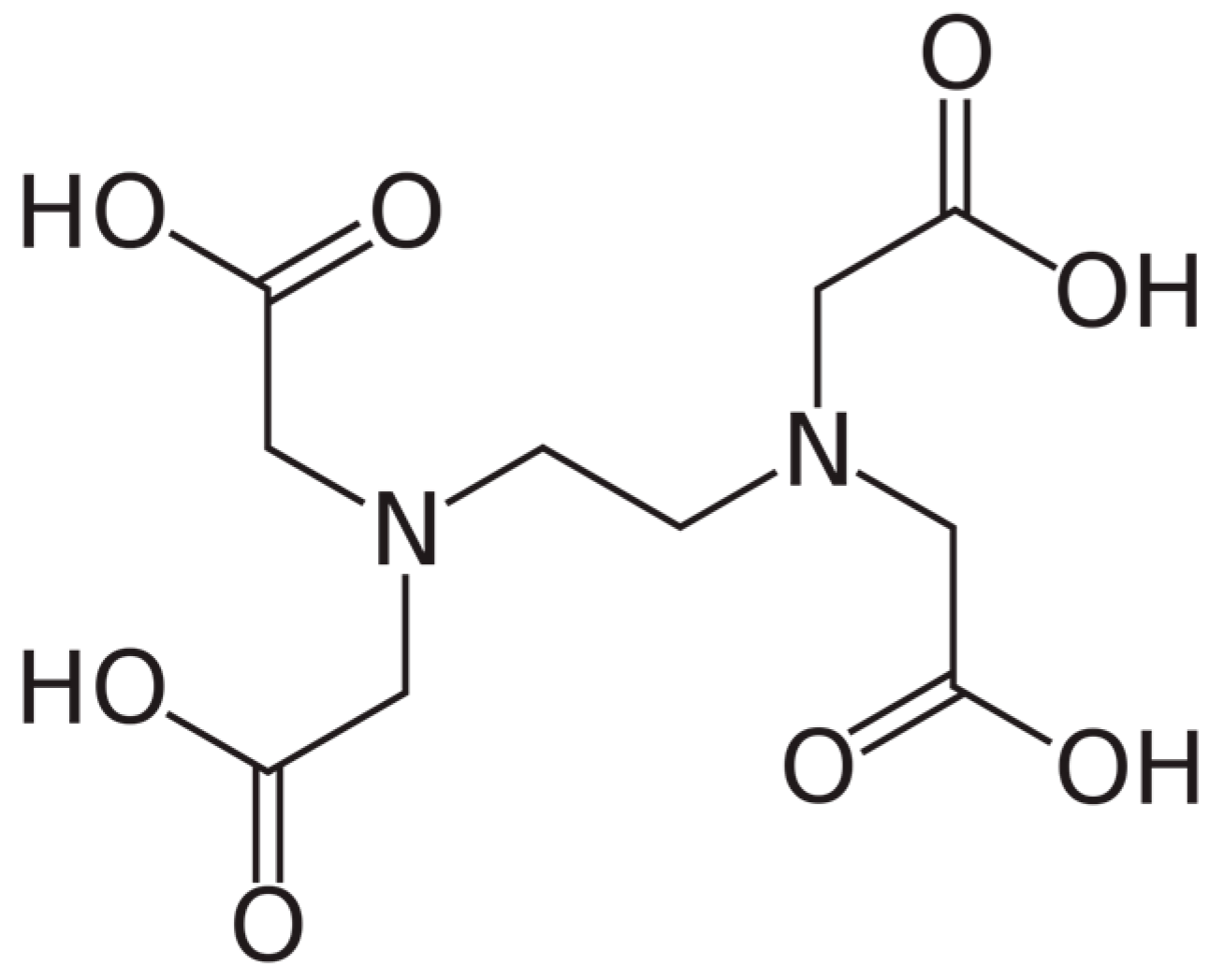
6. The Use of EDTA Chelation Therapy in the Context of Vascular Health
7. Clinical Evaluations of Vitamin K Supplementation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef]
- Kalra, S.S.; Shanahan, C.M. Vascular calcification and hypertension: Cause and effect. Ann. Med. 2012, 44 (Suppl. S1), S85–S92. [Google Scholar] [CrossRef]
- Greenland, P.; LaBree, L.; Azen, S.P.; Doherty, T.M.; Detrano, R.C. Coronary artery calcium score combined with Framingham score for risk prediction in asymptomatic individuals. JAMA 2004, 291, 210–215. [Google Scholar] [CrossRef]
- Demer, L.L.; Tintut, Y. Vascular calcification: Pathobiology of a multifaceted disease. Circulation 2008, 117, 2938–2948. [Google Scholar] [CrossRef]
- Grier, M.T.; Meyers, D.G. So much writing, so little science: A review of 37 years of literature on edetate sodium chelation therapy. Ann. Pharmacother. 1993, 27, 1504–1509. [Google Scholar] [CrossRef]
- Aaseth, J.; Crisponi, G.; Andersen, O. (Eds.) Chelation Therapy in the Treatment of Metal Intoxication; Academic Press: London, UK, 2016. [Google Scholar]
- Crisponi, G.; Nurchi, V.M.; Lachowicz, J.I.; Crespo-Alonso, M.; Zoroddu, M.A.; Peana, M. Kill or cure: Misuse of chelation therapyfor human diseases. Coord. Chem. Rev. 2015, 284, 278–285. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Spronk, H.M.; Skepper, J.N.; Hackeng, T.M.; Shanahan, C.M.; Vermeer, C.; Weissberg, P.L.; Proudfoot, D. Post-translational modifications regulate matrix Gla protein function: Importance for inhibition of vascular smooth muscle cell calcification. J. Thromb. Haemost. 2007, 5, 2503–2511. [Google Scholar] [CrossRef]
- Bjorklund, G.; Svanberg, E.; Dadar, M.; Card, D.J.; Chirumbolo, S.; Harrington, D.J.; Aaseth, J. The Role of Matrix Gla Protein (MGP) in Vascular Calcification. Curr. Med. Chem. 2020, 27, 1647–1660. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Uitto, J.; Reutelingsperger, C.P. Vitamin K-dependent carboxylation of matrix Gla-protein: A crucial switch to control ectopic mineralization. Trends Mol. Med. 2013, 19, 217–226. [Google Scholar] [CrossRef]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2016, 82, 42–49. [Google Scholar] [CrossRef]
- Wen, L.; Chen, J.; Duan, L.; Li, S. Vitamin K-dependent proteins involved in bone and cardiovascular health (Review). Mol. Med. Rep. 2018, 18, 3–15. [Google Scholar] [CrossRef]
- Alehagen, U.; Dahlstrom, U.; Lindahl, T.L. Low plasma concentrations of coagulation factors II, VII and XI indicate increased risk among elderly with symptoms of heart failure. Blood Coagul. Fibrinolysis 2010, 21, 62–69. [Google Scholar] [CrossRef]
- Chinetti, G.; Neels, J.G. Roles of Nuclear Receptors in Vascular Calcification. Int. J. Mol. Sci. 2021, 22, 6491. [Google Scholar] [CrossRef]
- Sheng, L.; Cao, W.; Cha, B.; Chen, Z.; Wang, F.; Liu, J. Serum osteocalcin level and its association with carotid atherosclerosis in patients with type 2 diabetes. Cardiovasc. Diabetol. 2013, 12, 22. [Google Scholar] [CrossRef]
- Halder, M.; Petsophonsakul, P.; Akbulut, A.C.; Pavlic, A.; Bohan, F.; Anderson, E.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K: Double Bonds beyond Coagulation Insights into Differences between Vitamin K1 and K2 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 896. [Google Scholar] [CrossRef]
- Kalampogias, A.; Siasos, G.; Oikonomou, E.; Tsalamandris, S.; Mourouzis, K.; Tsigkou, V.; Vavuranakis, M.; Zografos, T.; Deftereos, S.; Stefanadis, C.; et al. Basic Mechanisms in Atherosclerosis: The Role of Calcium. Med. Chem. 2016, 12, 103–113. [Google Scholar] [CrossRef]
- Shreya, D.; Zamora, D.I.; Patel, G.S.; Grossmann, I.; Rodriguez, K.; Soni, M.; Joshi, P.K.; Patel, S.C.; Sange, I. Coronary Artery Calcium Score—A Reliable Indicator of Coronary Artery Disease? Cureus 2021, 13, e20149. [Google Scholar] [CrossRef]
- Simes, D.C.; Viegas, C.S.B.; Araujo, N.; Marreiros, C. Vitamin K as a Powerful Micronutrient in Aging and Age-Related Diseases: Pros and Cons from Clinical Studies. Int. J. Mol. Sci. 2019, 20, 4150. [Google Scholar] [CrossRef]
- Booth, S.L. Vitamin K: Food composition and dietary intakes. Food Nutr. Res. 2012, 56. [Google Scholar] [CrossRef]
- Walther, B.; Karl, J.P.; Booth, S.L.; Boyaval, P. Menaquinones, bacteria, and the food supply: The relevance of dairy and fermented food products to vitamin K requirements. Adv. Nutr. 2013, 4, 463–473. [Google Scholar] [CrossRef]
- Bentley, R.; Meganathan, R. Biosynthesis of vitamin K (menaquinone) in bacteria. Microbiol. Rev. 1982, 46, 241–280. [Google Scholar] [CrossRef]
- Shearer, M.J.; Okano, T. Key Pathways and Regulators of Vitamin K Function and Intermediary Metabolism. Annu. Rev. Nutr. 2018, 38, 127–151. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Takada, T.; Kurauchi, R.; Tanaka, Y.; Komine, T.; Suzuki, H. Transporters for the Intestinal Absorption of Cholesterol, Vitamin, E, and Vitamin, K. J. Atheroscler. Thromb. 2017, 24, 347–359. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Vermeer, C. Differential lipoprotein transport pathways of K-vitamins in healthy subjects. Biochim. Biophys. Acta 2002, 1570, 27–32. [Google Scholar] [CrossRef]
- Ellis, J.L.; Fu, X.; Al Rajabi, A.; Grusak, M.A.; Shearer, M.J.; Naumova, E.N.; Saltzman, E.; Barger, K.; Booth, S.L. Plasma Response to Deuterium-Labeled Vitamin K Intake Varies by TG Response, but Not Age or Vitamin K Status, in Older and Younger Adults. J. Nutr. 2019, 149, 18–25. [Google Scholar] [CrossRef]
- Shea, M.K.; Berkner, K.L.; Ferland, G.; Fu, X.; Holden, R.M.; Booth, S.L. Perspective: Evidence before Enthusiasm—A Critical Review of the Potential Cardiovascular Benefits of Vitamin K. Adv. Nutr. 2021, 12, 632–646. [Google Scholar] [CrossRef]
- Ellis, J.L.; Fu, X.; Karl, J.P.; Hernandez, C.J.; Mason, J.B.; DeBose-Boyd, R.A.; Booth, S.L. Multiple Dietary Vitamin K Forms Are Converted to Tissue Menaquinone-4 in Mice. J. Nutr. 2022, 152, 981–993. [Google Scholar] [CrossRef]
- Thijssen, H.H.; Vervoort, L.M.; Schurgers, L.J.; Shearer, M.J. Menadione is a metabolite of oral vitamin K. Br. J. Nutr. 2006, 95, 260–266. [Google Scholar] [CrossRef]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (vitamin K3) is a catabolic product of oral phylloquinone (vitamin K1) in the intestine and a circulating precursor of tissue menaquinone-4 (vitamin K2) in rats. J. Biol. Chem. 2013, 288, 33071–33080. [Google Scholar] [CrossRef]
- Ellis, J.L.; Karl, J.P.; Oliverio, A.M.; Fu, X.; Soares, J.W.; Wolfe, B.E.; Hernandez, C.J.; Mason, J.B.; Booth, S.L. Dietary vitamin K is remodeled by gut microbiota and influences community composition. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Harshman, S.G.; Shea, M.K.; Fu, X.; Grusak, M.A.; Smith, D.; Lamon-Fava, S.; Kuliopulos, A.; Greenberg, A.; Booth, S.L. Atorvastatin Decreases Renal Menaquinone-4 Formation in C57BL/6 Male Mice. J. Nutr. 2019, 149, 416–421. [Google Scholar] [CrossRef]
- Park, J.; Pandya, V.R.; Ezekiel, S.J.; Berghuis, A.M. Phosphonate and Bisphosphonate Inhibitors of Farnesyl Pyrophosphate Synthases: A Structure-Guided Perspective. Front. Chem. 2020, 8, 612728. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, W.; Yang, P.; Zhu, S.; Luo, S.; Li, M. Menaquinone-4 prevents medication-related osteonecrosis of the jaw through the SIRT1 signaling-mediated inhibition of cellular metabolic stresses-induced osteoblast apoptosis. Free Radic. Biol. Med. 2023, 206, 33–49. [Google Scholar] [CrossRef]
- Ikeda, S.; Nomura, S.; Hanzawa, F.; Takahashi, S.; Oda, H.; Fujiwara, Y.; Uchida, T. Alpha-Tocopherol Intake Decreases Phylloquinone Concentration in Bone but Does Not Affect Bone Metabolism in Rats. J. Nutr. Sci. Vitaminol. 2018, 64, 243–250. [Google Scholar] [CrossRef]
- Sato, T.; Schurgers, L.J.; Uenishi, K. Comparison of menaquinone-4 and menaquinone-7 bioavailability in healthy women. Nutr. J. 2012, 11, 93. [Google Scholar] [CrossRef]
- Akbulut, A.C.; Pavlic, A.; Petsophonsakul, P.; Halder, M.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K2 Needs an RDI Separate from Vitamin K1. Nutrients 2020, 12, 1852. [Google Scholar] [CrossRef]
- Klapkova, E.; Cepova, J.; Dunovska, K.; Prusa, R. Determination of vitamins K(1), MK-4, and MK-7 in human serum of postmenopausal women by HPLC with fluorescence detection. J. Clin. Lab. Anal. 2018, 32, e22381. [Google Scholar] [CrossRef]
- Nie, S.; Yang, L.; Feng, J.; Lu, J.; Zhang, H.; Li, W.; Hu, Y.; Yang, X. Reference Range of Vitamin K Evaluating Indicators in Chinese Childbearing Women. Nutrients 2023, 15, 1977. [Google Scholar] [CrossRef]
- Vermeer, C.; Schurgers, L.J. A comprehensive review of vitamin K and vitamin K antagonists. Hematol. Oncol. Clin. N. Am. 2000, 14, 339–353. [Google Scholar] [CrossRef]
- Berkner, K.L. Vitamin K-dependent carboxylation. Vitam. Horm. 2008, 78, 131–156. [Google Scholar] [CrossRef]
- Tie, J.K.; Stafford, D.W. Structural and functional insights into enzymes of the vitamin K cycle. J. Thromb. Haemost. 2016, 14, 236–247. [Google Scholar] [CrossRef]
- Finnes, T.E.; Lofthus, C.M.; Meyer, H.E.; Sogaard, A.J.; Tell, G.S.; Apalset, E.M.; Gjesdal, C.; Grimnes, G.; Schei, B.; Blomhoff, R.; et al. A combination of low serum concentrations of vitamins K1 and D is associated with increased risk of hip fractures in elderly Norwegians: A NOREPOS study. Osteoporos. Int. 2016, 27, 1645–1652. [Google Scholar] [CrossRef]
- Aaseth, J.O.; Alexander, J. Postoperative Osteoporosis in Subjects with Morbid Obesity Undergoing Bariatric Surgery with Gastric Bypass or Sleeve Gastrectomy. Nutrients 2023, 15, 1302. [Google Scholar] [CrossRef]
- Schulman, S.; Wang, B.; Li, W.; Rapoport, T.A. Vitamin K epoxide reductase prefers ER membrane-anchored thioredoxin-like redox partners. Proc. Natl. Acad. Sci. USA 2010, 107, 15027–15032. [Google Scholar] [CrossRef]
- Rishavy, M.A.; Usubalieva, A.; Hallgren, K.W.; Berkner, K.L. Novel insight into the mechanism of the vitamin K oxidoreductase (VKOR): Electron relay through Cys43 and Cys51 reduces VKOR to allow vitamin K reduction and facilitation of vitamin K-dependent protein carboxylation. J. Biol. Chem. 2011, 286, 7267–7278. [Google Scholar] [CrossRef]
- Shearer, M.J. Vitamin K deficiency bleeding (VKDB) in early infancy. Blood Rev. 2009, 23, 49–59. [Google Scholar] [CrossRef]
- Tsugawa, N. Cardiovascular Diseases and Fat Soluble Vitamins: Vitamin D and Vitamin K. J. Nutr. Sci. Vitaminol. 2015, 61, S170–S172. [Google Scholar] [CrossRef]
- Danziger, J. Vitamin K-dependent proteins, warfarin, and vascular calcification. Clin. J. Am. Soc. Nephrol. 2008, 3, 1504–1510. [Google Scholar] [CrossRef]
- van Gorp, R.H.; Dijkgraaf, I.; Broker, V.; Bauwens, M.; Leenders, P.; Jennen, D.; Dweck, M.R.; Bucerius, J.; Briede, J.J.; van Ryn, J.; et al. Off-target effects of oral anticoagulants—vascular effects of vitamin K antagonist and non-vitamin K antagonist oral anticoagulant dabigatran etexilate. J. Thromb. Haemost. 2021, 19, 1348–1363. [Google Scholar] [CrossRef]
- Chen, Z.; Qureshi, A.R.; Parini, P.; Hurt-Camejo, E.; Ripsweden, J.; Brismar, T.B.; Barany, P.; Jaminon, A.M.; Schurgers, L.J.; Heimburger, O.; et al. Does statins promote vascular calcification in chronic kidney disease? Eur. J. Clin. Investig. 2017, 47, 137–148. [Google Scholar] [CrossRef]
- Xian, J.Z.; Lu, M.; Fong, F.; Qiao, R.; Patel, N.R.; Abeydeera, D.; Iriana, S.; Demer, L.L.; Tintut, Y. Statin Effects on Vascular Calcification: Microarchitectural Changes in Aortic Calcium Deposits in Aged Hyperlipidemic Mice. Arterioscler. Thromb. Vasc. Biol. 2021, 41, e185–e192. [Google Scholar] [CrossRef]
- Lee, S.E.; Chang, H.J.; Sung, J.M.; Park, H.B.; Heo, R.; Rizvi, A.; Lin, F.Y.; Kumar, A.; Hadamitzky, M.; Kim, Y.J.; et al. Effects of Statins on Coronary Atherosclerotic Plaques: The PARADIGM Study. JACC Cardiovasc. Imaging 2018, 11, 1475–1484. [Google Scholar] [CrossRef]
- Doran, A.C.; Terry, J.G.; Carr, J.J.; Linton, M.F. Statins and Atherosclerotic Lesion Microcalcification: A New Mechanism for Plaque Stability? Arterioscler. Thromb. Vasc. Biol. 2021, 41, 1306–1308. [Google Scholar] [CrossRef]
- Hirota, Y.; Suhara, Y. New Aspects of Vitamin K Research with Synthetic Ligands: Transcriptional Activity via SXR and Neural Differentiation Activity. Int. J. Mol. Sci. 2019, 20, 3006. [Google Scholar] [CrossRef]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grun, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 regulation of bone homeostasis is mediated by the steroid and xenobiotic receptor SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef]
- Azuma, K.; Casey, S.C.; Ito, M.; Urano, T.; Horie, K.; Ouchi, Y.; Kirchner, S.; Blumberg, B.; Inoue, S. Pregnane X receptor knockout mice display osteopenia with reduced bone formation and enhanced bone resorption. J. Endocrinol. 2010, 207, 257–263. [Google Scholar] [CrossRef]
- Yang, W.; Yu, Z.; Chiyoya, M.; Liu, X.; Daitoku, K.; Motomura, S.; Imaizumi, T.; Fukuda, I.; Furukawa, K.I.; Tsuji, M.; et al. Menaquinone-4 Accelerates Calcification of Human Aortic Valve Interstitial Cells in High-Phosphate Medium through PXR. J. Pharmacol. Exp. Ther. 2020, 372, 277–284. [Google Scholar] [CrossRef]
- Yu, Z.; Seya, K.; Chiyoya, M.; Daitoku, K.; Motomura, S.; Imaizumi, T.; Fukuda, I.; Furukawa, K.I. Warfarin calcifies human aortic valve interstitial cells at high-phosphate conditions via pregnane X receptor. J. Bone Miner. Metab. 2019, 37, 944–956. [Google Scholar] [CrossRef]
- Shioi, A.; Morioka, T.; Shoji, T.; Emoto, M. The Inhibitory Roles of Vitamin K in Progression of Vascular Calcification. Nutrients 2020, 12, 583. [Google Scholar] [CrossRef]
- Aaseth, J.; Skaug, M.A.; Cao, Y.; Andersen, O. Chelation in metal intoxication–Principles and paradigms. J. Trace Elem. Med. Biol. 2015, 31, 260–266. [Google Scholar] [CrossRef]
- Chen, H.G.; Sheng, L.T.; Zhang, Y.B.; Cao, A.L.; Lai, Y.W.; Kunutsor, S.K.; Jiang, L.; Pan, A. Association of vitamin K with cardiovascular events and all-cause mortality: A systematic review and meta-analysis. Eur. J. Nutr. 2019, 58, 2191–2205. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Dounousi, E.; Eleftheriadis, T.; Liakopoulos, V. Association of the Inactive Circulating Matrix Gla Protein with Vitamin K Intake, Calcification, Mortality, and Cardiovascular Disease: A Review. Int. J. Mol. Sci. 2019, 20, 628. [Google Scholar] [CrossRef]
- Wallin, R.; Cain, D.; Hutson, S.M.; Sane, D.C.; Loeser, R. Modulation of the binding of matrix Gla protein (MGP) to bone morphogenetic protein-2 (BMP-2). Thromb. Haemost. 2000, 84, 1039–1044. [Google Scholar]
- Zebboudj, A.F.; Shin, V.; Bostrom, K. Matrix GLA protein and BMP-2 regulate osteoinduction in calcifying vascular cells. J. Cell. Biochem. 2003, 90, 756–765. [Google Scholar] [CrossRef]
- Shea, M.K.; Barger, K.; Booth, S.L.; Matuszek, G.; Cushman, M.; Benjamin, E.J.; Kritchevsky, S.B.; Weiner, D.E. Vitamin K status, cardiovascular disease, and all-cause mortality: A participant-level meta-analysis of 3 US cohorts. Am. J. Clin. Nutr. 2020, 111, 1170–1177. [Google Scholar] [CrossRef]
- Belknap, E.L. EDTA in the treatment of lead poisoning. Ind. Med. Surg. 1952, 21, 305–306. [Google Scholar]
- Clarke, N.E.; Clarke, C.N.; Mosher, R.E. The in vivo dissolution of metastatic calcium; an approach to atherosclerosis. Am. J. Med. Sci. 1955, 229, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Lamas, G.A.; Goertz, C.; Boineau, R.; Mark, D.B.; Rozema, T.; Nahin, R.L.; Lindblad, L.; Lewis, E.F.; Drisko, J.; Lee, K.L.; et al. Effect of disodium EDTA chelation regimen on cardiovascular events in patients with previous myocardial infarction: The TACT randomized trial. JAMA 2013, 309, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Lamas, G.A.; Anstrom, K.J.; Navas-Acien, A.; Boineau, R.; Kim, H.; Rosenberg, Y.; Stylianou, M.; Jones, T.L.Z.; Joubert, B.R.; Santella, R.M.; et al. The trial to assess chelation therapy 2 (TACT2): Rationale and design. Am. Heart J. 2022, 252, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Waters, R.S.; Bryden, N.A.; Patterson, K.Y.; Veillon, C.; Anderson, R.A. EDTA chelation effects on urinary losses of cadmium, calcium, chromium, cobalt, copper, lead, magnesium, and zinc. Biol. Trace Elem. Res. 2001, 83, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Villarruz-Sulit, M.V.; Forster, R.; Dans, A.L.; Tan, F.N.; Sulit, D.V. Chelation therapy for atherosclerotic cardiovascular disease. Cochrane Database Syst. Rev. 2020, 5, CD002785. [Google Scholar] [CrossRef] [PubMed]
- Olszewer, E.; Carter, J.P. EDTA chelation therapy in chronic degenerative disease. Med. Hypotheses 1988, 27, 41–49. [Google Scholar] [CrossRef]
- Guldager, B.; Jelnes, R.; Jorgensen, S.J.; Nielsen, J.S.; Klaerke, A.; Mogensen, K.; Larsen, K.E.; Reimer, E.; Holm, J.; Ottesen, S. EDTA treatment of intermittent claudication—A double-blind, placebo-controlled study. J. Intern. Med. 1992, 231, 261–267. [Google Scholar] [CrossRef]
- van Rij, A.M.; Solomon, C.; Packer, S.G.; Hopkins, W.G. Chelation therapy for intermittent claudication. A double-blind, randomized, controlled trial. Circulation 1994, 90, 1194–1199. [Google Scholar] [CrossRef]
- Knudtson, M.L.; Wyse, D.G.; Galbraith, P.D.; Brant, R.; Hildebrand, K.; Paterson, D.; Richardson, D.; Burkart, C.; Burgess, E. Program to Assess Alternative Treatment Strategies to Achieve Cardiac Health, I. Chelation therapy for ischemic heart disease: A randomized controlled trial. JAMA 2002, 287, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Ujueta, F.; Arenas, I.A.; Escolar, E.; Diaz, D.; Boineau, R.; Mark, D.B.; Golden, P.; Lindblad, L.; Kim, H.; Lee, K.L.; et al. The effect of EDTA-based chelation on patients with diabetes and peripheral artery disease in the Trial to Assess Chelation Therapy (TACT). J. Diabetes Complicat. 2019, 33, 490–494. [Google Scholar] [CrossRef]
- Ujueta, F.; Navas-Acien, A.; Mann, K.K.; Prashad, R.; Lamas, G.A. Low-Level Metal Contamination and Chelation in Cardiovascular Disease—A Ripe Area for Toxicology Research. Toxicol. Sci. 2021, 181, 135–147. [Google Scholar] [CrossRef]
- Chappell, L.T.; Drisko, J.A. Protocol controversies for treating cardiovascular disease with EDTA chelation therapy. Townsend Lett. 2014, 181, 38–45. [Google Scholar]
- Nissen, S.E. Concerns about reliability in the Trial to Assess Chelation Therapy (TACT). JAMA 2013, 309, 1293–1294. [Google Scholar] [CrossRef]
- Pereira, L.C.; Pereira, F.A.; Sa, M.F.; Foss, M.C.; de Paula, F.J. Parathyroid hormone secretion in women in late menopause submitted to EDTA-induced hypocalcemia. Maturitas 2008, 59, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Mosekilde, L. Primary hyperparathyroidism and the skeleton. Clin. Endocrinol. 2008, 69, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Negru, A.G.; Pastorcici, A.; Crisan, S.; Cismaru, G.; Popescu, F.G.; Luca, C.T. The Role of Hypomagnesemia in Cardiac Arrhythmias: A Clinical Perspective. Biomedicines 2022, 10, 2356. [Google Scholar] [CrossRef] [PubMed]
- Willems, B.A.; Vermeer, C.; Reutelingsperger, C.P.; Schurgers, L.J. The realm of vitamin K dependent proteins: Shifting from coagulation toward calcification. Mol. Nutr. Food Res. 2014, 58, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- Donato, M.; Faggin, E.; Cinetto, F.; Felice, C.; Lupo, M.G.; Ferri, N.; Rattazzi, M. The Emerging Role of Nutraceuticals in Cardiovascular Calcification: Evidence from Preclinical and Clinical Studies. Nutrients 2021, 13, 2603. [Google Scholar] [CrossRef] [PubMed]
- Theuwissen, E.; Smit, E.; Vermeer, C. The role of vitamin K in soft-tissue calcification. Adv. Nutr. 2012, 3, 166–173. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Henning, A.L.; Venable, A.S. Oral Consumption of Vitamin K2 for 8 Weeks Associated with Increased Maximal Cardiac Output during Exercise. Altern. Ther. Health Med. 2017, 23, 26–32. [Google Scholar]
- Gast, G.C.; de Roos, N.M.; Sluijs, I.; Bots, M.L.; Beulens, J.W.; Geleijnse, J.M.; Witteman, J.C.; Grobbee, D.E.; Peeters, P.H.; van der Schouw, Y.T. A high menaquinone intake reduces the incidence of coronary heart disease. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 504–510. [Google Scholar] [CrossRef]
- Vissers, L.E.T.; Dalmeijer, G.W.; Boer, J.M.A.; Verschuren, W.M.M.; van der Schouw, Y.T.; Beulens, J.W.J. The relationship between vitamin K and peripheral arterial disease. Atherosclerosis 2016, 252, 15–20. [Google Scholar] [CrossRef]
- Ruiz-Leon, A.M.; Lapuente, M.; Estruch, R.; Casas, R. Clinical Advances in Immunonutrition and Atherosclerosis: A Review. Front. Immunol. 2019, 10, 837. [Google Scholar] [CrossRef]
- Shea, M.K.; Booth, S.L.; Miller, M.E.; Burke, G.L.; Chen, H.; Cushman, M.; Tracy, R.P.; Kritchevsky, S.B. Association between circulating vitamin K1 and coronary calcium progression in community-dwelling adults: The Multi-Ethnic Study of Atherosclerosis. Am. J. Clin. Nutr. 2013, 98, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Haugsgjerd, T.R.; Egeland, G.M.; Nygard, O.K.; Vinknes, K.J.; Sulo, G.; Lysne, V.; Igland, J.; Tell, G.S. Association of dietary vitamin K and risk of coronary heart disease in middle-age adults: The Hordaland Health Study Cohort. BMJ Open 2020, 10, e035953. [Google Scholar] [CrossRef] [PubMed]
- Bellinge, J.W.; Dalgaard, F.; Murray, K.; Connolly, E.; Blekkenhorst, L.C.; Bondonno, C.P.; Lewis, J.R.; Sim, M.; Croft, K.D.; Gislason, G.; et al. Vitamin K Intake and Atherosclerotic Cardiovascular Disease in the Danish Diet Cancer and Health Study. J. Am. Heart Assoc. 2021, 10, e020551. [Google Scholar] [CrossRef] [PubMed]
- Braam, L.A.; Hoeks, A.P.; Brouns, F.; Hamulyak, K.; Gerichhausen, M.J.; Vermeer, C. Beneficial effects of vitamins D and K on the elastic properties of the vessel wall in postmenopausal women: A follow-up study. Thromb. Haemost. 2004, 91, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Knapen, M.H.; Braam, L.A.; Drummen, N.E.; Bekers, O.; Hoeks, A.P.; Vermeer, C. Menaquinone-7 supplementation improves arterial stiffness in healthy postmenopausal women. A double-blind randomised clinical trial. Thromb. Haemost. 2015, 113, 1135–1144. [Google Scholar] [CrossRef]
- Lees, J.S.; Chapman, F.A.; Witham, M.D.; Jardine, A.G.; Mark, P.B. Vitamin K status, supplementation and vascular disease: A systematic review and meta-analysis. Heart 2019, 105, 938–945. [Google Scholar] [CrossRef]
- Bellinge, J.W.; Francis, R.J.; Lee, S.C.; Vickery, A.; Macdonald, W.; Gan, S.K.; Chew, G.T.; Phillips, M.; Lewis, J.R.; Watts, G.F.; et al. The effect of Vitamin-K1 and Colchicine on Vascular Calcification Activity in subjects with Diabetes Mellitus (ViKCoVaC): A double-blind 2 × 2 factorial randomized controlled trial. J. Nucl. Cardiol. 2022, 29, 1855–1866. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aaseth, J.O.; Alehagen, U.; Opstad, T.B.; Alexander, J. Vitamin K and Calcium Chelation in Vascular Health. Biomedicines 2023, 11, 3154. https://doi.org/10.3390/biomedicines11123154
Aaseth JO, Alehagen U, Opstad TB, Alexander J. Vitamin K and Calcium Chelation in Vascular Health. Biomedicines. 2023; 11(12):3154. https://doi.org/10.3390/biomedicines11123154
Chicago/Turabian StyleAaseth, Jan O., Urban Alehagen, Trine Baur Opstad, and Jan Alexander. 2023. "Vitamin K and Calcium Chelation in Vascular Health" Biomedicines 11, no. 12: 3154. https://doi.org/10.3390/biomedicines11123154
APA StyleAaseth, J. O., Alehagen, U., Opstad, T. B., & Alexander, J. (2023). Vitamin K and Calcium Chelation in Vascular Health. Biomedicines, 11(12), 3154. https://doi.org/10.3390/biomedicines11123154