
Modeling Central Nervous System Injury In Vitro: Current Status and Promising Future Strategies

Abstract
1. Introduction
2. In Vitro Models of Trauma
3. In Vitro Models of Cerebral Hypoxia/Ischemia
4. Cell Culture Platforms Used in Traumatic Brain Injury and Brain Ischemia In Vitro Models
4.1. Primary Cell Lines
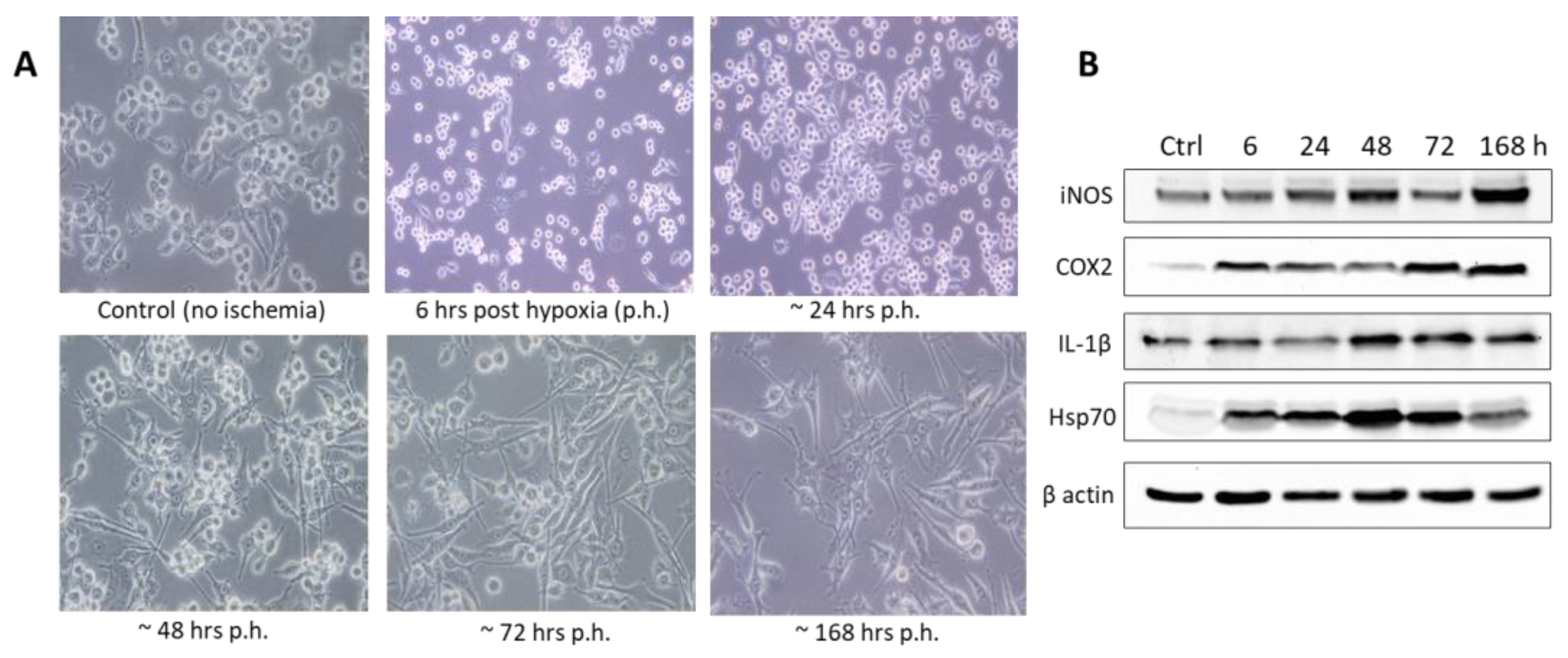
4.2. Immortalized Cell Lines
4.3. Co-Cultures, 3D Culture Models, and Brain Organoids
4.4. Organotypic Slice Cultures
4.5. Human Induced Pluripotent Stem Cells
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Capizzi, A.; Woo, J.; Verduzco-Gutierrez, M. Traumatic Brain Injury: An Overview of Epidemiology, Pathophysiology, and Medical Management. Med. Clin. N. Am. 2020, 104, 213–238. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-H.; Rosset, S.; Lee, T.-R.; Dragunow, M.; Park, T.; Shim, V. In Vitro Models of Traumatic Brain Injury: A Systematic Review. J. Neurotrauma 2021, 38, 2336–2372. [Google Scholar] [CrossRef] [PubMed]
- Kumaria, A. In Vitro Models as a Platform to Investigate Traumatic Brain Injury. Altern. Lab. Anim. 2017, 45, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, A.; Singh, N.K.; Firestein, B.L. Current Advances in in Vitro Models of Central Nervous System Trauma. Curr. Opin. Biomed. Eng. 2020, 14, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.J. Ischemic Stroke: Experimental Models and Reality. Acta Neuropathol. 2017, 133, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Ezzelarab, N.M.; Saleh, N.; Khalil, E.A.; Abdellatif, A. Experimental Models and Plant-based Therapy for Experimental Cerebral Ischemia (Review). Int. J. Funct. Nutr. 2020, 1, 5. [Google Scholar] [CrossRef]
- Holloway, P.M.; Gavins, F.N.E. Modeling Ischemic Stroke In Vitro: Status Quo and Future Perspectives. Stroke 2016, 47, 561–569. [Google Scholar] [CrossRef]
- Shaughness, M.; Byrnes, K. Assessment of the Effects of Stretch-Injury on Primary Rat Microglia. Mol. Neurobiol. 2021, 58, 3545–3560. [Google Scholar] [CrossRef]
- Hemphill, M.A.; Dauth, S.; Yu, C.J.; Dabiri, B.E.; Parker, K.K. Traumatic Brain Injury and the Neuronal Microenvironment: A Potential Role for Neuropathological Mechanotransduction. Neuron 2015, 85, 1177–1192. [Google Scholar] [CrossRef]
- Jowers, C.T.; Taberner, A.J.; Dragunow, M.; Anderson, I.A. The Cell Injury Device: A High-Throughput Platform for Traumatic Brain Injury Research. J. Neurosci. Methods 2013, 218, 1–8. [Google Scholar] [CrossRef]
- Cao, Y.; Risling, M.; Malm, E.; Sondén, A.; Bolling, M.F.; Sköld, M.K. Cellular High-Energy Cavitation Trauma—Description of a Novel In Vitro Trauma Model in Three Different Cell Types. Front. Neurol. 2016, 7, 10. [Google Scholar] [CrossRef]
- Nogueira, G.O.; Garcez, P.P.; Bardy, C.; Cunningham, M.O.; Sebollela, A. Modeling the Human Brain With Ex Vivo Slices and in Vitro Organoids for Translational Neuroscience. Front. Neurosci. 2022, 16, 838594. [Google Scholar] [CrossRef]
- Mršić-Pelčić, J.; Pilipović, K.; Pelčić, G.; Vitezić, D.; Župan, G. Decrease in Oxidative Stress Parameters after Post-Ischaemic Recombinant Human Erythropoietin Administration in the Hippocampus of Rats Exposed to Focal Cerebral Ischaemia. Basic Clin. Pharm. Toxicol. 2017, 121, 453–464. [Google Scholar] [CrossRef]
- Trotman-Lucas, M.; Gibson, C.L. A Review of Experimental Models of Focal Cerebral Ischemia Focusing on the Middle Cerebral Artery Occlusion Model. F1000Res 2021, 10, 242. [Google Scholar] [CrossRef]
- van der Kooij, M.A.; Groenendaal, F.; Kavelaars, A.; Heijnen, C.J.; van Bel, F. Neuroprotective Properties and Mechanisms of Erythropoietin in in Vitro and in Vivo Experimental Models for Hypoxia/Ischemia. Brain Res. Rev. 2008, 59, 22–33. [Google Scholar] [CrossRef]
- Porterfield, V. Neural Progenitor Cell Derivation Methodologies for Drug Discovery Applications. Assay Drug Dev. Technol. 2020, 18, 89–95. [Google Scholar] [CrossRef]
- Richard, M.J.P.; Saleh, T.M.; El Bahh, B.; Zidichouski, J.A. A Novel Method for Inducing Focal Ischemia in Vitro. J. Neurosci. Methods 2010, 190, 20–27. [Google Scholar] [CrossRef]
- Kurian, G.A.; Pemaih, B. Standardization of in Vitro Cell-Based Model for Renal Ischemia and Reperfusion Injury. Indian J. Pharm. Sci. 2014, 76, 348–353. [Google Scholar]
- Xie, M.; Wang, W.; Kimelberg, H.K.; Zhou, M. Oxygen and Glucose Deprivation-Induced Changes in Astrocyte Membrane Potential and Their Underlying Mechanisms in Acute Rat Hippocampal Slices. J. Cereb. Blood Flow Metab. 2008, 28, 456–467. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Turovskaya, M.V.; Gaidin, S.G.; Zinchenko, V.P. Cytokine IL-10, Activators of PI3-Kinase, Agonists of α-2 Adrenoreceptor and Antioxidants Prevent Ischemia-Induced Cell Death in Rat Hippocampal Cultures. Arch. Biochem. Biophys. 2017, 615, 35–43. [Google Scholar] [CrossRef]
- Turovskaya, M.V.; Turovsky, E.A.; Zinchenko, V.P.; Levin, S.G.; Shamsutdinova, A.A.; Godukhin, O.V. Repeated Brief Episodes of Hypoxia Modulate the Calcium Responses of Ionotropic Glutamate Receptors in Hippocampal Neurons. Neurosci. Lett. 2011, 496, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.J.; Wang, L.; Xu, Q.; McTiernan, C.F.; Shiva, S.; Tejero, J.; Gladwin, M.T. Carbon Monoxide Poisoning: Pathogenesis, Management, and Future Directions of Therapy. Am. J. Respir. Crit. Care Med. 2017, 195, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Blasi, E.; Barluzzi, R.; Bocchini, V.; Mazzolla, R.; Bistoni, F. Immortalization of Murine Microglial Cells by a V-Raf/v-Myc Carrying Retrovirus. J. Neuroimmunol. 1990, 27, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, G.; Brafman, D.A. The Emergence of Model Systems to Investigate the Link Between Traumatic Brain Injury and Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 813544. [Google Scholar] [CrossRef]
- Hamilton, K.A.; Santhakumar, V. Current Ex Vivo and in Vitro Approaches to Uncovering Mechanisms of Neurological Dysfunction after Traumatic Brain Injury. Curr. Opin. Biomed. Eng. 2020, 14, 18–24. [Google Scholar] [CrossRef]
- Shi, Y.; Inoue, H.; Wu, J.C.; Yamanaka, S. Induced Pluripotent Stem Cell Technology: A Decade of Progress. Nat. Rev. Drug Discov. 2017, 16, 115–130. [Google Scholar] [CrossRef]
- Juntunen, M.; Hagman, S.; Moisan, A.; Narkilahti, S.; Miettinen, S. In Vitro Oxygen-Glucose Deprivation-Induced Stroke Models with Human Neuroblastoma Cell- and Induced Pluripotent Stem Cell-Derived Neurons. Stem Cells Int. 2020, 2020, e8841026. [Google Scholar] [CrossRef]
- Liu, Q.; Jin, Z.; Xu, Z.; Yang, H.; Li, L.; Li, G.; Li, F.; Gu, S.; Zong, S.; Zhou, J.; et al. Antioxidant Effects of Ginkgolides and Bilobalide against Cerebral Ischemia Injury by Activating the Akt/Nrf2 Pathway in Vitro and in Vivo. Cell Stress Chaperones 2019, 24, 441–452. [Google Scholar] [CrossRef]
- Lockhart, E.M.; Warner, D.S.; Pearlstein, R.D.; Penning, D.H.; Mehrabani, S.; Boustany, R.M. Allopregnanolone Attenuates N-Methyl-D-Aspartate-Induced Excitotoxicity and Apoptosis in the Human NT2 Cell Line in Culture. Neurosci. Lett. 2002, 328, 33–36. [Google Scholar] [CrossRef]
- Roloff, F.; Scheiblich, H.; Dewitz, C.; Dempewolf, S.; Stern, M.; Bicker, G. Enhanced Neurite Outgrowth of Human Model (NT2) Neurons by Small-Molecule Inhibitors of Rho/ROCK Signaling. PLoS ONE 2015, 10, e0118536. [Google Scholar] [CrossRef]
- Wang, L.; Yin, Z.; Wang, F.; Han, Z.; Wang, Y.; Huang, S.; Hu, T.; Guo, M.; Lei, P. Hydrogen Exerts Neuroprotection by Activation of the MiR-21/PI3K/AKT/GSK-3 Beta Pathway in an in Vitro Model of Traumatic Brain Injury. J. Cell. Mol. Med. 2020, 24, 4061–4071. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Zhou, Y.; Liang, D.; He, H.; Liu, X.; Zhu, R.; Zhang, M.; Luo, X.; Wang, Y.; Huang, G. Exosomes Secreted From Bone Marrow Mesenchymal Stem Cells Attenuate Oxygen-Glucose Deprivation/Reoxygenation-Induced Pyroptosis in PC12 Cells by Promoting AMPK-Dependent Autophagic Flux. Front. Cell. Neurosci. 2020, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhang, Y.; Yuan, X.; Pan, Y.; Yang, L.; Zhao, Y.; Zhuo, R.; Chen, C.; Peng, L.; Li, W.; et al. Oleoylethanolamide Inhibits Glial Activation via Moudulating PPAR Alpha and Promotes Motor Function Recovery after Brain Ischemia. Pharmacol. Res. 2019, 141, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Sima, G.; Wu, C.; Fan, Y.; Tan, Y.; Wang, Z.; Cheng, G.; Li, J. Decreased MiR-17 in Glioma Cells Increased Cell Viability and Migration by Increasing the Expression of Cyclin D1, p-Akt and Akt. PLoS ONE 2018, 13, e0190515. [Google Scholar] [CrossRef] [PubMed]
- Barialai, L.; Strecker, M.I.; Luger, A.-L.; Jaeger, M.; Bruns, I.; Sittig, A.C.M.; Mildenberger, I.C.; Heller, S.M.; Delaidelli, A.; Lorenz, N.I.; et al. AMPK Activation Protects Astrocytes from Hypoxia-Induced Cell Death. Int. J. Mol. Med. 2020, 45, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, Z.; Zhu, X.; Lin, R.; Chen, L. Salidroside Reduces Cell Mobility via NF- κ B and MAPK Signaling in LPS-Induced BV2 Microglial Cells. Evid. Based Complement. Altern. Med 2014, 2014, 383821. [Google Scholar] [CrossRef]
- Smith, G.S.T.; Voyer-Grant, J.A.M.; Harauz, G. Monitoring Cleaved Caspase-3 Activity and Apoptosis of Immortalized Oligodendroglial Cells Using Live-Cell Imaging and Cleaveable Fluorogenic-Dye Substrates Following Potassium-Induced Membrane Depolarization. J. Vis. Exp. 2012, 59, e3422. [Google Scholar] [CrossRef]
- Fennema, E.; Rivron, N.; Rouwkema, J.; van Blitterswijk, C.; de Boer, J. Spheroid Culture as a Tool for Creating 3D Complex Tissues. Trends Biotechnol. 2013, 31, 108–115. [Google Scholar] [CrossRef]
- Zhuang, P.; Sun, A.X.; An, J.; Chua, C.K.; Chew, S.Y. 3D Neural Tissue Models: From Spheroids to Bioprinting. Biomaterials 2018, 154, 113–133. [Google Scholar] [CrossRef]
- Jorfi, M.; D’Avanzo, C.; Tanzi, R.E.; Kim, D.Y.; Irimia, D. Human Neurospheroid Arrays for In Vitro Studies of Alzheimer’s Disease. Sci. Rep. 2018, 8, 2450. [Google Scholar] [CrossRef]
- Eiraku, M.; Watanabe, K.; Matsuo-Takasaki, M.; Kawada, M.; Yonemura, S.; Matsumura, M.; Wataya, T.; Nishiyama, A.; Muguruma, K.; Sasai, Y. Self-Organized Formation of Polarized Cortical Tissues from ESCs and Its Active Manipulation by Extrinsic Signals. Cell Stem Cell 2008, 3, 519–532. [Google Scholar] [CrossRef]
- Wang, S.-N.; Wang, Z.; Wang, X.-Y.; Zhang, X.-P.; Xu, T.-Y.; Miao, C.-Y. Humanized Cerebral Organoids-Based Ischemic Stroke Model for Discovering of Potential Anti-Stroke Agents. Acta Pharmacol. Sin. 2022. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, L.; Kuang, Y.; Venkataramani, V.; Jin, F.; Hein, K.; Zafeiriou, M.P.; Lenz, C.; Moebius, W.; Kilic, E.; et al. Extracellular Vesicles Derived from Neural Progenitor Cells—A Preclinical Evaluation for Stroke Treatment in Mice. Transl. Stroke Res. 2021, 12, 185–203. [Google Scholar] [CrossRef]
- Ramirez, S.; Mukherjee, A.; Sepulveda, S.; Becerra-Calixto, A.; Bravo-Vasquez, N.; Gherardelli, C.; Chavez, M.; Soto, C. Modeling Traumatic Brain Injury in Human Cerebral Organoids. Cells 2021, 10, 2683. [Google Scholar] [CrossRef]
- El-Ali, J.; Sorger, P.K.; Jensen, K.F. Cells on Chips. Nature 2006, 442, 403–411. [Google Scholar] [CrossRef]
- Esch, E.W.; Bahinski, A.; Huh, D. Organs-on-Chips at the Frontiers of Drug Discovery. Nat. Rev. Drug Discov. 2015, 14, 248–260. [Google Scholar] [CrossRef]
- Kilic, O.; Pamies, D.; Lavell, E.; Schiapparelli, P.; Feng, Y.; Hartung, T.; Bal-Price, A.; Hogberg, H.T.; Quinones-Hinojosa, A.; Guerrero-Cazares, H.; et al. Brain-on-a-Chip Model Enables Analysis of Human Neuronal Differentiation and Chemotaxis. Lab. Chip. 2016, 16, 4152–4162. [Google Scholar] [CrossRef]
- Zheng, F.; Fu, F.; Cheng, Y.; Wang, C.; Zhao, Y.; Gu, Z. Organ-on-a-Chip Systems: Microengineering to Biomimic Living Systems. Small 2016, 12, 2253–2282. [Google Scholar] [CrossRef]
- Li, Q.; Han, X.; Wang, J. Organotypic Hippocampal Slices as Models for Stroke and Traumatic Brain Injury. Mol. Neurobiol. 2016, 53, 4226–4237. [Google Scholar] [CrossRef]
- Cimarosti, H.; Henley, J.M. Investigating the Mechanisms Underlying Neuronal Death in Ischemia Using in Vitro Oxygen-Glucose Deprivation: Potential Involvement of Protein SUMOylation. Neuroscientist 2008, 14, 626–636. [Google Scholar] [CrossRef]
- Noraberg, J.; Poulsen, F.R.; Blaabjerg, M.; Kristensen, B.W.; Bonde, C.; Montero, M.; Meyer, M.; Gramsbergen, J.B.; Zimmer, J. Organotypic Hippocampal Slice Cultures for Studies of Brain Damage, Neuroprotection and Neurorepair. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Lana, D.; Gerace, E.; Magni, G.; Cialdai, F.; Monici, M.; Mannaioni, G.; Giovannini, M.G. Hypoxia/Ischemia-Induced Rod Microglia Phenotype in CA1 Hippocampal Slices. Int. J. Mol. Sci. 2022, 23, 1422. [Google Scholar] [CrossRef] [PubMed]
- Ziemka-Nałęcz, M.; Stanaszek, L.; Zalewska, T. Oxygen-Glucose Deprivation Promotes Gliogenesis and Microglia Activation in Organotypic Hippocampal Slice Culture: Involvement of Metalloproteinases. Acta Neurobiol. Exp. (Wars) 2013, 73, 130–142. [Google Scholar] [PubMed]
- Landucci, E.; Pellegrini-Giampietro, D.E.; Facchinetti, F. Experimental Models for Testing the Efficacy of Pharmacological Treatments for Neonatal Hypoxic-Ischemic Encephalopathy. Biomedicines 2022, 10, 937. [Google Scholar] [CrossRef] [PubMed]
- Daviaud, N.; Garbayo, E.; Schiller, P.C.; Perez-Pinzon, M.; Montero-Menei, C.N. Organotypic Cultures as Tools for Optimizing Central Nervous System Cell Therapies. Exp. Neurol. 2013, 248, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Dzhala, V.; Staley, K.J. Acute and Chronic Efficacy of Bumetanide in an in Vitro Model of Posttraumatic Epileptogenesis. CNS Neurosci. 2015, 21, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Sherman, S.A.; Phillips, J.K.; Costa, J.T.; Cho, F.S.; Oungoulian, S.R.; Finan, J.D. Stretch Injury of Human Induced Pluripotent Stem Cell Derived Neurons in a 96 Well Format. Sci. Rep. 2016, 6, 34097. [Google Scholar] [CrossRef]
- Phillips, J.K.; Sherman, S.A.; Oungoulian, S.R.; Finan, J.D. Method for High Speed Stretch Injury of Human Induced Pluripotent Stem Cell-Derived Neurons in a 96-Well Format. J. Vis. Exp. 2018, 134, 57305. [Google Scholar] [CrossRef]
- Zander, N.E.; Piehler, T.; Hogberg, H.; Pamies, D. Explosive Blast Loading on Human 3D Aggregate Minibrains. Cell. Mol. Neurobiol. 2017, 37, 1331–1334. [Google Scholar] [CrossRef]
- Shoemaker, A.R.; Jones, I.E.; Jeffris, K.D.; Gabrielli, G.; Togliatti, A.G.; Pichika, R.; Martin, E.; Kiskinis, E.; Franz, C.K.; Finan, J.D. Biofidelic Dynamic Compression of Human Cortical Spheroids Reproduces Neurotrauma Phenotypes. Dis. Model. Mech. 2021, 14, dmm048916. [Google Scholar] [CrossRef]
- Chaves, R.S.; Tran, M.; Holder, A.R.; Balcer, A.M.; Dickey, A.M.; Roberts, E.A.; Bober, B.G.; Gutierrez, E.; Head, B.P.; Groisman, A.; et al. Amyloidogenic Processing of Amyloid Precursor Protein Drives Stretch-Induced Disruption of Axonal Transport in HiPSC-Derived Neurons. J. Neurosci. 2021, 41, 10034–10053. [Google Scholar] [CrossRef]
- Ramirez, S.; Mukherjee, A.; Sepulveda, S.E.; Gherardelli, C.; Becerra-Calixto, A.; Bravo-Vasquez, N.; Soto, C. Protocol for Controlled Cortical Impact in Human Cerebral Organoids to Model Traumatic Brain Injury. STAR Protoc. 2021, 2, 100987. [Google Scholar] [CrossRef]
- Shi, W.; Dong, P.; Kuss, M.A.; Gu, L.; Kievit, F.; Kim, H.J.; Duan, B. Design and Evaluation of an In Vitro Mild Traumatic Brain Injury Modeling System Using 3D Printed Mini Impact Device on the 3D Cultured Human IPSC Derived Neural Progenitor Cells. Adv. Heal. Mater. 2021, 10, e2100180. [Google Scholar] [CrossRef]
- Phillips, J.K.; Sherman, S.A.; Cotton, K.Y.; Heddleston, J.M.; Taylor, A.B.; Finan, J.D. Characterization of Neurite Dystrophy after Trauma by High Speed Structured Illumination Microscopy and Lattice Light Sheet Microscopy. J. Neurosci. Methods 2019, 312, 154–161. [Google Scholar] [CrossRef]
- Kokubu, Y.; Yamaguchi, T.; Kawabata, K. In Vitro Model of Cerebral Ischemia by Using Brain Microvascular Endothelial Cells Derived from Human Induced Pluripotent Stem Cells. Biochem. Biophys. Res. Commun. 2017, 486, 577–583. [Google Scholar] [CrossRef]
- Monteiro, S.P.; Voogd, E.; Muzzi, L.; Vecchis, G.D.; Mossink, B.; Levers, M.; Hassink, G.; Putten, M.V.; Feber, J.L.; Hofmeijer, J.; et al. Neuroprotective Effect of Hypoxic Preconditioning and Neuronal Activation in a in Vitro Human Model of the Ischemic Penumbra. J. Neural Eng. 2021, 18, 036016. [Google Scholar] [CrossRef]
- Räsänen, N.; Harju, V.; Joki, T.; Narkilahti, S. Practical Guide for Preparation, Computational Reconstruction and Analysis of 3D Human Neuronal Networks in Control and Ischaemic Conditions. Development 2022, 149, dev200012. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cell Line | Origin and Source | Selected References |
|---|---|---|
| SH-SY5Y | Human, neuroblastoma cells | [27,28] |
| NTera (NT2) | Human, neuronally committed teratocarcinoma cell line | [29,30] |
| PC12 | Rat, derived from a pheochromocytoma of the adrenal medulla | [31,32] |
| C6 | Rat, glioma cell line | [33,34] |
| SVG | Human, immortalized astrocytes | [35] |
| BV-2 | Mouse, murine microglial cell line | [23,36] |
| N19 | Mouse, immortalized oligodendrocytes | [37] |
| Model | Origin and Cell Type | References |
|---|---|---|
| TBI/stretch injury | Human iPSC-derived neurons | [58] |
| TBI/stretch injury | Human iPSC-derived neurons | [59] |
| TBI/blast injury | 3D aggregates of human iPSCs (minibrains) | [60] |
| TBI/compressive injury | Cortical spheroids derived from human iPSCs | [61] |
| TBI/stretch injury | Human iPSC-derived neurons | [62] |
| TBI/controlled cortical impact | Human iPSC-derived cerebral organoids | [44,63] |
| TBI/weight-drop model | Human iPSC-derived neural progenitor cells | [64] |
| TBI/neurite transection model | Human iPSC-derived neurons | [65] |
| Oxygen-glucose deprivation/reperfusion | Human brain-derived microvascular endothelial cells from iPSCs | [66] |
| Hypoxia model | Human iPSC-derived neurons | [67] |
| Oxygen-glucose deprivation/reperfusion | Human iPSC-derived neurons | [27] |
| Oxygen-glucose deprivation | Human iPSC-derived neurons in 3D culture | [68] |
| Hypoxia | Human iPSC-derived neurons | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilipović, K.; Harej Hrkać, A.; Kučić, N.; Mršić-Pelčić, J. Modeling Central Nervous System Injury In Vitro: Current Status and Promising Future Strategies. Biomedicines 2023, 11, 94. https://doi.org/10.3390/biomedicines11010094
Pilipović K, Harej Hrkać A, Kučić N, Mršić-Pelčić J. Modeling Central Nervous System Injury In Vitro: Current Status and Promising Future Strategies. Biomedicines. 2023; 11(1):94. https://doi.org/10.3390/biomedicines11010094
Chicago/Turabian StylePilipović, Kristina, Anja Harej Hrkać, Natalia Kučić, and Jasenka Mršić-Pelčić. 2023. "Modeling Central Nervous System Injury In Vitro: Current Status and Promising Future Strategies" Biomedicines 11, no. 1: 94. https://doi.org/10.3390/biomedicines11010094
APA StylePilipović, K., Harej Hrkać, A., Kučić, N., & Mršić-Pelčić, J. (2023). Modeling Central Nervous System Injury In Vitro: Current Status and Promising Future Strategies. Biomedicines, 11(1), 94. https://doi.org/10.3390/biomedicines11010094