
Autologous Platelet-Rich Growth Factor Reduces M1 Macrophages and Modulates Inflammatory Microenvironments to Promote Sciatic Nerve Regeneration

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. PRGF Preparation for In Vitro and In Vivo Usage
2.2. Culture and Treatment of THP-1 Cells and RT4 SCs
2.3. Gene Expression Measurement Using Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. Enzyme-Linked Immunosorbent Assay (ELISA) for TNFα Secretion
2.5. The Animal Model for Sciatic Nerve Regeneration
2.6. Paraffin Block Preparation and Immunohistochemistry (IHC)
2.7. Statistical Analysis
3. Results
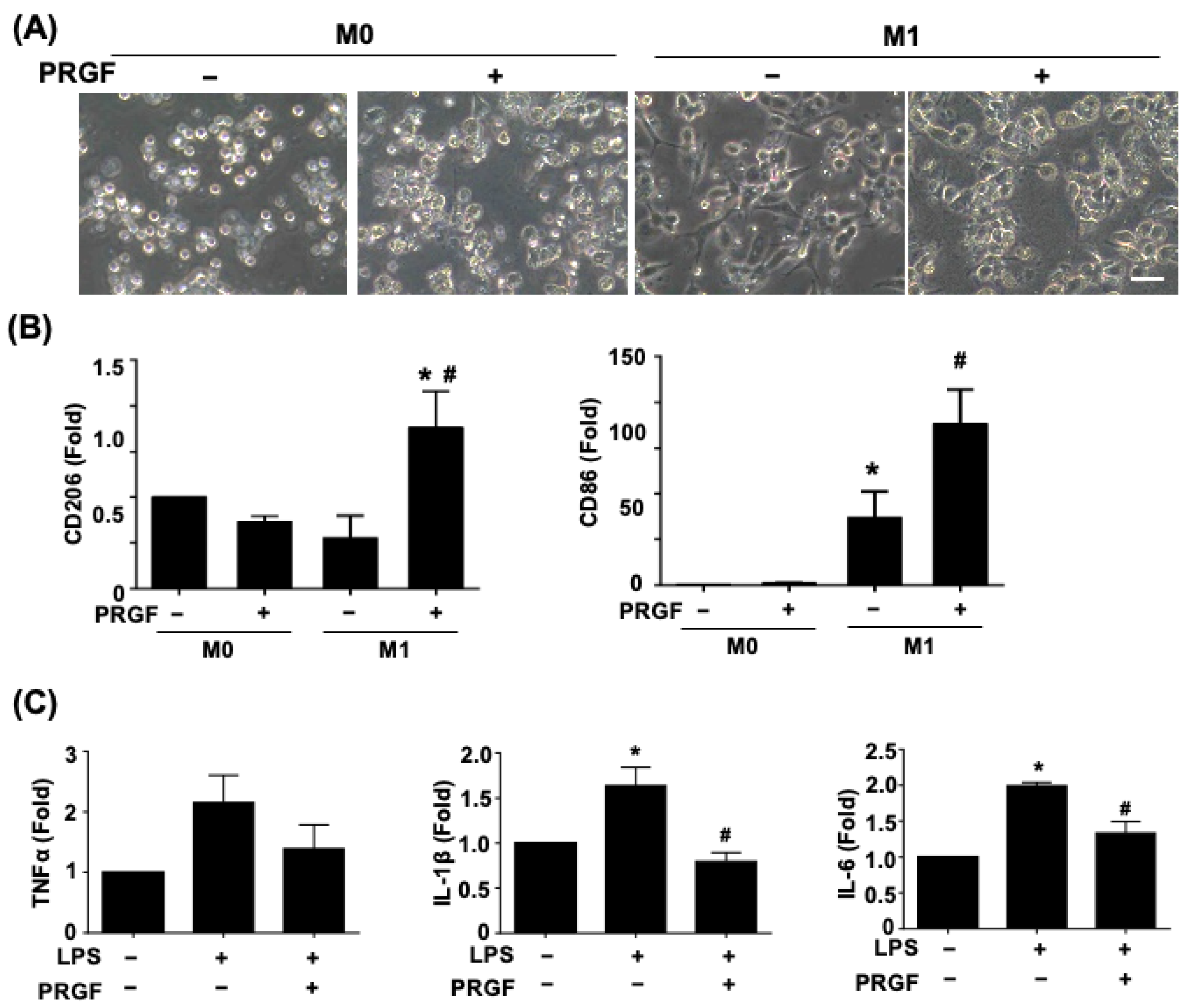
3.1. PRGF Attenuates the Pro-Inflammatory Cytokines during M1 Macrophage Induction
3.2. Regenerative Phenotypic Changes of Macrophages and SCs with PRGF Treatments
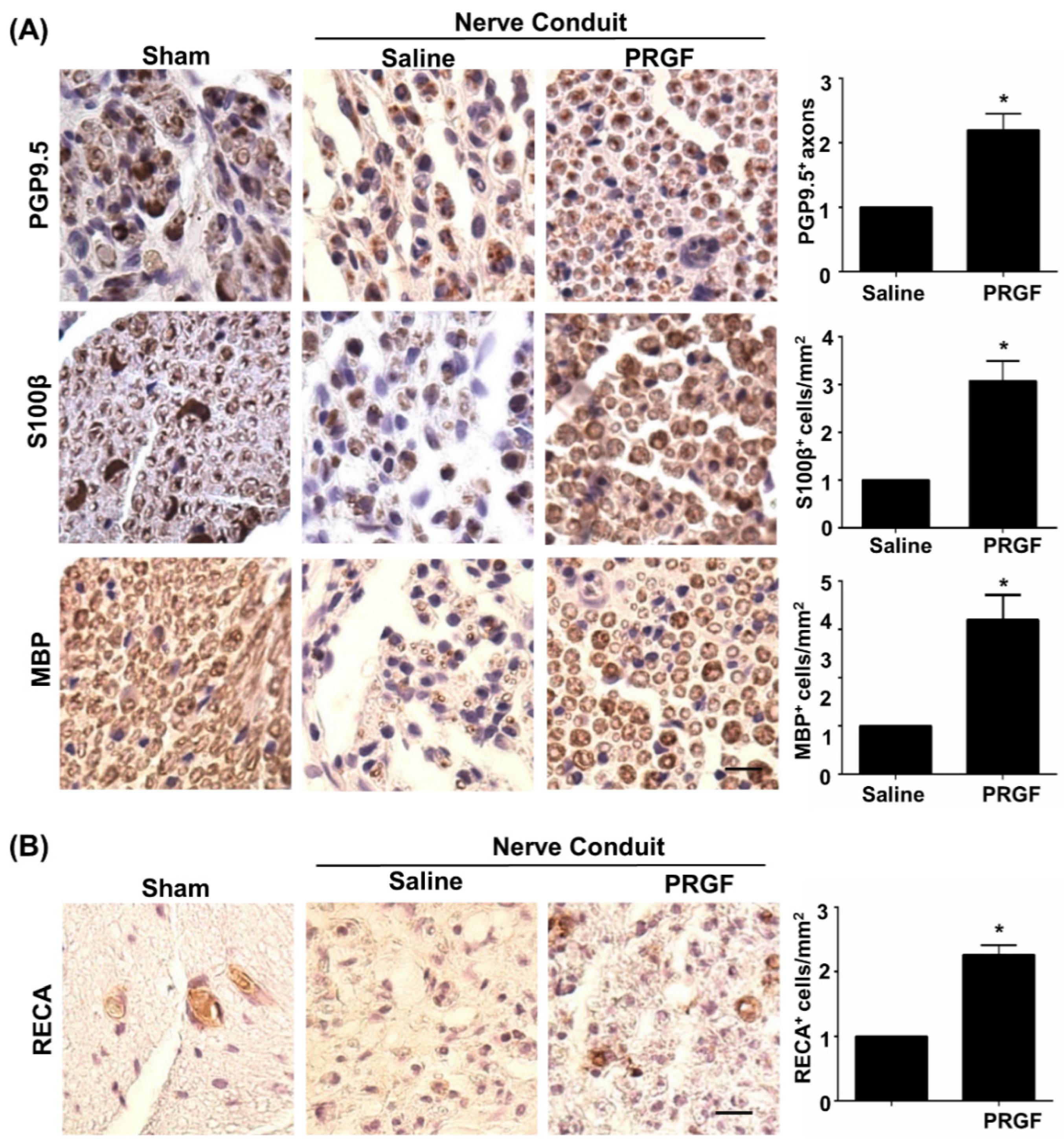
3.3. PRGF Alleviates Chronic Inflammation through Suppression of the M1 Macrophage during Nerve Regeneration
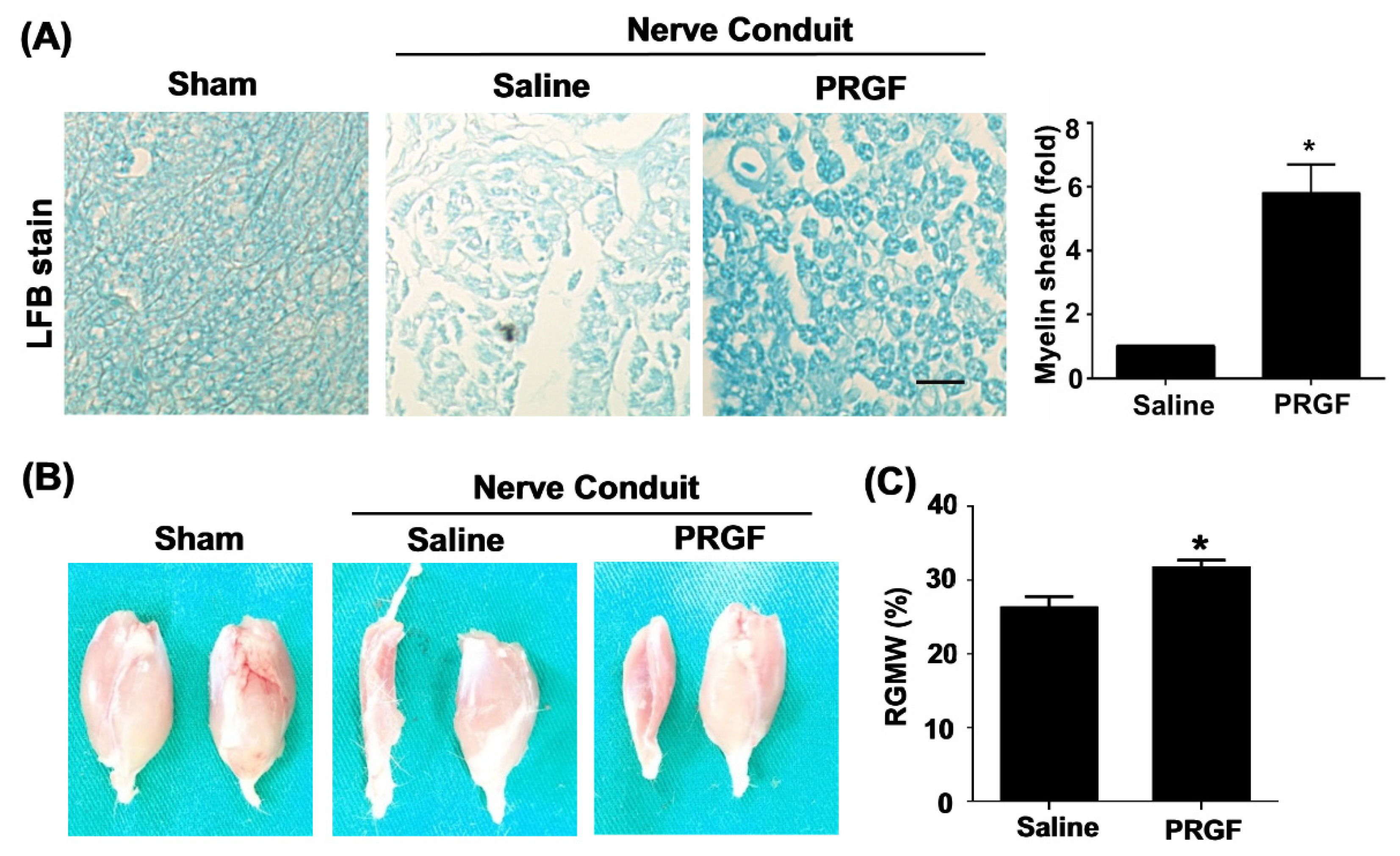
3.4. PRGF Fosters Nerve Regeneration and Reinnervation through an Increase in SCs Number, Regrown Axons, and Remyelination
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| ELISA | Enzyme-linked immunosorbent assay |
| FGF | Fibroblast growth factor |
| IL-1β | Interleukin-1β |
| IGF | Insulin-like growth factor |
| NGF | Nerve growth factor |
| PRP | Platelet-rich plasma |
| PRGF | Platelet-rich growth factors |
| PDGF | Platelet-derived growth factors |
| PBS | Phosphate-buffered saline |
| PGP9.5 | Protein gene product |
| RT | Room temperature |
| TGF-β | Transforming growth factor-β |
| RBCs | Red blood cells |
| SC | Schwann cell |
| Sprague Dawley | SD |
| TNFα | Tumor necrosis factor-α |
| MBP | Myelin basic protein |
| WBCs | White blood cells |
References
- Zigmond, R.E.; Echevarria, F.D. Macrophage biology in the peripheral nervous system after injury. Prog. Neurobiol. 2019, 173, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Msheik, Z.; El Massry, M.; Rovini, A.; Billet, F.; Desmouliere, A. The macrophage: A key player in the pathophysiology of peripheral neuropathies. J. Neuroinflamm. 2022, 19, 97. [Google Scholar] [CrossRef] [PubMed]
- Fregnan, F.; Muratori, L.; Simoes, A.R.; Giacobini-Robecchi, M.G.; Raimondo, S. Role of inflammatory cytokines in peripheral nerve injury. Neural Regen. Res. 2012, 7, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8, 110. [Google Scholar] [CrossRef]
- Liu, P.; Peng, J.; Han, G.H.; Ding, X.; Wei, S.; Gao, G.; Huang, K.; Chang, F.; Wang, Y. Role of macrophages in peripheral nerve injury and repair. Neural Regen. Res. 2019, 14, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Mokarram, N.; Merchant, A.; Mukhatyar, V.; Patel, G.; Bellamkonda, R.V. Effect of modulating macrophage phenotype on peripheral nerve repair. Biomaterials 2012, 33, 8793–8801. [Google Scholar] [CrossRef]
- Knoferle, J.; Ramljak, S.; Koch, J.C.; Tonges, L.; Asif, A.R.; Michel, U.; Wouters, F.S.; Heermann, S.; Krieglstein, K.; Zerr, I.; et al. TGF-beta 1 enhances neurite outgrowth via regulation of proteasome function and EFABP. Neurobiol. Dis. 2010, 38, 395–404. [Google Scholar] [CrossRef]
- Zhou, D.; Huang, C.; Lin, Z.; Zhan, S.; Kong, L.; Fang, C.; Li, J. Macrophage polarization and function with emphasis on the evolving roles of coordinated regulation of cellular signaling pathways. Cell Signal. 2014, 26, 192–197. [Google Scholar] [CrossRef]
- Huang, T.C.; Wu, H.L.; Chen, S.H.; Wang, Y.T.; Wu, C.C. Thrombomodulin facilitates peripheral nerve regeneration through regulating M1/M2 switching. J. Neuroinflamm. 2020, 17, 240. [Google Scholar] [CrossRef]
- Yadav, A.; Huang, T.C.; Chen, S.H.; Ramasamy, T.S.; Hsueh, Y.Y.; Lin, S.P.; Lu, F.I.; Liu, Y.H.; Wu, C.C. Sodium phenylbutyrate inhibits Schwann cell inflammation via HDAC and NFkappaB to promote axonal regeneration and remyelination. J. Neuroinflamm. 2021, 18, 238. [Google Scholar] [CrossRef]
- Gordon, T.; Sulaiman, O.; Boyd, J.G. Experimental strategies to promote functional recovery after peripheral nerve injuries. J. Peripher. Nerv. Syst. 2003, 8, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Wu, C.C.; Lin, S.C.; Tseng, W.L.; Huang, T.C.; Yadav, A.; Lu, F.I.; Liu, Y.H.; Lin, S.P.; Hsueh, Y.Y. Investigation of Neuropathology after Nerve Release in Chronic Constriction Injury of Rat Sciatic Nerve. Int. J. Mol. Sci. 2021, 22, 4746. [Google Scholar] [CrossRef]
- DeFrancesco-Lisowitz, A.; Lindborg, J.A.; Niemi, J.P.; Zigmond, R.E. The neuroimmunology of degeneration and regeneration in the peripheral nervous system. Neuroscience 2015, 302, 174–203. [Google Scholar] [CrossRef] [PubMed]
- Popovich, P.G.; Longbrake, E.E. Can the immune system be harnessed to repair the CNS? Nat. Rev. Neurosci. 2008, 9, 481–493. [Google Scholar] [CrossRef]
- Cui, Q. Actions of neurotrophic factors and their signaling pathways in neuronal survival and axonal regeneration. Mol. Neurobiol. 2006, 33, 155–179. [Google Scholar] [CrossRef]
- Hoke, A.; Redett, R.; Hameed, H.; Jari, R.; Zhou, C.; Li, Z.B.; Griffin, J.W.; Brushart, T.M. Schwann cells express motor and sensory phenotypes that regulate axon regeneration. J. Neurosci. 2006, 26, 9646–9655. [Google Scholar] [CrossRef]
- Zhou, Y.; Notterpek, L. Promoting peripheral myelin repair. Exp. Neurol. 2016, 283, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Xiao, G.; Zhang, K.; Ying, G.; Xu, H.; De Melo, B.A.G.; Li, S.; Liu, F.; Yetisen, A.K.; Jiang, N. Tacrolimus- and Nerve Growth Factor-Treated Allografts for Neural Tissue Regeneration. ACS Chem. Neurosci. 2019, 10, 1411–1419. [Google Scholar] [CrossRef]
- Liao, C.F.; Chen, C.C.; Lu, Y.W.; Yao, C.H.; Lin, J.H.; Way, T.D.; Yang, T.Y.; Chen, Y.S. Effects of endogenous inflammation signals elicited by nerve growth factor, interferon-gamma, and interleukin-4 on peripheral nerve regeneration. J. Biol. Eng. 2019, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Lu, S.Y.; Huang, T.C.; Huang, B.M.; Sun, H.S.; Yang, S.H.; Chuang, J.I.; Hsueh, Y.Y.; Wu, Y.T.; Wu, C.C. FGF9 induces functional differentiation to Schwann cells from human adipose derived stem cells. Theranostics 2020, 10, 2817–2831. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Lu, W.W.; Zhen, W.; Yang, D.; Peng, S. Novel biomaterial strategies for controlled growth factor delivery for biomedical applications. NPG Asia Mater. 2017, 9, e435. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X.; Wang, Y. Evaluation of Platelet-Rich Plasma Therapy for Peripheral Nerve Regeneration: A Critical Review of Literature. Front. Bioeng. Biotechnol. 2022, 10, 808248. [Google Scholar] [CrossRef] [PubMed]
- Weibrich, G.; Hansen, T.; Kleis, W.; Buch, R.; Hitzler, W.E. Effect of platelet concentration in platelet-rich plasma on peri-implant bone regeneration. Bone 2004, 34, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Graziani, F.; Ivanovski, S.; Cei, S.; Ducci, F.; Tonetti, M.; Gabriele, M. The in vitro effect of different PRP concentrations on osteoblasts and fibroblasts. Clin. Oral Implant. Res. 2006, 17, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Pascual, C.; Perez-Gonzalez, R.; Orive, G.; Carro, E. Intranasal PRGF-Endoret enhances neuronal survival and attenuates NF-kappaB-dependent inflammation process in a mouse model of Parkinson’s disease. J. Control. Release 2015, 203, 170–180. [Google Scholar] [CrossRef]
- Anitua, E.; Sanchez, M.; Orive, G.; Andia, I. The potential impact of the preparation rich in growth factors (PRGF) in different medical fields. Biomaterials 2007, 28, 4551–4560. [Google Scholar] [CrossRef]
- Anitua, E.; Zalduendo, M.M.; Prado, R.; Alkhraisat, M.H.; Orive, G. Morphogen and proinflammatory cytokine release kinetics from PRGF-Endoret fibrin scaffolds: Evaluation of the effect of leukocyte inclusion. J. Biomed. Mater. Res. A 2015, 103, 1011–1020. [Google Scholar] [CrossRef]
- Nishiyama, K.; Okudera, T.; Watanabe, T.; Isobe, K.; Suzuki, M.; Masuki, H.; Okudera, H.; Uematsu, K.; Nakata, K.; Kawase, T. Basic characteristics of plasma rich in growth factors (PRGF): Blood cell components and biological effects. Clin. Exp. Dent. Res. 2016, 2, 96–103. [Google Scholar] [CrossRef]
- Anitua, E.; Pino, A.; Aspe, L.; Martinez, M.; Garcia, A.; Goni, F.; Troya, M. Anti-inflammatory effect of different PRGF formulations on cutaneous surface. J. Tissue Viability 2021, 30, 183–189. [Google Scholar] [CrossRef]
- Sanchez, M.; Anitua, E.; Delgado, D.; Prado, R.; Sanchez, P.; Fiz, N.; Guadilla, J.; Azofra, J.; Pompei, O.; Orive, G.; et al. Ultrasound-guided plasma rich in growth factors injections and scaffolds hasten motor nerve functional recovery in an ovine model of nerve crush injury. J. Tissue Eng. Regen. Med. 2017, 11, 1619–1629. [Google Scholar] [CrossRef]
- Asai, T. Nursing practice and medical dispute. 11. keypoints in prevention of basic medical accidents. Kango Gijutsu 1986, 32, 1976–1977. [Google Scholar] [PubMed]
- Torul, D.; Bereket, M.C.; Onger, M.E.; Altun, G. Comparison of the Regenerative Effects of Platelet-Rich Fibrin and Plasma Rich in Growth Factors on Injured Peripheral Nerve: An Experimental Study. J. Oral Maxillofac. Surg. 2018, 76, 1823.e1–1823.e12. [Google Scholar] [CrossRef] [PubMed]
- Escobar, G.; Escobar, A.; Ascui, G.; Tempio, F.I.; Ortiz, M.C.; Perez, C.A.; Lopez, M.N. Pure platelet-rich plasma and supernatant of calcium-activated P-PRP induce different phenotypes of human macrophages. Regen. Med. 2018, 13, 427–441. [Google Scholar] [CrossRef]
- Hudgens, J.L.; Sugg, K.B.; Grekin, J.A.; Gumucio, J.P.; Bedi, A.; Mendias, C.L. Platelet-Rich Plasma Activates Proinflammatory Signaling Pathways and Induces Oxidative Stress in Tendon Fibroblasts. Am. J. Sports Med. 2016, 44, 1931–1940. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Liu, W.; Zhou, Y.; Jiang, X.; Liu, Y.; Yang, Q.; Shao, L. Concentrated growth factor regulates the macrophage-mediated immune response. Regen. Biomater. 2021, 8, rbab049. [Google Scholar] [CrossRef]
- Uchiyama, R.; Toyoda, E.; Maehara, M.; Wasai, S.; Omura, H.; Watanabe, M.; Sato, M. Effect of Platelet-Rich Plasma on M1/M2 Macrophage Polarization. Int. J. Mol. Sci. 2021, 22, 2336. [Google Scholar] [CrossRef]
- Zhao, B.; Pan, Y.; Xu, H.; Song, X. Kindlin-1 Regulates Astrocyte Activation and Pain Sensitivity in Rats With Neuropathic Pain. Reg. Anesth. Pain Med. 2018, 43, 547–553. [Google Scholar] [CrossRef]
- Huang, C.F.; Chang, Y.J.; Hsueh, Y.Y.; Huang, C.W.; Wang, D.H.; Huang, T.C.; Wu, Y.T.; Su, F.C.; Hughes, M.; Chuong, C.M.; et al. Assembling Composite Dermal Papilla Spheres with Adipose-derived Stem Cells to Enhance Hair Follicle Induction. Sci. Rep. 2016, 6, 26436. [Google Scholar] [CrossRef]
- Liu, C.; Tsai, A.L.; Li, P.C.; Huang, C.W.; Wu, C.C. Endothelial differentiation of bone marrow mesenchyme stem cells applicable to hypoxia and increased migration through Akt and NFkappaB signals. Stem Cell Res. Ther. 2017, 8, 29. [Google Scholar] [CrossRef]
- Kroner, A.; Greenhalgh, A.D.; Zarruk, J.G.; Passos Dos Santos, R.; Gaestel, M.; David, S. TNF and increased intracellular iron alter macrophage polarization to a detrimental M1 phenotype in the injured spinal cord. Neuron 2014, 83, 1098–1116. [Google Scholar] [CrossRef]
- Kopper, T.J.; Zhang, B.; Bailey, W.M.; Bethel, K.E.; Gensel, J.C. The effects of myelin on macrophage activation are phenotypic specific via cPLA2 in the context of spinal cord injury inflammation. Sci. Rep. 2021, 11, 6341. [Google Scholar] [CrossRef] [PubMed]
- Siqueira Mietto, B.; Kroner, A.; Girolami, E.I.; Santos-Nogueira, E.; Zhang, J.; David, S. Role of IL-10 in Resolution of Inflammation and Functional Recovery after Peripheral Nerve Injury. J. Neurosci. 2015, 35, 16431–16442. [Google Scholar] [CrossRef] [PubMed]
- Buttner, R.; Schulz, A.; Reuter, M.; Akula, A.K.; Mindos, T.; Carlstedt, A.; Riecken, L.B.; Baader, S.L.; Bauer, R.; Morrison, H. Inflammaging impairs peripheral nerve maintenance and regeneration. Aging Cell 2018, 17, e12833. [Google Scholar] [CrossRef] [PubMed]
- Ydens, E.; Cauwels, A.; Asselbergh, B.; Goethals, S.; Peeraer, L.; Lornet, G.; Almeida-Souza, L.; Van Ginderachter, J.A.; Timmerman, V.; Janssens, S. Acute injury in the peripheral nervous system triggers an alternative macrophage response. J. Neuroinflamm. 2012, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Lana, J.F.; Huber, S.C.; Purita, J.; Tambeli, C.H.; Santos, G.S.; Paulus, C.; Annichino-Bizzacchi, J.M. Leukocyte-rich PRP versus leukocyte-poor PRP—The role of monocyte/macrophage function in the healing cascade. J. Clin. Orthop. Trauma 2019, 10, S7–S12. [Google Scholar] [CrossRef]
- Nishio, H.; Saita, Y.; Kobayashi, Y.; Takaku, T.; Fukusato, S.; Uchino, S.; Wakayama, T.; Ikeda, H.; Kaneko, K. Platelet-rich plasma promotes recruitment of macrophages in the process of tendon healing. Regen. Ther. 2020, 14, 262–270. [Google Scholar] [CrossRef]
- Fang, S.Y.; Huang, C.W.; Huang, T.C.; Yadav, A.; Chiu, J.J.; Wu, C.C. Reduction in MicroRNA-4488 Expression Induces NFkappaB Translocation in Venous Endothelial Cells Under Arterial Flow. Cardiovasc. Drugs Ther. 2021, 35, 61–71. [Google Scholar] [CrossRef]
- Lichtenfels, M.; Colome, L.; Sebben, A.D.; Braga-Silva, J. Effect of Platelet Rich Plasma and Platelet Rich Fibrin on sciatic nerve regeneration in a rat model. Microsurgery 2013, 33, 383–390. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jeon, W.J.; Kim, D.H.; Rhyu, I.J.; Kim, Y.H.; Youn, I.; Park, J.W. An inside-out vein graft filled with platelet-rich plasma for repair of a short sciatic nerve defect in rats. Neural Regen. Res. 2014, 9, 1351–1357. [Google Scholar] [CrossRef]
- Yu, W.; Wang, J.; Yin, J. Platelet-rich plasma: A promising product for treatment of peripheral nerve regeneration after nerve injury. Int. J. Neurosci. 2011, 121, 176–180. [Google Scholar] [CrossRef]
- Sariguney, Y.; Yavuzer, R.; Elmas, C.; Yenicesu, I.; Bolay, H.; Atabay, K. Effect of platelet-rich plasma on peripheral nerve regeneration. J. Reconstr. Microsurg. 2008, 24, 159–167. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, H.; Kantarci, A.; Deady, J.; Hasturk, H.; Liu, H.; Alshahat, M.; Van Dyke, T.E. Platelet-rich plasma: Growth factors and pro- and anti-inflammatory properties. J. Periodontol. 2007, 78, 661–669. [Google Scholar] [CrossRef]
- Wang, Z.; Mudalal, M.; Sun, Y.; Liu, Y.; Wang, J.; Wang, Y.; Sun, X.; Zhou, Y. The Effects of Leukocyte-Platelet Rich Fibrin (L-PRF) on Suppression of the Expressions of the Pro-Inflammatory Cytokines, and Proliferation of Schwann Cell, and Neurotrophic Factors. Sci. Rep. 2020, 10, 2421. [Google Scholar] [CrossRef]
- Tohidnezhad, M.; Bayer, A.; Rasuo, B.; Hock, J.V.P.; Kweider, N.; Fragoulis, A.; Sonmez, T.T.; Jahr, H.; Pufe, T.; Lippross, S. Platelet-Released Growth Factors Modulate the Secretion of Cytokines in Synoviocytes under Inflammatory Joint Disease. Mediat. Inflamm. 2017, 2017, 1046438. [Google Scholar] [CrossRef]
- Jain, R.K.; Au, P.; Tam, J.; Duda, D.G.; Fukumura, D. Engineering vascularized tissue. Nat. Biotechnol. 2005, 23, 821–823. [Google Scholar] [CrossRef]
- Novosel, E.C.; Kleinhans, C.; Kluger, P.J. Vascularization is the key challenge in tissue engineering. Adv. Drug Deliv. Rev. 2011, 63, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Cattin, A.L.; Burden, J.J.; Van Emmenis, L.; Mackenzie, F.E.; Hoving, J.J.; Garcia Calavia, N.; Guo, Y.; McLaughlin, M.; Rosenberg, L.H.; Quereda, V.; et al. Macrophage-Induced Blood Vessels Guide Schwann Cell-Mediated Regeneration of Peripheral Nerves. Cell 2015, 162, 1127–1139. [Google Scholar] [CrossRef]
- Kim, E.S.; Kim, J.J.; Park, E.J. Angiogenic factor-enriched platelet-rich plasma enhances in vivo bone formation around alloplastic graft material. J. Adv. Prosthodont. 2010, 2, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, J.; Putnam, A.J. Sculpting the blank slate: How fibrin’s support of vascularization can inspire biomaterial design. Acta Biomater. 2014, 10, 1515–1523. [Google Scholar] [CrossRef]
- Chang, Y.J.; Li, Y.S.; Wu, C.C.; Wang, K.C.; Huang, T.C.; Chen, Z.; Chien, S. Extracellular MicroRNA-92a Mediates Endothelial Cell-Macrophage Communication. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 2492–2504. [Google Scholar] [CrossRef]
- Anitua, E.; Pelacho, B.; Prado, R.; Aguirre, J.J.; Sanchez, M.; Padilla, S.; Aranguren, X.L.; Abizanda, G.; Collantes, M.; Hernandez, M.; et al. Infiltration of plasma rich in growth factors enhances in vivo angiogenesis and improves reperfusion and tissue remodeling after severe hind limb ischemia. J. Control. Release 2015, 202, 31–39. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (F)/Reverse (R) | Primer Sequence |
|---|---|---|
| TNFα | F | TCAACCTCCTCTCTGCCATC |
| R | CCAAAGTAGACCTGCCCAGA | |
| IL1β | F | CTGTCCTGCGTGTTGAAAGA |
| R | CTGCTTGAGAGGTGCTGATG | |
| IL6 | F | AGGAGACTTGCCTGGTGAAA |
| R | CAGGGGTGGTTATTGCATCT | |
| CD86 | F | GACGCGGCTTTTATCTTCAC |
| R | CCCTCTCCATTGTGTTGGTT | |
| CD206 | F | GATGGGTGTCCGAATCTCAG |
| R | TTCCACCTGCTCCATAAACC | |
| GAPDH | F | CATCAAGAAGGTGGTGAAGC |
| R | TGACAAAGTGGTCGTTGAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, A.; Ramasamy, T.S.; Lin, S.-C.; Chen, S.-H.; Lu, J.; Liu, Y.-H.; Lu, F.-I.; Hsueh, Y.-Y.; Lin, S.-P.; Wu, C.-C. Autologous Platelet-Rich Growth Factor Reduces M1 Macrophages and Modulates Inflammatory Microenvironments to Promote Sciatic Nerve Regeneration. Biomedicines 2022, 10, 1991. https://doi.org/10.3390/biomedicines10081991
Yadav A, Ramasamy TS, Lin S-C, Chen S-H, Lu J, Liu Y-H, Lu F-I, Hsueh Y-Y, Lin S-P, Wu C-C. Autologous Platelet-Rich Growth Factor Reduces M1 Macrophages and Modulates Inflammatory Microenvironments to Promote Sciatic Nerve Regeneration. Biomedicines. 2022; 10(8):1991. https://doi.org/10.3390/biomedicines10081991
Chicago/Turabian StyleYadav, Anjali, Thamil Selvee Ramasamy, Sheng-Che Lin, Szu-Han Chen, Jean Lu, Ya-Hsin Liu, Fu-I Lu, Yuan-Yu Hsueh, Shau-Ping Lin, and Chia-Ching Wu. 2022. "Autologous Platelet-Rich Growth Factor Reduces M1 Macrophages and Modulates Inflammatory Microenvironments to Promote Sciatic Nerve Regeneration" Biomedicines 10, no. 8: 1991. https://doi.org/10.3390/biomedicines10081991
APA StyleYadav, A., Ramasamy, T. S., Lin, S.-C., Chen, S.-H., Lu, J., Liu, Y.-H., Lu, F.-I., Hsueh, Y.-Y., Lin, S.-P., & Wu, C.-C. (2022). Autologous Platelet-Rich Growth Factor Reduces M1 Macrophages and Modulates Inflammatory Microenvironments to Promote Sciatic Nerve Regeneration. Biomedicines, 10(8), 1991. https://doi.org/10.3390/biomedicines10081991