
Contribution of the Environment, Epigenetic Mechanisms and Non-Coding RNAs in Psoriasis

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Microbiome and Environmental Risk Factors
2.1. Skin and Gut Microbiome
2.2. Lifestyle
3. DNA Methylation
3.1. DNA Methylation in KCs
3.2. DNA Methylation in Immune Cells
4. Histone Modifications
4.1. Histone Modifications in KCs
4.2. Histone Modifications in Immune Cells
5. Non-Coding RNAs
5.1. MiRNAs in KCs
5.2. MiRNAs in Immune Cells and Serum
5.3. LncRNAs in PsO
6. Therapeutic Approaches Targeting the Epigenetic Mechanisms
7. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, Regional, and Worldwide Epidemiology of Psoriasis: Systematic Analysis and Modelling Study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Mehta, M.D.; Schupp, C.W.; Gondo, G.C.; Bell, S.J.; Griffiths, C.E.M. Psoriasis Prevalence in Adults in the United States. JAMA Dermatol. 2021, 157, 940. [Google Scholar] [CrossRef] [PubMed]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef] [PubMed]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of Psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef]
- Chang, H.-W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.; et al. Alteration of the Cutaneous Microbiome in Psoriasis and Potential Role in Th17 Polarization. Microbiome 2018, 6, 154. [Google Scholar] [CrossRef]
- Fuentes-Duculan, J.; Bonifacio, K.; Kunjravia, N.; Cueto, I.; Li, X.; Garcet, S.; Krueger, J.G. Autoantigens ADAMTSL5 and LL-37 Are Significantly Upregulated in Active Psoriasis and Associated with Dendritic Cells and Macrophages. J. Investig. Dermatol. 2017, 137, S1. [Google Scholar] [CrossRef]
- Hänsel, A.; Günther, C.; Ingwersen, J.; Starke, J.; Schmitz, M.; Bachmann, M.; Meurer, M.; Rieber, E.P.; Schäkel, K. Human Slan (6-Sulfo LacNAc) Dendritic Cells Are Inflammatory Dermal Dendritic Cells in Psoriasis and Drive Strong T 17/T 1 T-Cell Responses. J. Allergy Clin. Immunol. 2011, 127, 787–794. [Google Scholar] [CrossRef]
- Fitch, E.; Harper, E.; Skorcheva, I.; Kurtz, S.E.; Blauvelt, A. Pathophysiology of Psoriasis: Recent Advances on IL-23 and Th17 Cytokines. Curr. Rheumatol. Rep. 2007, 9, 461–467. [Google Scholar] [CrossRef]
- Di Cesare, A.; Di Meglio, P.; Nestle, F.O. The IL-23/Th17 Axis in the Immunopathogenesis of Psoriasis. J. Investig. Dermatol. 2009, 129, 1339–1350. [Google Scholar] [CrossRef]
- Banno, T.; Gazel, A.; Blumenberg, M. Effects of Tumor Necrosis Factor-α (TNFα) in Epidermal Keratinocytes Revealed Using Global Transcriptional Profiling. J. Biol. Chem. 2004, 279, 32633–32642. [Google Scholar] [CrossRef]
- Chen, X.; Hamano, R.; Subleski, J.J.; Hurwitz, A.A.; Howard, O.M.Z.; Oppenheim, J.J. Expression of Costimulatory TNFR2 Induces Resistance of CD4 + FoxP3 − Conventional T Cells to Suppression by CD4 + FoxP3 + Regulatory T Cells. J. Immunol. 2010, 185, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.P.; Erpenbeck, L. The Interleukin-23/Interleukin-17 Axis Links Adaptive and Innate Immunity in Psoriasis. Front. Immunol. 2018, 9, 1323. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, L.C.; Stuart, P.E.; Tian, C.; Gudjonsson, J.E.; Das, S.; Zawistowski, M.; Ellinghaus, E.; Barker, J.N.; Chandran, V.; Dand, N.; et al. Large Scale Meta-Analysis Characterizes Genetic Architecture for Common Psoriasis Associated Variants. Nat. Commun. 2017, 8, 15382. [Google Scholar] [CrossRef]
- Naldi, L. Psoriasis and Smoking: Links and Risks. PTT 2016, 6, 65. [Google Scholar] [CrossRef]
- Wolters, M. Diet and Psoriasis: Experimental Data and Clinical Evidence. Br. J. Dermatol. 2005, 153, 706–714. [Google Scholar] [CrossRef]
- Søyland, E.; Heier, I.; Rodríguez-Gallego, C.; Mollnes, T.E.; Johansen, F.-E.; Holven, K.B.; Halvorsen, B.; Aukrust, P.; Jahnsen, F.L.; de la Rosa Carrillo, D.; et al. Sun Exposure Induces Rapid Immunological Changes in Skin and Peripheral Blood in Patients with Psoriasis: Immunological Changes in Sun-Exposed Patients with Psoriasis. Br. J. Dermatol. 2011, 164, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Schuebel, K.; Gitik, M.; Domschke, K.; Goldman, D. Making Sense of Epigenetics. Int. J. Neuropsychopharmacol. 2016, 19, pyw058. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.-Z.; Jiang, J.; Duan, C.-G. The Crosstalk Between Epigenetic Mechanisms and Alternative RNA Processing Regulation. Front. Genet. 2020, 11, 998. [Google Scholar] [CrossRef]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in Cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef]
- Mazzone, R.; Zwergel, C.; Artico, M.; Taurone, S.; Ralli, M.; Greco, A.; Mai, A. The Emerging Role of Epigenetics in Human Autoimmune Disorders. Clin. Epigenet. 2019, 11, 34. [Google Scholar] [CrossRef]
- Hsu, D.K.; Fung, M.A.; Chen, H.-L. Role of Skin and Gut Microbiota in the Pathogenesis of Psoriasis, an Inflammatory Skin Disease. Med. Microecol. 2020, 4, 100016. [Google Scholar] [CrossRef]
- Roszkiewicz, M.; Dopytalska, K.; Szymańska, E.; Jakimiuk, A.; Walecka, I. Environmental Risk Factors and Epigenetic Alternations in Psoriasis. Ann. Agric. Environ. Med. 2020, 27, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.; Travers, J.B.; Giorno, R.; Norris, D.A.; Skinner, R.; Aelion, J.; Kazemi, L.V.; Kim, M.H.; Trumble, A.E.; Kotb, M. Evidence for a Streptococcal Superantigen-Driven Process in Acute Guttate Psoriasis. J. Clin. Investig. 1995, 96, 2106–2112. [Google Scholar] [CrossRef]
- Benhadou, F.; Mintoff, D.; Schnebert, B.; Thio, H. Psoriasis and Microbiota: A Systematic Review. Diseases 2018, 6, 47. [Google Scholar] [CrossRef]
- Paulino, L.C.; Tseng, C.-H.; Blaser, M.J. Analysis of Malassezia Microbiota in Healthy Superficial Human Skin and in Psoriatic Lesions by Multiplex Real-Time PCR: Malassezia Microbiota in Health and Psoriasis by QPCR. FEMS Yeast Res. 2008, 8, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Allhorn, M.; Arve, S.; Brüggemann, H.; Lood, R. A Novel Enzyme with Antioxidant Capacity Produced by the Ubiquitous Skin Colonizer Propionibacterium Acnes. Sci. Rep. 2016, 6, 36412. [Google Scholar] [CrossRef] [PubMed]
- Baroni, A.; Orlando, M.; Donnarumma, G.; Farro, P.; Iovene, M.R.; Tufano, M.A.; Buommino, E. Toll-like Receptor 2 (TLR2) Mediates Intracellular Signalling in Human Keratinocytes in Response to Malassezia Furfur. Arch. Derm. Res. 2006, 297, 280–288. [Google Scholar] [CrossRef]
- Fyhrquist, N.; Muirhead, G.; Prast-Nielsen, S.; Jeanmougin, M.; Olah, P.; Skoog, T.; Jules-Clement, G.; Feld, M.; Barrientos-Somarribas, M.; Sinkko, H.; et al. Microbe-Host Interplay in Atopic Dermatitis and Psoriasis. Nat. Commun. 2019, 10, 4703. [Google Scholar] [CrossRef]
- Reizis, B.; Colonna, M.; Trinchieri, G.; Barrat, F.; Gilliet, M. Plasmacytoid Dendritic Cells: One-Trick Ponies or Workhorses of the Immune System? Nat. Rev. Immunol. 2011, 11, 558–565. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Gómez, J.; Delgado, S.; Requena-López, S.; Queiro-Silva, R.; Margolles, A.; Coto, E.; Sánchez, B.; Coto-Segura, P. Gut Microbiota Dysbiosis in a Cohort of Patients with Psoriasis. Br. J. Dermatol. 2019, 181, 1287–1295. [Google Scholar] [CrossRef]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M.; Marmon, S.; Neimann, A.; Brusca, S.; Patel, T.; et al. Decreased Bacterial Diversity Characterizes the Altered Gut Microbiota in Patients With Psoriatic Arthritis, Resembling Dysbiosis in Inflammatory Bowel Disease: Gut Microbiota in PsA. Arthritis Rheumatol. 2015, 67, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.; Thaiss, C.A.; Levy, M.; Elinav, E. The Cross Talk between Microbiota and the Immune System: Metabolites Take Center Stage. Curr. Opin. Immunol. 2014, 30, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.I.; Kapila, R. Dietary Metabolites Derived from Gut Microbiota: Critical Modulators of Epigenetic Changes in Mammals. Nutr. Rev. 2017, 75, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Remely, M.; Lovrecic, L.; de la Garza, A.L.; Migliore, L.; Peterlin, B.; Milagro, F.I.; Martinez, A.J.; Haslberger, A.G. Therapeutic Perspectives of Epigenetically Active Nutrients: Therapeutic Epigenetically Active Nutrients. Br. J. Pharm. 2015, 172, 2756–2768. [Google Scholar] [CrossRef] [PubMed]
- Fujii, R.; Yamada, H.; Munetsuna, E.; Yamazaki, M.; Mizuno, G.; Ando, Y.; Maeda, K.; Tsuboi, Y.; Ohashi, K.; Ishikawa, H.; et al. Dietary Fish and ω-3 Polyunsaturated Fatty Acids Are Associated with Leukocyte ABCA1 DNA Methylation Levels. Nutrition 2021, 81, 110951. [Google Scholar] [CrossRef]
- Karimi, M.; Vedin, I.; Freund Levi, Y.; Basun, H.; Faxén Irving, G.; Eriksdotter, M.; Wahlund, L.-O.; Schultzberg, M.; Hjorth, E.; Cederholm, T.; et al. DHA-Rich n–3 Fatty Acid Supplementation Decreases DNA Methylation in Blood Leukocytes: The OmegAD Study. Am. J. Clin. Nutr. 2017, 106, 1157–1165. [Google Scholar] [CrossRef]
- Naldi, L.; Chatenoud, L.; Linder, D.; Belloni Fortina, A.; Peserico, A.; Virgili, A.R.; Bruni, P.L.; Ingordo, V.; Scocco, G.L.; Solaroli, C.; et al. Cigarette Smoking, Body Mass Index, and Stressful Life Events as Risk Factors for Psoriasis: Results from an Italian Case–Control Study. J. Investig. Dermatol. 2005, 125, 61–67. [Google Scholar] [CrossRef]
- Setty, A.R.; Curhan, G.; Choi, H.K. Obesity, Waist Circumference, Weight Change, and the Risk of Psoriasis in Women. ARCH Intern. Med. 2007, 167, 6. [Google Scholar] [CrossRef]
- Snekvik, I.; Smith, C.H.; Nilsen, T.I.L.; Langan, S.M.; Modalsli, E.H.; Romundstad, P.R.; Saunes, M. Obesity, Waist Circumference, Weight Change, and Risk of Incident Psoriasis: Prospective Data from the HUNT Study. J. Investig. Dermatol. 2017, 137, 2484–2490. [Google Scholar] [CrossRef]
- Zhu, K.-J.; Zhang, C.; Li, M.; Zhu, C.-Y.; Shi, G.; Fan, Y.-M. Leptin Levels in Patients with Psoriasis: A Meta-Analysis. Clin. Exp. Dermatol. 2013, 38, 478–483. [Google Scholar] [CrossRef]
- Kong, Y.; Zhang, S.; Wu, R.; Su, X.; Peng, D.; Zhao, M.; Su, Y. New Insights into Different Adipokines in Linking the Pathophysiology of Obesity and Psoriasis. Lipids Health Dis. 2019, 18, 171. [Google Scholar] [CrossRef] [PubMed]
- Kunz, M.; Simon, J.C.; Saalbach, A. Psoriasis: Obesity and Fatty Acids. Front. Immunol. 2019, 10, 1807. [Google Scholar] [CrossRef] [PubMed]
- Farkas, Á.; Kemény, L. Alcohol, Liver, Systemic Inflammation and Skin: A Focus on Patients with Psoriasis. Ski. Pharm. Physiol. 2013, 26, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Joehanes, R.; Just, A.C.; Marioni, R.E.; Pilling, L.C.; Reynolds, L.M.; Mandaviya, P.R.; Guan, W.; Xu, T.; Elks, C.E.; Aslibekyan, S.; et al. Epigenetic Signatures of Cigarette Smoking. Circ. Cardiovasc. Genet. 2016, 9, 436–447. [Google Scholar] [CrossRef]
- Sundar, I.K.; Nevid, M.Z.; Friedman, A.E.; Rahman, I. Cigarette Smoke Induces Distinct Histone Modifications in Lung Cells: Implications for the Pathogenesis of COPD and Lung Cancer. J. Proteome Res. 2014, 13, 982–996. [Google Scholar] [CrossRef]
- Izzotti, A.; Larghero, P.; Longobardi, M.; Cartiglia, C.; Camoirano, A.; Steele, V.E.; De Flora, S. Dose-Responsiveness and Persistence of MicroRNA Expression Alterations Induced by Cigarette Smoke in Mouse Lung. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2011, 717, 9–16. [Google Scholar] [CrossRef]
- Bi, H.; Zhou, J.; Wu, D.; Gao, W.; Li, L.; Yu, L.; Liu, F.; Huang, M.; Adcock, I.M.; Barnes, P.J.; et al. Microarray Analysis of Long Non-Coding RNAs in COPD Lung Tissue. Inflamm. Res. 2015, 64, 119–126. [Google Scholar] [CrossRef]
- Evers, A.W.M.; Verhoeven, E.W.M.; Kraaimaat, F.W.; De Jong, E.M.G.J.; De Brouwer, S.J.M.; Schalkwijk, J.; Sweep, F.C.G.J.; Van De Kerkhof, P.C.M. How Stress Gets under the Skin: Cortisol and Stress Reactivity in Psoriasis: Stress Reactivity in Psoriasis. Br. J. Dermatol. 2010, 163, 986–991. [Google Scholar] [CrossRef]
- Thomassin, H. Glucocorticoid-Induced DNA Demethylation and Gene Memory during Development. EMBO J. 2001, 20, 1974–1983. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Zill, J.M.; Dirmaier, J.; Augustin, M.; Dwinger, S.; Christalle, E.; Härter, M.; Mrowietz, U. Psychosocial Distress of Patients with Psoriasis: Protocol for an Assessment of Care Needs and the Development of a Supportive Intervention. JMIR Res. Protoc. 2018, 7, e22. [Google Scholar] [CrossRef] [PubMed]
- Vockley, C.M.; D’Ippolito, A.M.; McDowell, I.C.; Majoros, W.H.; Safi, A.; Song, L.; Crawford, G.E.; Reddy, T.E. Direct GR Binding Sites Potentiate Clusters of TF Binding across the Human Genome. Cell 2016, 166, 1269–1281.e19. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y.; Roy, B.; Lugli, G.; Rizavi, H.; Zhang, H.; Smalheiser, N.R. Chronic Corticosterone-Mediated Dysregulation of MicroRNA Network in Prefrontal Cortex of Rats: Relevance to Depression Pathophysiology. Transl. Psychiatry 2015, 5, e682. [Google Scholar] [CrossRef] [PubMed]
- Rutter, K.J.; Watson, R.E.B.; Cotterell, L.F.; Brenn, T.; Griffiths, C.E.M.; Rhodes, L.E. Severely Photosensitive Psoriasis: A Phenotypically Defined Patient Subset. J. Investig. Dermatol. 2009, 129, 2861–2867. [Google Scholar] [CrossRef] [PubMed]
- Cicarma, E.; Mørk, C.; Porojnicu, A.C.; Juzeniene, A.; Tam, T.T.T.; Dahlback, A.; Moan, J. Influence of Narrowband UVB Phototherapy on Vitamin D and Folate Status: Narrowband UVB, Vitamin D and Folate. Exp. Dermatol. 2009, 19, e67–e72. [Google Scholar] [CrossRef] [PubMed]
- Sloka, S.; Silva, C.; Wang, J.; Yong, V.W. Predominance of Th2 Polarization by Vitamin D through a STAT6-Dependent Mechanism. J. Neuroinflamm. 2011, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Komuves, L.; Yu, Q.-C.; Elalieh, H.; Ng, D.C.; Leary, C.; Chang, S.; Crumrine, D.; Bikle, D.D.; Yoshizawa, T.; et al. Lack of the Vitamin D Receptor Is Associated with Reduced Epidermal Differentiation and Hair Follicle Growth. J. Investig. Dermatol. 2002, 118, 11–16. [Google Scholar] [CrossRef]
- Fetahu, I.S.; Höbaus, J.; Kállay, E. Vitamin D and the Epigenome. Front. Physiol. 2014, 5. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J. DNA Methyltransferases and Their Roles in Tumorigenesis. Biomark. Res. 2017, 5, 1. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacol 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, Y.; Cui, L.; Shi, Y.; Guo, C. Advances in the Pathogenesis of Psoriasis: From Keratinocyte Perspective. Cell Death Dis. 2022, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in Inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef]
- Christmann, C.; Zenker, S.; Martens, L.; Hübner, J.; Loser, K.; Vogl, T.; Roth, J. Interleukin 17 Promotes Expression of Alarmins S100A8 and S100A9 During the Inflammatory Response of Keratinocytes. Front. Immunol. 2021, 11, 599947. [Google Scholar] [CrossRef]
- Roberson, E.D.O.; Liu, Y.; Ryan, C.; Joyce, C.E.; Duan, S.; Cao, L.; Martin, A.; Liao, W.; Menter, A.; Bowcock, A.M. A Subset of Methylated CpG Sites Differentiate Psoriatic from Normal Skin. J. Investig. Dermatol. 2012, 132, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhao, M.; Liang, G.; Yin, G.; Huang, D.; Su, F.; Zhai, H.; Wang, L.; Su, Y.; Lu, Q. Whole-Genome DNA Methylation in Skin Lesions from Patients with Psoriasis Vulgaris. J. Autoimmun. 2013, 41, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A.; Senapati, S.; Roy, S.; Chatterjee, G.; Chatterjee, R. Epigenome-Wide DNA Methylation Regulates Cardinal Pathological Features of Psoriasis. Clin. Epigenet. 2018, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Yin, G.; An, P.; Wang, C.; Liu, R.; Yang, Y.; Yan, X.; Li, J.; Li, X.; Zhang, K. DNA Methylation of Dermal MSCs in Psoriasis: Identification of Epigenetically Dysregulated Genes. J. Dermatol. Sci. 2013, 72, 103–109. [Google Scholar] [CrossRef]
- Zhou, F.; Wang, W.; Shen, C.; Li, H.; Zuo, X.; Zheng, X.; Yue, M.; Zhang, C.; Yu, L.; Chen, M.; et al. Epigenome-Wide Association Analysis Identified Nine Skin DNA Methylation Loci for Psoriasis. J. Investig. Dermatol. 2016, 136, 779–787. [Google Scholar] [CrossRef]
- Zhang, P.; Su, Y.; Chen, H.; Zhao, M.; Lu, Q. Abnormal DNA Methylation in Skin Lesions and PBMCs of Patients with Psoriasis Vulgaris. J. Dermatol. Sci. 2010, 60, 40–42. [Google Scholar] [CrossRef]
- Bai, J.; Liu, Z.; Xu, Z.; Ke, F.; Zhang, L.; Zhu, H.; Lou, F.; Wang, H.; Fei, Y.; Shi, Y.-L.; et al. Epigenetic Downregulation of SFRP4 Contributes to Epidermal Hyperplasia in Psoriasis. J. Immunol. 2015, 194, 4185–4198. [Google Scholar] [CrossRef]
- Tian, F.; Mauro, T.M.; Li, Z. The Pathological Role of Wnt5a in Psoriasis and Psoriatic Arthritis. J. Cell Mol. Med. 2019, 23, 5876–5883. [Google Scholar] [CrossRef] [PubMed]
- Ruchusatsawat, K.; Wongpiyabovorn, J.; Shuangshoti, S.; Hirankarn, N.; Mutirangura, A. SHP-1 Promoter 2 Methylation in Normal Epithelial Tissues and Demethylation in Psoriasis. J. Mol. Med. 2006, 84, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Ruchusatsawat, K.; Wongpiyabovorn, J.; Protjaroen, P.; Chaipipat, M.; Shuangshoti, S.; Thorner, P.S.; Mutirangura, A. Parakeratosis in Skin Is Associated with Loss of Inhibitor of Differentiation 4 via Promoter Methylation. Hum. Pathol. 2011, 42, 1878–1887. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, Z.-Q.; Cui, P.-G.; Yao, X.; Li, Y.-M.; Li, A.-S.; Gong, J.-Q.; Cao, Y.-H. The Methylation Pattern of P16 INK4a Gene Promoter in Psoriatic Epidermis and Its Clinical Significance. Br. J. Derm. 2008, 158, 987–993. [Google Scholar] [CrossRef]
- Sheng, Y.; Wen, L.; Zheng, X.; Li, M.; Wang, D.; Chen, S.; Li, R.; Tang, L.; Zhou, F. CYP2S1 Might Regulate Proliferation and Immune Response of Keratinocyte in Psoriasis. Epigenetics 2021, 16, 618–628. [Google Scholar] [CrossRef]
- Maeda, T.; Hanna, A.N.; Sim, A.B.; Chua, P.P.; Chong, M.T.; Tron, V.A. GADD45 Regulates G2/M Arrest, DNA Repair, and Cell Death in Keratinocytes Following Ultraviolet Exposure. J. Investig. Dermatol. 2002, 119, 22–26. [Google Scholar] [CrossRef]
- Rodríguez-Jiménez, P.; Fernández-Messina, L.; Ovejero-Benito, M.C.; Chicharro, P.; Vera-Tomé, P.; Vara, A.; Cibrian, D.; Martínez-Fleta, P.; Jiménez-Fernández, M.; Sánchez-García, I.; et al. Growth Arrest and DNA Damage-Inducible Proteins (GADD45) in Psoriasis. Sci. Rep. 2021, 11, 14579. [Google Scholar] [CrossRef]
- Liu, S.G.; Luo, G.P.; Qu, Y.B.; Chen, Y.F. Indirubin Inhibits Wnt/β-Catenin Signal Pathway via Promoter Demethylation of WIF-1. BMC Complement Med. 2020, 20, 250. [Google Scholar] [CrossRef]
- Cai, Y.; Fleming, C.; Yan, J. New Insights of T Cells in the Pathogenesis of Psoriasis. Cell Mol. Immunol. 2012, 9, 302–309. [Google Scholar] [CrossRef]
- Han, J.; Park, S.-G.; Bae, J.-B.; Choi, J.; Lyu, J.-M.; Park, S.H.; Kim, H.S.; Kim, Y.-J.; Kim, S.; Kim, T.-Y. The Characteristics of Genome-Wide DNA Methylation in Naïve CD4+ T Cells of Patients with Psoriasis or Atopic Dermatitis. Biochem. Biophys. Res. Commun. 2012, 422, 157–163. [Google Scholar] [CrossRef]
- Vecellio, M.; Paraboschi, E.M.; Ceribelli, A.; Isailovic, N.; Motta, F.; Cardamone, G.; Robusto, M.; Asselta, R.; Brescianini, S.; Sacrini, F.; et al. DNA Methylation Signature in Monozygotic Twins Discordant for Psoriatic Disease. Front. Cell Dev. Biol. 2021, 9, 778677. [Google Scholar] [CrossRef] [PubMed]
- Charras, A.; Garau, J.; Hofmann, S.R.; Carlsson, E.; Cereda, C.; Russ, S.; Abraham, S.; Hedrich, C.M. DNA Methylation Patterns in CD8+ T Cells Discern Psoriasis From Psoriatic Arthritis and Correlate With Cutaneous Disease Activity. Front. Cell Dev. Biol. 2021, 9, 746145. [Google Scholar] [CrossRef] [PubMed]
- Ngalamika, O.; Liang, G.; Zhao, M.; Yu, X.; Yang, Y.; Yin, H.; Liu, Y.; Yung, S.; Chan, T.M.; Lu, Q. Peripheral Whole Blood FOXP3 TSDR Methylation: A Potential Marker in Severity Assessment of Autoimmune Diseases and Chronic Infections. Immunol. Investig. 2015, 44, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, R.; Li, X.; Yin, G.; Niu, X. Promoter Methylation Status of P15 and P21 Genes in HPP-CFCs of Bone Marrow of Patients with Psoriasis. Eur. J. Dermatol. 2009, 19, 141–146. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, R.; Li, X.; Yin, G.; Niu, X.; Hou, R. The MRNA Expression and Promoter Methylation Status of the P16 Gene in Colony-Forming Cells with High Proliferative Potential in Patients with Psoriasis. Clin. Exp. Derm. 2007, 32, 702–708. [Google Scholar] [CrossRef]
- Li, X.; Zhao, X.; Xing, J.; Li, J.; He, F.; Hou, R.; Wang, Q.; Yin, G.; Li, X.; Zhang, K. Different Epigenome Regulation and Transcriptome Expression of CD4 + and CD8 + T Cells from Monozygotic Twins Discordant for Psoriasis. Australas. J. Derm. 2020, 61. [Google Scholar] [CrossRef]
- MacAlpine, D.M.; Almouzni, G. Chromatin and DNA Replication. Cold Spring Harb. Perspect. Biol. 2013, 5, a010207. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Li, H.; Yao, Q.; Mariscal, A.G.; Wu, X.; Hülse, J.; Pedersen, E.; Helin, K.; Waisman, A.; Vinkel, C.; Thomsen, S.F.; et al. Epigenetic Control of IL-23 Expression in Keratinocytes Is Important for Chronic Skin Inflammation. Nat. Commun. 2018, 9, 1420. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, L.; Ke, Y.; Lei, J.; Shen, S.; Shao, S.; Zhang, C.; Zhu, Z.; Dang, E.; Wang, G. EZH2-Dependent Epigenetic Modulation of Histone H3 Lysine-27 Contributes to Psoriasis by Promoting Keratinocyte Proliferation. Cell Death Dis. 2020, 11, 826. [Google Scholar] [CrossRef]
- Busch, F.; Mobasheri, A.; Shayan, P.; Stahlmann, R.; Shakibaei, M. Sirt-1 Is Required for the Inhibition of Apoptosis and Inflammatory Responses in Human Tenocytes. J. Biol. Chem. 2012, 287, 25770–25781. [Google Scholar] [CrossRef] [PubMed]
- McCormick, R.; McDonagh, B.; Goljanek-Whysall, K. MicroRNA-SIRT-1 Interactions: Key Regulators of Adult Skeletal Muscle Homeostasis?: Perspectives. J. Physiol. 2017, 595, 3253–3254. [Google Scholar] [CrossRef]
- Hwang, Y.-J.; Na, J.-I.; Byun, S.-Y.; Kwon, S.-H.; Yang, S.-H.; Lee, H.-S.; Choi, H.-R.; Cho, S.; Youn, S.W.; Park, K.-C. Histone Deacetylase 1 and Sirtuin 1 Expression in Psoriatic Skin: A Comparison between Guttate and Plaque Psoriasis. Life 2020, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Cao, G.; Sun, G.; Zhu, L.; Tian, Y.; Song, Y.; Guo, C.; Wang, X.; Zhong, J.; Zhou, W.; et al. GLS1-Mediated Glutaminolysis Unbridled by MALT1 Protease Promotes Psoriasis Pathogenesis. J. Clin. Investig. 2020, 130, 5180–5196. [Google Scholar] [CrossRef] [PubMed]
- Masalha, M.; Ben-Dov, I.Z.; Ram, O.; Meningher, T.; Jacob-Hirsch, J.; Kassem, R.; Sidi, Y.; Avni, D. H3K27Ac Modification and Gene Expression in Psoriasis. J. Dermatol. Sci. 2021, 103, 93–100. [Google Scholar] [CrossRef]
- Liao, Y.; Su, Y.; Wu, R.; Zhang, P.; Feng, C. Overexpression of Wilms Tumor 1 Promotes IL-1β Expression by Upregulating Histone Acetylation in Keratinocytes. Int. Immunopharmacol. 2021, 96, 107793. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, Y.; Zhang, H.; Zhang, Y.; Gao, L.; Tong, X.; Xie, Y.; Hu, Q.; Chen, C.; Ding, S.; et al. RPL22 Overexpression Promotes Psoriasis-Like Lesion by Inducing Keratinocytes Abnormal Biological Behavior. Front. Immunol. 2021, 12, 699900. [Google Scholar] [CrossRef]
- Ovejero-Benito, M.C.; Reolid, A.; Sánchez-Jiménez, P.; Saiz-Rodríguez, M.; Muñoz-Aceituno, E.; Llamas-Velasco, M.; Martín-Vilchez, S.; Cabaleiro, T.; Román, M.; Ochoa, D.; et al. Histone Modifications Associated with Biological Drug Response in Moderate-to-severe Psoriasis. Exp. Derm. 2018, 27, 1361–1371. [Google Scholar] [CrossRef]
- Liu, Z.; Cao, W.; Xu, L.; Chen, X.; Zhan, Y.; Yang, Q.; Liu, S.; Chen, P.; Jiang, Y.; Sun, X.; et al. The Histone H3 Lysine-27 Demethylase Jmjd3 Plays a Critical Role in Specific Regulation of Th17 Cell Differentiation. J. Mol. Cell Biol. 2015, 7, 505–516. [Google Scholar] [CrossRef]
- Zhang, P.; Su, Y.; Zhao, M.; Huang, W.; Lu, Q. Abnormal Histone Modifications in PBMCs from Patients with Psoriasis Vulgaris. Eur. J. Dermatol. 2011, 21, 552–557. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and Properties of a Novel Potential Biomarker for Cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-C.; Pan, H.-F.; Leng, R.-X.; Wang, D.-G.; Li, X.-P.; Li, X.-M.; Ye, D.-Q. Emerging Role of Long Noncoding RNAs in Autoimmune Diseases. Autoimmun. Rev. 2015, 14, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.-W.; Zhang, W.-J.; Wang, Y.; Tan, L.-P.; Bao, Y.-L.; Song, Z.-B.; Yu, C.-L.; Wang, S.-Y.; Liu, L.; Li, Y.-X. Convallatoxin Induces HaCaT Cell Necroptosis and Ameliorates Skin Lesions in Psoriasis-like Mouse Models. Biomed. Pharmacother. 2020, 121, 109615. [Google Scholar] [CrossRef] [PubMed]
- Chowdhari, S.; Sardana, K.; Saini, N. MiR-4516, a MicroRNA Downregulated in Psoriasis Inhibits Keratinocyte Motility by Targeting Fibronectin/Integrin A9 Signaling. Biochim. Et Biophys. Acta (BBA)—Mol. Basis Dis. 2017, 1863, 3142–3152. [Google Scholar] [CrossRef]
- Ichihara, A.; Jinnin, M.; Yamane, K.; Fujisawa, A.; Sakai, K.; Masuguchi, S.; Fukushima, S.; Maruo, K.; Ihn, H. MicroRNA-Mediated Keratinocyte Hyperproliferation in Psoriasis Vulgaris: MicroRNA-Mediated Keratinocyte Hyperproliferation in Psoriasis. Br. J. Dermatol. 2011, 165, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Qiao, M.; Li, R.H.; Zhao, X.T.; Wang, X.Y.; Sun, Q. Downregulation of MiR-145-5p Contributes to Hyperproliferation of Keratinocytes and Skin Inflammation in Psoriasis. Br. J. Dermatol. 2019, 180, 365–372. [Google Scholar] [CrossRef]
- Guinea-Viniegra, J.; Jiménez, M.; Schonthaler, H.B.; Navarro, R.; Delgado, Y.; José Concha-Garzón, M.; Tschachler, E.; Obad, S.; Daudén, E.; Wagner, E.F. Targeting MiR-21 to Treat Psoriasis. Sci. Transl. Med. 2014, 6. [Google Scholar] [CrossRef]
- Magenta, A.; D’Agostino, M.; Sileno, S.; Di Vito, L.; Uras, C.; Abeni, D.; Martino, F.; Barillà, F.; Madonna, S.; Albanesi, C.; et al. The Oxidative Stress-Induced MiR-200c Is Upregulated in Psoriasis and Correlates with Disease Severity and Determinants of Cardiovascular Risk. Oxidative Med. Cell. Longev. 2019, 2019, 8061901. [Google Scholar] [CrossRef]
- Huang, C.; Zhong, W.; Ren, X.; Huang, X.; Li, Z.; Chen, C.; Jiang, B.; Chen, Z.; Jian, X.; Yang, L.; et al. MiR-193b-3p–ERBB4 Axis Regulates Psoriasis Pathogenesis via Modulating Cellular Proliferation and Inflammatory-Mediator Production of Keratinocytes. Cell Death Dis. 2021, 12, 963. [Google Scholar] [CrossRef]
- Pan, M.; Huang, Y.; Zhu, X.; Lin, X.; Luo, D. MiR-125b-mediated Regulation of Cell Proliferation through the Jagged-1/Notch Signaling Pathway by Inhibiting BRD4 Expression in Psoriasis. Mol. Med. Rep. 2019. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Brodin, P.; Wei, T.; Meisgen, F.; Eidsmo, L.; Nagy, N.; Kemeny, L.; Ståhle, M.; Sonkoly, E.; Pivarcsi, A. MiR-125b, a MicroRNA Downregulated in Psoriasis, Modulates Keratinocyte Proliferation by Targeting FGFR2. J. Investig. Dermatol. 2011, 131, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; An, J.; Hua, Y.; Li, Z.; Yan, N.; Fan, W.; Su, C. MicroRNA-194 Regulates Keratinocyte Proliferation and Differentiation by Targeting Grainyhead-like 2 in Psoriasis. Pathol.—Res. Pract. 2017, 213, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Rongna, A.; Yu, P.; Hao, S.; Li, Y. MiR-876-5p Suppresses Cell Proliferation by Targeting Angiopoietin-1 in the Psoriasis. Biomed. Pharmacother. 2018, 103, 1163–1169. [Google Scholar] [CrossRef]
- Zheng, Y.; Cai, B.; Li, X.; Li, D.; Yin, G. MiR-125b-5p and MiR-181b-5p Inhibit Keratinocyte Proliferation in Skin by Targeting Akt3. Eur. J. Pharmacol. 2019, 862, 172659. [Google Scholar] [CrossRef]
- Jiang, M.; Sun, Z.; Dang, E.; Li, B.; Fang, H.; Li, J.; Gao, L.; Zhang, K.; Wang, G. TGFβ/SMAD/MicroRNA-486-3p Signaling Axis Mediates Keratin 17 Expression and Keratinocyte Hyperproliferation in Psoriasis. J. Investig. Dermatol. 2017, 137, 2177–2186. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, F.; Tian, Q.; Dong, J.; Chen, L.; Hu, R. Involvement of MiR-214-3p/FOXM1 Axis During the Progression of Psoriasis. Inflammation 2022, 45, 267–278. [Google Scholar] [CrossRef]
- Yan, S.; Xu, Z.; Lou, F.; Zhang, L.; Ke, F.; Bai, J.; Liu, Z.; Liu, J.; Wang, H.; Zhu, H.; et al. NF-ΚB-Induced MicroRNA-31 Promotes Epidermal Hyperplasia by Repressing Protein Phosphatase 6 in Psoriasis. Nat. Commun. 2015, 6, 7652. [Google Scholar] [CrossRef]
- Zhang, W.; Yi, X.; An, Y.; Guo, S.; Li, S.; Song, P.; Chang, Y.; Zhang, S.; Gao, T.; Wang, G.; et al. MicroRNA-17-92 Cluster Promotes the Proliferation and the Chemokine Production of Keratinocytes: Implication for the Pathogenesis of Psoriasis. Cell Death Dis. 2018, 9, 567. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Y.; Xia, Y.; Huo, J.; Zhang, Y.; Yang, P.; Zhang, Y.; Wang, X. Repression of MiR-142–3p Alleviates Psoriasis-like Inflammation by Repressing Proliferation and Promoting Apoptosis of Keratinocytes via Targeting Sema3A. Mol. Cell. Probes 2020, 52, 101573. [Google Scholar] [CrossRef]
- Xu, L.; Leng, H.; Shi, X.; Ji, J.; Fu, J.; Leng, H. MiR-155 Promotes Cell Proliferation and Inhibits Apoptosis by PTEN Signaling Pathway in the Psoriasis. Biomed. Pharmacother. 2017, 90, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, F.; Cao, H.; Yang, J. MiR-223 Regulates Proliferation and Apoptosis of IL-22-Stimulated HaCat Human Keratinocyte Cell Lines via the PTEN/Akt Pathway. Life Sci. 2019, 230, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Wei, T.; Janson, P.C.J.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel Regulators Involved in the Pathogenesis of Psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [PubMed]
- Zibert, J.R.; Løvendorf, M.B.; Litman, T.; Olsen, J.; Kaczkowski, B.; Skov, L. MicroRNAs and Potential Target Interactions in Psoriasis. J. Dermatol. Sci. 2010, 58, 177–185. [Google Scholar] [CrossRef]
- Fu, D.; Yu, W.; Li, M.; Wang, H.; Liu, D.; Song, X.; Li, Z.; Tian, Z. MicroRNA-138 Regulates the Balance of Th1/Th2 via Targeting RUNX3 in Psoriasis. Immunol. Lett. 2015, 166, 55–62. [Google Scholar] [CrossRef]
- Zheng, Y.-Z.; Chen, C.-F.; Jia, L.-Y.; Yu, T.-G.; Sun, J.; Wang, X.-Y. Correlation between MicroRNA-143 in Peripheral Blood Mononuclear Cells and Disease Severity in Patients with Psoriasis Vulgaris. Oncotarget 2017, 8, 51288–51295. [Google Scholar] [CrossRef]
- Collaborative Association Study of Psoriasis (CASP); Genetic Analysis of Psoriasis Consortium; Psoriasis Association Genetics Extension; Wellcome Trust Case Control Consortium 2; Tsoi, L.C.; Spain, S.L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; et al. Identification of 15 New Psoriasis Susceptibility Loci Highlights the Role of Innate Immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef]
- García-Rodríguez, S.; Arias-Santiago, S.; Blasco-Morente, G.; Orgaz-Molina, J.; Rosal-Vela, A.; Navarro, P.; Magro-Checa, C.; Martínez-López, A.; Ruiz, J.-C.; Raya, E.; et al. Increased Expression of MicroRNA-155 in Peripheral Blood Mononuclear Cells from Psoriasis Patients Is Related to Disease Activity. J. Eur. Acad. Derm. Venereol. 2017, 31, 312–322. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, L.; Liang, G.; Zhang, P.; Deng, X.; Tang, Q.; Zhai, H.; Chang, C.C.; Su, Y.; Lu, Q. Up-Regulation of MicroRNA-210 Induces Immune Dysfunction via Targeting FOXP3 in CD4+ T Cells of Psoriasis Vulgaris. Clin. Immunol. 2014, 150, 22–30. [Google Scholar] [CrossRef]
- Duan, Y.; Zou, J.; Mao, J.; Guo, D.; Wu, M.; Xu, N.; Zhou, J.; Zhang, Y.; Guo, W.; Jin, W. Plasma MiR-126 Expression Correlates with Risk and Severity of Psoriasis and Its High Level at Baseline Predicts Worse Response to Tripterygium Wilfordii Hook F in Combination with Acitretin. Biomed. Pharmacother. 2019, 115, 108761. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Chen, X.-Y.; Li, J.; Zhang, H.-Y.; Liu, J.; Sun, L.-D. MiR-200a Expression in CD4+ T Cells Correlates with the Expression of Th17/Treg Cells and Relevant Cytokines in Psoriasis Vulgaris: A Case Control Study. Biomed. Pharmacother. 2017, 93, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Borska, L.; Andrys, C.; Chmelarova, M.; Kovarikova, H.; Krejsek, J.; Hamakova, K.; Beranek, M.; Palicka, V.; Kremlacek, J.; Borsky, P.; et al. Roles of MiR-31 and Endothelin-1 in Psoriasis Vulgaris: Pathophysiological Functions and Potential Biomarkers. Physiol. Res. 2017, 66, 987–992. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, S.; Arias-Santiago, S.; Orgaz-Molina, J.; Magro-Checa, C.; Valenzuela, I.; Navarro, P.; Naranjo-Sintes, R.; Sancho, J.; Zubiaur, M. Abnormal Levels of Expression of Plasma MicroRNA-33 in Patients With Psoriasis. Actas Dermo-Sifiliográficas (Engl. Ed.) 2014, 105, 497–503. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, W.; Wei, C.; Wang, L.; Zhu, G.; Shi, Q.; Li, S.; Ge, R.; Li, K.; Gao, L.; et al. Serum and Skin Levels of MiR-369-3p in Patients with Psoriasis and Their Correlation with Disease Severity. Eur. J. Dermatol. 2013, 23, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.-Y.; Zhang, K.; Lu, W.-J.; Xu, G.-W.; Zhang, J.-F.; Tang, Z.-L. LncRNA MEG3 Influences the Proliferation and Apoptosis of Psoriasis Epidermal Cells by Targeting MiR-21/Caspase-8. BMC Mol. Cell Biol. 2019, 20, 46. [Google Scholar] [CrossRef]
- Qiao, M.; Li, R.; Zhao, X.; Yan, J.; Sun, Q. Up-Regulated LncRNA-MSX2P1 Promotes the Growth of IL-22-Stimulated Keratinocytes by Inhibiting MiR-6731-5p and Activating S100A7. Exp. Cell Res. 2018, 363, 243–254. [Google Scholar] [CrossRef]
- Abdallah, H.Y.; Tawfik, N.Z.; Soliman, N.H.; Eldeen, L.A.T. The LncRNA PRINS-MiRNA-MRNA Axis Gene Expression Profile as a Circulating Biomarker Panel in Psoriasis. Mol. Diagn. 2022. [Google Scholar] [CrossRef]
- Ahmed Shehata, W.; Maraee, A.; Abd El Monem Ellaithy, M.; Tayel, N.; Abo-Ghazala, A.; Mohammed El-Hefnawy, S. Circulating Long Noncoding RNA Growth Arrest-specific Transcript 5 as a Diagnostic Marker and Indicator of Degree of Severity in Plaque Psoriasis. Int. J. Derm. 2021, 60, 973–979. [Google Scholar] [CrossRef]
- Liu, X.; Frost, J.; Bowcock, A.; Zhang, W. Canonical and Interior Circular RNAs Function as Competing Endogenous RNAs in Psoriatic Skin. Int. J. Mol. Sci. 2021, 22, 5182. [Google Scholar] [CrossRef]
- Yazici, S.; Guner, R.; Akyol, M.; Tuzemen Bayyurt, E.; Arslan, S. Research on Hotair and 7SL-RNA Gene Expression Levels in Psoriasis Vulgaris. Indian J. Derm. 2021, 66, 704. [Google Scholar] [CrossRef]
- Moldovan, L.I.; Tsoi, L.C.; Ranjitha, U.; Hager, H.; Weidinger, S.; Gudjonsson, J.E.; Kjems, J.; Kristensen, L.S. Characterization of Circular RNA Transcriptomes in Psoriasis and Atopic Dermatitis Reveals Disease-specific Expression Profiles. Exp. Derm. 2021, 30, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, F.; Hua, M.; Guo, J.; Nong, Y.; Tang, Q.; Zhong, F.; Qin, L. Knockdown of LncRNA MIR31HG Inhibits Cell Proliferation in Human HaCaT Keratinocytes. Biol. Res. 2018, 51, 30. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, H.; Huang, L.; Kong, Y.; Chen, F.; Liang, J.; Yu, H.; Yao, Z. H19 LncRNA Regulates Keratinocyte Differentiation by Targeting MiR-130b-3p. Cell Death Dis. 2017, 8, e3174. [Google Scholar] [CrossRef] [PubMed]
- Szegedi, K.; Sonkoly, E.; Nagy, N.; Németh, I.B.; Bata-Csörgő, Z.; Kemény, L.; Dobozy, A.; Széll, M. The Anti-Apoptotic Protein G1P3 Is Overexpressed in Psoriasis and Regulated by the Non-Coding RNA, PRINS. Exp. Dermatol. 2010, 19, 269–278. [Google Scholar] [CrossRef]
- Cai, P.; Otten, A.B.C.; Cheng, B.; Ishii, M.A.; Zhang, W.; Huang, B.; Qu, K.; Sun, B.K. A Genome-Wide Long Noncoding RNA CRISPRi Screen Identifies PRANCR as a Novel Regulator of Epidermal Homeostasis. Genome Res. 2020, 30, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; George, T.L.; Otterson, G.A.; Verschraegen, C.; Wen, H.; Carbone, D.; Herman, J.; Bertino, E.M.; He, K. Advances in Epigenetic Therapeutics with Focus on Solid Tumors. Clin. Epigenet. 2021, 13, 83. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, J.-S.; Park, S.-Y.; Lee, Y.-J. Resveratrol Induces Human Keratinocyte Damage via the Activation of Class III Histone Deacetylase, Sirt1. Oncol. Rep. 2016, 35, 524–529. [Google Scholar] [CrossRef]
- Samuelov, L.; Bochner, R.; Magal, L.; Malovitski, K.; Sagiv, N.; Nousbeck, J.; Keren, A.; Fuchs-Telem, D.; Sarig, O.; Gilhar, A.; et al. Vorinostat, a Histone Deacetylase Inhibitor, as a Potential Novel Treatment for Psoriasis. Exp. Dermatol. 2022, 31, 567–576. [Google Scholar] [CrossRef]
- Jorn Bovenschen, H.; van de Kerkhof, P.C.; van Erp, P.E.; Woestenenk, R.; Joosten, I.; Koenen, H.J.P.M. Foxp3+ Regulatory T Cells of Psoriasis Patients Easily Differentiate into IL-17A-Producing Cells and Are Found in Lesional Skin. J. Investig. Dermatol. 2011, 131, 1853–1860. [Google Scholar] [CrossRef]
- Tao, R.; de Zoeten, E.F.; Özkaynak, E.; Chen, C.; Wang, L.; Porrett, P.M.; Li, B.; Turka, L.A.; Olson, E.N.; Greene, M.I.; et al. Deacetylase Inhibition Promotes the Generation and Function of Regulatory T Cells. Nat. Med. 2007, 13, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chowdhury, S.; Choudhary, V.; Chen, X.; Bollag, W.B. Keratinocyte Aquaporin-3 Expression Induced by Histone Deacetylase Inhibitors Is Mediated in Part by Peroxisome Proliferator-activated Receptors (PPARs). Exp. Derm. 2020, 29, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Bollag, W.B.; Aitkens, L.; White, J.; Hyndman, K.A. Aquaporin-3 in the Epidermis: More than Skin Deep. Am. J. Physiol.-Cell Physiol. 2020, 318, C1144–C1153. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Wu, R.; Zhang, S.; Kong, Y.; Liu, Z.; Wu, H.; Wang, H.; Su, Y.; Zhao, M.; Lu, Q. Topical Administration of Nanocarrier MiRNA-210 Antisense Ameliorates Imiquimod-induced Psoriasis-like Dermatitis in Mice. J. Dermatol. 2020, 47, 147–154. [Google Scholar] [CrossRef]
- Lifshiz Zimon, R.; Lerman, G.; Elharrar, E.; Meningher, T.; Barzilai, A.; Masalha, M.; Chintakunta, R.; Hollander, E.; Goldbart, R.; Traitel, T.; et al. Ultrasound Targeting of Q-Starch/MiR-197 Complexes for Topical Treatment of Psoriasis. J. Control. Release 2018, 284, 103–111. [Google Scholar] [CrossRef] [PubMed]
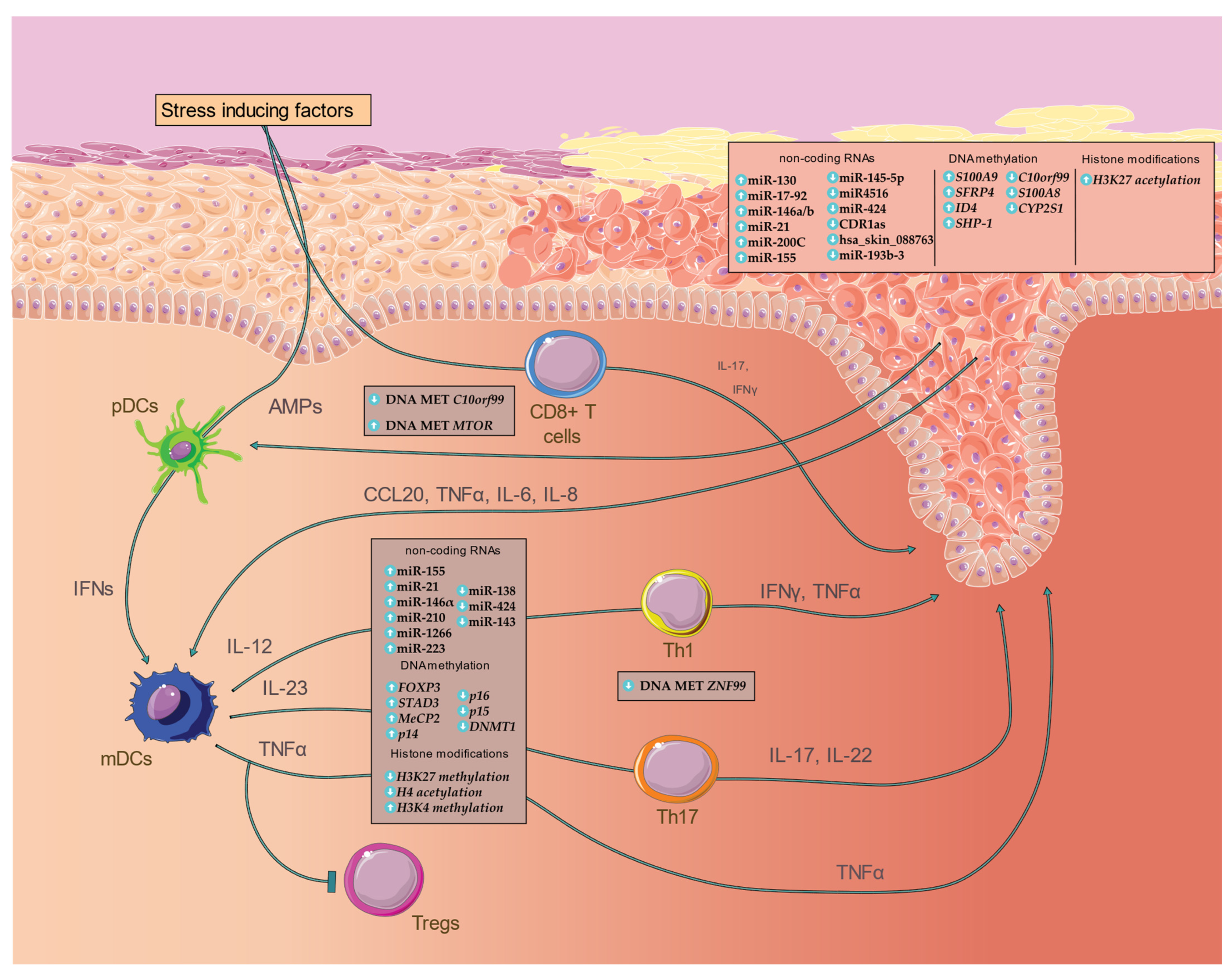

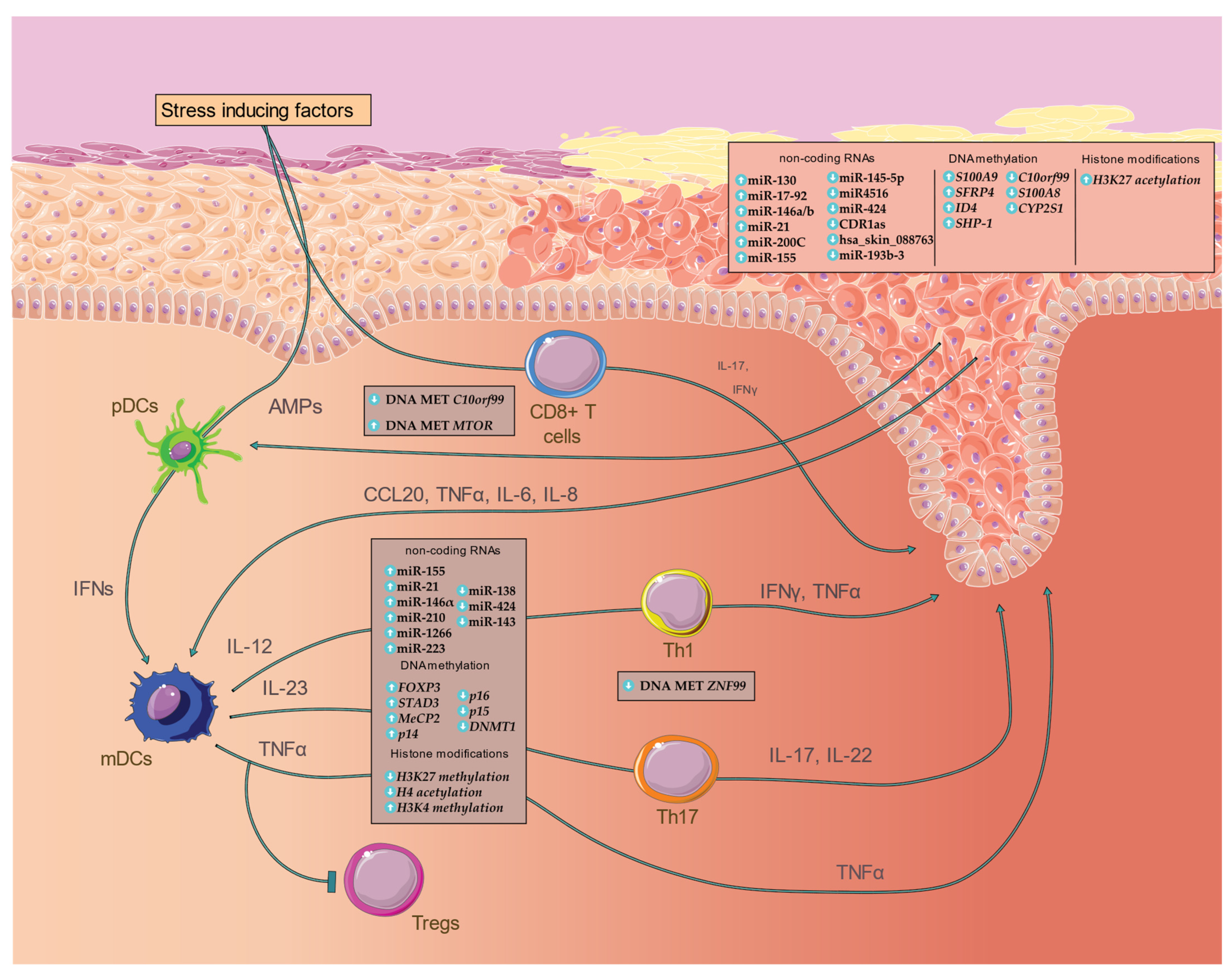{kind=link}
| Author, Year | ncRNA | Expression | Direct Targets (Indirectly Affected Genes) | Cell Type |
|---|---|---|---|---|
| Yan et al., 2018 [107] | miR-145-5p | Down | MLK3, (NF-κB, STAT3, AKT, GSK, MAPK, MTOR) | HEKs |
| Chowdhari et al., 2017 [105] | miR-4516 | Down | FN1, ITG9, STAT3 | HaCaT |
| Ichihara et al., 2011 [106] | miR-424 | Down | (MEK1, CCNE1) | HEKs |
| Huang et al., 2021 [110] | miR-193b-3 | Down | ERBB4, (STAT3, NF-κB, IL-6, CXCL1, CCL20, BD-2) | HaCaT |
| Pan et al., 2019 [111] | miR-125b | Down | BRD4, (JAG1) | HaCaT |
| Xu et al., 2011 [112] | miR-125b | Down | FGFR2 | HEKs |
| Yu et al., 2017 [113] | miR-194 | Down | GRHL2, SOX5 | HEKs |
| Rongna et al., 2018 [114] | miR-876-5p | Down | ANGPT1 | HaCaT |
| Zheng et al., 2019 [115] | miR-181b-5p | Down | AKT3 | HEKs |
| Zheng et al., 2019 [115] | miR-125b-5 | Down | AKT3 | HEKs |
| Jiang et al., 2017 [116] | miR-486-3p | Down | K17 | HaCaT |
| Zhao et al., 2022 [117] | miR-214-3p | Down | FOXM1 | HaCaT |
| Yan et al., 2015 [118] | miR-31 | Up | PPP6C | HEKs |
| Guinea-Viniegra et al., 2014 [108] | miR-21 | Up | TIMP3 | HEKs |
| Zhang et al., 2018 [119] | miR-17-92 | Up | CDKN2B, SOCS1 | HEKs |
| Zhang et al., 2020 [120] | miR-142-3p | Up | SEMA3A | HaCaT |
| Xu et al., 2017 [121] | miR-155 | Up | (PTEN, PIP3, AKT, BAX, Bcl-2) | HaCaT |
| Wang et al., 2019 [122] | miR-223 | Up | PTEN | HaCaT |
| Sonkoly et al., 2007 [123] | miR-203 | Up | SOCS3, (STAT3) | HEKs |
| Zibert et al., 2010 [124] | miR-221, miR-222 | Up | (TIMP3) | HEKs |
| Author, Year | ncRNA | Expression | Direct Targets (Indirectly Affected Genes) | Cell Type |
|---|---|---|---|---|
| Immune cells | ||||
| Zheng et al., 2017 [126] | miR-143 | Down | BCL2 | PBMCs |
| Fu et al., 2015 [125] | miR-138 | Down | RUNX3 | CD4+ T cells |
| Garcıa–Rodrıguez et al., 2016 [128] | miR-146a | Up | TRAF6, IRAK1, (NF-κΒ, IL-6, TNFα) | PBMCs |
| Garcıa–Rodrıguez et al., 2016 [128] | miR-21 | Up | PDCD4 | PBMCs |
| Garcıa–Rodrıguez et al., 2016 [128] | miR-155 | Up | SOCS1 | PBMCs |
| Serum levels | ||||
| Duan et al., 2019 [130] | miR-126 | Down | - | Plasma |
| Ichihara et al., 2011 [106] | miR-424 | Down | (MEK1, CCNE1) | Plasma |
| Zhao et al., 2014 [129] | miR-210 | Up | FOXP3, (IFN-γ, IL-17, IL-10, TGF-β) | Plasma |
| Magenta et al., 2019 [109] | miR-200c | Up | SIRT1, eNOS, FOXO1 | Plasma |
| Wang et al., 2017 [131] | miR-200a | Up | - | Plasma |
| Borska et al., 2017 [132] | miR-31 | Up | - | Plasma |
| García-Rodríguez et al., 2014 [133] | miR-33 | Up | ABCA1 | Plasma |
| Guo et al., 2013 [134] | miR-369-3p | Up | - | Plasma |
| Author, Year | Non-Coding RNA | Expression | Target Genes (Indirect Targets) | Cell Type |
|---|---|---|---|---|
| Liu et al., 2021 [139] | CDR1as | Down | (EGR3, GATA6, GATA3, FOXN3) | HEKs |
| Liu et al., 2021 [139] | hsa_skin_088763 | Down | (GATA6, SIK2, IL17RD, EGR3, FAS, LRIG1, PPARGC1A) | HEKs |
| Jia et al., 2019 [135] | MEG3 | Down | miR-21 | HaCaT |
| Liu et al., 2021 [139] | hsa_skin_05227 | Down | GATA6 | HEKs |
| Yazıcı et al., 2021 [140] | 7SL-RNA | Down | - | HEKs |
| Moldovan et al., 2020 [141] | circEXOC6B | Down | - | HEKs |
| Moldovan et al., 2020 [142] | circSLC8A | Down | - | HEKs |
| Moldovan et al., 2020 [142] | circRHOBTB | Down | - | HEKs |
| Qiao et al., 2018 [136] | MSX2P1 | Up | miR-6731, (S100A7) | HaCaT, NHEK |
| Gao et al., 2018 [142] | MIR31HG | Up | - | HaCaT |
| Li et al., 2017 [143] | lncRNA-H19 | Up | miR-130b, (DSG1) | HEKs |
| Szegedi et al., 2010 [144] | PRINS | Up | - | HeLa |
| Cai et al., 2019 [145] | PRANCR | Up | - | HEKs |
| Moldovan et al., 2020 [141] | cTNFRSF21, | Up | - | HEKs |
| Moldovan et al., 2020 [141] | circDOCK1 | Up | - | HEKs |
| Moldovan et al., 2020 [141] | circARAP2 | Up | - | HEKs |
| Moldovan et al., 2020 [141] | circDDX21 | Up | - | HEKs |
| Moldovan et al., 2020 [141] | circZRANB1 | Up | - | HEKs |
| Author, Year | Non-Coding RNA | Expression | Target Genes (Indirect Targets) | Cell Type |
|---|---|---|---|---|
| Abdallah et al., 2022 [137] | PRINS | Down | miR-124, miR-203a, miR-129, miR-146a, miR-9, GIP3 | Plasma |
| Shehata et al., 2021 [138] | GAS5 | Up | - | Plasma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonatos, C.; Grafanaki, K.; Asmenoudi, P.; Xiropotamos, P.; Nani, P.; Georgakilas, G.K.; Georgiou, S.; Vasilopoulos, Y. Contribution of the Environment, Epigenetic Mechanisms and Non-Coding RNAs in Psoriasis. Biomedicines 2022, 10, 1934. https://doi.org/10.3390/biomedicines10081934
Antonatos C, Grafanaki K, Asmenoudi P, Xiropotamos P, Nani P, Georgakilas GK, Georgiou S, Vasilopoulos Y. Contribution of the Environment, Epigenetic Mechanisms and Non-Coding RNAs in Psoriasis. Biomedicines. 2022; 10(8):1934. https://doi.org/10.3390/biomedicines10081934
Chicago/Turabian StyleAntonatos, Charalabos, Katerina Grafanaki, Paschalia Asmenoudi, Panagiotis Xiropotamos, Paraskevi Nani, Georgios K. Georgakilas, Sophia Georgiou, and Yiannis Vasilopoulos. 2022. "Contribution of the Environment, Epigenetic Mechanisms and Non-Coding RNAs in Psoriasis" Biomedicines 10, no. 8: 1934. https://doi.org/10.3390/biomedicines10081934
APA StyleAntonatos, C., Grafanaki, K., Asmenoudi, P., Xiropotamos, P., Nani, P., Georgakilas, G. K., Georgiou, S., & Vasilopoulos, Y. (2022). Contribution of the Environment, Epigenetic Mechanisms and Non-Coding RNAs in Psoriasis. Biomedicines, 10(8), 1934. https://doi.org/10.3390/biomedicines10081934