
Vitamin D-Dimer: A Possible Biomolecule Modulator in Cytotoxic and Phagocytosis Processes?

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Vitamin D Binding Protein (VDBP)
2.3. Labeling of VitD-dgVDBP
2.4. Macrophage Phagocytosis and Superoxide Anion Radical Production Analysis
2.5. Apoptosis Measurements on Malignant Prostate Cell Line in the Absence or Presence of dgVDBP or VitD-dgVDBP
2.6. Flow Cytometry of T Cells in the Absence or Presence of VitD-dgVDBP or Labeled VitD-dgVDBP
2.7. Statistical Analysis
3. Results
3.1. Macrophage Phagocytosis and Superoxide Anion Radical Production Analysis
3.2. Apoptosis Measurements on Malignant Prostate Cell Line in the Absence or Presence of dgVDBP or VitD-dgVDBP
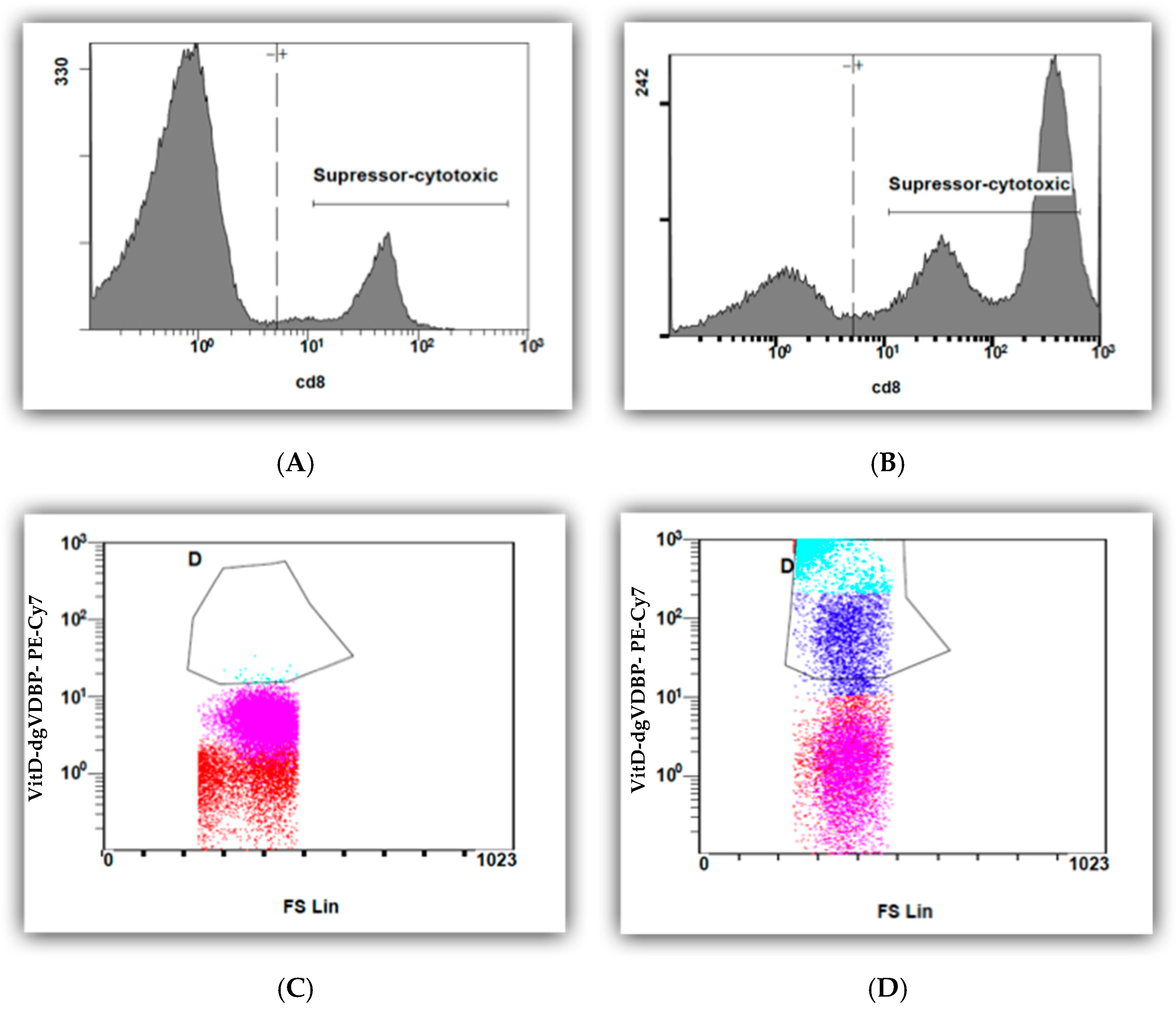
3.3. Flow Cytometry of T Cells in the Absence or Presence of Labeled VitD-dgVDBP
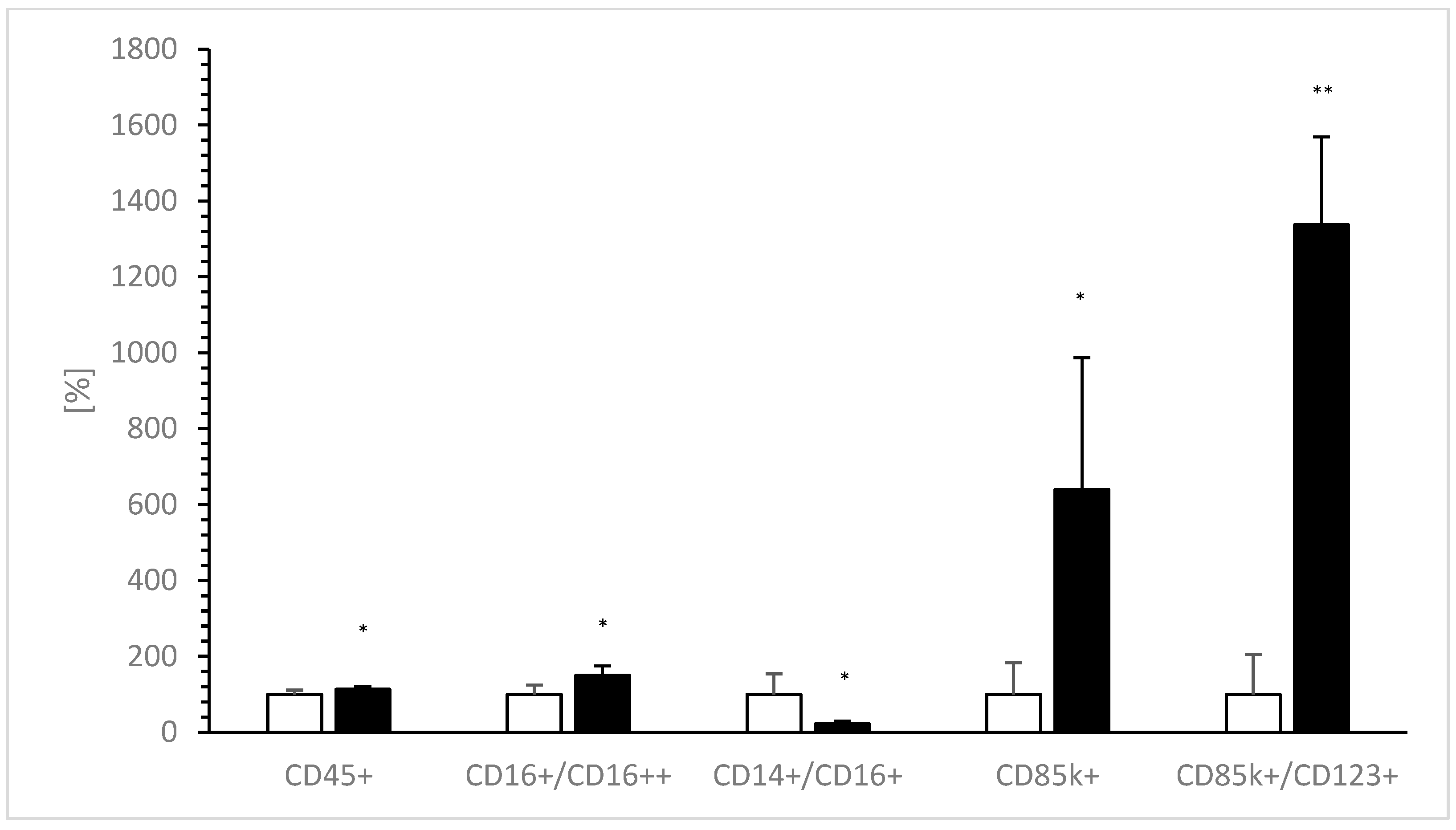
3.4. Flow Cytometry of nPBMCs in the Absence or Presence of VitD-dgVDBP
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef]
- Krutzik, S.R.; Hewison, M.; Liu, P.T.; Robles, J.A.; Stenger, S.; Adams, J.S.; Modlin, R.L. IL-15 links TLR2/1-induced macrophage differentiation to the vitamin D-dependent antimicrobial pathway. J. Immunol. 2008, 181, 7115–7120. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.J.; O’Garra, A. 1α,25-Dihydroxyvitamin D3 Has a Direct Effect on Naive CD4+ T Cells to Enhance the Development of Th2 Cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef]
- Hu, J.; Wan, Y. Tolerogenic dendritic cells and their potential applications. Immunology 2011, 132, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Pantalena, L.-C.; Liu, X.K.; Gaffen, S.L.; Liu, H.; Rohowsky-Kochan, C.; Ichiyama, K.; Yoshimura, A.; Steinman, L.; Christakos, S.; et al. 1,25-Dihydroxyvitamin D 3 Ameliorates Th17 Autoimmunity via Transcriptional Modulation of Interleukin-17A. Mol. Cell. Biol. 2011, 31, 3653–3669. [Google Scholar] [CrossRef] [PubMed]
- Korf, H.; Wenes, M.; Stijlemans, B.; Takiishi, T.; Robert, S.; Miani, M.; Eizirik, D.L.; Gysemans, C.; Mathieu, C. 1,25-Dihydroxyvitamin D3 curtails the inflammatory and T cell stimulatory capacity of macrophages through an IL-10-dependent mechanism. Immunobiology 2012, 217, 1292–1300. [Google Scholar] [CrossRef]
- Penna, G.; Adorini, L. 1α,25-Dihydroxyvitamin D3Inhibits Differentiation, Maturation, Activation, and Survival of Dendritic Cells Leading to Impaired Alloreactive T Cell Activation. J. Immunol. 2000, 164, 2405–2411. [Google Scholar] [CrossRef]
- Pichler, J.; Gerstmayr, M.; Szépfalusi, Z.; Urbanek, R.; Peterlik, M.; Willheim, M. 1α,25(OH)2D3 Inhibits Not Only Th1 But Also Th2 Differentiation in Human Cord Blood T Cells. Pediatr. Res. 2002, 52, 12–18. [Google Scholar] [CrossRef][Green Version]
- Otterbein, L.R.; Cosio, C.; Graceffa, P.; Dominguez, R. Crystal structures of the vitamin D-binding protein and its complex with actin: Structural basis of the actin-scavenger system. Proc. Natl. Acad. Sci. USA 2002, 99, 8003–8008. [Google Scholar] [CrossRef]
- Bouillon, R.; Schuit, F.; Antonio, L.; Rastinejad, F. Vitamin D Binding Protein: A Historic Overview. Front. Endocrinol. 2019, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Kongsbak, M.; von Essen, M.R.; Levring, T.B.; Schjerling, P.; Woetmann, A.; Ødum, N.; Bonefeld, C.M.; Geisler, C. Vitamin D-binding protein controls T cell responses to vitamin D. BMC Immunol. 2014, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.S.; Assar, S.; Harnpanich, D.; Bouillon, R.; Lambrechts, D.; Prentice, A.; Schoenmakers, I. 25(OH)D2 Half-Life Is Shorter Than 25(OH)D3 Half-Life and Is Influenced by DBP Concentration and Genotype. J. Clin. Endocrinol. Metab. 2014, 99, 3373–3381. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Pauwels, S. The vitamin D-binding protein. Vitam. D 2018. [Google Scholar] [CrossRef]
- White, P.; Cooke, N. The Multifunctional Properties and Characteristics of Vitamin D-binding Protein. Trends Endocrinol. Metab. 2000, 11, 320–327. [Google Scholar] [CrossRef]
- Chesney, R.W. Interactions of vitamin D and the proximal tubule. Pediatr. Nephrol. 2016, 31, 7–14. [Google Scholar] [CrossRef]
- Yamamoto, N.; Kumashiro, R. Conversion of vitamin D3 binding protein (group-specific component) to a macrophage activating factor by the stepwise action of beta-galactosidase of B cells and sialidase of T cells. J. Immunol. 1993, 151, 2794–2802. [Google Scholar]
- Kanda, S.; Mochizuki, Y.; Miyata, Y.; Kanetake, H.; Yamamoto, N. Effects of vitamin D(3)-binding protein-derived macrophage activating factor (GcMAF) on angiogenesis. J. Natl. Cancer Inst. 2002, 94, 1311–1319. [Google Scholar] [CrossRef]
- Yamamoto, N.; Naraparaju, V.; Orchard, P. Defective lymphocyte glycosidases in the macrophage activation cascade of juvenile osteopetrosis. Blood 1996, 88, 1473–1478. [Google Scholar] [CrossRef]
- Yamamoto, N.; Naraparaju, V.R. Vitamin D3-Binding Protein as a Precursor for Macrophage Activating Factor in the Inflammation-Primed Macrophage Activation Cascade in Rats. Cell. Immunol. 1996, 170, 161–167. [Google Scholar] [CrossRef]
- Greilberger, J.; Greilberger, M.; Petek, T.; Philipp, S.; Bettina, L.; Reichl, H.; Kamel, A.; Herwig, R. Effective Increase of Serum Vitamin D3 by IV Application of a Cholecalciferol-N-Acetyl-Galactosamine-Stabilized Dimer: A Clinical Murine Trial Study. Clin. Lab. 2019, 65, 181114. [Google Scholar] [CrossRef] [PubMed]
- Rheinländer, A.; Schraven, B.; Bommhardt, U. CD45 in human physiology and clinical medicine. Immunol. Lett. 2018, 196, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Greilberger, J.; Herwig, R. Vitamin D-Deglycosylated Vitamin D Binding Protein Dimer: Positive Synergistic Effects on Recognition, Activation, Phagocytosis and Oxidative Stress on Macrophages. Clin. Lab. 2020, 66, 191121. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.B. Vitamin D, cancer, and dysregulated phosphate metabolism. Endocrine 2019, 65, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Cannell, J.; Grant, W.B. What is the role of vitamin D in autism? Derm. -Endocrinol. 2013, 5, 199–204. [Google Scholar] [CrossRef]
- Garcion, E.; Wion-Barbot, N.; Montero-Menei, C.N.; Berger, F.; Wion, D. New clues about vitamin D functions in the nervous system. Trends Endocrinol. Metab. 2002, 13, 100–105. [Google Scholar] [CrossRef]
- Ginanjar, E.; Setiati, S.S.; Setiyohadi, B. Vitamin D and autoimmune disease. Acta Med. Indones 2007, 39, 133–141. [Google Scholar]
- Lin, L.; Zhang, L.; Li, C.; Gai, Z.; Li, Y. Vitamin D and Vitamin D Receptor: New Insights in the Treatment of Hypertension. Curr. Protein Pept. Sci. 2019, 20, 984–995. [Google Scholar] [CrossRef]
- Molina, P.; Carrero, J.J.; Bover, J.; Chauveau, P.; Mazzaferro, S.; Torres, P.U.; European Renal Nutrition (ERN) and Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD) Working Groups of the European Renal Association–European Dialysis Transplant Association (ERA-EDTA). Vitamin D, a modulator of musculoskeletal health in chronic kidney disease. J. Cachexia Sarcopenia Muscle 2017, 8, 686–701. [Google Scholar] [CrossRef]
- Norman, A.W.; Ishizuka, S.; Okamura, W.H. Ligands for the vitamin D endocrine system: Different shapes function as agonists and antagonists for genomic and rapid response receptors or as a ligand for the plasma vitamin D binding protein. J. Steroid Biochem. Mol. Biol. 2001, 76, 49–59. [Google Scholar] [CrossRef]
- Bouillon, R.; VAN Baelen, H.; Rombauts, W.; DE Moor, P. The Purification and Characterisation of the Human-Serum Binding Protein for the 25-Hydroxycholecalciferol (Transcalciferin) Identity with Group-Specific Component. JBIC J. Biol. Inorg. Chem. 1976, 66, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Carvalho, A.; Meraz-Ríos, M.A.; Santos-Argumedo, L.; Bajaña, S.; Soldevila, G.; Moreno-García, M.E.; Sánchez-Torres, C. CD16+ human monocyte-derived dendritic cells matured with different and unrelated stimuli promote similar allogeneic Th2 responses: Regulation by pro- and anti-inflammatory cytokines. Int. Immunol. 2004, 16, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Torres, C.; García-Romo, G.S.; Cornejo-Cortés, M.A.; Rivas-Carvalho, A.; Sanchez-Schmitz, G. CD16+ and CD16− human blood monocyte subsets differentiate in vitro to dendritic cells with different abilities to stimulate CD4+ T cells. Int. Immunol. 2001, 13, 1571–1581. [Google Scholar] [CrossRef]
- Randolph, G.J.; Sanchez-Schmitz, G.; Liebman, R.M.; Schäkel, K. The CD16+ (FcγRIII+) Subset of Human Monocytes Preferentially Becomes Migratory Dendritic Cells in a Model Tissue Setting. J. Exp. Med. 2002, 196, 517–527. [Google Scholar] [CrossRef]
- Kabanov, D.S.; Vwedenskaya, O.Y.; Fokina, M.A.; Morozova, E.M.; Grachev, S.; Prokhorenko, I.R. Impact of CD14 on Reactive Oxygen Species Production from Human Leukocytes Primed byEscherichia coliLipopolysaccharides. Oxidative Med. Cell. Longev. 2019, 2019, 6043245. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.R. The CD45 tyrosine phosphatase: A positive and negative regulator of immune cell function. Semin. Immunol. 2000, 12, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Schuette, V.; Embgenbroich, M.; Ulas, T.; Welz, M.; Schulte-Schrepping, J.; Draffehn, A.M.; Quast, T.; Koch, K.; Nehring, M.; König, J.; et al. Mannose receptor induces T-cell tolerance via inhibition of CD45 and up-regulation of CTLA-4. Proc. Natl. Acad. Sci. USA 2016, 113, 10649–10654. [Google Scholar] [CrossRef]
- Siftar, Z. IFCC Professional Scientific Exchange Programme Expression of CD85, a Killer-Cell Inhibitory Receptor (KIR) Molecule on T Cells in B-Chronic Lymphocytic Leukemia (B-CLL). EJIFCC 2000, 12, 117–119. [Google Scholar]
- MacDonald, K.; Munster, D.J.; Clark, G.J.; Dzionek, A.; Schmitz, J.; Hart, D.N.J. Characterization of human blood dendritic cell subsets. Blood 2002, 100, 4512–4520. [Google Scholar] [CrossRef]
- Rovati, B.; Mariucci, S.; Manzoni, M.; Bencardino, K.; Danova, M. Flow cytometric detection of circulating dendritic cells in healthy subjects. Eur. J. Histochem. 2008, 52, 45–52. [Google Scholar] [CrossRef]
- Gregory, K.J.; Zhao, B.; Bielenberg, D.R.; Dridi, S.; Wu, J.; Jiang, W.; Huang, B.; Pirie-Shepherd, S.; Fannon, M. Vitamin D Binding Protein-Macrophage Activating Factor Directly Inhibits Proliferation, Migration, and uPAR Expression of Prostate Cancer Cells. PLoS ONE 2010, 5, e13428. [Google Scholar] [CrossRef] [PubMed]
- Thyer, L.; Ward, E.; Smith, R.; Fiore, M.G.; Magherini, S.; Branca, J.J.V.; Morucci, G.; Gulisano, M.; Ruggiero, M.; Pacini, S. A Novel Role for a Major Component of the Vitamin D Axis: Vitamin D Binding Protein-Derived Macrophage Activating Factor Induces Human Breast Cancer Cell Apoptosis through Stimulation of Macrophages. Nutrients 2013, 5, 2577–2589. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD3 (T Cells) % | CD4 (TH) % | CD8 (TC) % | CD19 (B Cells) % | CD14 (Monocyte) % | CD56/CD16 (NK Cells) % | PROPIDIUM IODIDE % Viability | HLA Type (ABC Type) | HLA-DR% (CLASS II) |
|---|---|---|---|---|---|---|---|---|
| 56.4 | 43.2 | 14.6 | 9.3 | 14.6 | 1.7 | 87 | HLA-A2 NEG, B27+ | 24.7 |
| 0 h Incubation | 12 h Incubation with VitD-dgVDBP | ||||
|---|---|---|---|---|---|
| Mean (% Lymph) | SD (% Lymph) | Mean (% Lymph) | SD (% Lymph) | p | |
| CD3+ | 61.9 | 5.7 | 68.5 | 4.7 | n.s. |
| CD4+ | 55.7 | 3.7 | 60.8 | 3.3 | n.s. |
| CD8+ | 15.6 | 8.2 | 13.6 | 4.6 | n.s. |
| CD4/CD8 | 2.1 | 1.1 | 2.0 | 1.2 | n.s. |
| CD4/CD8++ | 0.5 | 0.3 | 0.6 | 0.3 | n.s. |
| CD45+ | 84.7 | 9.5 | 96.0 | 6.0 | p < 0.05 |
| CD16+ | 10.4 | 4.0 | 8.4 | 4.5 | n.s. |
| CD16++ | 4.5 | 2.9 | 6.2 | 3.4 | n.s. |
| CD16+/CD16++ | 0.4 | 0.1 | 0.6 | 0.1 | p < 0.05 |
| CD14+ | 18.2 | 6.0 | 17.6 | 6.1 | n.s. |
| CD14+/CD16+ | 3.1 | 1.7 | 0.7 | 0.2 | p < 0.05 |
| CD14+CD16+ | 14.6 | 4.7 | 20.5 | 6.7 | n.s. |
| CD85k+ | 3.8 | 3.2 | 24.3 | 13.2 | p < 0.05 |
| CD123+ | 52.4 | 11.2 | 61.9 | 7.3 | n.s. |
| CD85k+/CD123+ | 3.5 | 3.7 | 46.8 | 8.1 | p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herwig, R.; Erlbacher, K.; Ibrahimagic, A.; Kacar, M.; Brajshori, N.; Beqiri, P.; Greilberger, J. Vitamin D-Dimer: A Possible Biomolecule Modulator in Cytotoxic and Phagocytosis Processes? Biomedicines 2022, 10, 1785. https://doi.org/10.3390/biomedicines10081785
Herwig R, Erlbacher K, Ibrahimagic A, Kacar M, Brajshori N, Beqiri P, Greilberger J. Vitamin D-Dimer: A Possible Biomolecule Modulator in Cytotoxic and Phagocytosis Processes? Biomedicines. 2022; 10(8):1785. https://doi.org/10.3390/biomedicines10081785
Chicago/Turabian StyleHerwig, Ralf, Katharina Erlbacher, Amela Ibrahimagic, Mehtap Kacar, Naime Brajshori, Petrit Beqiri, and Joachim Greilberger. 2022. "Vitamin D-Dimer: A Possible Biomolecule Modulator in Cytotoxic and Phagocytosis Processes?" Biomedicines 10, no. 8: 1785. https://doi.org/10.3390/biomedicines10081785
APA StyleHerwig, R., Erlbacher, K., Ibrahimagic, A., Kacar, M., Brajshori, N., Beqiri, P., & Greilberger, J. (2022). Vitamin D-Dimer: A Possible Biomolecule Modulator in Cytotoxic and Phagocytosis Processes? Biomedicines, 10(8), 1785. https://doi.org/10.3390/biomedicines10081785