
Estradiol and Estrogen-like Alternative Therapies in Use: The Importance of the Selective and Non-Classical Actions

 ,
, 
Abstract
1. Introduction
2. Estrogen Synthesis and Receptors
2.1. Forms and Synthesis of Estradiol
2.2. Estrogen Receptors and Their Localization
3. Mechanism of Action
3.1. Classical Estrogen Actions
3.2. Non-Classical Estrogen Actions
4. The Problem with E2 and Hormone Replacement Therapy (HRT)
5. Possible New Therapies: SERMs, Phytoestrogens, SERDs, and ANGELS
5.1. Selective Estrogen-Receptor Modulators (SERMs)
5.2. Phytoestrogens
5.3. Selective Estrogen-Receptor Downregulators (SERDs)
5.4. Activators of Non-Genomic Estrogen-like Signaling (ANGELS)
5.5. Aromatase Inhibitors
5.6. Future Direction of HRT
6. Estrogen Effects with Therapeutic Consequences
6.1. Main Role of E2 on Reproduction; Contraceptives
6.2. Menopausal Symptoms
6.2.1. Effects in the Brain
6.2.1.1. Hot Flushes
6.2.1.2. Mood Swings
6.2.1.3. Neuroprotection
6.2.2. Changes at the Periphery
6.2.2.1. Cardiovascular Effects
6.2.2.2. Metabolic Disturbances
6.2.2.3. Atrophy of the Outer Barrier (Skin, Mucosa) and Hair
6.2.3. Musculoskeletal System
6.2.3.1. Osteoporosis: Role of E2 in Bone Integrity
6.3. Role of E2 in Cancer
6.3.1. Breast Cancer
6.3.2. Ovarian Cancer
6.3.3. Endometrial Cancer
6.3.4. Prostate Cancer
6.4. General Health Benefit: Antiviral Effect
7. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veler, C.D.; Thayer, S.; Doisy, E.A. The Preparation of the Crystalline Follicular Ovarian Hormone: Theelin. J. Biol. Chem. 1930, 87, 357–371. [Google Scholar] [CrossRef]
- Huffman, M.N.; Thayer, S.A.; Doisy, E.A. The isolation of α-dihydrotheelin from human placenta. J. Biol. Chem. 1940, 133, 567–571. [Google Scholar] [CrossRef]
- Butenandt, A. Uber Progynon ein krytallisiertes weibliches Sexualhormone. Naturwissenschaften 1929, 17, 879. [Google Scholar] [CrossRef]
- Tata, J.R. One hundred years of hormones. EMBO Rep. 2005, 6, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S.; Wagner, J.S.; Roberts, R.M.; Lubahn, D.B. Intraovarian actions of oestrogen. Reproduction 2001, 122, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Munk, A.J.L.; Zoeller, A.C.; Hennig, J. Fluctuations of estradiol during women’s menstrual cycle: Influences on reactivity towards erotic stimuli in the late positive potential. Psychoneuroendocrinology 2018, 91, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Micevych, P.E.; Mermelstein, P.G.; Sinchak, K. Estradiol Membrane-Initiated Signaling in the Brain Mediates Reproduction. Trends Neurosci. 2017, 40, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.D. Menopause. Lancet 2008, 371, 760–770. [Google Scholar] [CrossRef]
- Bosworth, H.B.; Bastian, L.A.; Kuchibhatla, M.N.; Steffens, D.C.; McBride, C.M.; Skinner, C.S.; Rimer, B.K.; Siegler, I.C. Depressive symptoms, menopausal status, and climacteric symptoms in women at midlife. Psychosom. Med. 2001, 63, 603–608. [Google Scholar] [CrossRef]
- Weise, M.; De-Levi, S.; Barnes, K.M.; Gafni, R.I.; Abad, V.; Baron, J. Effects of estrogen on growth plate senescence and epiphyseal fusion. Proc. Natl. Acad. Sci. USA 2001, 98, 6871–6876. [Google Scholar] [CrossRef] [PubMed]
- Klein-Nulend, J.; van Oers, R.F.; Bakker, A.D.; Bacabac, R.G. Bone cell mechanosensitivity, estrogen deficiency, and osteoporosis. J. Biomech. 2015, 48, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Levin, V.A.; Jiang, X.; Kagan, R. Estrogen therapy for osteoporosis in the modern era. Osteoporos. Int. 2018, 29, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.J.; Hewitt, S.C.; Arao, Y.; Korach, K.S. Estrogen Hormone Biology. Curr. Top. Dev. Biol. 2017, 125, 109–146. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.A.; Korach, K.S. Estrogen receptors and human disease: An update. Arch. Toxicol. 2012, 86, 1491–1504. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Z.; Liu, K.; Liu, J.; Chai, S.; Chen, G.; Wen, S.; Ming, T.; Wang, J.; Ma, Y.; et al. Activation of the G Protein-Coupled Estrogen Receptor Prevented the Development of Acute Colitis by Protecting the Crypt Cell. J. Pharmacol. Exp. Ther. 2021, 376, 281–293. [Google Scholar] [CrossRef]
- Callard, G.V.; Tarrant, A.M.; Novillo, A.; Yacci, P.; Ciaccia, L.; Vajda, S.; Chuang, G.-Y.; Kozakov, D.; Greytak, S.R.; Sawyer, S.; et al. Evolutionary origins of the estrogen signaling system: Insights from amphioxus. J. Steroid Biochem. Mol. Biol. 2011, 127, 176–188. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5757, Estradiol. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Estradiol (accessed on 10 January 2022).
- Li, R.; Cui, J.; Shen, Y. Brain sex matters: Estrogen in cognition and Alzheimer’s disease. Mol. Cell. Endocrinol. 2014, 389, 13–21. [Google Scholar] [CrossRef]
- Hanukoglu, I. Steroidogenic enzymes: Structure, function, and role in regulation of steroid hormone biosynthesis. J. Steroid Biochem. Mol. Biol. 1992, 43, 779–804. [Google Scholar] [CrossRef]
- Hall, J.E.; Hall, M.E. Guyton and Hall: Textbook of Medical Physiology; Elsevier: Philadelphia, PA, USA, 2016. [Google Scholar]
- Sato, T.; Miyagawa, S.; Iguchi, T. Subchapter 94G—Estradiol-17β. In Handbook of Hormones; Takei, Y., Ando, H., Tsutsui, K., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 520–524. ISBN 978-0-12-801028-0. [Google Scholar]
- Thomas, M.P.; Potter, B.V.L. The structural biology of oestrogen metabolism. J. Steroid Biochem. Mol. Biol. 2013, 137, 27–49. [Google Scholar] [CrossRef]
- Beato, M.; Klug, J. Steroid hormone receptors: An update. Hum. Reprod. Update 2000, 6, 225–236. [Google Scholar] [CrossRef]
- Atanassova, N. Hydrohysteroid Dehydrogenases—Biological Role and Clinical Importance—Review; Canuto, Y.K.E.-R.A., Ed.; IntechOpen: Rijeka, Croatia, 2012; p. 6. [Google Scholar]
- Reed, B.G.; Carr, B.R. The Normal Menstrual Cycle and the Control of Ovulation; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; EndoText: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Gebhart, V.M.; Caldwell, J.D.; Rodewald, A.; Kalyvianaki, K.; Kampa, M.; Jirikowski, G.F. Estrogen receptors and sex hormone binding globulin in neuronal cells and tissue. Steroids 2019, 142, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yates, S.R. Dissolved organic matter and estrogen interactions regulate estrogen removal in the aqueous environment: A review. Sci. Total Environ. 2018, 640–641, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.V. Basic guides to the mechanism of estrogen action. Recent Prog. Horm. Res. 1962, 18, 387–414. [Google Scholar]
- Green, S.; Walter, P.; Kumar, V.; Krust, A.; Bornert, J.M.; Argos, P.; Chambon, P. Human oestrogen receptor cDNA: Sequence, expression and homology to v-erb-A. Nature 1986, 320, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Greene, G.L.; Gilna, P.; Waterfield, M.; Baker, A.; Hort, Y.; Shine, J. Sequence and expression of human estrogen receptor complementary DNA. Science 1986, 231, 1150–1154. [Google Scholar] [CrossRef]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef]
- Mosselman, S.; Polman, J.; Dijkema, R. ER beta: Identification and characterization of a novel human estrogen receptor. FEBS Lett. 1996, 392, 49–53. [Google Scholar] [CrossRef]
- Gustafsson, J.A. Estrogen receptor beta--a new dimension in estrogen mechanism of action. J. Endocrinol. 1999, 163, 379–383. [Google Scholar] [CrossRef]
- Evans, R.M. The steroid and thyroid hormone receptor superfamily. Science 1988, 240, 889–895. [Google Scholar] [CrossRef]
- Tsai, M.J.; O’Malley, B.W. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu. Rev. Biochem. 1994, 63, 451–486. [Google Scholar] [CrossRef]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Ström, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, G.B.; Tremblay, A.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Labrie, F.; Giguere, V. Cloning, chromosomal localization, and functional analysis of the murine estrogen receptor beta. Mol. Endocrinol. 1997, 11, 353–365. [Google Scholar] [CrossRef]
- Ruff, M.; Gangloff, M.; Wurtz, J.M.; Moras, D. Estrogen receptor transcription and transactivation: Structure-function relationship in DNA- and ligand-binding domains of estrogen receptors. Breast Cancer Res. 2000, 2, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Weiser, M.J.; Foradori, C.D.; Handa, R.J. Estrogen receptor beta in the brain: From form to function. Brain Res. Rev. 2008, 57, 309–320. [Google Scholar] [CrossRef]
- Shughrue, P.J.; Lane, M.V.; Merchenthaler, I. Comparative distribution of estrogen receptor-α and-β mRNA in the rat central nervous system. J. Comp. Neurol. 1997, 388, 507–525. [Google Scholar] [CrossRef]
- Saimi, M.; Moriya, S.; Li, Z.-L.; Miyaso, H.; Nagahori, K.; Kawata, S.; Omotehara, T.; Ogawa, Y.; Hino, H.; Miyazawa, K.; et al. Cytonuclear Estrogen Receptor Alpha Regulates Proliferation and Migration of Endometrial Carcinoma Cells. Tokai J. Exp. Clin. Med. 2021, 46, 7–16. [Google Scholar]
- Rosenfeld, C.S.; Cooke, P.S. Endocrine disruption through membrane estrogen receptors and novel pathways leading to rapid toxicological and epigenetic effects. J. Steroid Biochem. Mol. Biol. 2019, 187, 106–117. [Google Scholar] [CrossRef]
- Levin, E.R. Membrane estrogen receptors signal to determine transcription factor function. Steroids 2018, 132, 1–4. [Google Scholar] [CrossRef]
- Christensen, A.; Micevych, P. CAV1 siRNA reduces membrane estrogen receptor-α levels and attenuates sexual receptivity. Endocrinology 2012, 153, 3872–3877. [Google Scholar] [CrossRef] [PubMed]
- Boulware, M.I.; Kordasiewicz, H.; Mermelstein, P.G. Caveolin proteins are essential for distinct effects of membrane estrogen receptors in neurons. J. Neurosci. 2007, 27, 9941–9950. [Google Scholar] [CrossRef] [PubMed]
- Sołtysik, K.; Czekaj, P. ERα36—Another piece of the estrogen puzzle. Eur. J. Cell Biol. 2015, 94, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Carmeci, C.; Thompson, D.A.; Ring, H.Z.; Francke, U.; Weigel, R.J. Identification of a gene (GPR30) with homology to the G-protein-coupled receptor superfamily associated with estrogen receptor expression in breast cancer. Genomics 1997, 45, 607–617. [Google Scholar] [CrossRef]
- Filardo, E.J.; Thomas, P. GPR30: A seven-transmembrane-spanning estrogen receptor that triggers EGF release. Trends Endocrinol. Metab. 2005, 16, 362–367. [Google Scholar] [CrossRef]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R., Jr. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef]
- Filardo, E.; Quinn, J.; Pang, Y.; Graeber, C.; Shaw, S.; Dong, J.; Thomas, P. Activation of the novel estrogen receptor G protein-coupled receptor 30 (GPR30) at the plasma membrane. Endocrinology 2007, 148, 3236–3245. [Google Scholar] [CrossRef]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef]
- Filardo, E.J.; Thomas, P. Minireview: G Protein-Coupled Estrogen Receptor-1, GPER-1: Its Mechanism of Action and Role in Female Reproductive Cancer, Renal and Vascular Physiology. Endocrinology 2012, 153, 2953–2962. [Google Scholar] [CrossRef]
- Shoval, G.; Shbiro, L.; Hershkovitz, L.; Hazut, N.; Zalsman, G.; Mechoulam, R.; Weller, A. Prohedonic effect of cannabidiol in a rat model of depression. Neuropsychobiology 2016, 73, 123–129. [Google Scholar] [CrossRef]
- Sarchielli, E.; Guarnieri, G.; Idrizaj, E.; Squecco, R.; Mello, T.; Comeglio, P.; Gallina, P.; Maggi, M.; Vannelli, G.B.; Morelli, A. The G protein-coupled oestrogen receptor, GPER1, mediates direct anti-inflammatory effects of oestrogens in human cholinergic neurones from the nucleus basalis of Meynert. J. Neuroendocrinol. 2020, 32, e12837. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, B.W.; Tsai, M.-J. Molecular pathways of steroid receptor action. Biol. Reprod. 1992, 46, 163–167. [Google Scholar] [CrossRef]
- Klinge, C.M. Estrogen receptor interaction with co-activators and co-repressors. Steroids 2000, 65, 227–251. [Google Scholar] [CrossRef]
- Nilsson, S.; Makela, S.; Treuter, E.; Tujague, M.; Thomsen, J.; Andersson, G.; Enmark, E.; Pettersson, K.; Warner, M.; Gustafsson, J.-Å. Mechanisms of estrogen action. Physiol. Rev. 2001, 81, 1535–1565. [Google Scholar] [CrossRef]
- Beliakoff, J.; Whitesell, L. Hsp90: An emerging target for breast cancer therapy. Anticancer Drugs 2004, 15, 651–662. [Google Scholar] [CrossRef]
- Chakravarti, D.; LaMorte, V.J.; Nelson, M.C.; Nakajima, T.; Schulman, I.G.; Juguilon, H.; Montminy, M.; Evans, R.M. Role of CBP/P300 in nuclear receptor signalling. Nature 1996, 383, 99. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.J.; Tschugguel, W.; Schneeberger, C.; Huber, J.C. Production and actions of estrogens. N. Engl. J. Med. 2002, 346, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Le Dily, F.; Beato, M. Signaling by Steroid Hormones in the 3D Nuclear Space. Int. J. Mol. Sci. 2018, 19, 306. [Google Scholar] [CrossRef]
- O’lone, R.; Frith, M.C.; Karlsson, E.K.; Hansen, U. Genomic targets of nuclear estrogen receptors. Mol. Endocrinol. 2004, 18, 1859–1875. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, G.; Yasar, P.; Olgun, C.E.; Karakaya, B.; Kars, G.; Razizadeh, N.; Yavuz, K.; Turan, G.; Muyan, M. Dynamic transcriptional events mediated by estrogen receptor alpha. Front. Biosci. 2019, 24, 245–276. [Google Scholar] [CrossRef]
- Bryant, D.N.; Bosch, M.A.; Ronnekleiv, O.K.; Dorsa, D.M. 17-Beta estradiol rapidly enhances extracellular signal-regulated kinase 2 phosphorylation in the rat brain. Neuroscience 2005, 133, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Wade, C.B.; Dorsa, D.M. Estrogen activation of cyclic adenosine 5′-monophosphate response element-mediated transcription requires the extracellularly regulated kinase/mitogen-activated protein kinase pathway. Endocrinology 2003, 144, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Glidewell-Kenney, C.; Hurley, L.A.; Pfaff, L.; Weiss, J.; Levine, J.E.; Jameson, J.L. Nonclassical estrogen receptor alpha signaling mediates negative feedback in the female mouse reproductive axis. Proc. Natl. Acad. Sci. USA 2007, 104, 8173–8177. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, M.A.; Glidewell-Kenney, C.; Jimenez, M.A.; Ahearn, P.C.; Weiss, J.; Jameson, J.L.; Levine, J.E. New insights into the classical and non-classical actions of estrogen: Evidence from estrogen receptor knock-out and knock-in mice. Mol. Cell. Endocrinol. 2008, 290, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Szego, C.M.; Davis, J.S. Adenosine 3′,5′-monophosphate in rat uterus: Acute elevation by estrogen. Proc. Natl. Acad. Sci. USA 1967, 58, 1711–1718. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuhanna, I.S.; Galcheva-Gargova, Z.; Karas, R.H.; Mendelsohn, M.E.; Shaul, P.W. Estrogen receptor alpha mediates the nongenomic activation of endothelial nitric oxide synthase by estrogen. J. Clin. Investig. 1999, 103, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.; Steelman, L.S.; Lee, J.T.; Shelton, J.G.; Navolanic, P.M.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Signal transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine receptors to transcription factors: Potential targeting for therapeutic intervention. Leukemia 2003, 17, 1263–1293. [Google Scholar] [CrossRef] [PubMed]
- Mhyre, A.J.; Dorsa, D.M. Estrogen activates rapid signaling in the brain: Role of estrogen receptor alpha and estrogen receptor beta in neurons and glia. Neuroscience 2006, 138, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Barabas, K.; Szego, E.M.; Kaszas, A.; Nagy, G.M.; Juhasz, G.D.; Abraham, I.M. Sex differences in oestrogen-induced p44/42 MAPK phosphorylation in the mouse brain in vivo. J Neuroendocr. 2006, 18, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Haynes, M.P.; Li, L.; Sinha, D.; Russell, K.S.; Hisamoto, K.; Baron, R.; Collinge, M.; Sessa, W.C.; Bender, J.R. Src kinase mediates phosphatidylinositol 3-kinase/Akt-dependent rapid endothelial nitric-oxide synthase activation by estrogen. J. Biol. Chem. 2003, 278, 2118–2123. [Google Scholar] [CrossRef] [PubMed]
- Hisamoto, K.; Ohmichi, M.; Kurachi, H.; Hayakawa, J.; Kanda, Y.; Nishio, Y.; Adachi, K.; Tasaka, K.; Miyoshi, E.; Fujiwara, N.; et al. Estrogen induces the Akt-dependent activation of endothelial nitric-oxide synthase in vascular endothelial cells. J. Biol. Chem. 2001, 276, 3459–3467. [Google Scholar] [CrossRef] [PubMed]
- Simoncini, T.; Fornari, L.; Mannella, P.; Varone, G.; Caruso, A.; Liao, J.K.; Genazzani, A.R. Novel non-transcriptional mechanisms for estrogen receptor signaling in the cardiovascular system. Interaction of estrogen receptor alpha with phosphatidylinositol 3-OH kinase. Steroids 2002, 67, 935–939. [Google Scholar] [CrossRef]
- Chen, D.-B.; Bird, I.M.; Zheng, J.; Magness, R.R. Membrane estrogen receptor-dependent extracellular signal-regulated kinase pathway mediates acute activation of endothelial nitric oxide synthase by estrogen in uterine artery endothelial cells. Endocrinology 2004, 145, 113–125. [Google Scholar] [CrossRef]
- Sirohi, V.K.; Medrano, T.I.; Mesa, A.M.; Kannan, A.; Bagchi, I.C.; Cooke, P.S. Regulation of AKT Signaling in Mouse Uterus. Endocrinology 2022, 163, bqab233. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, X.; Liu, P.; Li, P.; Xu, X.; Chen, Y.; Cheng, Y.; Zhu, D.; Fu, X. 17β-estradiol promotes angiogenesis through non-genomic activation of Smad1 signaling in endometriosis. Vasc. Pharmacol. 2021, 142, 106932. [Google Scholar] [CrossRef] [PubMed]
- Ohshiro, K.; Schwartz, A.M.; Levine, P.H.; Kumar, R. Alternate estrogen receptors promote invasion of inflammatory breast cancer cells via non-genomic signaling. PLoS ONE 2012, 7, e30725. [Google Scholar] [CrossRef] [PubMed]
- Brindle, P.; Linke, S.; Montminy, M. Protein-kinase-A-dependent activator in transcription factor CREB reveals new role for CREM repressers. Nature 1993, 364, 821. [Google Scholar] [CrossRef] [PubMed]
- Shaywitz, A.J.; Greenberg, M.E. CREB: A stimulus-induced transcription factor activated by a diverse array of extracellular signals. Annu. Rev. Biochem. 1999, 68, 821–861. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, F.N.; Lucas, T.F.G.; Lazari, M.F.M.; Porto, C.S. Estrogen receptor ESR1 mediates activation of ERK1/2, CREB, and ELK1 in the corpus of the epididymis. J. Mol. Endocrinol. 2015, 54, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Aronica, S.M.; Kraus, W.L.; Katzenellenbogen, B.S. Estrogen action via the cAMP signaling pathway: Stimulation of adenylate cyclase and cAMP-regulated gene transcription. Proc. Natl. Acad. Sci. USA 1994, 91, 8517–8521. [Google Scholar] [CrossRef] [PubMed]
- Chaban, V.V.; Lakhter, A.J.; Micevych, P. A membrane estrogen receptor mediates intracellular calcium release in astrocytes. Endocrinology 2004, 145, 3788–3795. [Google Scholar] [CrossRef] [PubMed]
- Mareş Miceli, A. Replacement Hormone Therapy for Gender Dysphoria and Congenital Sexual Anomalies. Adv. Exp. Med. Biol. 2020, 1242, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.A. Feminine Forever; J.B. Lippincott Co.: New York, NY, USA, 1966. [Google Scholar]
- Lobo, R.A.; Pickar, J.H.; Stevenson, J.C.; Mack, W.J.; Hodis, H.N. Back to the future: Hormone replacement therapy as part of a prevention strategy for women at the onset of menopause. Atherosclerosis 2016, 254, 282–290. [Google Scholar] [CrossRef]
- Grodstein, F.; Stampfer, M. The epidemiology of coronary heart disease and estrogen replacement in postmenopausal women. Prog. Cardiovasc. Dis. 1995, 38, 199–210. [Google Scholar] [CrossRef]
- Hulley, S.; Grady, D.; Bush, T.; Furberg, C.; Herrington, D.; Riggs, B.; Vittinghoff, E. Randomized trial of estrogen plus progestin for secondary prevention of coronary heart disease in postmenopausal women. Heart and Estrogen/progestin Replacement Study (HERS) Research Group. JAMA 1998, 280, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.L.; Limacher, M.; Assaf, A.R.; Bassford, T.; Beresford, S.A.; Black, H.; Bonds, D.; Brunner, R.; Brzyski, R.; Caan, B.; et al. Effects of conjugated equine estrogen in postmenopausal women with hysterectomy: The Women’s Health Initiative randomized controlled trial. JAMA 2004, 291, 1701–1712. [Google Scholar] [CrossRef] [PubMed]
- Shumaker, S.A.; Legault, C.; Rapp, S.R.; Thal, L.; Wallace, R.B.; Ockene, J.K.; Hendrix, S.L.; Jones, B.N., 3rd; Assaf, A.R.; Jackson, R.D.; et al. Estrogen plus progestin and the incidence of dementia and mild cognitive impairment in postmenopausal women: The Women’s Health Initiative Memory Study: A randomized controlled trial. JAMA 2003, 289, 2651–2662. [Google Scholar] [CrossRef] [PubMed]
- Shumaker, S.A.; Legault, C.; Kuller, L.; Rapp, S.R.; Thal, L.; Lane, D.S.; Fillit, H.; Stefanick, M.L.; Hendrix, S.L.; Lewis, C.E.; et al. Conjugated equine estrogens and incidence of probable dementia and mild cognitive impairment in postmenopausal women: Women’s Health Initiative Memory Study. JAMA 2004, 291, 2947–2958. [Google Scholar] [CrossRef] [PubMed]
- Chlebowski, R.T.; Anderson, G.L.; Aragaki, A.K.; Manson, J.E.; Stefanick, M.L.; Pan, K.; Barrington, W.; Kuller, L.H.; Simon, M.S.; Lane, D.; et al. Association of Menopausal Hormone Therapy With Breast Cancer Incidence and Mortality During Long-term Follow-up of the Women’s Health Initiative Randomized Clinical Trials. JAMA 2020, 324, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Tan, C.-C.; Xu, W.; Hu, H.; Cao, X.-P.; Dong, Q.; Tan, L.; Yu, J.-T. The Prevalence of Dementia: A Systematic Review and Meta-Analysis. J. Alzheimers Dis. 2020, 73, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.T.; Maki, P.M.; McDermott, M.P. Cognition and mood in perimenopause: A systematic review and meta-analysis. J. Steroid Biochem. Mol. Biol. 2014, 142, 90–98. [Google Scholar] [CrossRef]
- Irvine, K.; Laws, K.R.; Gale, T.M.; Kondel, T.K. Greater cognitive deterioration in women than men with Alzheimer’s disease: A meta analysis. J. Clin. Exp. Neuropsychol. 2012, 34, 989–998. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Alves, S.E. Estrogen actions in the central nervous system. Endocr. Rev. 1999, 20, 279–307. [Google Scholar] [CrossRef]
- Roof, R.L.; Hall, E.D. Gender differences in acute CNS trauma and stroke: Neuroprotective effects of estrogen and progesterone. J. Neurotrauma 2000, 17, 367–388. [Google Scholar] [CrossRef]
- Pike, C.J. Sex and the development of Alzheimer’s disease. J. Neurosci. Res. 2017, 95, 671–680. [Google Scholar] [CrossRef]
- Xu, W.; Tan, L.; Wang, H.-F.; Jiang, T.; Tan, M.-S.; Tan, L.; Zhao, Q.-F.; Li, J.-Q.; Wang, J.; Yu, J.-T. Meta-analysis of modifiable risk factors for Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2015, 86, 1299–1306. [Google Scholar] [CrossRef]
- Stute, P.; Wienges, J.; Koller, A.-S.; Giese, C.; Wesemüller, W.; Janka, H.; Baumgartner, S. Cognitive health after menopause: Does menopausal hormone therapy affect it? Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101565. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Jackson, J.W.; Grodstein, F.; Blacker, D.; Weuve, J. Postmenopausal hormone therapy is not associated with risk of all-cause dementia and Alzheimer’s disease. Epidemiol. Rev. 2014, 36, 83–103. [Google Scholar] [CrossRef]
- Shumaker, S.A.; Reboussin, B.A.; Espeland, M.A.; Rapp, S.R.; McBee, W.L.; Dailey, M.; Bowen, D.; Terrell, T.; Jones, B.N. The Women’s Health Initiative Memory Study (WHIMS): A trial of the effect of estrogen therapy in preventing and slowing the progression of dementia. Control. Clin. Trials 1998, 19, 604–621. [Google Scholar] [CrossRef]
- Espeland, M.A.; Rapp, S.R.; Shumaker, S.A.; Brunner, R.; Manson, J.E.; Sherwin, B.B.; Hsia, J.; Margolis, K.L.; Hogan, P.E.; Wallace, R.; et al. Conjugated equine estrogens and global cognitive function in postmenopausal women: Women’s Health Initiative Memory Study. JAMA 2004, 291, 2959–2968. [Google Scholar] [CrossRef] [PubMed]
- Craig, M.C.; Maki, P.M.; Murphy, D.G.M. The Women’s Health Initiative Memory Study: Findings and implications for treatment. Lancet. Neurol. 2005, 4, 190–194. [Google Scholar] [CrossRef]
- Coker, L.H.; Espeland, M.A.; Rapp, S.R.; Legault, C.; Resnick, S.M.; Hogan, P.; Gaussoin, S.; Dailey, M.; Shumaker, S.A. Postmenopausal hormone therapy and cognitive outcomes: The Women’s Health Initiative Memory Study (WHIMS). J. Steroid Biochem. Mol. Biol. 2010, 118, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Krug, R.; Beier, L.; Lämmerhofer, M.; Hallschmid, M. Distinct and Convergent Beneficial Effects of Estrogen and Insulin on Cognitive Function in Healthy Young Men. J. Clin. Endocrinol. Metab. 2022, 107, e582–e593. [Google Scholar] [CrossRef] [PubMed]
- Plouffe, L.J. Selective estrogen receptor modulators (SERMs) in clinical practice. J. Soc. Gynecol. Investig. 2000, 7, S38–S46. [Google Scholar] [CrossRef]
- Dickey, R.P.; Holtkamp, D.E. Development, pharmacology and clinical experience with clomiphene citrate. Hum. Reprod. Update 1996, 2, 483–506. [Google Scholar] [CrossRef] [PubMed]
- Trost, L.W.; Khera, M. Alternative treatment modalities for the hypogonadal patient. Curr. Urol. Rep. 2014, 15, 417. [Google Scholar] [CrossRef]
- Uyar, Y.; Koltan, S.O.; Pögün, S.; Vatansever, S.; Caglar, H. The effect of clomiphene citrate on osteoporosis in ovariectomized rats. Arch. Gynecol. Obstet. 2008, 278, 107–114. [Google Scholar] [CrossRef]
- Kacker, R.; Conners, W.; Zade, J.; Morgentaler, A. Bone mineral density and response to treatment in men younger than 50 years with testosterone deficiency and sexual dysfunction or infertility. J. Urol. 2014, 191, 1072–1076. [Google Scholar] [CrossRef]
- Jordan, V.C.; Collins, M.M.; Rowsby, L.; Prestwich, G. A monohydroxylated metabolite of tamoxifen with potent antioestrogenic activity. J. Endocrinol. 1977, 75, 305–316. [Google Scholar] [CrossRef]
- World Health Organization. World Health Organization Model List of Essential Medicines, 21st List, 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- “Tamoxifen Citrate—Drug Usage Statistics” ClinCalc. Archived from the Original on 22 September 2020. Available online: https://clincalc.com/DrugStats/Drugs/Tamoxifen (accessed on 18 February 2021).
- Osborne, C.K.; Boldt, D.H.; Clark, G.M.; Trent, J.M. Effects of tamoxifen on human breast cancer cell cycle kinetics: Accumulation of cells in early G1 phase. Cancer Res. 1983, 43, 3583–3585. [Google Scholar]
- Early Breast Cancer Trialists’ Collaborative Group. Tamoxifen for early breast cancer: An overview of the randomised trials. Lancet 1998, 351, 1451–1467. [Google Scholar] [CrossRef]
- Davies, C.; Godwin, J.; Gray, R.; Clarke, M.; Cutter, D.; Darby, S.; McGale, P.; Pan, H.C.; Taylor, C.; Wang, Y.C.; et al. Relevance of breast cancer hormone receptors and other factors to the efficacy of adjuvant tamoxifen: Patient-level meta-analysis of randomised trials. Lancet 2011, 378, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, L.K.; Suri, V.; Gupta, K.R.; Sahdev, S. Tamoxifen: An alternative to clomiphene in women with polycystic ovary syndrome. J. Hum. Reprod. Sci. 2011, 4, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Michael, H.; Härkönen, P.L.; Kangas, L.; Väänänen, H.K.; Hentunen, T.A. Differential effects of selective oestrogen receptor modulators (SERMs) tamoxifen, ospemifene and raloxifene on human osteoclasts in vitro. Br. J. Pharmacol. 2007, 151, 384–395. [Google Scholar] [CrossRef]
- Kristensen, B.; Ejlertsen, B.; Dalgaard, P.; Larsen, L.; Holmegaard, S.N.; Transbøl, I.; Mouridsen, H.T. Tamoxifen and bone metabolism in postmenopausal low-risk breast cancer patients: A randomized study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1994, 12, 992–997. [Google Scholar] [CrossRef]
- Vehmanen, L.; Elomaa, I.; Blomqvist, C.; Saarto, T. Tamoxifen treatment after adjuvant chemotherapy has opposite effects on bone mineral density in premenopausal patients depending on menstrual status. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 675–680. [Google Scholar] [CrossRef]
- Ramin, C.; May, B.J.; Roden, R.B.S.; Orellana, M.M.; Hogan, B.C.; McCullough, M.S.; Petry, D.; Armstrong, D.K.; Visvanathan, K. Evaluation of osteopenia and osteoporosis in younger breast cancer survivors compared with cancer-free women: A prospective cohort study. Breast Cancer Res. 2018, 20, 134. [Google Scholar] [CrossRef]
- Early Breast Cancer Trialists’ Collaborative Group. Effects of adjuvant tamoxifen and of cytotoxic therapy on mortality in early breast cancer. An overview of 61 randomized trials among 28,896 women. N. Engl. J. Med. 1988, 319, 1681–1692. [Google Scholar] [CrossRef]
- Franco Rodríguez, N.E.; Dueñas Jiménez, J.M.; De la Torre Valdovinos, B.; López Ruiz, J.R.; Hernández Hernández, L.; Dueñas Jiménez, S.H. Tamoxifen favoured the rat sensorial cortex regeneration after a penetrating brain injury. Brain Res. Bull. 2013, 98, 64–75. [Google Scholar] [CrossRef]
- Gonzalez, G.A.; Hofer, M.P.; Syed, Y.A.; Amaral, A.I.; Rundle, J.; Rahman, S.; Zhao, C.; Kotter, M.R.N. Tamoxifen accelerates the repair of demyelinated lesions in the central nervous system. Sci. Rep. 2016, 6, 31599. [Google Scholar] [CrossRef]
- Baez-Jurado, E.; Rincón-Benavides, M.A.; Hidalgo-Lanussa, O.; Guio-Vega, G.; Ashraf, G.M.; Sahebkar, A.; Echeverria, V.; Garcia-Segura, L.M.; Barreto, G.E. Molecular mechanisms involved in the protective actions of Selective Estrogen Receptor Modulators in brain cells. Front. Neuroendocrinol. 2019, 52, 44–64. [Google Scholar] [CrossRef] [PubMed]
- Seoud, M.A.; Johnson, J.; Weed, J.C.J. Gynecologic tumors in tamoxifen-treated women with breast cancer. Obstet. Gynecol. 1993, 82, 165–169. [Google Scholar] [CrossRef]
- Ellis, A.J.; Hendrick, V.M.; Williams, R.; Komm, B.S. Selective estrogen receptor modulators in clinical practice: A safety overview. Expert Opin. Drug Saf. 2015, 14, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Yuan, Z.Y.; Peng, R.J.; Zeng, Y.D.; Shi, Y.X.; Teng, X.Y.; Liu, D.G.; Bai, B.; Wang, S.S. Efficacy and tolerability of toremifene and tamoxifen therapy in premenopausal patients with operable breast cancer: A retrospective analysis. Curr. Oncol. 2013, 20, 196–204. [Google Scholar] [CrossRef]
- Shibutani, S.; Ravindernath, A.; Terashima, I.; Suzuki, N.; Laxmi, Y.R.; Kanno, Y.; Suzuki, M.; Apak, T.I.; Sheng, J.J.; Duffel, M.W. Mechanism of lower genotoxicity of toremifene compared with tamoxifen. Cancer Res. 2001, 61, 3925–3931. [Google Scholar]
- Ettinger, B.; Black, D.M.; Mitlak, B.H.; Knickerbocker, R.K.; Nickelsen, T.; Genant, H.K.; Christiansen, C.; Delmas, P.D.; Zanchetta, J.R.; Stakkestad, J.; et al. Reduction of vertebral fracture risk in postmenopausal women with osteoporosis treated with raloxifene: Results from a 3-year randomized clinical trial. Multiple Outcomes of Raloxifene Evaluation (MORE) Investigators. JAMA 1999, 282, 637–645. [Google Scholar] [CrossRef]
- Martino, S.; Cauley, J.A.; Barrett-Connor, E.; Powles, T.J.; Mershon, J.; Disch, D.; Secrest, R.J.; Cummings, S.R. Continuing outcomes relevant to Evista: Breast cancer incidence in postmenopausal osteoporotic women in a randomized trial of raloxifene. J. Natl. Cancer Inst. 2004, 96, 1751–1761. [Google Scholar] [CrossRef]
- Wickerham, D.L.; Tan-Chiu, E. Breast cancer chemoprevention: Current status and future directions. Semin. Oncol. 2001, 28, 253–259. [Google Scholar] [CrossRef]
- Lewis-Wambi, J.S.; Kim, H.; Curpan, R.; Grigg, R.; Sarker, M.A.; Jordan, V.C. The selective estrogen receptor modulator bazedoxifene inhibits hormone-independent breast cancer cell growth and down-regulates estrogen receptor α and cyclin D1. Mol. Pharmacol. 2011, 80, 610–620. [Google Scholar] [CrossRef]
- Wardell, S.E.; Nelson, E.R.; Chao, C.A.; McDonnell, D.P. Bazedoxifene exhibits antiestrogenic activity in animal models of tamoxifen-resistant breast cancer: Implications for treatment of advanced disease. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 2420–2431. [Google Scholar] [CrossRef]
- Christiansen, C.; Chesnut, C.H., 3rd; Adachi, J.D.; Brown, J.P.; Fernandes, C.E.; Kung, A.W.; Palacios, S.; Levine, A.B.; Chines, A.A.; Constantine, G.D. Safety of bazedoxifene in a randomized, double-blind, placebo- and active-controlled Phase 3 study of postmenopausal women with osteoporosis. BMC Musculoskelet. Disord. 2010, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Kouremenos, K.; Tull, D.; Maggi, A.; Schroeder, A.; Gibbons, A.; Kulkarni, J.; Sundram, S.; Du, X. Bazedoxifene—A promising brain active SERM that crosses the blood brain barrier and enhances spatial memory. Psychoneuroendocrinology 2020, 121, 104830. [Google Scholar] [CrossRef] [PubMed]
- Pinkerton, J.V. Selective Estrogen Receptor Modulators in Gynecology Practice. Clin. Obstet. Gynecol. 2021, 64, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Zheng, H.; Dahllund, J.; Laine, A.; Cockcroft, N.; Peng, Z.; Koskinen, M.; Hemminki, K.; Kangas, L.; Väänänen, K.; et al. Selective estrogenic effects of a novel triphenylethylene compound, FC1271a, on bone, cholesterol level, and reproductive tissues in intact and ovariectomized rats. Endocrinology 2000, 141, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H. Selective estrogen receptor modulators (SERMS): Keys to understanding their function. Menopause 2020, 27, 1171–1176. [Google Scholar] [CrossRef]
- Cummings, S.R.; Ensrud, K.; Delmas, P.D.; LaCroix, A.Z.; Vukicevic, S.; Reid, D.M.; Goldstein, S.; Sriram, U.; Lee, A.; Thompson, J.; et al. Lasofoxifene in postmenopausal women with osteoporosis. N. Engl. J. Med. 2010, 362, 686–696. [Google Scholar] [CrossRef]
- Pinkerton, J.V.; Thomas, S. Use of SERMs for treatment in postmenopausal women. J. Steroid Biochem. Mol. Biol. 2014, 142, 142–154. [Google Scholar] [CrossRef]
- Hwang, C.S.; Kwak, H.S.; Lim, H.J.; Lee, S.H.; Kang, Y.S.; Choe, T.B.; Hur, H.G.; Han, K.O. Isoflavone metabolites and their in vitro dual functions: They can act as an estrogenic agonist or antagonist depending on the estrogen concentration. J. Steroid Biochem. Mol. Biol. 2006, 101, 246–253. [Google Scholar] [CrossRef]
- Setchell, K.D.; Zimmer-Nechemias, L.; Cai, J.; Heubi, J.E. Exposure of infants to phyto-oestrogens from soy-based infant formula. Lancet 1997, 350, 23–27. [Google Scholar] [CrossRef]
- Najaf Najafi, M.; Ghazanfarpour, M. Effect of phytoestrogens on sexual function in menopausal women: A systematic review and meta-analysis. Climacteric 2018, 21, 437–445, 1076. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.E.; Nickell, L.A.; Thompson, L.U.; Szalai, J.P.; Kiss, A.; Hilditch, J.R. A randomized controlled trial of the effect of dietary soy and flaxseed muffins on quality of life and hot flashes during menopause. Menopause 2006, 13, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Gao, Y.H. Inhibitory effect of genistein on bone resorption in tissue culture. Biochem. Pharmacol. 1998, 55, 71–76. [Google Scholar] [CrossRef]
- Jin, X.; Sun, J.; Yu, B.; Wang, Y.; Sun, W.J.; Yang, J.; Huang, S.H.; Xie, W.L. Daidzein stimulates osteogenesis facilitating proliferation, differentiation, and antiapoptosis in human osteoblast-like MG-63 cells via estrogen receptor-dependent MEK/ERK and PI3K/Akt activation. Nutr. Res. 2017, 42, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Mao, Z.; Brinton, R.D. A select combination of clinically relevant phytoestrogens enhances estrogen receptor beta-binding selectivity and neuroprotective activities in vitro and in vivo. Endocrinology 2009, 150, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Mao, Z.; Chen, S.; Schneider, L.S.; Brinton, R.D. Early intervention with an estrogen receptor β-selective phytoestrogenic formulation prolongs survival, improves spatial recognition memory, and slows progression of amyloid pathology in a female mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2013, 37, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.; Maier, C. Phytoestrogens and breast cancer: In vitro anticancer activities of isoflavones, lignans, coumestans, stilbenes and their analogs and derivatives. Biomed. Pharmacother. 2018, 107, 1648–1666. [Google Scholar] [CrossRef]
- Squadrito, F.; Altavilla, D.; Squadrito, G.; Saitta, A.; Cucinotta, D.; Minutoli, L.; Deodato, B.; Ferlito, M.; Campo, G.M.; Bova, A.; et al. Genistein supplementation and estrogen replacement therapy improve endothelial dysfunction induced by ovariectomy in rats. Cardiovasc. Res. 2000, 45, 454–462. [Google Scholar] [CrossRef]
- Polini, N.; Rauschemberger, M.B.; Mendiberri, J.; Selles, J.; Massheimer, V. Effect of genistein and raloxifene on vascular dependent platelet aggregation. Mol. Cell. Endocrinol. 2007, 267, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.K.; Bihani, T. Selective estrogen receptor modulators (SERMs) and selective estrogen receptor degraders (SERDs) in cancer treatment. Pharmacol. Ther. 2018, 186, 1–24. [Google Scholar] [CrossRef]
- Wardell, S.E.; Marks, J.R.; McDonnell, D.P. The turnover of estrogen receptor α by the selective estrogen receptor degrader (SERD) fulvestrant is a saturable process that is not required for antagonist efficacy. Biochem. Pharmacol. 2011, 82, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Fulvestrant as a reference antiestrogen and estrogen receptor (ER) degrader in preclinical studies: Treatment dosage, efficacy, and implications on development of new ER-targeting agents. Transl. Cancer Res. 2020, 9, 4464–4468. [Google Scholar] [CrossRef]
- Cristofanilli, M.; Turner, N.C.; Bondarenko, I.; Ro, J.; Im, S.-A.; Masuda, N.; Colleoni, M.; DeMichele, A.; Loi, S.; Verma, S.; et al. Fulvestrant plus palbociclib versus fulvestrant plus placebo for treatment of hormone-receptor-positive, HER2-negative metastatic breast cancer that progressed on previous endocrine therapy (PALOMA-3): Final analysis of the multicentre, double-blind, phas. Lancet Oncol. 2016, 17, 425–439. [Google Scholar] [CrossRef]
- Covens, A.L.; Filiaci, V.; Gersell, D.; Lutman, C.V.; Bonebrake, A.; Lee, Y.-C. Phase II study of fulvestrant in recurrent/metastatic endometrial carcinoma: A Gynecologic Oncology Group Study. Gynecol. Oncol. 2011, 120, 185–188. [Google Scholar] [CrossRef]
- McDonnell, D.P.; Wardell, S.E.; Norris, J.D. Oral Selective Estrogen Receptor Downregulators (SERDs), a Breakthrough Endocrine Therapy for Breast Cancer. J. Med. Chem. 2015, 58, 4883–4887. [Google Scholar] [CrossRef] [PubMed]
- Shagufta; Ahmad, I.; Mathew, S.; Rahman, S. Recent progress in selective estrogen receptor downregulators (SERDs) for the treatment of breast cancer. RSC Med. Chem. 2020, 11, 438–454. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C.; Kousteni, S.; Jilka, R.L. Sex steroids and bone. Recent Prog. Horm. Res. 2002, 57, 385–409. [Google Scholar] [CrossRef]
- Kwakowsky, A.; Koszegi, Z.; Cheong, R.Y.; Abraham, I.M. Neuroprotective effects of non-classical estrogen-like signaling activators: From mechanism to potential implications. CNS Neurol. Disord. Drug Targets 2013, 12, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Fuchs, I.; Altmann, H.; Klewer, M.; Schwarz, G.; Bohlmann, R.; Nguyen, D.; Zorn, L.; Vonk, R.; Prelle, K.; et al. In vivo characterization of estrogen receptor modulators with reduced genomic versus nongenomic activity in vitro. J. Steroid Biochem. Mol. Biol. 2008, 111, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Wessler, S.; Otto, C.; Wilck, N.; Stangl, V.; Fritzemeier, K.-H. Identification of estrogen receptor ligands leading to activation of non-genomic signaling pathways while exhibiting only weak transcriptional activity. J. Steroid Biochem. Mol. Biol. 2006, 98, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Koszegi, Z.; Szego, É.M.; Cheong, R.Y.; Tolod-Kemp, E.; Ábrahám, I.M. Postlesion estradiol treatment increases cortical cholinergic innervations via estrogen receptor-α dependent nonclassical estrogen signaling in vivo. Endocrinology 2011, 152, 3471–3482. [Google Scholar] [CrossRef] [PubMed]
- Kwakowsky, A.; Potapov, K.; Kim, S.; Peppercorn, K.; Tate, W.P.; Ábrahám, I.M. Treatment of beta amyloid 1-42 (Aβ 1-42)-induced basal forebrain cholinergic damage by a non-classical estrogen signaling activator in vivo. Sci. Rep. 2016, 6, 21101. [Google Scholar] [CrossRef] [PubMed]
- Bologa, C.G.; Revankar, C.M.; Young, S.M.; Edwards, B.S.; Arterburn, J.B.; Kiselyov, A.S.; Parker, M.A.; Tkachenko, S.E.; Savchuck, N.P.; Sklar, L.A.; et al. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat. Chem. Biol. 2006, 2, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.R.; Fredette, N.C.; Howard, T.A.; Hu, C.; Ramesh, C.; Daniel, C.; Amann, K.; Arterburn, J.B.; Barton, M.; Prossnitz, E.R. G protein-coupled estrogen receptor protects from atherosclerosis. Sci. Rep. 2014, 4, 7564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, Y.; Guo, L.; Zhou, J.; Niu, J.; Wang, P.; Qiang, Y.; Liu, K.; Wen, Y.; Zhang, L.; et al. GPER1 Modulates Synaptic Plasticity During the Development of Temporal Lobe Epilepsy in Rats. Neurochem. Res. 2021, 46, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Subramanian, S.; Dziennis, S.; Jia, J.; Uchida, M.; Akiyoshi, K.; Migliati, E.; Lewis, A.D.; Vandenbark, A.A.; Offner, H.; et al. Estradiol and G1 Reduce Infarct Size and Improve Immunosuppression after Experimental Stroke. J. Immunol. 2010, 184, 4087–4094. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.-C.; Chen, C.-H.; Chou, Y.-S.; Ho, M.-L.; Chang, J.-K. G Protein-Coupled Estrogen Receptor Mediates Cell Proliferation through the cAMP/PKA/CREB Pathway in Murine Bone Marrow Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 6490. [Google Scholar] [CrossRef] [PubMed]
- Scaling, A.L.; Prossnitz, E.R.; Hathaway, H.J. GPER mediates estrogen-induced signaling and proliferation in human breast epithelial cells and normal and malignant breast. Horm. Cancer 2014, 5, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.K.; Burai, R.; Ramesh, C.; Petrie, W.K.; Alcon, S.N.; Nayak, T.K.; Bologa, C.G.; Leitao, A.; Brailoiu, E.; Deliu, E.; et al. In vivo effects of a GPR30 antagonist. Nat. Chem. Biol. 2009, 5, 421–427. [Google Scholar] [CrossRef]
- Dennis, M.K.; Field, A.S.; Burai, R.; Ramesh, C.; Petrie, W.K.; Bologa, C.G.; Oprea, T.I.; Yamaguchi, Y.; Hayashi, S.-I.; Sklar, L.A.; et al. Identification of a GPER/GPR30 antagonist with improved estrogen receptor counterselectivity. J. Steroid Biochem. Mol. Biol. 2011, 127, 358–366. [Google Scholar] [CrossRef]
- Lønning, P.E. The potency and clinical efficacy of aromatase inhibitors across the breast cancer continuum. Ann. Oncol. 2011, 22, 503–514. [Google Scholar] [CrossRef]
- Awouters, M.; Vanderschueren, D.; Antonio, L. Aromatase inhibitors and selective estrogen receptor modulators: Unconventional therapies for functional hypogonadism? Andrology 2020, 8, 1590–1597. [Google Scholar] [CrossRef]
- Sanderson, J.T.; Hordijk, J.; Denison, M.S.; Springsteel, M.F.; Nantz, M.H.; van den Berg, M. Induction and Inhibition of Aromatase (CYP19) Activity by Natural and Synthetic Flavonoid Compounds in H295R Human Adrenocortical Carcinoma Cells. Toxicol. Sci. 2004, 82, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Reeve, J. Future prospects for hormone replacement therapy. Br. Med. Bull. 1992, 48, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Cho, M.K.; Cho, Y.J.; Chun, S.; Hong, S.H.; Hwang, K.R.; Jeon, G.H.; Joo, J.K.; Kim, S.K.; Lee, D.O.; et al. The 2020 Menopausal Hormone Therapy Guidelines. J. Menopausal Med. 2020, 26, 69–98. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Oung, T.; Yung, S.; Flagg, R.A.; Woller, M.J. Neuroendocrine mechanisms for reproductive senescence in the female rat: Gonadotropin-releasing hormone neurons. Endocrine 2000, 13, 315–323. [Google Scholar] [CrossRef]
- Oyola, M.G.; Handa, R.J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- Haisenleder, D.J.; Dalkin, A.C.; Ortolano, G.A.; Marshall, J.C.; Shupnik, M.A. A pulsatile gonadotropin-releasing hormone stimulus is required to increase transcription of the gonadotropin subunit genes: Evidence for differential regulation of transcription by pulse frequency in vivo. Endocrinology 1991, 128, 509–517. [Google Scholar] [CrossRef]
- Herbison, A.E.; Porteous, R.; Pape, J.-R.; Mora, J.M.; Hurst, P.R. Gonadotropin-releasing hormone neuron requirements for puberty, ovulation, and fertility. Endocrinology 2008, 149, 597–604. [Google Scholar] [CrossRef]
- Marques, P.; Skorupskaite, K.; George, J.T.; Anderson, R.A. Physiology of GNRH and Gonadotropin Secretion. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Barabás, K.; Szabó-Meleg, E.; Ábrahám, I.M. Effect of Inflammation on Female Gonadotropin-Releasing Hormone (GnRH) Neurons: Mechanisms and Consequences. Int. J. Mol. Sci. 2020, 21, 529. [Google Scholar] [CrossRef]
- Herbison, A.E. A simple model of estrous cycle negative and positive feedback regulation of GnRH secretion. Front. Neuroendocrinol. 2020, 57, 100837. [Google Scholar] [CrossRef] [PubMed]
- Park, O.K.; Ramirez, V.D. Spontaneous changes in LHRH release during the rat estrous cycle, as measured with repetitive push-pull perfusions of the pituitary gland in the same female rats. Neuroendocrinology 1989, 50, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Kwakowsky, A.; Herbison, A.E.; Ábrahám, I.M. The role of cAMP response element-binding protein in estrogen negative feedback control of gonadotropin-releasing hormone neurons. J. Neurosci. 2012, 32, 11309–11317. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.K.; Chiappa, S.A.; Fink, G.; Sherwood, N.M. Gonadotropin-releasing hormone surge in pro-oestrous rats. Nature 1976, 264, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Clarke, I.J.; Smith, J.T.; Henry, B.A.; Oldfield, B.J.; Stefanidis, A.; Millar, R.P.; Sari, I.P.; Chng, K.; Fabre-Nys, C.; Caraty, A.; et al. Gonadotropin-inhibitory hormone is a hypothalamic peptide that provides a molecular switch between reproduction and feeding. Neuroendocrinology 2012, 95, 305–316. [Google Scholar] [CrossRef]
- Herbison, A.E. Rapid actions of oestrogen on gonadotropin-releasing hormone neurons; from fantasy to physiology? J. Physiol. 2009, 587, 5025–5030. [Google Scholar] [CrossRef]
- Mills, R.H.; Sohn, R.K.; Micevych, P.E. Estrogen-induced mu-opioid receptor internalization in the medial preoptic nucleus is mediated via neuropeptide Y-Y1 receptor activation in the arcuate nucleus of female rats. J. Neurosci. 2004, 24, 947–955. [Google Scholar] [CrossRef]
- Foresta, C.; De Toni, L.; Di Mambro, A.; Ferlin, A.; Perilli, L.; Bertuzzi, I.; Galan, A.; Zuccarello, D. Role of estrogen receptors in menstrual cycle-related neoangiogenesis and their influence on endothelial progenitor cell physiology. Fertil. Steril. 2010, 93, 220–228. [Google Scholar] [CrossRef]
- Mesiano, S. Roles of estrogen and progesterone in human parturition. Front. Horm. Res. 2001, 27, 86–104. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. The G-protein-coupled estrogen receptor GPER in health and disease. Nat. Rev. Endocrinol. 2011, 7, 715–726. [Google Scholar] [CrossRef]
- Kolkova, Z.; Noskova, V.; Ehinger, A.; Hansson, S.; Casslén, B. G protein-coupled estrogen receptor 1 (GPER, GPR 30) in normal human endometrium and early pregnancy decidua. Mol. Hum. Reprod. 2010, 16, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Prossnitz, E.R.; Roy, S.K. Expression of G protein-coupled receptor 30 in the hamster ovary: Differential regulation by gonadotropins and steroid hormones. Endocrinology 2007, 148, 4853–4864. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Prossnitz, E.R.; Roy, S.K. G protein-coupled receptor 30 expression is required for estrogen stimulation of primordial follicle formation in the hamster ovary. Endocrinology 2008, 149, 4452–4461. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Wang, Y.; Yang, F.; Li, J.; Qin, X. Melatonin rescues the reproductive toxicity of low-dose glyphosate-based herbicide during mouse oocyte maturation via the GPER signaling pathway. J. Pineal Res. 2021, 70, e12718. [Google Scholar] [CrossRef]
- Li, Y.-R.; Ren, C.-E.; Zhang, Q.; Li, J.-C.; Chian, R.-C. Expression of G protein estrogen receptor (GPER) on membrane of mouse oocytes during maturation. J. Assist. Reprod. Genet. 2013, 30, 227–232. [Google Scholar] [CrossRef]
- Pavlik, R.; Wypior, G.; Hecht, S.; Papadopoulos, P.; Kupka, M.; Thaler, C.; Wiest, I.; Pestka, A.; Friese, K.; Jeschke, U. Induction of G protein-coupled estrogen receptor (GPER) and nuclear steroid hormone receptors by gonadotropins in human granulosa cells. Histochem. Cell Biol. 2011, 136, 289–299. [Google Scholar] [CrossRef]
- Maiti, K.; Paul, J.W.; Read, M.; Chan, E.C.; Riley, S.C.; Nahar, P.; Smith, R. G-1-activated membrane estrogen receptors mediate increased contractility of the human myometrium. Endocrinology 2011, 152, 2448–2455. [Google Scholar] [CrossRef]
- Lauring, J.R.; Lehman, E.B.; Deimling, T.A.; Legro, R.S.; Chuang, C.H. Combined hormonal contraception use in reproductive-age women with contraindications to estrogen use. Am. J. Obstet. Gynecol. 2016, 215, 330.e1–330.e7. [Google Scholar] [CrossRef]
- Paton, D.M. Estetrol and drospirenone: A novel oral contraceptive. Drugs Today 2022, 58, 1–8. [Google Scholar] [CrossRef]
- Valéra, M.-C.; Noirrit-Esclassan, E.; Dupuis, M.; Fontaine, C.; Lenfant, F.; Briaux, A.; Cabou, C.; Garcia, C.; Lairez, O.; Foidart, J.-M.; et al. Effect of estetrol, a selective nuclear estrogen receptor modulator, in mouse models of arterial and venous thrombosis. Mol. Cell. Endocrinol. 2018, 477, 132–139. [Google Scholar] [CrossRef]
- Monteleone, P.; Mascagni, G.; Giannini, A.; Genazzani, A.R.; Simoncini, T. Symptoms of menopause—Global prevalence, physiology and implications. Nat. Rev. Endocrinol. 2018, 14, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Woods, N.F.; Mitchell, E.S. Symptoms during the perimenopause: Prevalence, severity, trajectory, and significance in women’s lives. Am. J. Med. 2005, 118, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Burger, H.G.; Hale, G.E.; Robertson, D.M.; Dennerstein, L. A review of hormonal changes during the menopausal transition: Focus on findings from the Melbourne Women’s Midlife Health Project. Hum. Reprod. Update 2007, 13, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.G.; Bromberger, J.T.; Buysse, D.J.; Hall, M.H.; Hardin, K.A.; Kravitz, H.M.; Matthews, K.A.; Rasor, M.O.; Utts, J.; Gold, E. Evaluation of the association of menopausal status with delta and beta EEG activity during sleep. Sleep 2011, 34, 1561–1568. [Google Scholar] [CrossRef]
- Rossmanith, W.G.; Ruebberdt, W. What causes hot flushes? The neuroendocrine origin of vasomotor symptoms in the menopause. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2009, 25, 303–314. [Google Scholar] [CrossRef]
- Modi, M.; Dhillo, W.S. The neuroendocrinology of the preoptic area in menopause: Symptoms and therapeutic strategies. Handb. Clin. Neurol. 2021, 179, 455–460. [Google Scholar] [CrossRef]
- Rometo, A.M.; Rance, N.E. Changes in prodynorphin gene expression and neuronal morphology in the hypothalamus of postmenopausal women. J. Neuroendocrinol. 2008, 20, 1376–1381. [Google Scholar] [CrossRef]
- Rometo, A.M.; Krajewski, S.J.; Voytko, M.L.; Rance, N.E. Hypertrophy and increased kisspeptin gene expression in the hypothalamic infundibular nucleus of postmenopausal women and ovariectomized monkeys. J. Clin. Endocrinol. Metab. 2007, 92, 2744–2750. [Google Scholar] [CrossRef]
- Sandoval-Guzmán, T.; Stalcup, S.T.; Krajewski, S.J.; Voytko, M.L.; Rance, N.E. Effects of ovariectomy on the neuroendocrine axes regulating reproduction and energy balance in young cynomolgus macaques. J. Neuroendocrinol. 2004, 16, 146–153. [Google Scholar] [CrossRef]
- Padilla, S.L.; Johnson, C.W.; Barker, F.D.; Patterson, M.A.; Palmiter, R.D. A Neural Circuit Underlying the Generation of Hot Flushes. Cell Rep. 2018, 24, 271–277. [Google Scholar] [CrossRef]
- Hansen, K.R.; Knowlton, N.S.; Thyer, A.C.; Charleston, J.S.; Soules, M.R.; Klein, N.A. A new model of reproductive aging: The decline in ovarian non-growing follicle number from birth to menopause. Hum. Reprod. 2008, 23, 699–708. [Google Scholar] [CrossRef]
- Rance, N.E.; Dacks, P.A.; Mittelman-Smith, M.A.; Romanovsky, A.A.; Krajewski-Hall, S.J. Modulation of body temperature and LH secretion by hypothalamic KNDy (kisspeptin, neurokinin B and dynorphin) neurons: A novel hypothesis on the mechanism of hot flushes. Front. Neuroendocrinol. 2013, 34, 211–227. [Google Scholar] [CrossRef] [PubMed]
- Brennan, A.; Hickey, M. Management of Early Menopause/Premature Ovarian Insufficiency in Women with or at High Risk of Breast Cancer. Semin. Reprod. Med. 2020, 38, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Del Pup, L.; Sánchez-Borrego, R. Ospemifene efficacy and safety data in women with vulvovaginal atrophy. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2020, 36, 569–577. [Google Scholar] [CrossRef]
- Di Donato, V.; Schiavi, M.C.; Iacobelli, V.; D’oria, O.; Kontopantelis, E.; Simoncini, T.; Muzii, L.; Benedetti Panici, P. Ospemifene for the treatment of vulvar and vaginal atrophy: A meta-analysis of randomized trials. Part II: Evaluation of tolerability and safety. Maturitas 2019, 121, 93–100. [Google Scholar] [CrossRef]
- Rattanatantikul, T.; Maiprasert, M.; Sugkraroek, P.; Bumrungpert, A. Efficacy and Safety of Nutraceutical on Menopausal Symptoms in Post-Menopausal Women: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Diet. Suppl. 2022, 19, 168–183. [Google Scholar] [CrossRef] [PubMed]
- Frigo, M.; de Barros, E.; Dos Santos, P.C.B.; Peres, G.L.; Weber, J.; Zanelatto, C.; Koehnlein, E.A. Effects of a Cereal Bar with a Combination of Phytoestrogens on the Climacteric Symptoms: A Placebo-Controlled, Randomized Trial. J. Am. Coll. Nutr. 2021, 41, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Dunneram, Y.; Chung, H.-F.; Cade, J.E.; Greenwood, D.C.; Dobson, A.J.; Mitchell, E.S.; Woods, N.F.; Brunner, E.J.; Yoshizawa, T.; Anderson, D.; et al. Soy intake and vasomotor menopausal symptoms among midlife women: A pooled analysis of five studies from the InterLACE consortium. Eur. J. Clin. Nutr. 2019, 73, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Bitto, A.; Arcoraci, V.; Alibrandi, A.; D’Anna, R.; Corrado, F.; Atteritano, M.; Minutoli, L.; Altavilla, D.; Squadrito, F. Visfatin correlates with hot flashes in postmenopausal women with metabolic syndrome: Effects of genistein. Endocrine 2017, 55, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, L.M.; Mazyck, P.J.; Hunter, M.H. Premenstrual syndrome. Am. Fam. Physician 2003, 67, 1743–1752. [Google Scholar] [PubMed]
- Dutta, A.; Sharma, A. Prevalence of premenstrual syndrome and premenstrual dysphoric disorder in India: A systematic review and meta-analysis. Health Promot. Perspect. 2021, 11, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Kuehner, C.; Nayman, S. Premenstrual Exacerbations of Mood Disorders: Findings and Knowledge Gaps. Curr. Psychiatry Rep. 2021, 23, 78. [Google Scholar] [CrossRef] [PubMed]
- Bucciarelli, V.; Nasi, M.; Bianco, F.; Seferovic, J.; Ivkovic, V.; Gallina, S.; Mattioli, A.V. Depression pandemic and cardiovascular risk in the COVID-19 era and long COVID syndrome: Gender makes a difference. Trends Cardiovasc. Med. 2022, 32, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.P.; Anderson, B.; Bharucha, A.E. Sex- and Gender-Related Differences in Common Functional Gastroenterologic Disorders. Mayo Clin. Proc. 2021, 96, 1071–1089. [Google Scholar] [CrossRef]
- Marver, J.E.; McGlinchey, E.A. Sex differences in insomnia and risk for psychopathology in adolescence. Curr. Opin. Psychol. 2020, 34, 63–67. [Google Scholar] [CrossRef]
- Price, M.E.; McCool, B.A. Structural, functional, and behavioral significance of sex and gonadal hormones in the basolateral amygdala: A review of preclinical literature. Alcohol 2022, 98, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.T.; Lichter, J.B.; Perry, A.N.; Castillo, S.A.; Keane, B.; Cushing, B.S.; Solomon, N.G. Medial amygdala ERα expression influences monogamous behaviour of male prairie voles in the field. Proc. Biol. Sci. 2021, 288, 20210318. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Clinard, C.T.; Dulka, B.N.; Grizzell, J.A.; Loewen, A.L.; Campbell, A.V.; Adler, S.G. Gonadal steroid hormone receptors in the medial amygdala contribute to experience-dependent changes in stress vulnerability. Psychoneuroendocrinology 2021, 129, 105249. [Google Scholar] [CrossRef] [PubMed]
- Rasia-Filho, A.A.; Dalpian, F.; Menezes, I.C.; Brusco, J.; Moreira, J.E.; Cohen, R.S. Dendritic spines of the medial amygdala: Plasticity, density, shape, and subcellular modulation by sex steroids. Histol. Histopathol. 2012, 27, 985–1011. [Google Scholar] [CrossRef]
- Gordon, J.L.; Girdler, S.S.; Meltzer-Brody, S.E.; Stika, C.S.; Thurston, R.C.; Clark, C.T.; Prairie, B.A.; Moses-Kolko, E.; Joffe, H.; Wisner, K.L. Ovarian hormone fluctuation, neurosteroids, and HPA axis dysregulation in perimenopausal depression: A novel heuristic model. Am. J. Psychiatry 2015, 172, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Montagne, A.; Zhao, Z. Alzheimer’s pathogenic mechanisms and underlying sex difference. Cell. Mol. Life Sci. 2021, 78, 4907–4920. [Google Scholar] [CrossRef]
- Genazzani, A.R.; Pluchino, N.; Luisi, S.; Luisi, M. Estrogen, cognition and female ageing. Hum. Reprod. Update 2007, 13, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Karkhaneh, A.; Ansari, M.; Emamgholipour, S.; Rafiee, M.H. The effect of 17β-estradiol on gene expression of calcitonin gene-related peptide and some pro-inflammatory mediators in peripheral blood mononuclear cells from patients with pure menstrual migraine. Iran. J. Basic Med. Sci. 2015, 18, 894–901. [Google Scholar] [PubMed]
- Wöber, C.; Brannath, W.; Schmidt, K.; Kapitan, M.; Rudel, E.; Wessely, P.; Wöber-Bingöl, C. Prospective analysis of factors related to migraine attacks: The PAMINA study. Cephalalgia 2007, 27, 304–314. [Google Scholar] [CrossRef]
- Azcoitia, I.; Yague, J.G.; Garcia-Segura, L.M. Estradiol synthesis within the human brain. Neuroscience 2011, 191, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Hojo, Y.; Kawato, S. Neurosteroids in adult hippocampus of male and female rodents: Biosynthesis and actions of sex steroids. Front. Endocrinol. 2018, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-G.; Wang, R.; Tang, H.; Dong, Y.; Chan, A.; Sareddy, G.R.; Vadlamudi, R.K.; Brann, D.W. Brain-derived estrogen exerts anti-inflammatory and neuroprotective actions in the rat hippocampus. Mol. Cell. Endocrinol. 2014, 389, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Caruso, D.; Pesaresi, M.; Abbiati, F.; Calabrese, D.; Giatti, S.; Garcia-Segura, L.M.; Melcangi, R.C. Comparison of plasma and cerebrospinal fluid levels of neuroactive steroids with their brain, spinal cord and peripheral nerve levels in male and female rats. Psychoneuroendocrinology 2013, 38, 2278–2290. [Google Scholar] [CrossRef]
- Fokidis, H.B.; Adomat, H.H.; Kharmate, G.; Hosseini-Beheshti, E.; Guns, E.S.; Soma, K.K. Regulation of local steroidogenesis in the brain and in prostate cancer: Lessons learned from interdisciplinary collaboration. Front. Neuroendocrinol. 2015, 36, 108–129. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Zhang, Q.; Tang, F.L.; Pratap, U.P.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Neuron-Derived Estrogen Is Critical for Astrocyte Activation and Neuroprotection of the Ischemic Brain. J. Neurosci. 2020, 40, 7355–7374. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Dubal, D.B.; Zhu, H.; Yu, J.; Rau, S.W.; Shughrue, P.J.; Merchenthaler, I.; Kindy, M.S.; Wise, P.M. Estrogen receptor alpha, not beta, is a critical link in estradiol-mediated protection against brain injury. Proc. Natl. Acad. Sci. USA 2001, 98, 1952–1957. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, L.; Colón, J.M.; Santiago, J.M.; Torrado, A.I.; Meléndez, M.; Segarra, A.C.; Rodríguez-Orengo, J.F.; Miranda, J.D. Tamoxifen and estradiol improved locomotor function and increased spared tissue in rats after spinal cord injury: Their antioxidant effect and role of estrogen receptor alpha. Brain Res. 2014, 1561, 11–22. [Google Scholar] [CrossRef]
- Sellers, K.J.; Erli, F.; Raval, P.; Watson, I.A.; Chen, D.; Srivastava, D.P. Rapid modulation of synaptogenesis and spinogenesis by 17β-estradiol in primary cortical neurons. Front. Cell. Neurosci. 2015, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, Q.; Yu, C.-Y.; Wang, F.; Shao, Y.; Sun, K.-S.; Sun, T.; Liu, J. G Protein-Coupled Estrogen Receptor 1 Knockout Deteriorates MK-801-Induced Learning and Memory Impairment in Mice. Front. Behav. Neurosci. 2020, 14, 157. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Varma, A.; Barry, J.; Vertegel, A.; Banik, N. Nanoparticle Estrogen in Rat Spinal Cord Injury Elicits Rapid Anti-Inflammatory Effects in Plasma, Cerebrospinal Fluid, and Tissue. J. Neurotrauma 2015, 32, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Khaksari, M.; Abbasloo, E.; Dehghan, F.; Soltani, Z.; Asadikaram, G. The brain cytokine levels are modulated by estrogen following traumatic brain injury: Which estrogen receptor serves as modulator? Int. Immunopharmacol. 2015, 28, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Choi, H.Y.; Na, W.H.; Ju, B.G.; Yune, T.Y. 17β-estradiol inhibits MMP-9 and SUR1/TrpM4 expression and activation and thereby attenuates BSCB disruption/hemorrhage after spinal cord injury in male rats. Endocrinology 2015, 156, 1838–1850. [Google Scholar] [CrossRef] [PubMed]
- Soltani, Z.; Khaksari, M.; Jafari, E.; Iranpour, M.; Shahrokhi, N. Is genistein neuroprotective in traumatic brain injury? Physiol. Behav. 2015, 152, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Khalil, R.A. Matrix Metalloproteinases in Normal Pregnancy and Preeclampsia. Prog. Mol. Biol. Transl. Sci. 2017, 148, 87–165. [Google Scholar] [CrossRef]
- Turner, R.J.; Sharp, F.R. Implications of MMP9 for Blood Brain Barrier Disruption and Hemorrhagic Transformation Following Ischemic Stroke. Front. Cell. Neurosci. 2016, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.J.; Rønnekleiv, O.K. Control of CNS neuronal excitability by estrogens via membrane-initiated signaling. Mol. Cell. Endocrinol. 2009, 308, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, Q.; Hu, F.; Wang, J.; Ding, H.; Gao, R.; Xiao, H.; Wang, L. Effects of estradiol on voltage-gated potassium channels in mouse dorsal root ganglion neurons. J. Membr. Biol. 2014, 247, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Meseke, M.; Neumüller, F.; Brunne, B.; Li, X.; Anstötz, M.; Pohlkamp, T.; Rogalla, M.M.; Herz, J.; Rune, G.M.; Bender, R.A. Distal Dendritic Enrichment of HCN1 Channels in Hippocampal CA1 Is Promoted by Estrogen, but Does Not Require Reelin. eNeuro 2018, 5, ENEURO.0258-18.2018. [Google Scholar] [CrossRef]
- Fester, L.; Prange-Kiel, J.; Zhou, L.; Blittersdorf, B.V.; Böhm, J.; Jarry, H.; Schumacher, M.; Rune, G.M. Estrogen-regulated synaptogenesis in the hippocampus: Sexual dimorphism in vivo but not in vitro. J. Steroid Biochem. Mol. Biol. 2012, 131, 24–29. [Google Scholar] [CrossRef]
- Ye, Z.; Cudmore, R.H.; Linden, D.J. Estrogen-Dependent Functional Spine Dynamics in Neocortical Pyramidal Neurons of the Mouse. J. Neurosci. 2019, 39, 4874–4888. [Google Scholar] [CrossRef]
- Bailey, M.E.; Wang, A.C.J.; Hao, J.; Janssen, W.G.M.; Hara, Y.; Dumitriu, D.; Hof, P.R.; Morrison, J.H. Interactive effects of age and estrogen on cortical neurons: Implications for cognitive aging. Neuroscience 2011, 191, 148–158. [Google Scholar] [CrossRef]
- Chen, J.-R.; Yan, Y.-T.; Wang, T.-J.; Chen, L.-J.; Wang, Y.-J.; Tseng, G.-F. Gonadal hormones modulate the dendritic spine densities of primary cortical pyramidal neurons in adult female rat. Cereb. Cortex 2009, 19, 2719–2727. [Google Scholar] [CrossRef]
- Batallán Burrowes, A.A.; Sundarakrishnan, A.; Bouhour, C.; Chapman, C.A. G protein-coupled estrogen receptor-1 enhances excitatory synaptic responses in the entorhinal cortex. Hippocampus 2021, 31, 1191–1201. [Google Scholar] [CrossRef]
- Huang, G.Z.; Woolley, C.S. Estradiol acutely suppresses inhibition in the hippocampus through a sex-specific endocannabinoid and mGluR-dependent mechanism. Neuron 2012, 74, 801–808. [Google Scholar] [CrossRef]
- Cersosimo, M.G.; Benarroch, E.E. Estrogen actions in the nervous system: Complexity and clinical implications. Neurology 2015, 85, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Godó, S.; Barabás, K.; Lengyel, F.; Ernszt, D.; Kovács, T.; Kecskés, M.; Varga, C.; Jánosi, T.Z.; Makkai, G.; Kovács, G.; et al. Single-Molecule Imaging Reveals Rapid Estradiol Action on the Surface Movement of AMPA Receptors in Live Neurons. Front. Cell Dev. Biol. 2021, 9, 708715. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, D.H.; Zhang, S.; Gan, W.-B. Dendritic spine dynamics. Annu. Rev. Physiol. 2009, 71, 261–282. [Google Scholar] [CrossRef] [PubMed]
- Kramár, E.A.; Babayan, A.H.; Gall, C.M.; Lynch, G. Estrogen promotes learning-related plasticity by modifying the synaptic cytoskeleton. Neuroscience 2013, 239, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Al Sweidi, S.; Sanchez, M.G.; Bourque, M.; Morissette, M.; Dluzen, D.; Di Paolo, T. Oestrogen receptors and signalling pathways: Implications for neuroprotective effects of sex steroids in Parkinson’s disease. J. Neuroendocr. 2012, 24, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Barreto, G.E.; McGovern, A.J.; Garcia-Segura, L.M. Role of Neuroglobin in the Neuroprotective Actions of Estradiol and Estrogenic Compounds. Cells 2021, 10, 1907. [Google Scholar] [CrossRef]
- Exertier, C.; Montemiglio, L.C.; Freda, I.; Gugole, E.; Parisi, G.; Savino, C.; Vallone, B. Neuroglobin, clues to function and mechanism. Mol. Asp. Med. 2022, 84, 101055. [Google Scholar] [CrossRef]
- Khombi Shooshtari, M.; Farbood, Y.; Mansouri, S.M.T.; Badavi, M.; Khorsandi, L.S.; Ghasemi Dehcheshmeh, M.; Sarkaki, A.R. Neuroprotective Effects of Chrysin Mediated by Estrogenic Receptors Following Cerebral Ischemia and Reperfusion in Male Rats. Basic Clin. Neurosci. 2021, 12, 149–162. [Google Scholar] [CrossRef]
- Finney, C.A.; Shvetcov, A.; Westbrook, R.F.; Morris, M.J.; Jones, N.M. Tamoxifen offers long-term neuroprotection after hippocampal silent infarct in male rats. Horm. Behav. 2021, 136, 105085. [Google Scholar] [CrossRef]
- Finney, C.A.; Shvetcov, A.; Westbrook, R.F.; Morris, M.J.; Jones, N.M. The selective estrogen receptor modulator tamoxifen protects against subtle cognitive decline and early markers of injury 24 h after hippocampal silent infarct in male Sprague-Dawley rats. Horm. Behav. 2021, 134, 105016. [Google Scholar] [CrossRef]
- Brinton, R.D. Estrogen-induced plasticity from cells to circuits: Predictions for cognitive function. Trends Pharmacol. Sci. 2009, 30, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Galea, L.A.M.; Uban, K.A.; Epp, J.R.; Brummelte, S.; Barha, C.K.; Wilson, W.L.; Lieblich, S.E.; Pawluski, J.L. Endocrine regulation of cognition and neuroplasticity: Our pursuit to unveil the complex interaction between hormones, the brain, and behaviour. Can. J. Exp. Psychol. Can. Psychol. Expérimentale 2008, 62, 247. [Google Scholar] [CrossRef] [PubMed]
- Guetta, V.; Quyyumi, A.A.; Prasad, A.; Panza, J.A.; Waclawiw, M.; Cannon, R.O. 3rd The role of nitric oxide in coronary vascular effects of estrogen in postmenopausal women. Circulation 1997, 96, 2795–2801. [Google Scholar] [CrossRef] [PubMed]
- Barrett-Connor, E.; Bush, T.L. Estrogen and coronary heart disease in women. JAMA 1991, 265, 1861–1867. [Google Scholar] [CrossRef] [PubMed]
- Guetta, V.; Cannon, R.O. 3rd Cardiovascular effects of estrogen and lipid-lowering therapies in postmenopausal women. Circulation 1996, 93, 1928–1937. [Google Scholar] [CrossRef]
- Grodstein, F.; Stampfer, M.J.; Manson, J.E.; Colditz, G.A.; Willett, W.C.; Rosner, B.; Speizer, F.E.; Hennekens, C.H. Postmenopausal estrogen and progestin use and the risk of cardiovascular disease. N. Engl. J. Med. 1996, 335, 453–461. [Google Scholar] [CrossRef]
- Stevenson, J.C.; Tsiligiannis, S.; Panay, N. Cardiovascular Risk in Perimenopausal Women. Curr. Vasc. Pharmacol. 2019, 17, 591–594. [Google Scholar] [CrossRef]
- Williams, J.K.; Adams, M.R.; Herrington, D.M.; Clarkson, T.B. Short-term administration of estrogen and vascular responses of atherosclerotic coronary arteries. J. Am. Coll. Cardiol. 1992, 20, 452–457. [Google Scholar] [CrossRef][Green Version]
- Venkov, C.D.; Rankin, A.B.; Vaughan, D.E. Identification of authentic estrogen receptor in cultured endothelial cells. A potential mechanism for steroid hormone regulation of endothelial function. Circulation 1996, 94, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Losordo, D.W.; Kearney, M.; Kim, E.A.; Jekanowski, J.; Isner, J.M. Variable expression of the estrogen receptor in normal and atherosclerotic coronary arteries of premenopausal women. Circulation 1994, 89, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Pare, G.; Krust, A.; Karas, R.H.; Dupont, S.; Aronovitz, M.; Chambon, P.; Mendelsohn, M.E. Estrogen receptor-alpha mediates the protective effects of estrogen against vascular injury. Circ. Res. 2002, 90, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Zancan, V.; Santagati, S.; Bolego, C.; Vegeto, E.; Maggi, A.; Puglisi, L. 17Beta-estradiol decreases nitric oxide synthase II synthesis in vascular smooth muscle cells. Endocrinology 1999, 140, 2004–2009. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maggi, A.; Cignarella, A.; Brusadelli, A.; Bolego, C.; Pinna, C.; Puglisi, L. Diabetes undermines estrogen control of inducible nitric oxide synthase function in rat aortic smooth muscle cells through overexpression of estrogen receptor-beta. Circulation 2003, 108, 211–217. [Google Scholar] [CrossRef]
- Hodgin, J.B.; Krege, J.H.; Reddick, R.L.; Korach, K.S.; Smithies, O.; Maeda, N. Estrogen receptor alpha is a major mediator of 17beta-estradiol’s atheroprotective effects on lesion size in Apoe-/- mice. J. Clin. Investig. 2001, 107, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.R.; Roy, T.; Cox, B.E. Mechanisms modulating estrogen-induced uterine vasodilation. Vascul. Pharmacol. 2002, 38, 115–125. [Google Scholar] [CrossRef]
- Magness, R.R.; Phernetton, T.M.; Zheng, J. Systemic and uterine blood flow distribution during prolonged infusion of 17beta-estradiol. Am. J. Physiol. 1998, 275, H731-43. [Google Scholar] [CrossRef] [PubMed]
- Critchley, H.O.; Brenner, R.M.; Henderson, T.A.; Williams, K.; Nayak, N.R.; Slayden, O.D.; Millar, M.R.; Saunders, P.T. Estrogen receptor beta, but not estrogen receptor alpha, is present in the vascular endothelium of the human and nonhuman primate endometrium. J. Clin. Endocrinol. Metab. 2001, 86, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Jobe, S.O.; Ramadoss, J.; Koch, J.M.; Jiang, Y.; Zheng, J.; Magness, R.R. Estradiol-17beta and its cytochrome P450- and catechol-O-methyltransferase-derived metabolites stimulate proliferation in uterine artery endothelial cells: Role of estrogen receptor-alpha versus estrogen receptor-beta. Hypertension 2010, 55, 1005–1011. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, L.; Wang, W.; Magness, R.R.; Chen, D. Estrogen-responsive nitroso-proteome in uterine artery endothelial cells: Role of endothelial nitric oxide synthase and estrogen receptor-β. J. Cell. Physiol. 2012, 227, 146–159. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lindner, V.; Kim, S.K.; Karas, R.H.; Kuiper, G.G.; Gustafsson, J.A.; Mendelsohn, M.E. Increased expression of estrogen receptor-beta mRNA in male blood vessels after vascular injury. Circ. Res. 1998, 83, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Darblade, B.; Pendaries, C.; Krust, A.; Dupont, S.; Fouque, M.-J.; Rami, J.; Chambon, P.; Bayard, F.; Arnal, J.-F. Estradiol alters nitric oxide production in the mouse aorta through the alpha-, but not beta-, estrogen receptor. Circ. Res. 2002, 90, 413–419. [Google Scholar] [CrossRef]
- Zuloaga, K.L.; O’Connor, D.T.; Handa, R.J.; Gonzales, R.J. Estrogen receptor beta dependent attenuation of cytokine-induced cyclooxygenase-2 by androgens in human brain vascular smooth muscle cells and rat mesenteric arteries. Steroids 2012, 77, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Park, J.; Shim, M.K.; Rhyu, M.-R.; Yoon, B.-K.; Kim, K.S.; Lee, Y. Indazole-Cl inhibits hypoxia-induced cyclooxygenase-2 expression in vascular smooth muscle cells. J. Mol. Endocrinol. 2019, 63, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, X.; Cong, B.; You, X.; Wang, Y.; Wang, W.; Ni, X. Estrogenic action on arterial smooth muscle: Permissive for maintenance of CRHR2 expression. Endocrinology 2012, 153, 1915–1924. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.; Chen, Y.-X.; Siebert, T.; O’Brien, E.R. Heat shock protein 27: Clue to understanding estrogen-mediated atheroprotection? Trends Cardiovasc. Med. 2010, 20, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Pastore, M.B.; Jobe, S.O.; Ramadoss, J.; Magness, R.R. Estrogen receptor-α and estrogen receptor-β in the uterine vascular endothelium during pregnancy: Functional implications for regulating uterine blood flow. Semin. Reprod. Med. 2012, 30, 46–61. [Google Scholar] [CrossRef]
- Morales, D.E.; McGowan, K.A.; Grant, D.S.; Maheshwari, S.; Bhartiya, D.; Cid, M.C.; Kleinman, H.K.; Schnaper, H.W. Estrogen promotes angiogenic activity in human umbilical vein endothelial cells in vitro and in a murine model. Circulation 1995, 91, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Magness, R.R.; Phernetton, T.M.; Gibson, T.C.; Chen, D.-B. Uterine blood flow responses to ICI 182 780 in ovariectomized oestradiol-17beta-treated, intact follicular and pregnant sheep. J. Physiol. 2005, 565, 71–83. [Google Scholar] [CrossRef]
- Kim, J.G.; Leem, Y.-E.; Kwon, I.; Kang, J.-S.; Bae, Y.M.; Cho, H. Estrogen modulates serotonin effects on vasoconstriction through Src inhibition. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cairrão, E.; Alvarez, E.; Carvas, J.M.; Santos-Silva, A.J.; Verde, I. Non-genomic vasorelaxant effects of 17β-estradiol and progesterone in rat aorta are mediated by L-type Ca2+ current inhibition. Acta Pharmacol. Sin. 2012, 33, 615–624. [Google Scholar] [CrossRef]
- Geary, G.G.; Krause, D.N.; Duckles, S.P. Estrogen reduces mouse cerebral artery tone through endothelial NOS- and cyclooxygenase-dependent mechanisms. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H511-9. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.; Shay, J.; Jiang, C.; Moss, J. Nitric oxide accounts for dose-dependent estrogen-mediated coronary relaxation after acute estrogen withdrawal. Circulation 1994, 90, 1964–1968. [Google Scholar] [CrossRef] [PubMed]
- Salom, J.B.; Burguete, M.C.; Pérez-Asensio, F.J.; Centeno, J.M.; Torregrosa, G.; Alborch, E. Acute relaxant effects of 17-beta-estradiol through non-genomic mechanisms in rabbit carotid artery. Steroids 2002, 67, 339–346. [Google Scholar] [CrossRef]
- Goetz, R.M.; Thatte, H.S.; Prabhakar, P.; Cho, M.R.; Michel, T.; Golan, D.E. Estradiol induces the calcium-dependent translocation of endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1999, 96, 2788–2793. [Google Scholar] [CrossRef] [PubMed]
- Nechmad, A.; Merin, G.; Schwalb, H.; Shimon, D.V.; Borman, J.B.; Milgalter, E.; Mosseri, M. Estrogen induces nitric oxide-mediated vasodilation of human mammary arteries in vitro. Nitric Oxide Biol. Chem. 1998, 2, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Scheidegger, K.J.; Cenni, B.; Picard, D.; Delafontaine, P. Estradiol decreases IGF-1 and IGF-1 receptor expression in rat aortic smooth muscle cells. Mechanisms for its atheroprotective effects. J. Biol. Chem. 2000, 275, 38921–38928. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Thomas, P. Additive effects of low concentrations of estradiol-17β and progesterone on nitric oxide production by human vascular endothelial cells through shared signaling pathways. J. Steroid Biochem. Mol. Biol. 2017, 165, 258–267. [Google Scholar] [CrossRef]
- Mirdamadi, M.; Kothencz, A.; Szűcs, E.; Benyhe, S.; Szécsi, M.; Gáspár, R. Non-genomic actions of sex hormones on pregnant uterine contractility in rats: An in vitro study at term. Life Sci. 2020, 263, 118584. [Google Scholar] [CrossRef]
- Grodstein, F.; Manson, J.E.; Colditz, G.A.; Willett, W.C.; Speizer, F.E.; Stampfer, M.J. A prospective, observational study of postmenopausal hormone therapy and primary prevention of cardiovascular disease. Ann. Intern. Med. 2000, 133, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Hsia, J.; Johnson, K.C.; Rossouw, J.E.; Assaf, A.R.; Lasser, N.L.; Trevisan, M.; Black, H.R.; Heckbert, S.R.; Detrano, R.; et al. Estrogen plus progestin and the risk of coronary heart disease. N. Engl. J. Med. 2003, 349, 523–534. [Google Scholar] [CrossRef]
- Hulley, S.; Furberg, C.; Barrett-Connor, E.; Cauley, J.; Grady, D.; Haskell, W.; Knopp, R.; Lowery, M.; Satterfield, S.; Schrott, H.; et al. Noncardiovascular disease outcomes during 6.8 years of hormone therapy: Heart and Estrogen/progestin Replacement Study follow-up (HERS II). JAMA 2002, 288, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Roa-Díaz, Z.M.; Raguindin, P.F.; Bano, A.; Laine, J.E.; Muka, T.; Glisic, M. Menopause and cardiometabolic diseases: What we (don’t) know and why it matters. Maturitas 2021, 152, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.G.; Anekwe, C.; Washington, K.; Akam, E.Y.; Wang, E.; Stanford, F.C. Weight regulation in menopause. Menopause 2021, 28, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Sutton-Tyrrell, K.; Zhao, X.; Santoro, N.; Lasley, B.; Sowers, M.; Johnston, J.; Mackey, R.; Matthews, K. Reproductive hormones and obesity: 9 years of observation from the Study of Women’s Health Across the Nation. Am. J. Epidemiol. 2010, 171, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Janssen, I.; Powell, L.H.; Kazlauskaite, R.; Dugan, S.A. Testosterone and visceral fat in midlife women: The Study of Women’s Health Across the Nation (SWAN) fat patterning study. Obesity 2010, 18, 604–610. [Google Scholar] [CrossRef]
- Leeners, B.; Geary, N.; Tobler, P.N.; Asarian, L. Ovarian hormones and obesity. Hum. Reprod. Update 2017, 23, 300–321. [Google Scholar] [CrossRef]
- Rooyackers, O.E.; Nair, K.S. Hormonal regulation of human muscle protein metabolism. Annu. Rev. Nutr. 1997, 17, 457–485. [Google Scholar] [CrossRef]
- Török, B.; Csikota, P.; Fodor, A.; Balázsfi, D.; Ferenczi, S.; Demeter, K.; Tóth, Z.E.; Könczöl, K.; Perna, J.C.; Farkas, I.; et al. Rescue of Vasopressin Synthesis in Magnocellular Neurons of the Supraoptic Nucleus Normalises Acute Stress-Induced Adrenocorticotropin Secretion and Unmasks an Effect on Social Behaviour in Male Vasopressin-Deficient Brattleboro Rats. Int. J. Mol. Sci. 2022, 23, 1357. [Google Scholar] [CrossRef]
- Alemany, M. Estrogens and the regulation of glucose metabolism. World J. Diabetes 2021, 12, 1622–1654. [Google Scholar] [CrossRef]
- Shibata, Y.; Shiga, Y.; Suematsu, Y.; Tashiro, K.; Kawahira, Y.; Morita, K.; Kuwano, T.; Sugihara, M.; Miura, S.-I. Association Between Major Adverse Cardiovascular Events and the Ratio of Subcutaneous Fat Area to Visceral Fat Area in Patients Who Have Undergone Multidetector Row Computed Tomography. Circ. Rep. 2021, 3, 674–681. [Google Scholar] [CrossRef]
- Xu, B.; Lovre, D.; Mauvais-Jarvis, F. Effect of selective estrogen receptor modulators on metabolic homeostasis. Biochimie 2016, 124, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.N.; Khaksari, M.; Sepehri, G.; Karam, G.A.; Raji-Amirhasani, A.; Azizian, H. The effects of alone and combination tamoxifen, raloxifene and estrogen on lipid profile and atherogenic index of ovariectomized type 2 diabetic rats. Life Sci. 2020, 263, 118573. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, E.L.; Shearin, J.; Koegler, F.H.; Cameron, J.L. Selective estrogen receptor modulator promotes weight loss in ovariectomized female rhesus monkeys (Macaca mulatta) by decreasing food intake and increasing activity. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E759-67. [Google Scholar] [CrossRef]
- Romani, A.M.P. The controversy on the beneficial effect of phytoestrogens in diabetic treatment in postmenopausal women. Biochem. Pharmacol. 2021, 190, 114619. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Franke, A.A.; Rosner, B.A.; Giovannucci, E.; van Dam, R.M.; Tworoger, S.S.; Hu, F.B.; Sun, Q. Urinary isoflavonoids and risk of type 2 diabetes: A prospective investigation in US women. Br. J. Nutr. 2015, 114, 1694–1701. [Google Scholar] [CrossRef][Green Version]
- Lephart, E.D.; Naftolin, F. Menopause and the Skin: Old Favorites and New Innovations in Cosmeceuticals for Estrogen-Deficient Skin. Dermatol. Ther. 2021, 11, 53–69. [Google Scholar] [CrossRef]
- Calleja-Agius, J.; Brincat, M. The effect of menopause on the skin and other connective tissues. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2012, 28, 273–277. [Google Scholar] [CrossRef]
- Patriarca, M.T.; de Moraes, A.R.B.; Nader, H.B.; Petri, V.; Martins, J.R.M.; Gomes, R.C.T.; Soares, J.M.J. Hyaluronic acid concentration in postmenopausal facial skin after topical estradiol and genistein treatment: A double-blind, randomized clinical trial of efficacy. Menopause 2013, 20, 336–341. [Google Scholar] [CrossRef]
- Nanashima, N.; Horie, K.; Maeda, H.; Tomisawa, T.; Kitajima, M.; Nakamura, T. Blackcurrant Anthocyanins Increase the Levels of Collagen, Elastin, and Hyaluronic Acid in Human Skin Fibroblasts and Ovariectomized Rats. Nutrients 2018, 10, 495. [Google Scholar] [CrossRef]
- Pomari, E.; Dalla Valle, L.; Pertile, P.; Colombo, L.; Thornton, M.J. Intracrine sex steroid synthesis and signaling in human epidermal keratinocytes and dermal fibroblasts. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 508–524. [Google Scholar] [CrossRef]
- Briggs, P. Consensus statement—Urogenital atrophy. Post Reprod. Health 2022, 28, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Pingarron, C.; de Lafuente, P.; Ierullo, A.M.; Poyo Torcal, S.; Maroto Díaz, C.J.; Palacios, S. Ospemifene in clinical practice for vulvo-vaginal atrophy: Results at 3 months of follow-up of use. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2021, 37, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Archer, D.F. Ospemifene: Less venous thrombosis than other selective estrogen receptor modulators in postmenopausal women with vulvo vaginal atrophy. Menopause 2020, 27, 846–847. [Google Scholar] [CrossRef] [PubMed]
- Wiik, A.; Ekman, M.; Johansson, O.; Jansson, E.; Esbjörnsson, M. Expression of both oestrogen receptor alpha and beta in human skeletal muscle tissue. Histochem. Cell Biol. 2009, 131, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Lowe, D.A.; Baltgalvis, K.A.; Greising, S.M. Mechanisms behind estrogen’s beneficial effect on muscle strength in females. Exerc. Sport Sci. Rev. 2010, 38, 61–67. [Google Scholar] [CrossRef]
- Cosma, M.; Bailey, J.; Miles, J.M.; Bowers, C.Y.; Veldhuis, J.D. Pituitary and/or peripheral estrogen-receptor alpha regulates follicle-stimulating hormone secretion, whereas central estrogenic pathways direct growth hormone and prolactin secretion in postmenopausal women. J. Clin. Endocrinol. Metab. 2008, 93, 951–958. [Google Scholar] [CrossRef]
- McMillin, S.L.; Minchew, E.C.; Lowe, D.A.; Spangenburg, E.E. Skeletal muscle wasting: The estrogen side of sexual dimorphism. Am. J. Physiol. Cell Physiol. 2022, 322, C24–C37. [Google Scholar] [CrossRef]
- Lee, S.; Kim, M.; Hong, S.; Kim, E.J.; Kim, J.H.; Sohn, Y.; Jung, H.S. Effects of Sparganii Rhizoma on Osteoclast Formation and Osteoblast Differentiation and on an OVX-Induced Bone Loss Model. Front. Pharmacol. 2022, 12, 797892. [Google Scholar] [CrossRef]
- Crandall, C.J.; Larson, J.C.; Lacroix, A.Z.; Robbins, J.A.; Wactawski-Wende, J.; Johnson, K.C.; Sattari, M.; Stone, K.L.; Weitlauf, J.C.; Gure, T.R.; et al. Risk of Subsequent Fractures in Postmenopausal Women After Nontraumatic vs Traumatic Fractures. JAMA Intern. Med. 2021, 181, 1055–1063. [Google Scholar] [CrossRef]
- Gustafsson, K.L.; Farman, H.; Henning, P.; Lionikaite, V.; Movérare-Skrtic, S.; Wu, J.; Ryberg, H.; Koskela, A.; Gustafsson, J.-Å.; Tuukkanen, J.; et al. The role of membrane ERα signaling in bone and other major estrogen responsive tissues. Sci. Rep. 2016, 6, 29473. [Google Scholar] [CrossRef]
- Clowes, J.A.; Riggs, B.L.; Khosla, S. The role of the immune system in the pathophysiology of osteoporosis. Immunol. Rev. 2005, 208, 207–227. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C.; O’Brien, C.A.; Almeida, M. The role of estrogen and androgen receptors in bone health and disease. Nat. Rev. Endocrinol. 2013, 9, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Ge, F.; Che, Y.; Wu, S.; Dong, X.; Song, D. Metabolomics coupled with pathway analysis provides insights into sarco-osteoporosis metabolic alterations and estrogen therapeutic effects in mice. Biomolecules 2022, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, J.E.; Anderson, G.L.; Prentice, R.L.; LaCroix, A.Z.; Kooperberg, C.; Stefanick, M.L.; Jackson, R.D.; Beresford, S.A.A.; Howard, B.V.; Johnson, K.C.; et al. Risks and benefits of estrogen plus progestin in healthy postmenopausal women: Principal results from the women’s health initiative randomized controlled trial. JAMA 2002, 288, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Liedert, A.; Nemitz, C.; Haffner-Luntzer, M.; Schick, F.; Jakob, F.; Ignatius, A. Effects of Estrogen Receptor and Wnt Signaling Activation on Mechanically Induced Bone Formation in a Mouse Model of Postmenopausal Bone Loss. Int. J. Mol. Sci. 2020, 21, 8301. [Google Scholar] [CrossRef]
- Jackson, R.L.; Greiwe, J.S.; Schwen, R.J. Emerging evidence of the health benefits of S-equol, an estrogen receptor β agonist. Nutr. Rev. 2011, 69, 432–448. [Google Scholar] [CrossRef]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and Androgens in Skeletal Physiology and Pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef]
- Kousteni, S.; Bellido, T.; Plotkin, L.I.; O’Brien, C.A.; Bodenner, D.L.; Han, L.; Han, K.; DiGregorio, G.B.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S.; et al. Nongenotropic, sex-nonspecific signaling through the estrogen or androgen receptors: Dissociation from transcriptional activity. Cell 2001, 104, 719–730. [Google Scholar] [CrossRef]
- Khosla, S.; Oursler, J.; Monroe, D.G. Estrogen and the Skeleton. Trends Endocrinol. Metab. 2012, 23, 576–581. [Google Scholar] [CrossRef]
- Streicher, C.; Heyny, A.; Andrukhova, O.; Haigl, B.; Slavic, S.; Schuler, C.; Kollmann, K.; Kantner, I.; Sexl, V.; Kleiter, M.; et al. Estrogen Regulates Bone Turnover by Targeting RANKL Expression in Bone Lining Cells. Sci. Rep. 2017, 7, 6460. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Schoppet, M. Clinical implications of the osteoprotegerin/RANKL/RANK system for bone and vascular diseases. JAMA 2004, 292, 490–495. [Google Scholar] [CrossRef]
- Föger-Samwald, U.; Dovjak, P.; Azizi-Semrad, U.; Kerschan-Schindl, K.; Pietschmann, P. Osteoporosis: Pathophysiology and therapeutic options. EXCLI J. 2020, 19, 1017. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.Y.; Hilal, A.; Maini, I. What traditional, complementary, and integrative medicine recommendations exist across osteoporosis clinical practice guidelines? A systematic review and quality assessment. Integr. Med. Res. 2022, 11, 100803. [Google Scholar] [CrossRef] [PubMed]
- Palacios, S. Medical treatment of osteoporosis. Climacteric 2022, 25, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jeong, H.E.; Baek, Y.-H.; Cho, S.W.; Lim, H.; Shin, J.-Y. Treatment pattern in postmenopausal women with osteoporosis: A population-based cohort study in South Korea. J. Bone Miner. Metab. 2022, 40, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Fine, A.; Busza, A.; Allen, L.M.; Kelly, C.; Wolfman, W.; Jacobson, M.; Lega, I.C. Comparing estrogen-based hormonal contraceptives and hormone therapy on bone mineral density in women with premature ovarian insufficiency: A systematic review. Menopause 2022, 29, 351–359. [Google Scholar] [CrossRef]
- Gu, Y.; Zhou, J.; Wang, Q.; Fan, W.; Yin, G. Ginsenoside Rg1 promotes osteogenic differentiation of rBMSCs and healing of rat tibial fractures through regulation of GR-dependent BMP-2/SMAD signaling. Sci. Rep. 2016, 6, 25282. [Google Scholar] [CrossRef]
- Khosla, S. Update on Estrogens and the Skeleton. J. Clin. Endocrinol. Metab. 2010, 95, 3569. [Google Scholar] [CrossRef]
- Bustamante-Barrientos, F.A.; Méndez-Ruette, M.; Ortloff, A.; Luz-Crawford, P.; Rivera, F.J.; Figueroa, C.D.; Molina, L.; Bátiz, L.F. The Impact of Estrogen and Estrogen-Like Molecules in Neurogenesis and Neurodegeneration: Beneficial or Harmful? Front. Cell. Neurosci. 2021, 15, 636176. [Google Scholar] [CrossRef] [PubMed]
- Folkerd, E.J.; Dowsett, M. Influence of sex hormones on cancer progression. J. Clin. Oncol. 2010, 28, 4038–4044. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Fry, E.A. Aberrant Splicing of Estrogen Receptor, HER2, and CD44 Genes in Breast Cancer. Genet. Epigenet. 2015, 7, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Matsushima-Nishiwaki, R.; Yamada, N.; Hattori, Y.; Hosokawa, Y.; Tachi, J.; Hori, T.; Kozawa, O. SERMs (selective estrogen receptor modulator), acting as estrogen receptor β agonists in hepatocellular carcinoma cells, inhibit the transforming growth factor-α-induced migration via specific inhibition of AKT signaling pathway. PLoS ONE 2022, 17, e0262485. [Google Scholar] [CrossRef] [PubMed]
- Hernando, C.; Ortega-Morillo, B.; Tapia, M.; Moragón, S.; Martínez, M.T.; Eroles, P.; Garrido-Cano, I.; Adam-Artigues, A.; Lluch, A.; Bermejo, B.; et al. Oral Selective Estrogen Receptor Degraders (SERDs) as a Novel Breast Cancer Therapy: Present and Future from a Clinical Perspective. Int. J. Mol. Sci. 2021, 22, 7812. [Google Scholar] [CrossRef] [PubMed]
- Louie, M.C.; Sevigny, M.B. Steroid hormone receptors as prognostic markers in breast cancer. Am. J. Cancer Res. 2017, 7, 1617. [Google Scholar] [PubMed]
- Brufsky, A.M.; Dickler, M.N. Estrogen Receptor-Positive Breast Cancer: Exploiting Signaling Pathways Implicated in Endocrine Resistance. Oncologist 2018, 23, 528–539. [Google Scholar] [CrossRef]
- Hua, H.; Zhang, H.; Kong, Q.; Jiang, Y. Mechanisms for estrogen receptor expression in human cancer. Exp. Hematol. Oncol. 2018, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Belachew, E.B.; Sewasew, D.T. Molecular Mechanisms of Endocrine Resistance in Estrogen-Positive Breast Cancer. Front. Endocrinol. 2021, 12, 599586. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Oesterreich, S.; Chamness, G.C.; McGuire, W.L.; Fuqua, S.A. Biological and clinical implications of heat shock protein 27,000 (Hsp27): A review. J. Natl. Cancer Inst. 1993, 85, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Osborne, C.K.; Wakeling, A.; Nicholson, R.I. Fulvestrant: An oestrogen receptor antagonist with a novel mechanism of action. Br. J. Cancer 2004, 90 (Suppl. 1), S2–S6. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Wang, Y.; Zhao, X.; Qi, X. Hormone therapy for ovarian cancer: Emphasis on mechanisms and applications (Review). Oncol. Rep. 2021, 46, 223. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, T.; Brown, W.D.; Sapi, E.; Aschkenazi, S.; Muñoz, A.; Mor, G. Absence of estrogen receptor-beta expression in metastatic ovarian cancer. Obstet. Gynecol. 2000, 96, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Cluzet, V.; Devillers, M.M.; Petit, F.; Pierre, A.; Giton, F.; Airaud, E.; L’Hôte, D.; Leary, A.; Genestie, C.; Treilleux, I.; et al. Estradiol promotes cell survival and induces Greb1 expression in granulosa cell tumors of the ovary through an ERα-dependent mechanism. J. Pathol. 2022, 256, 335–348. [Google Scholar] [CrossRef]
- Sieh, W.; Köbel, M.; Longacre, T.A.; Bowtell, D.D.; de Fazio, A.; Goodman, M.T.; Høgdall, E.; Deen, S.; Wentzensen, N.; Moysich, K.B.; et al. Hormone-receptor expression and ovarian cancer survival: An Ovarian Tumor Tissue Analysis consortium study. Lancet. Oncol. 2013, 14, 853–862. [Google Scholar] [CrossRef]
- Park, S.H.; Cheung, L.W.T.; Wong, A.S.T.; Leung, P.C.K. Estrogen regulates Snail and Slug in the down-regulation of E-cadherin and induces metastatic potential of ovarian cancer cells through estrogen receptor alpha. Mol. Endocrinol. 2008, 22, 2085–2098. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.K.L.; Leung, T.H.Y.; Chan, D.W.; Wei, N.; Lau, G.T.Y.; Liu, S.S.; Siu, M.K.Y.; Ngan, H.Y.S. Targeting estrogen receptor subtypes (ERα and ERβ) with selective ER modulators in ovarian cancer. J. Endocrinol. 2014, 221, 325–336. [Google Scholar] [CrossRef]
- Hoffmann, M.; Gogola, J.; Ptak, A. Apelin abrogates the stimulatory effects of 17β-estradiol and insulin-like growth factor-1 on proliferation of epithelial and granulosa ovarian cancer cell lines via crosstalk between APLNR and ERα/IGF1R. Mol. Biol. Rep. 2019, 46, 6325–6338. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Papaioannidou, P. Estrogen receptor beta and ovarian cancer: A key to pathogenesis and response to therapy. Arch. Gynecol. Obstet. 2016, 293, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Schüler-Toprak, S.; Weber, F.; Skrzypczak, M.; Ortmann, O.; Treeck, O. Estrogen receptor β is associated with expression of cancer associated genes and survival in ovarian cancer. BMC Cancer 2018, 18, 981. [Google Scholar] [CrossRef] [PubMed]
- Pinton, G.; Nilsson, S.; Moro, L. Targeting estrogen receptor beta (ERβ) for treatment of ovarian cancer: Importance of KDM6B and SIRT1 for ERβ expression and functionality. Oncogenesis 2018, 7, 15. [Google Scholar] [CrossRef]
- Liu, J.; Viswanadhapalli, S.; Garcia, L.; Zhou, M.; Nair, B.C.; Kost, E.; Rao Tekmal, R.; Li, R.; Rao, M.K.; Curiel, T.; et al. Therapeutic utility of natural estrogen receptor beta agonists on ovarian cancer. Oncotarget 2017, 8, 50002. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.K.L.; Siu, M.K.Y.; Jiang, Y.-X.; Wang, J.-J.; Leung, T.H.Y.; Ngan, H.Y.S. Estrogen receptor modulators genistein, daidzein and ERB-041 inhibit cell migration, invasion, proliferation and sphere formation via modulation of FAK and PI3K/AKT signaling in ovarian cancer. Cancer Cell Int. 2018, 18, 65. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Zaino, R.J.; Norris, H.J. Endometrial Carcinoma. In Blaustein’s Pathology of the Female Genital Tract; Springer: Berlin/Heidelberg, Germany, 1994; pp. 439–486. [Google Scholar] [CrossRef]
- Ellenson, L.H.; Ronnett, B.M.; Kurman, R.J. Precursor Lesions of Endometrial Carcinoma. In Blaustein’s Pathology of the Female Genital Tract; Springer: Berlin/Heidelberg, Germany, 2011; pp. 359–391. [Google Scholar] [CrossRef]
- Amant, F.; Mirza, M.R.; Koskas, M.; Creutzberg, C.L. Cancer of the corpus uteri. Int. J. Gynecol. Obstet. 2018, 143, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.J.; Thiel, K.W.; Leslie, K.K. Past, present, and future of hormonal therapy in recurrent endometrial cancer. Int. J. Women’s Health 2014, 6, 429–435. [Google Scholar] [CrossRef][Green Version]
- Jongen, V.; Briët, J.; de Jong, R.; ten Hoor, K.; Boezen, M.; van der Zee, A.; Nijman, H.; Hollema, H. Expression of estrogen receptor-alpha and -beta and progesterone receptor-A and -B in a large cohort of patients with endometrioid endometrial cancer. Gynecol. Oncol. 2009, 112, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.M.; Baker, W.H. Progestational Agents in the Treatment of Carcinoma of the Endometrium. N. Engl. J. Med. 2010, 264, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Hammes, S.R.; Levin, E.R. Impact of estrogens in males and androgens in females. J. Clin. Investig. 2019, 129, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Huggins, C.; Hodges, C.V. Studies on prostatic cancer. I. The effect of castration, of estrogen and of androgen injection on serum phosphatases in metastatic carcinoma of the prostate. CA Cancer J. Clin. 1972, 22, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Bonkhoff, H. Estrogen receptor signaling in prostate cancer: Implications for carcinogenesis and tumor progression. Prostate 2018, 78, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Bonkhoff, H.; Fixemer, T.; Hunsicker, I.; Remberger, K. Estrogen receptor expression in prostate cancer and premalignant prostatic lesions. Am. J. Pathol. 1999, 155, 641–647. [Google Scholar] [CrossRef]
- Qu, L.G.; Wardan, H.; Davis, I.D.; Pezaro, C.; Sluka, P. Effects of estrogen receptor signaling on prostate cancer carcinogenesis. Transl. Res. 2020, 222, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Fixemer, T.; Remberger, K.; Bonkhoff, H. Differential expression of the estrogen receptor beta (ERβ) in human prostate tissue, premalignant changes, and in primary, metastatic, and recurrent prostatic adenocarcinoma. Prostate 2003, 54, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Horvath, L.G.; Henshall, S.M.; Lee, C.S.; Head, D.R.; Quinn, D.I.; Makela, S.; Delprado, W.; Golovsky, D.; Brenner, P.C.; O’Neill, G.; et al. Frequent loss of estrogen receptor-β expression in prostate cancer. Cancer Res. 2001, 61, 5331–5335. [Google Scholar] [PubMed]
- Mishra, S.; Tai, Q.; Gu, X.; Schmitz, J.; Poullard, A.; Fajardo, R.J.; Mahalingam, D.; Chen, X.; Zhu, X.; Sun, L.Z. Estrogen and estrogen receptor alpha promotes malignancy and osteoblastic tumorigenesis in prostate cancer. Oncotarget 2015, 6, 44388–44402. [Google Scholar] [CrossRef]
- Bardin, A.; Boulle, N.; Lazennec, G.; Vignon, F.; Pujol, P. Loss of ERbeta expression as a common step in estrogen-dependent tumor progression. Endocr. Relat. Cancer 2004, 11, 537–551. [Google Scholar] [CrossRef]
- McPherson, S.J.; Hussain, S.; Balanathan, P.; Hedwards, S.L.; Niranjan, B.; Grant, M.; Chandrasiri, U.P.; Toivanen, R.; Wang, Y.; Taylor, R.A.; et al. Estrogen receptor-beta activated apoptosis in benign hyperplasia and cancer of the prostate is androgen independent and TNFalpha mediated. Proc. Natl. Acad. Sci. USA 2010, 107, 3123–3128. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Goodin, S.; Doyle-Lindrud, S.; Silberberg, J.; Kane, M.; Metzger, D.; Eddy, S.; Shih, W.; DiPaola, R.S. Transdermal estradiol in castrate and chemotherapy resistant prostate cancer. Med. Sci. Monit. 2012, 18, CR260–CR264. [Google Scholar] [CrossRef][Green Version]
- Beer, T.M.; Ryan, C.; Alumkal, J.; Ryan, C.W.; Sun, J.; Eilers, K.M. A phase II study of paclitaxel poliglumex in combination with transdermal estradiol for the treatment of metastatic castration-resistant prostate cancer after docetaxel chemotherapy. Anticancer Drugs 2010, 21, 433–438. [Google Scholar] [CrossRef]
- Fujimura, T.; Takahashi, S.; Kume, H.; Urano, T.; Takayama, K.; Yamada, Y.; Suzuki, M.; Fukuhara, H.; Nakagawa, T.; Inoue, S.; et al. Toremifene, a selective estrogen receptor modulator, significantly improved biochemical recurrence in bone metastatic prostate cancer: A randomized controlled phase II a trial. BMC Cancer 2015, 15, 836. [Google Scholar] [CrossRef]
- Pritchard, T.; Rosengren, R.J.; Greish, K.; Taurin, S. Raloxifene nanomicelles reduce the growth of castrate-resistant prostate cancer. J. Drug Target. 2016, 24, 441–449. [Google Scholar] [CrossRef]
- Thelen, P.; Wuttke, W.; Seidlová-Wuttke, D. Phytoestrogens selective for the estrogen receptor beta exert anti-androgenic effects in castration resistant prostate cancer. J. Steroid Biochem. Mol. Biol. 2014, 139, 290–293. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, K.; Chen, L.; Yin, B.; Song, Y. Is phytoestrogen intake associated with decreased risk of prostate cancer? A systematic review of epidemiological studies based on 17,546 cases. Andrology 2016, 4, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Montoya, M.C.; Krysan, D.J. Repurposing Estrogen Receptor Antagonists for the Treatment of Infectious Disease. MBio 2018, 9, e02272-18. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.A.; Barnes, A.B.; Wiehle, R.D.; Fontenot, G.K.; Hoenen, T.; White, J.M. Clomiphene and Its Isomers Block Ebola Virus Particle Entry and Infection with Similar Potency: Potential Therapeutic Implications. Viruses 2016, 8, 206. [Google Scholar] [CrossRef] [PubMed]
- Ghasemnejad-Berenji, M.; Pashapour, S.; Ghasemnejad-Berenji, H. Therapeutic potential for clomiphene, a selective estrogen receptor modulator, in the treatment of COVID-19. Med. Hypotheses 2020, 145, 110354. [Google Scholar] [CrossRef] [PubMed]












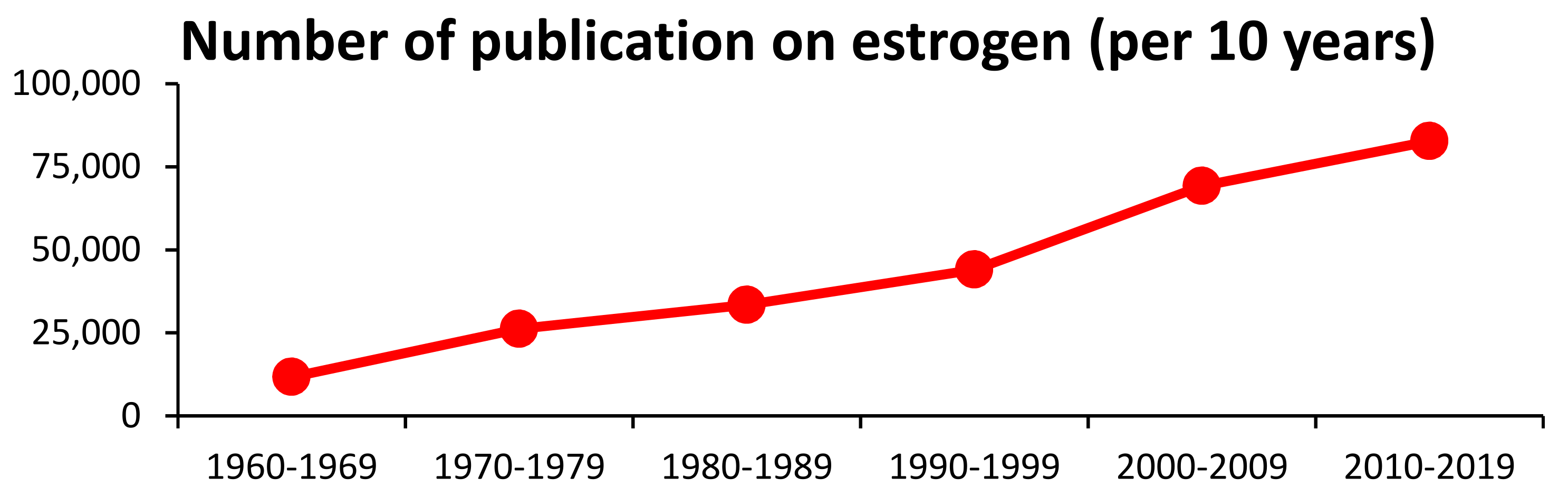{kind=link}
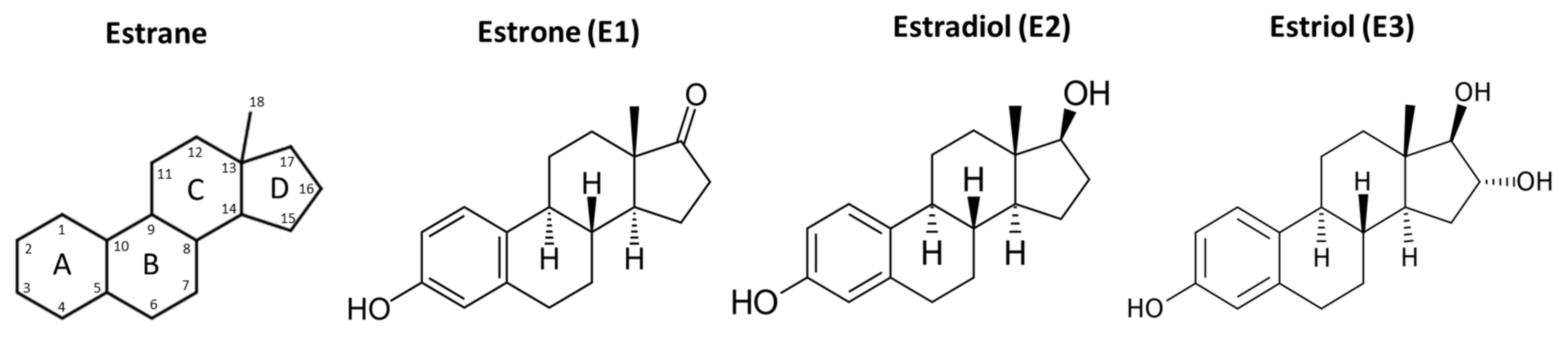{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Brain (VMS) | Cardiovascular System | Bone | Breast | Endometrium | Therapy | References |
|---|---|---|---|---|---|---|---|
| Estrogen | + | + | + | + | + | Hormonal contraception, hormone replacement therapy (HRT), feminizing hormone therapy in transgender woman. | refer Section 6 |
| Clomiphene citrate | −/+ (dose-dependent) | + | + | − | − | Treatment of female infertility, and male hypogonadism. | [111,112,113,114] |
| Tamoxifen | − | + | + | − | + | Adjuvant treatment of ER+ breast cancer, prevention in high-risked women. | [115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130] |
| Toremifene | − | + | + | − | + (weak) | Adjuvant treatment of ER+ breast cancer. | [131,132,133] |
| Raloxifene | − | + | + | − | neutral | Treatment of osteoporosis in postmenopause, or in women with high risk of ER+ breast cancer. | [134,135,136] |
| Bazedoxifene | − | + | + | − | − | Treatment of osteoporosis in postmenopausal women. | [137,138,139,140] |
| Ospemifene | − | + | + | − | −/+ | Treatment of genitourinary syndrome of menopause. | [141,142,143] |
| Lasofoxifene | − (weak) | + | + | − | −/+ | Treatment of osteoporosis and vaginal atrophy. | [144,145] |
| Phyto-estrogens | + | + | + | − | − | Treatment of menopausal symptoms. | [146,147,148,149,150,151,152,153,154,155,156,157] |
| Fulvestrant | − | − | − | − | − | Treatment of advanced ER+ breast cancer. | [158,159,160,161,162,163,164] |
| ANGELS | + | + | + | ? | − | Not in therapy. | [166,167,168,169,170] |
| G-1 | + | + | + | + | + | Not in therapy. | [171,172,173,174,175,176] |
| G-15 and G-36 | − | − | ? | − | − | Not in therapy. | [176,177,178] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farkas, S.; Szabó, A.; Hegyi, A.E.; Török, B.; Fazekas, C.L.; Ernszt, D.; Kovács, T.; Zelena, D. Estradiol and Estrogen-like Alternative Therapies in Use: The Importance of the Selective and Non-Classical Actions. Biomedicines 2022, 10, 861. https://doi.org/10.3390/biomedicines10040861
Farkas S, Szabó A, Hegyi AE, Török B, Fazekas CL, Ernszt D, Kovács T, Zelena D. Estradiol and Estrogen-like Alternative Therapies in Use: The Importance of the Selective and Non-Classical Actions. Biomedicines. 2022; 10(4):861. https://doi.org/10.3390/biomedicines10040861
Chicago/Turabian StyleFarkas, Szidónia, Adrienn Szabó, Anita Emőke Hegyi, Bibiána Török, Csilla Lea Fazekas, Dávid Ernszt, Tamás Kovács, and Dóra Zelena. 2022. "Estradiol and Estrogen-like Alternative Therapies in Use: The Importance of the Selective and Non-Classical Actions" Biomedicines 10, no. 4: 861. https://doi.org/10.3390/biomedicines10040861
APA StyleFarkas, S., Szabó, A., Hegyi, A. E., Török, B., Fazekas, C. L., Ernszt, D., Kovács, T., & Zelena, D. (2022). Estradiol and Estrogen-like Alternative Therapies in Use: The Importance of the Selective and Non-Classical Actions. Biomedicines, 10(4), 861. https://doi.org/10.3390/biomedicines10040861