

Epithelial Cell Adhesion Molecule (EpCAM) Expression Can Be Modulated via NFκB

 , , , ,
, , , ,  , , , , , , and
, , , , , , and
Abstract
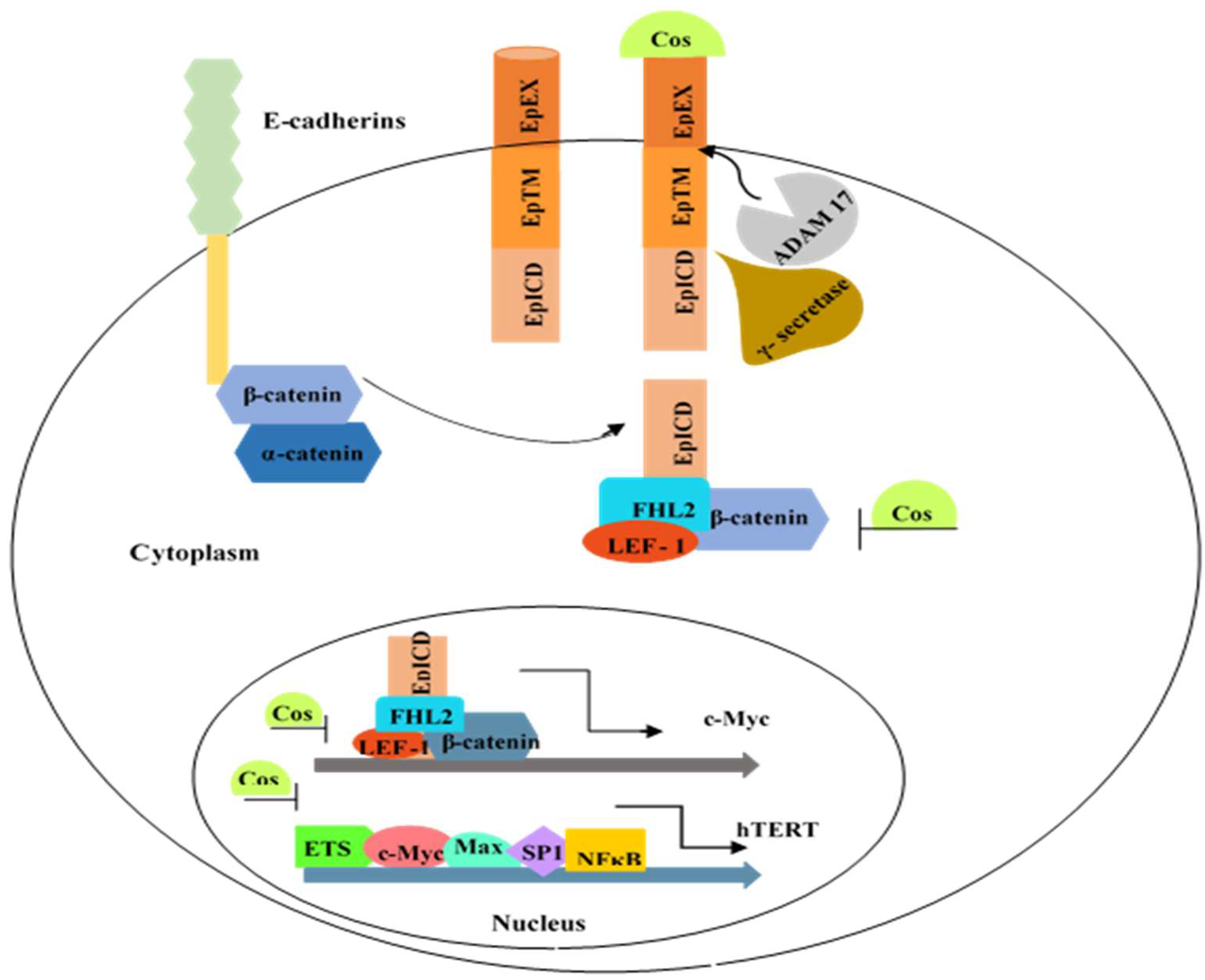
1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. RNA Isolation
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. In Vitro Treatment of Jurkat Cells with Costunolide
2.6. Quantitative Real Time PCR
2.7. Measurement of Telomere Length
2.8. Docking of Costunolide with EpCAM and Myc Proteins
2.9. Computational Analysis
2.10. Statistical Analysis
3. Results
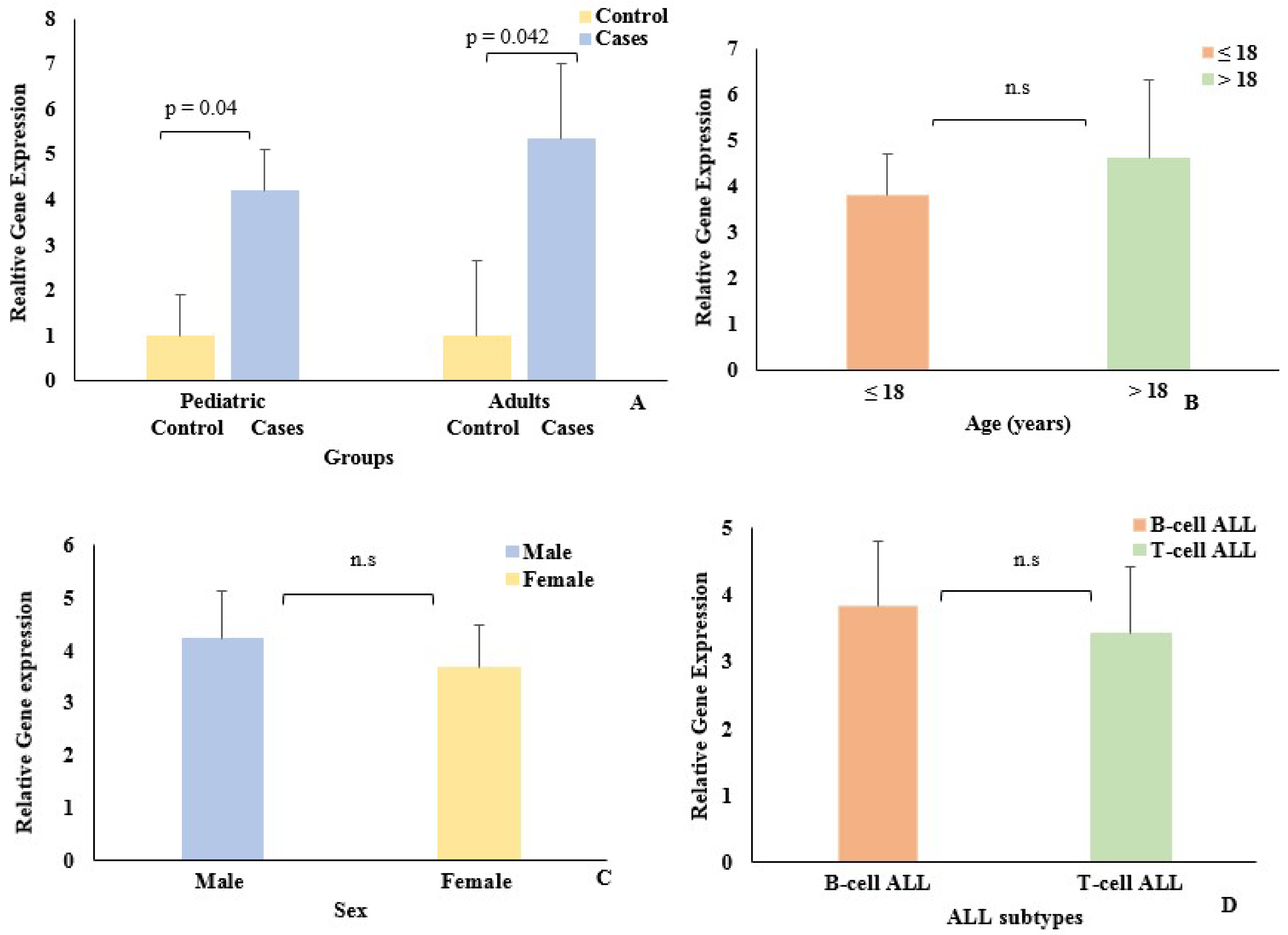
3.1. Association of EpCAM Gene Expression with Demographic Data
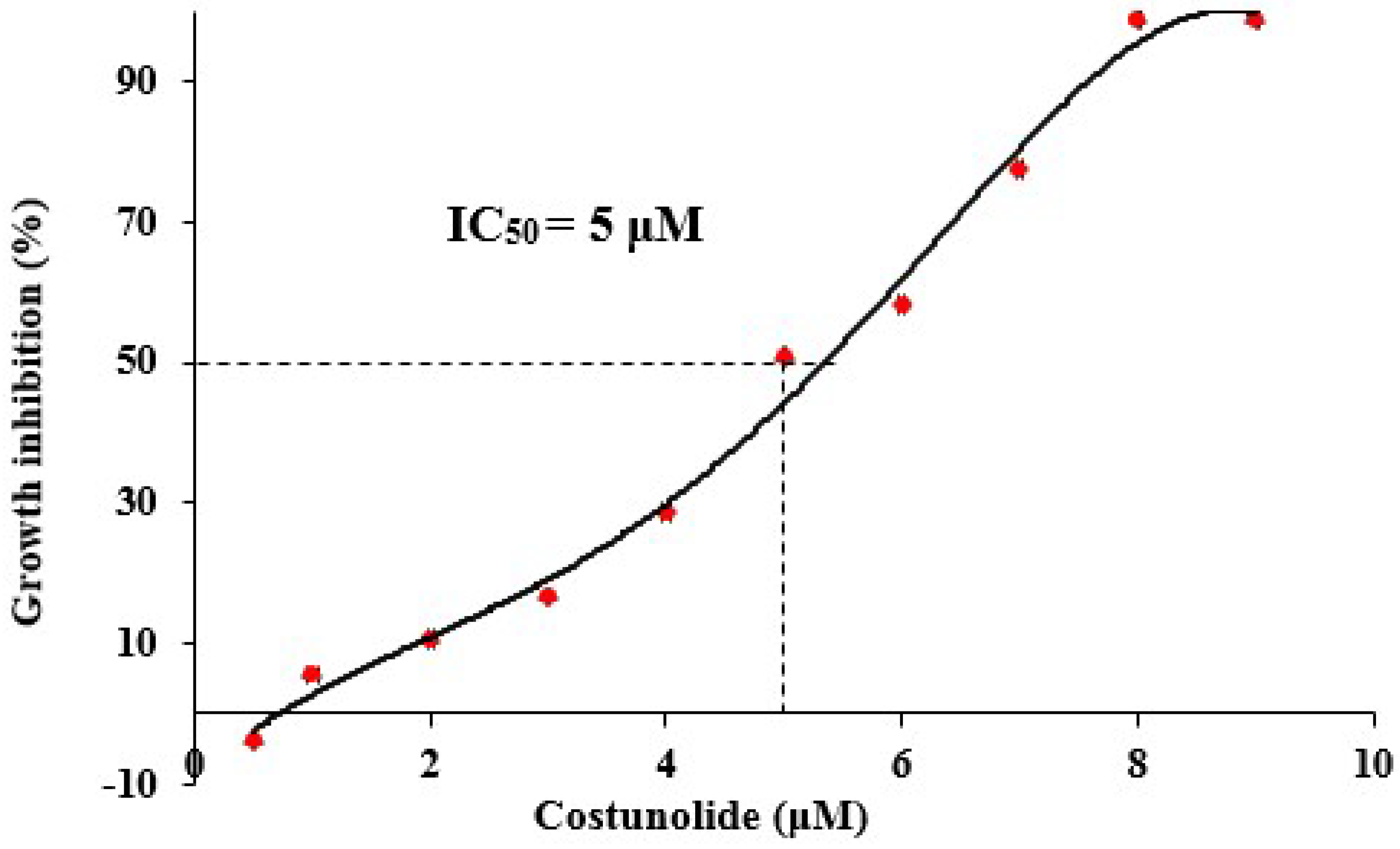
3.2. Effect of Costunolide on Jurkat Cell Viability
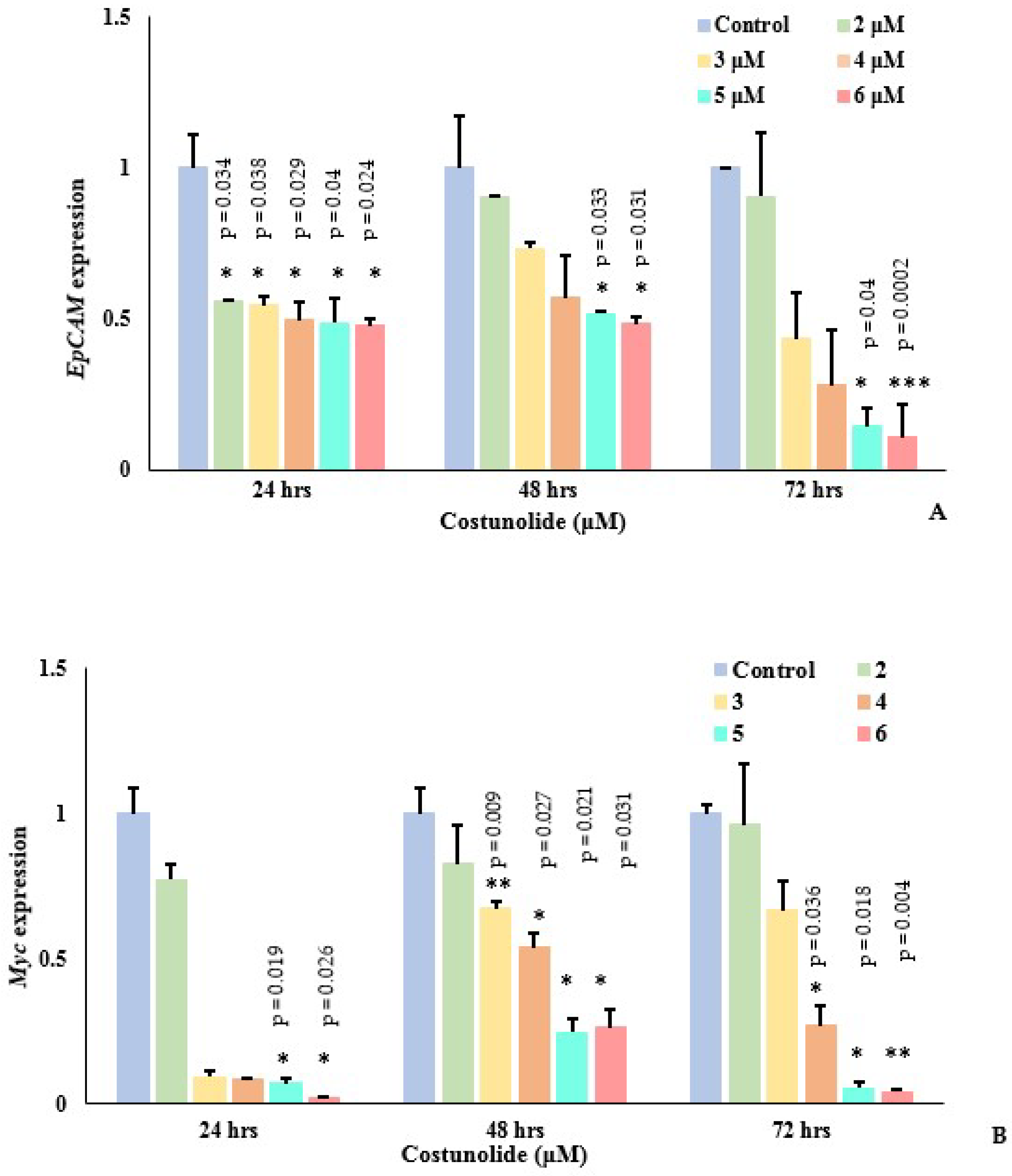
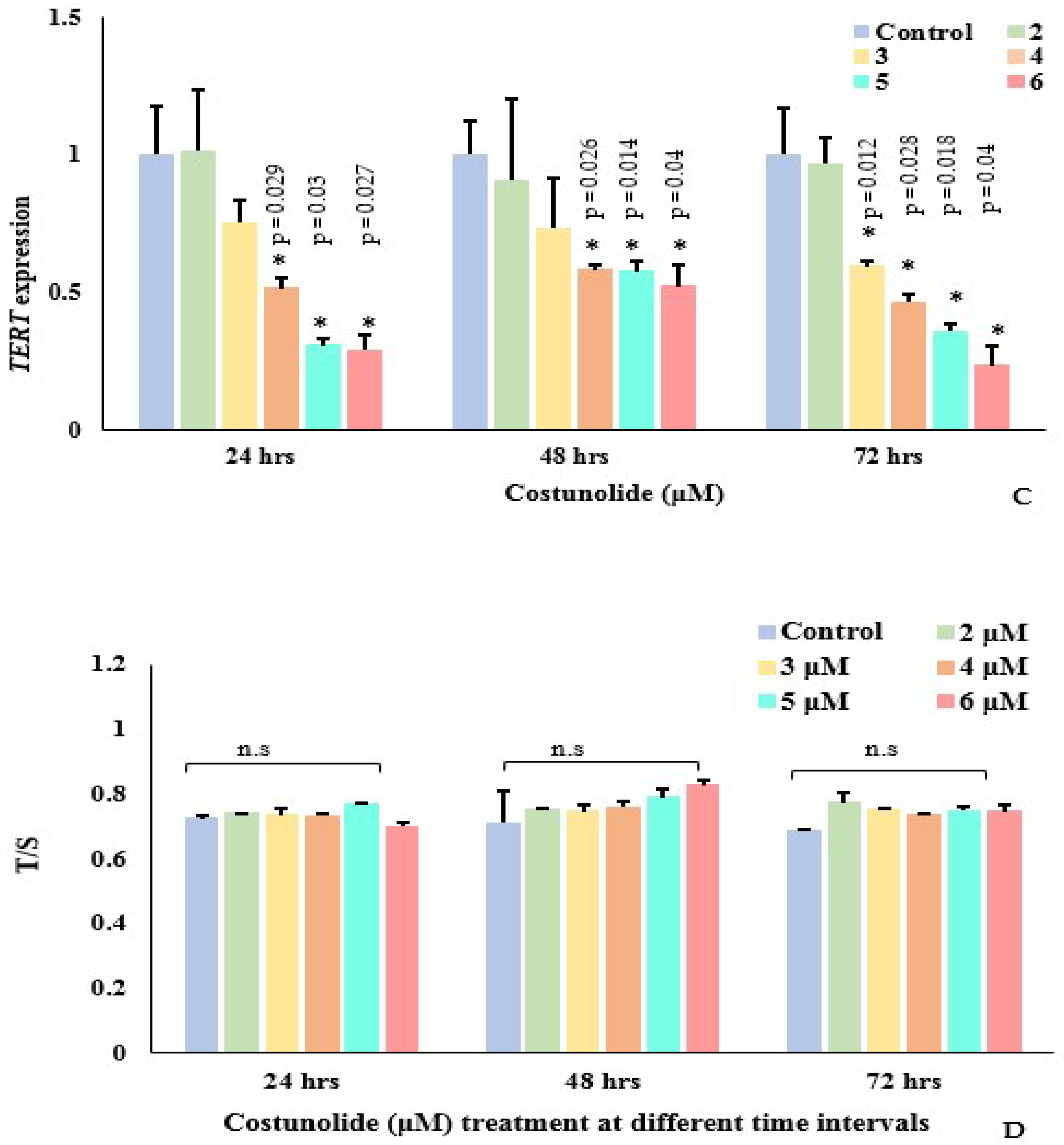
3.3. Effect of Costunolide on Genes (EpCAM, Myc and TERT) Expression and Telomere Length
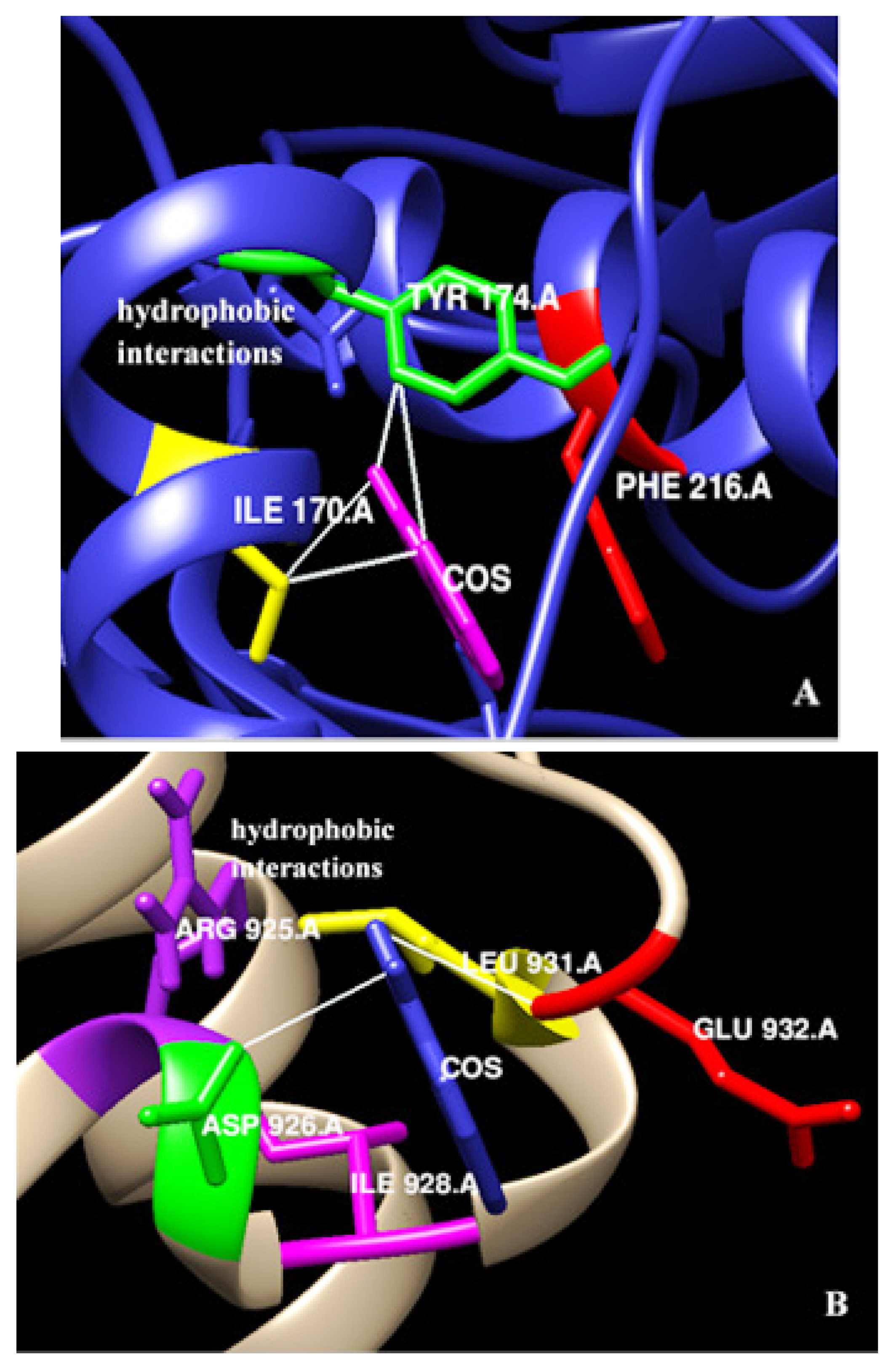
3.4. In-Silico Interaction of Costunolide with EpCAM and Myc
3.5. Identification of Transcription Factor Binding Sites within EpCAM, Myc, and TERT Promoter Region
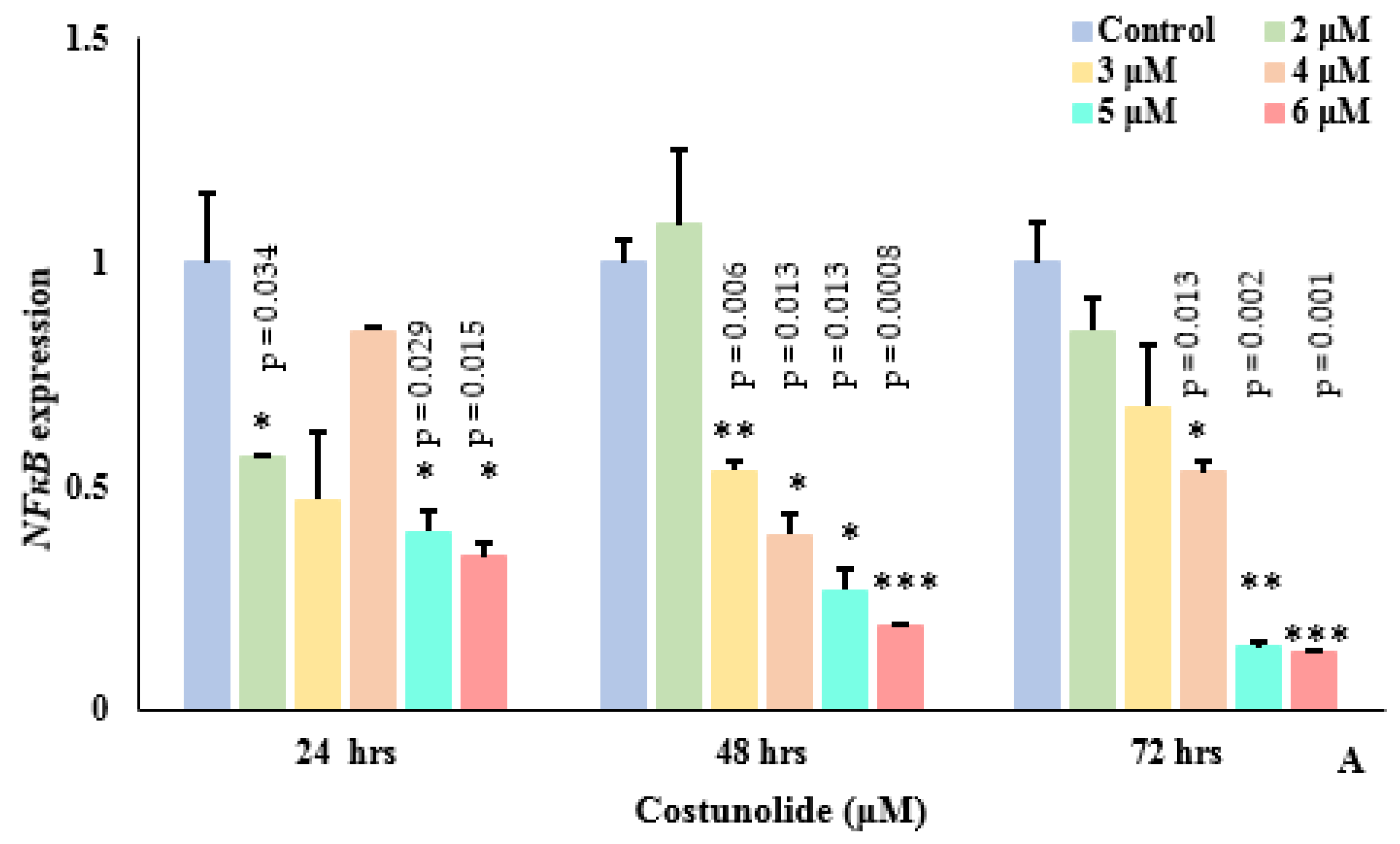
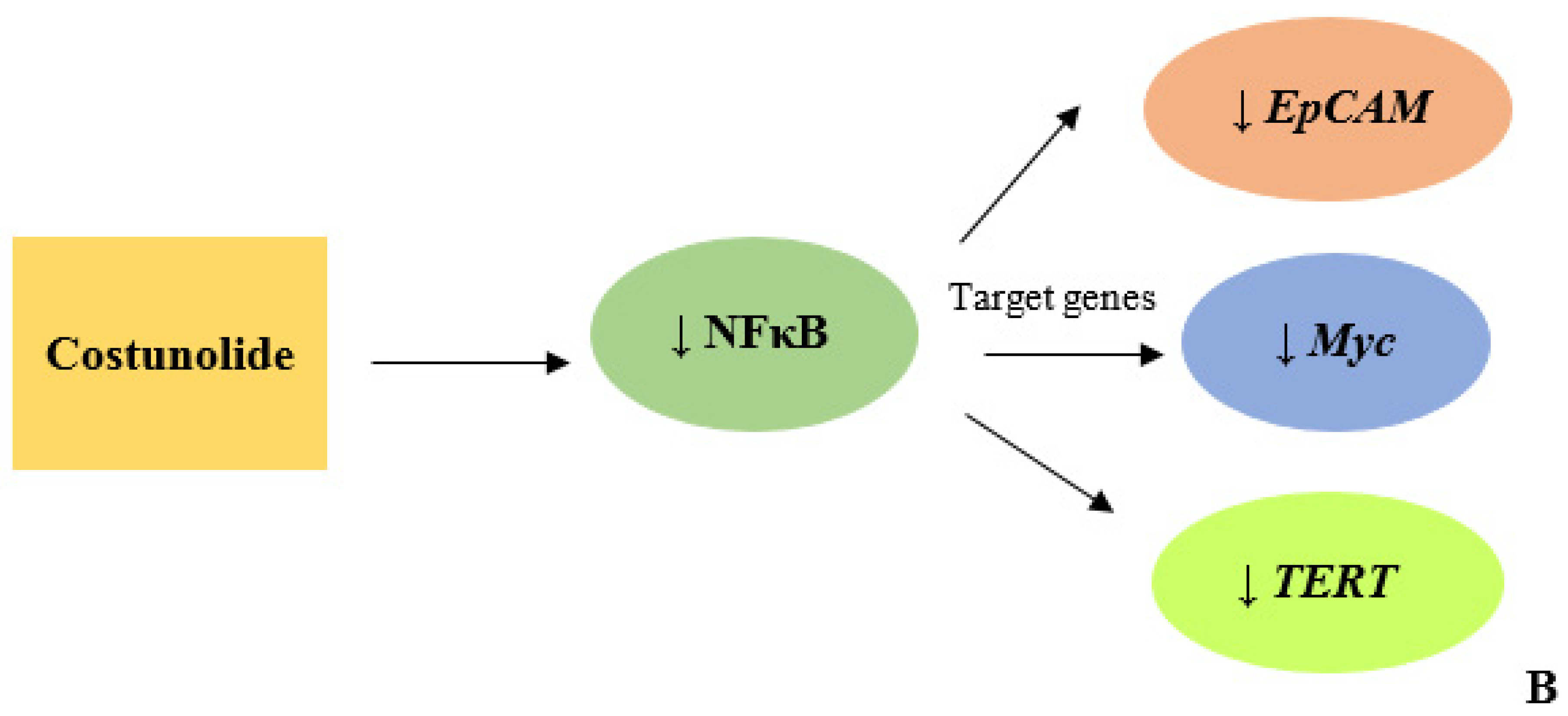
3.6. Costunolide Reduced NFκB Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, D.R.; Weber, J.K.; Yin, W.; Huynh, T.; Duan, W.; Zhou, R. In silico design and validation of high-affinity RNA aptamers targeting epithelial cellular adhesion molecule dimers. Proc. Natl. Acad. Sci. USA 2020, 117, 8486–8493. [Google Scholar] [CrossRef] [PubMed]
- Massoner, P.; Thomm, T.; Mack, B.; Untergasser, G.; Martowicz, A.; Bobowski, K.; Klocker, H.; Gires, O.; Puhr, M. EpCAM is overexpressed in local and metastatic prostate cancer, suppressed by chemotherapy and modulated by MET-associated miRNA-200c/205. Br. J. Cancer 2014, 111, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Fagotto, F.; Aslemarz, A. EpCAM cellular functions in adhesion and migration, and potential impact on invasion: A critical review. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2020, 1874, 188436. [Google Scholar] [CrossRef] [PubMed]
- Martowicz, A.; Rainer, J.; Lelong, J.; Spizzo, G.; Gastl, G.; Untergasser, G. EpCAM overexpression prolongs proliferative capacity of primary human breast epithelial cells and supports hyperplastic growth. Mol. Cancer 2013, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Schnell, U.; Cirulli, V.; Giepmans, B.N. EpCAM: Structure and function in health and disease. Biochim. Biophys. Acta 2013, 1828, 1989–2001. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, X.; Gao, J.; Sun, Y.; Liu, T.; Yan, Q.; Yang, X. The role of epithelial cell adhesion molecule N-glycosylation on apoptosis in breast cancer cells. Tumor Biol. 2017, 39, 1–8. [Google Scholar] [CrossRef]
- Zheng, X.; Fan, X.; Fu, B.; Zheng, M.; Zhang, A.; Zhong, K.; Yan, J.; Sun, R.; Tian, Z.; Wei, H. EpCAM inhibition sensitizes chemoresistant leukemia to immune surveillance. Cancer Res. 2017, 77, 482–493. [Google Scholar] [CrossRef]
- Gaber, A.; Lenarčič, B.; Pavšič, M. Current view on EpCAM structural biology. Cells 2020, 9, 1361. [Google Scholar] [CrossRef]
- Mal, A.; Dey, P.; Hayes, R.M.; McCarthy, J.V.; Ray, A.; De, A. In Silico Identification of Potential Phosphorylation in the Cytoplasmic Domain of Epithelial Cell Adhesion Molecule. ACS Omega 2020, 5, 30808–30816. [Google Scholar] [CrossRef]
- Dratwa, M.; Wysoczańska, B.; Łacina, P.; Kubik, T.; Bogunia-Kubik, K. TERT—Regulation and Roles in Cancer Formation. Front. Immunol. 2020, 11, 589929. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Z.; Xie, Y.; Hu, H. Antitumor activity and mechanism of costunolide and dehydrocostus lactone: Two natural sesquiterpene lactones from the Asteraceae family. Biomed. Pharm. 2020, 125, 109955. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Choi, B.Y. Costunolide—A bioactive sesquiterpene lactone with diverse therapeutic potential. Int. J. Mol. Sci. 2019, 20, 2926. [Google Scholar] [CrossRef] [PubMed]
- Tabata, K.; Nishimura, Y.; Takeda, T.; Kurita, M.; Uchiyama, T.; Suzuki, T. Sesquiterpene lactones derived from Saussurealappa induce apoptosis and inhibit invasion and migration in neuroblastoma cells. J. Pharmacol. Sci. 2015, 127, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, Y.; Fan, J.; Lin, X.; Liu, C.; Xu, Y.; Su, C. Costunolide and dehydrocostus lactone combination treatment inhibit breast cancer by inducing cell cycle arrest and apoptosis through c-Myc/p53 and AKT/14-3-3 pathway. Sci. Rep. 2017, 7, 1–16. [Google Scholar]
- Ramesh, A.; Ghosal, S.; Muralidharan, D.; Parvathi, V.D.; Dharmarajan, A. Reviewing Leads That Promote Apoptosis-Cancer Treatment Strategies. Biomed. J. Sci. Tech. Res. 2021, 36, 28283–28290. [Google Scholar]
- Lee, S.H.; Cho, Y.C.; Lim, J.S. Costunolide, a sesquiterpene lactone, suppresses skin cancer via induction of apoptosis and blockage of cell proliferation. Int. J. Mol. Sci. 2021, 2, 2075. [Google Scholar] [CrossRef]
- Choi, Y.K.; Cho, S.G.; Woo, S.M.; Yun, Y.J.; Jo, J.; Kim, W.; Shin, Y.C.; Ko, S.G. Saussurealappa Clarke-derived costunolide prevents TNFα-induced breast cancer cell migration and invasion by inhibiting NF-κB activity. Evid.-Based Complement. Altern. Med. 2013, 2013, 936257. [Google Scholar] [CrossRef]
- Wang, J.; Yu, Z.; Wang, C.; Tian, X.; Huo, X.; Wang, Y.; Sun, C.; Feng, L.; Ma, J.; Zhang, B.; et al. Dehydrocostus lactone, a natural sesquiterpene lactone, suppresses the biological characteristics of glioma, through inhibition of the NF-κB/COX-2 signaling pathway by targeting IKKβ. Am. J. Cancer Res. 2017, 7, 1270. [Google Scholar]
- Jakobs, A.; Steinmann, S.; Henrich, S.M.; Schmidt, T.J.; Klempnauer, K.H. Helenalin acetate, a natural sesquiterpene lactone with anti-inflammatory and anti-cancer activity, disrupts the cooperation of CCAAT box/enhancer-binding protein β (C/EBPβ) and co-activator p300. J. Biol. Chem. 2016, 291, 26098–26108. [Google Scholar] [CrossRef]
- Siedle, B.; García-Piñeres, A.J.; Murillo, R.; Schulte-Mönting, J.; Castro, V.; Rüngeler, P.; Klaas, C.A.; Da Costa, F.B.; Kisiel, W.; Merfort, I. Quantitative structure-activity relationship of sesquiterpene lactones as inhibitors of the transcription Factor NF-κB. J. Med. Chem. 2004, 47, 6042–6054. [Google Scholar] [CrossRef]
- Hu, M.; Liu, L.; Yao, W. Activation of p53 by costunolide blocks glutaminolysis and inhibits proliferation in human colorectal cancer cells. Gene 2018, 678, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yi, J.; Park, S.; Zhang, H.; Kim, E.; Park, S.; Kwon, W.; Jang, S.; Zhang, X.; Chen, H.; et al. Costunolide suppresses melanoma growth via the AKT/mTOR pathway in vitro and in vivo. Am. J. Cancer Res. 2021, 11, 1410. [Google Scholar] [PubMed]
- Jin, X.; Wang, C. Costunolide inhibits osteosarcoma growth and metastasis via suppressing STAT3 signal pathway. Biomed. Pharmacother. 2020, 121, 109659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lu, T.; Wang, G.D.; Ma, C.; Zhou, Y.F. Costunolide, an active sesquiterpene lactone, induced apoptosis via ROS-mediated ER stress and JNK pathway in human U2OS cells. Biomed. Pharmacother. 2016, 80, 253–259. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef]
- Nettle, D.; Monaghan, P.; Gillespie, R.; Brilot, B.; Bedford, T.; Bateson, M. An experimental demonstration that early-life competitive disadvantage accelerates telomere loss. Proc. R. Soc. B Biol. Sci. 2015, 282, 20141610. [Google Scholar] [CrossRef]
- Pavšič, M.; Gunčar, G.; Djinović-Carugo, K.; Lenarčič, B. Crystal structure and its bearing towards an understanding of key biological functions of EpCAM. Nat. Commun. 2014, 5, 4764. [Google Scholar] [CrossRef]
- Zhao, Q.; Assimopoulou, A.N.; Klauck, S.M.; Damianakos, H.; Chinou, I.; Kretschmer, N.; Rios, J.-L.; Papageorgiou, V.P.; Bauer, R.; Efferth, T. Inhibition of MYC with involvement of ERK/JNK/MAPK and AKT pathways as a novel mechanism for shikonin and its derivatives in killing leukemia cells. Oncotarget 2015, 6, 38934. [Google Scholar] [CrossRef]
- Dreos, R.; Ambrosini, G.; Périer, R.C.; Bucher, P. The Eukaryotic Promoter Database: Expansion of EPDnew and new promoter analysis tools. Nucleic Acids Res. 2015, 43, D92–D96. [Google Scholar] [CrossRef]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Nowsheen, S. Sustained proliferation in cancer: Mechanisms and novel therapeutic targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef]
- Warneke, V.S.; Behrens, H.M.; Haag, J.; Krüger, S.; Simon, E.; Mathiak, M.; Ebert, M.P.A.; Röcken, C. Members of the EpCAMsignalling pathway are expressed in gastric cancer tissue and are correlated with patient prognosis. Br. J. Cancer 2013, 109, 2217. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vallera, D.A.; Zhang, B.; Gleason, M.K.; Oh, S.; Weiner, L.M.; Kaufman, D.; McCullar, V.; Miller, J.S.; Verneris, M.R. Heterodimeric bispecific single-chain variable-fragment antibodies against EpCAM and CD16 induce effective antibody-dependent cellular cytotoxicity against human carcinoma cells. Cancer Biother. Radiopharm. 2013, 28, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.; Werner, S.; Pantel, K. Biology and clinical relevance of EpCAM. Cell Stress. 2019, 3, 165. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Sa, N.; Yu, L.; Lu, S.; Wang, H.; Xu, W. Epithelial cell adhesion molecule is overexpressed in hypopharyngeal carcinoma and suppresses the metastasis and proliferation of the disease when downregulated. Oncol. Lett. 2014, 8, 175–182. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sen, S.; Carnelio, S. Expression of epithelial cell adhesion molecule (Ep CAM) in oral squamous cell carcinoma. Histopathology 2016, 68, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Hekimian, K.; Meisezahl, S.; Trompelt, K.; Rabenstein, C.; Pachmann, K. Epithelial cell dissemination and readhesion: Analysis of factors contributing to metastasis formation in breast cancer. Int. Sch. Res. Not. 2012, 2012, 601810. [Google Scholar] [CrossRef]
- Choi, J.-H.; Seo, B.-R.; Seo, S.-H.; Lee, K.-T.; Park, J.-H.; Park, H.-J.; Choi, J.-W.; Itoh, Y.; Miyamoto, K.-I. Costunolide induces differentiation of human leukemia HL-60 cells. Arch. Pharmacal Res. 2002, 25, 480–484. [Google Scholar] [CrossRef]
- Kanno, S.I.; Kitajima, Y.; Kakuta, M.; Osanai, Y.; Kurauchi, K.; Ujibe, M.; Ishikawa, M. Costunolide-induced apoptosis is caused by receptor-mediated pathway and inhibition of telomerase activity in NALM-6 cells. Biol. Pharm. Bull. 2008, 31, 1024–1028. [Google Scholar] [CrossRef]
- Zach, L.O.; Beery, E.; Lahav, M.; Uziel, O. The effects of Rapamycin on Telomerase Activity and Regulation in Cancer Cells. Br. J. Cancer Res. 2019, 2, 334–340. [Google Scholar]
- Huang, E.; Tedone, E.; O’Hara, R.; Cornelius, C.; Lai, T.-P.; Ludlow, A.; Wright, W.; Shay, J.W. The maintenance of telomere length in CD28+ T cells during T lymphocyte stimulation. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Zhdanov, D.D.; Pokrovsky, V.S.; Pokrovskaya, M.V.; Alexandrova, S.S.; Eldarov, M.A.; Grishin, D.V.; Basharov, M.M.; Gladilina, Y.A.; Podobed, O.V.; Sokolov, N.N. Inhibition of telomerase activity and induction of apoptosis by Rhodospirillum rubrum l-asparaginase in cancer Jurkat cell line and normal human CD4+ T lymphocytes. Cancer Med. 2017, 6, 2697–2712. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Kang, X.; He, L.; Zhou, J.; Zhou, J.; Sturm, M.B.; Beer, D.G.; Kuick, R.; Nancarrow, D.J.; Appelman, H.D.; et al. Identification of tumor specific peptide as EpCAM ligand and its potential diagnostic and therapeutic clinical application. Mol. Pharm. 2019, 16, 2199–2213. [Google Scholar] [CrossRef] [PubMed]
- Alshaer, W.; Ababneh, N.; Hatmal, M.; Izmirli, H.; Choukeife, M.; Shraim, A.; Sharar, N.; Abu-Shiekah, A.; Odeh, F.; Al Bawab, A.; et al. Selection and targeting of EpCAM protein by ssDNA aptamer. PLoS ONE 2017, 12, e0189558. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Huangfu, Y.; Ge, J.; Wu, B.; Li, W.; Wang, X.; Zhao, L. Computational study on natural compounds inhibitor of c-Myc. Medicine 2020, 99, e23342. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Xie, Y.; Sun, X.; Zhang, M.; Zhou, J.; Liu, L.; Gao, J.; Xu, K. Identification of a novel c-Myc inhibitor 7594-0037 by structure-based virtual screening and investigation of its anti-cancer effect on multiple myeloma. Drug Des. Dev. Ther. 2020, 14, 3983. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Hubbard, A.K.; Giardina, C. NF-κB regulates transcription of the mouse telomerase catalytic subunit. J. Biol. Chem. 2000, 275, 36671–36675. [Google Scholar] [CrossRef]
- La Rosa, F.A.; Pierce, J.W.; Sonenshein, G.E. Differential regulation of the c-myc oncogene promoter by the NF-kappa B rel family of transcription factors. Mol. Cell. Biol. 1994, 14, 1039–1044. [Google Scholar]
- Huang, H.; Ma, L.; Li, J.; Yu, Y.; Zhang, D.; Wei, J.; Jin, H.; Xu, D.; Gao, J.; Huang, C. NF-κB1 inhibits c-Myc protein degradation through suppression of FBW7 expression. Oncotarget 2014, 5, 493. [Google Scholar] [CrossRef]
- Herreros-Pomares, A.; Aguilar-Gallardo, C.; Calabuig-Fariñas, S.; Sirera, R.; Jantus-Lewintre, E.; Camps, C. EpCAM duality becomes this molecule in a new Dr. Jekyll and Mr. Hyde tale. Crit. Rev. Oncol. Hematol. 2018, 126, 52–63. [Google Scholar] [CrossRef]
- Van Der Gun, B.T.F.; De Groote, M.L.; Kazemier, H.G.; Arendzen, A.J.; Terpstra, P.; Ruiters, M.H.J.; McLaughlin, P.M.J.; Rots, M.G. Transcription factors and molecular epigenetic marks underlying EpCAM overexpression in ovarian cancer. Br. J. Cancer 2011, 105, 312–319. [Google Scholar] [CrossRef]
- NF-kB Target Genes. Available online: https://www.bu.edu/nf-kb/gene-resources/target-genes/ (accessed on 10 November 2022).
- Nan, L.; Nam, H.H.; Choo, B.K. Costunolide inhibits inflammation in LPS-induced RAW264. 7 cells and ameliorates gastric acid reflux-induced esophageal injury in rat model. Appl. Biol. Chem. 2020, 63, 1–9. [Google Scholar] [CrossRef]
- Mao, J. Protective effects of costunolide against D-galactosamine and lipopolysaccharide-induced acute liver injury in mice. Front. Pharmacol. 2018, 9, 1469. [Google Scholar] [CrossRef] [PubMed]
- Pitchai, D.; Roy, A.; Banu, S. In Vitro and In Silico Evaluation of NF-κB Targeted Costunolide Action on Estrogen Receptor-Negative Breast Cancer Cells—A Comparison with Normal Breast Cells. Phytother. Res. 2014, 28, 1499–1505. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cases vs. Control | ||||||
|---|---|---|---|---|---|---|
| Pediatric | Adults | |||||
| Characteristics | Controls | Cases | p Value | Controls | Cases | p Value |
| Subjects (N) | 50 | 153 | - | 39 | 62 | - |
| Age (years) | 2.77 (3.25) | 8.58 (4.92) | 0.0026 | 30.0 (9.04) | 31.3 (9.12) | 0.5983 |
| Sex (Male) (%) | 31 (62.0%) | 100 (65.4%) | 0.830 | 31 (79.5%) | 49 (79.0%) | 0.797 |
| ALL immunophenotypes | ||||||
| B-cell ALL117 (76.5%)45 (72.5%)0.3413 | ||||||
| T-cell ALL36 (23.5%)17 (27.7%)0.9218 | ||||||
| NFκB Subunits | EpCAM | Myc | TERT |
|---|---|---|---|
| NFκB1/p50 | −916, −691, −134 bps | −161, −7 bps | −742, −727, −694, −692 bps |
| p65/RelA | −393 bps | −259 bps | −599, −598 bps |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zia, S.; Tehreem, K.; Batool, S.; Ishfaq, M.; Mirza, S.B.; Khan, S.; Almashjary, M.N.; Hazzazi, M.S.; Qanash, H.; Shaikh, A.; et al. Epithelial Cell Adhesion Molecule (EpCAM) Expression Can Be Modulated via NFκB. Biomedicines 2022, 10, 2985. https://doi.org/10.3390/biomedicines10112985
Zia S, Tehreem K, Batool S, Ishfaq M, Mirza SB, Khan S, Almashjary MN, Hazzazi MS, Qanash H, Shaikh A, et al. Epithelial Cell Adhesion Molecule (EpCAM) Expression Can Be Modulated via NFκB. Biomedicines. 2022; 10(11):2985. https://doi.org/10.3390/biomedicines10112985
Chicago/Turabian StyleZia, Saadiya, Komal Tehreem, Sidra Batool, Mehreen Ishfaq, Shaher Bano Mirza, Shahrukh Khan, Majed N. Almashjary, Mohannad S. Hazzazi, Husam Qanash, Ahmad Shaikh, and et al. 2022. "Epithelial Cell Adhesion Molecule (EpCAM) Expression Can Be Modulated via NFκB" Biomedicines 10, no. 11: 2985. https://doi.org/10.3390/biomedicines10112985
APA StyleZia, S., Tehreem, K., Batool, S., Ishfaq, M., Mirza, S. B., Khan, S., Almashjary, M. N., Hazzazi, M. S., Qanash, H., Shaikh, A., Baty, R. S., Jafri, I., Alsubhi, N. H., Alrefaei, G. I., Sami, R., & Shahid, R. (2022). Epithelial Cell Adhesion Molecule (EpCAM) Expression Can Be Modulated via NFκB. Biomedicines, 10(11), 2985. https://doi.org/10.3390/biomedicines10112985